La estación de cosecha afecta el contenido de semillas y anatomía del fruto de berenjena violeta
THE HARVEST SEASON AFFECTS THE SEED CONTENT AND ANATOMY OF VIOLET EGGPLANT FRUIT
La estación de cosecha afecta el contenido de semillas y anatomía del fruto de berenjena violeta
Revista Iberoamericana de Tecnología Postcosecha, vol. 18, núm. 2, pp. 129-136, 2017
Asociación Iberoamericana de Tecnología Postcosecha, S.C.
Recepción: 02 Agosto 2017
Aprobación: 16 Noviembre 2017
Resumen: La berenjena es un cultivo anual, su periodo de cosecha comienza en primavera (noviembre-diciembre) y culmina en otoño (mayo), ya su requerimiento de alta luminosidad y clima cálido determina la extensión de dicho ciclo. La polinización y posterior desarrollo de semillas, como así también la anatomía celular de los frutos en distintas etapas del crecimiento, son factores de calidad del fruto que podrían ser afectados por los cambios de temperatura durante las estaciones; aunque no existen trabajos hasta el momento en donde se determine dicha relación en berenjena. Por ello, el objetivo del presente trabajo fue evaluar el efecto del estado de crecimiento y estación de cosecha sobre la microestructura y contenido de semillas de berenjena. Se trabajó con berenjena violeta en dos estados de crecimiento definidos por la longitud de los frutos: baby o E1 (9cm) y comercial o E2 (17cm); cosechados en tres estaciones dentro de su ciclo de producción: primavera (Pdiciembre), verano (V-marzo) y otoño (O-mayo). Se analizó la tasa de crecimiento, la anatomía del tejido epidérmico y endocárpico mediante microscopia óptica, y el espesor de la cutícula y contenido de semillas mediante análisis de imágenes (ImageJ). La velocidad de crecimiento disminuyó con el avance de las estaciones, tardándose en O entre 2 y 3 veces más en alcanzar los E1 y E2, respectivamente, que en P y V. En cuanto a la estructura celular, con el avance del crecimiento se observó un aumento del tamaño celular y de los espacios intercelulares. A su vez, los frutos en O presentaron células menos turgentes, con una mayor desorganización y alcanzaron un menor espesor de cutícula. El tamaño de las semillas aumenta con el crecimiento del fruto, no así el número. El número y tamaño de semillas fue mayor en los frutos cosechados en V e intermedio en frutos cosechados en O. Por los resultados observados podemos concluir que la estación de cosecha es un factor determinante. Los frutos cosechados en O fueron los más afectados en su anatomía celular y los cosechados en V presentaron mayor desarrollo de semillas.
Palabras clave: análisis de imágenes, microscopia, ImageJ, epicarpio, endocarpio.
Abstract: The eggplant is an annual crop, its harvest period begins in spring (November-December) and culminates in autumn (May), and the extent of this cycle requirement was determined by high luminosity and warm climate determines. The pollination and subsequent development of seeds, as well as the cellular anatomy of the fruits at different growth stages, are fruit quality factors that could be affected by the temperature changes during the seasons; although no works were found in eggplant. The aim of the present work was to evaluate the effect of growth stage and harvest season on the anatomy and content of eggplant seeds. Violet eggplant was harvested with two growth stages defined by their length: baby or E1 (9 cm) and commercial or E2 (17 cm); harvested at three seasons within its production cycle: spring (December), summer (March) and autumn (May). Several determinations were made: growth rate, anatomy of the epidermic and endocarpic tissues (optical microscopy), cuticle thickness and seed content (image analysis by ImageJ). The growth rate decreased with seasons, taking in Autumn 2 to 3 times more to reach the E1 and E2, respectively, than in spring and summer. An increase in cell size and intercellular spaces was observed with the advance of growth. In turn, fruit harvested in autumn showed less turgid cells, with a greatest disorganization and smallest thickness of cuticle. The seeds size increased with the growth of the fruit, while the seed number remained unchanged. The number and size of seeds were higher in the fruits harvested in summer. While in autumn, the lowest seed growth was observed. Results showed that the harvesting season is a determining factor. Fruit harvested in autumn were the most affected at the cellular anatomy, and fruit harvested in summer showed a high seeds development.
Keywords: image analysis, microscopy, ImageJ, epicarp, endocarp.
INTRODUCCION
La berenjena es un fruto de climas cálidos, con alto requerimiento lumínico. Llega a tolerar temperaturas de hasta 40-45 ºC, aunque la óptima está comprendida entre 23- 25 ºC. A pesar de ser un cultivo anual, hacia el final del ciclo productivo nos encontramos frente a una planta más agotada, con diferente tasa de crecimiento y actividad metabólica; que combinada con la temperatura ambiente puede ser otro factor clave en influir sobre la calidad del fruto, principalmente los aspectos de sabor y firmeza. En general, las semillas son las responsables del sabor amargo de los frutos, por lo cual un contenido excesivo de las mismas no sería de gran aceptación. A su vez, la firmeza es considerada un aspecto clave y determinante de la aceptación por parte del consumidor, ya que sería rechazado tanto un fruto excesivamente blando como rígido.
Los frutos se cosechan generalmente cuando han alcanzado un 80% de su tamaño final, en estado inmaduro (Lawande y Chavan, 1998); aunque actualmente existe una tendencia en el consumo de frutos baby o de crecimiento reducido, aportándole un valor agregado. Los estados de crecimiento influyen sobre la calidad de un fruto, y algunos factores ya fueron estudiados, como lo es el contenido nutricional de los mismos y la firmeza (Zaro y col. 2014). Sin embargo, aún no se ha estudiado cómo se ve afectado el contenido de semillas ni la microestructura del tejido a lo largo del crecimiento del fruto. La polinización es un proceso determinante para el contenido de semillas de un fruto, y la misma depende en gran medida del genotipo, pero también de las condiciones climáticas como la temperatura, la humedad, la energía luminosa y el fotoperiodo (Kowalska, 2008). Así también la estructura celular podría ser afectada por las condiciones climáticas y en consecuencia la firmeza del fruto; más aún en condiciones de bajas temperaturas que podrían causar daño por frio ya en el cultivo. Aun siendo la temperatura de cultivo un factor que juega un rol fundamental sobre los parámetros de calidad mencionados, no existen trabajos hasta la fecha en donde se estudie su efecto en berenjena. Por ello en el presente trabajo se estudió cómo se ve afectada la calidad de frutos de berenjena en dos estados de crecimiento: baby (E1) y comercial (E2) cosechados en tres estaciones desde el inicio hasta el final de su ciclo productivo: primavera, verano y otoño; estudiando específicamente la microestructura como factor determinante de la firmeza, y el contenido de semillas a través del análisis de imágenes como herramienta actual.
MATERIALES Y METODOS
Material vegetal. Se emplearon berenjenas violetas (Solanum melongena L.) cv. Monarca producidas en invernaderos de La Plata (Buenos Aires, Argentina). Frutos recién cuajados se marcaron y midieron los días (days after fruit set o DAFS) necesarios para alcanzar distintos estadios de crecimiento definidos por la longitud de los frutos desde 0,09 a 0,21 m. Se definieron y cosecharon dos estados de crecimiento: baby o Estado 1 (E1: 0,09 m) y comercial o Estado 2 (E2: 0,17 m) para ser estudiados en tres estaciones a lo largo del ciclo productivo: primavera-P (diciembre 2015), verano-V (marzo 2016) y otoño-O (mayo 2016). El rango de temperatura ambiente fue de 14-26 ºC, 14-25 ºC y 5-15 ºC en primavera, verano y otoño, respectivamente. Treinta frutos de cada estado y estación de cosecha fueron cosechados y transportados al laboratorio para ser estudiados. El análisis de imágenes fue empleado en el presente trabajo para analizar la composición de semillas y microestructura del tejido.
Análisis anatómico
Fijación. La cutícula y la anatomía del tejido de berenjena fueron analizadas por métodos de microscopía. Cinco rodajas de frutos distintos fueron cortadas de la zona ecuatorial del fruto y fijadas inmediatamente con FAA70 (formol: ácido acético glacial: etanol 70%) hasta su análisis (Johansen, 1940).
Espesor de cutícula y anatomía del tejido epidérmico. Para el estudio del tejido epidérmico se cortaron secciones longitudinales de la zona más periférica, que incluye a la piel, de las rodajas fijadas (Figura 1). Para la detección de la cutícula se introdujeron los cortes por 15 min en Sudan IV y luego se diferenciaron en etanol 70% por 1 min. Las secciones se montaron en un gel de glicerina. El color rojo indica la presencia de componentes lipofílicos, en este caso la cutícula del fruto. Se usó un microscopio de luz (Gemalux), las imágenes se capturaron con una cámara (Moticam 1000), una amplitud de 100x y 400x y se analizaron mediante el software Image Plus 2.0. El espesor fue expresado en µm.

Figura 1
Imagen de una rodaja de la zona ecuatorial de berenjena en donde se diferencia al tejido epidérmico (1), del endocárpico: externo (2) y central (3)
Anatomía del tejido endocárpico de la pulpa. Secciones del endocarpio de la región externa (a partir de los 8 mm en dirección de la piel hacia el centro, sin incluirlo) y central de la pulpa (“corazón” del fruto) de las rodajas fijadas fueron cortadas manualmente (Figura 1). Dichas secciones se decoloraron, lavaron y montaron en gel de glicerina con 80% de safranina. Se usó el mismo microscopio de luz, cámara y analizador de imágenes anterior, y una magnificación de 100x.
Número y tamaño ocupado por semillas en la rodaja. Para determinar el contenido de semillas quince rodajas de diferentes frutos fueron cortadas de la zona ecuatorial (5 mm de espesor) y escaneadas (Scanner HP DeskJet F4480) 30 min después del corte a fin de permitir el pardeado de las mismas y con ello una mejor detección por parte del software. Se determinó el número y tamaño de semillas, empleando un software de análisis de imágenes (Image J2.0). Los resultados se expresaron en m2 .
Análisis estadístico
Se definió un diseño factorial y los resultados fueron analizados mediante ANOVA. Las medias fueron comparadas mediante el test LSD de Fisher con P<0,05 Empleando el software INFOSTAT.
RESULTADOS Y DISCUSION
Crecimiento de los frutos
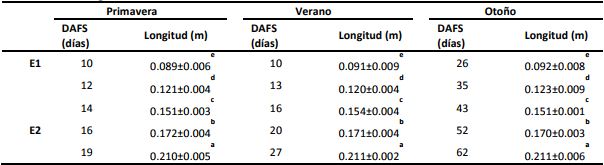
Aunque la berenjena puede ser consumida en sus distintos estados de crecimiento, los productores cosechan a la berenjena cuando han alcanzado un 80% de su tamaño final, siendo éstos los frutos de tamaño comercial. Sin embargo, actualmente podemos encontrar también en el mercado frutos de tamaño pequeño conocidos como baby que le otorgan un valor agregado al producto. En nuestro trabajo definimos a los estados, por su ongitud, en baby o E1 (0,09 m) y comercial o E2 (0,17 m). Ha sido reportado que se tarda entre 15-35 días para alcanzar el estado comercial, pero en el presente trabajo observamos que depende de la estación de cosecha. En P y V se necesitaron entre 10 y 20 DAFS para llegar a los tamaños E1 y E2, respectivamente; mientras que en O se necesitaron 26 y 52 DAFS respectivamente, siendo cerca de 3 veces más lento (Tabla 1). Así podría estimarse que los frutos crecieron cerca de 1 cm/día en P y V, y 0,3 cm/día en el O.
Anatomía celular del tejido epidérmico y endocárpico de los frutos. La anatomía y fisiología de los vegetales puede verse afectada tanto por el propio crecimiento del fruto como por factores característicos de cada estación climática tales como exposición a la luz solar, temperatura ambiente y amplitud térmica. Sin embargo, la anatomía de la berenjena durante el crecimiento del fruto y/o la temporada de cosecha no ha sido descripta previamente. En el presente trabajo, la anatomía de la pulpa de berenjena se evaluó mediante microscopía óptica sobre rodajas de la zona ecuatorial de la fruta. Para más detalle el tejido se diferenció en epidérmico, que comprende epidermis, epicarpio y las primeras capas celulares del endocarpio (Figura 2 A-F); y endocárpico, a partir de la anterior hasta el centro de la rodaja, dividiéndose este último a su vez en: sección externa (zona alrededor de los 8 mm en dirección de la piel hacia el centro) (Figura 2 G-L) y sección interna (endocarpio profundo, en el corazón del fruto) (Figura 2 M-Q).
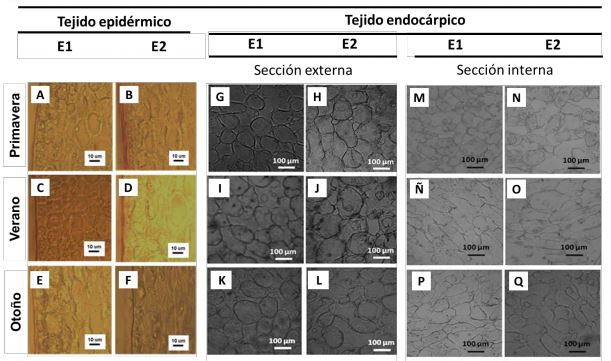
Figura 2
Imágenes de microscopia óptica de tejido epidérmico (A-F) y endocárpico de berenjena seccionado en periférico (A-I) y central (J-Q) en tres los dos tamaños estudiados: baby o E1 y comercial o E2; cosechados en tres estaciones diferentes: Primavera, Verano y Otoño. Magnificación del tejido epidérmico 400X y del endocárpico 100X
Dentro de lo que denominamos tejido epidérmico (Figura 2 A-F) podemos encontrar: la epidermis, constituida por una capa de células redondeadas, empaquetadas y cubiertas por una cutícula de naturaleza lipídica; el epicarpio, el cual se extiende a través de unas pocas capas celulares adyacentes y muestra un aspecto compacto; y por último se visualizan unas primeras capas del endocarpio, en donde las células comienzan a diferenciarse, adquieren una forma más alargada y un mayor tamaño, como así también aparecen moderados espacios intercelulares. En las estaciones P y V (Figura 2A-B y 2C-D) se observó un aumento en el espesor de la cutícula y en el tamaño celular con el crecimiento del fruto. En cambio, en la estación O (Figura 2E-F) el espesor de la cutícula no pareciera modificarse con el crecimiento del fruto, hacia el E2; y a su vez se observó un menor número y tamaño de las células del epicarpio y con cierta pérdida de integridad celular. Este proceso se manifestó también por una deformación y pérdida de globosidad de las primeras capas celulares del endocarpio.
Por otro lado, a medida que avanzamos hacia el centro del fruto, dentro de lo que denominamos tejido endocárpico (Figura 2 GQ), las células se encuentran interconectadas por ramificaciones dando lugar a la formación de espacios intercelulares, que llegan a ser, en la sección interna del fruto, grandes lagunas o cámaras aerenquimáticas debido al gran crecimiento y alargamiento de las células y sus ramificaciones. Por lo que dentro de lo que denominamos sección externa del tejido endocárpico (Figura 2G-L) se observó un tejido más compacto, con células más redondeadas, ramificaciones más cortas y menores espacios intercelulares que los hallados en la sección interna (Figura 2M-Q). Al igual que lo observado para el tejido epidérmico, en el tejido endocárpico tanto externo como interno se observó un incremento en el tamaño celular, como así también en los espacios intercelulares con el crecimiento del fruto para todas las estaciones estudiadas. En P y V, al igual que en el tejido epidérmico, los frutos se comportaron de manera similar, no observándose mayores diferencias de la anatomía celular entre dichas estaciones. Sin embargo, en la estación O se observó una pérdida de la integridad y mayor desorganización celular tanto en el tejido endocárpico externo (Figura 2 K-L) como interno (Figura 2 P-Q). En similitud a lo hallado por otros autores (Zhang y Chen, 2006) a medida que las berenjenas crecen el tejido se torna más esponjoso. Este comportamiento resulta similar al trastorno relacionado con un almacenamiento a baja temperatura observado en los frutos de carozo llamado leatheriness, donde el fruto muestra una textura seca y con alto grado de engrosamiento de la pared celular (Luza y col., 1992; Ju y col., 2001).
Finalmente, podemos decir que la anatomía celular de los frutos cosechados en O se vio afectada por las condiciones estacionales, tanto en el tejido epidérmico como endocárpico externo e interno.
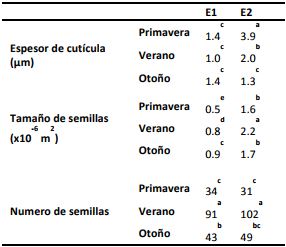
Espesor de cutícula. Cuando el fruto crece en tamaño, la cutícula continúa recubriéndolo a expensa de un continuo proceso de síntesis y deposición de diversos componentes que le otorgará la extensión y rigidez necesaria. Es así que la cutícula podrá engrosarse o no dependiendo de diversos factores, aunque en general tiende a engrosarse a medida que el fruto se desarrolla. Su espesor puede variar entre los 0,5 y 15 µm dependiendo de la especie vegetal y el estado de desarrollo del fruto (Jenks y col., 2002). Sin embargo, no se encuentran estudios sobre los cambios cuticulares durante la ontogenia del fruto de berenjena, lo cual presenta un alto interés para un posterior manejo y calidad de los frutos. En nuestro trabajo observamos un aumento hasta en tres veces del espesor de la cutícula con el crecimiento del fruto, aunque la magnitud dependió de la estación de cosecha estudiada (Tabla 2). Se observó un comportamiento diferencial según la estación de cosecha para los frutos en el estado de crecimiento E2. Así, en P y V el espesor de la cutícula se incrementó entre 3 y 2 veces, respectivamente, desde el fruto E1 al E2 (Tabla 2); mientras que, en la estación O el espesor de la cutícula se mantuvo constante en un valor aproximado de 1,4 µm (Tabla 2). Estas variaciones se evidencian claramente en las micrografías de los frutos de la Figura 2 A-F. Se ha reportado que la síntesis de compuestos cuticulares puede ser afectada por factores ambientales (Jetter y col., 2006; Stark y Tian, 2006) ya que las células epidérmicas requiere gran cantidad de energía y la coordinación de actividad de numerosas enzimas para la biosíntesis de la cutícula (Bargel y col., 2006; Yeats y col., 2010; Tafolla-Arellano y col., 2013). Así, durante el O las bajas temperaturas reinantes podrían ser las responsables del menor espesor de cutícula, debido a una menor biosíntesis de la misma.
Contenido de semillas. A través del análisis de imágenes se calculó el número y tamaño de las semillas.
El tamaño de las semillas se incrementó con el crecimiento del fruto, siendo la estación de cosecha un factor determinante para evaluar dicho parámetro. El tamaño de semilla en frutos E2 fue un 75% mayor en V que en P y O (Tabla 2 y Figura 3), indicando que la disponibilidad de nutrientes fue más eficiente en esta estación de cosecha.
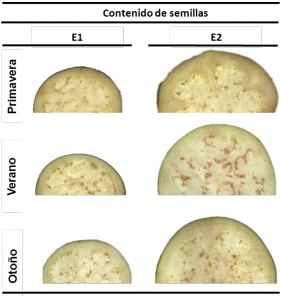
Figura 3
Imágenes de rodajas de berenjena baby o E1 y comercial o E2 cosechadas en tres estaciones diferentes: Primavera, Verano y Otoño
El número de semillas se mantuvo constante a lo largo del crecimiento del fruto, lo cual se condice con el hecho de que, en general, el número de semillas es definido por la polinización al momento del cuajado del fruto (Kowalska, 2008). El número de semillas fue entre 3 y 2 veces mayor en V respecto de P y O, respectivamente (Tabla 2 y Figura 3). Esta marcada diferencia puede deberse a que tanto el tamaño como número de las semillas están relacionados con la polinización y fertilización de las flores, procesos que se ven favorecidos con temperaturas ambientales templadas (Kowalska, 2008; Polverente y col., 2005; Lawande y Chavan, 1998) como las registradas en el V. El número de semillas de frutos cosechados en O fue intermedio entre P y V (Tabla 2 y Figura 3), debido tal vez a que las temperaturas ambientales son aún elevadas al inicio de esta estación, tiempo al cual debiera sucederse la polinización y fertilización. En resumen, en la estación V se obtuvieron frutos con mayor tamaño y número de semillas. En la estación O, los frutos presentaron un número intermedio de semillas.
CONCLUSION
El crecimiento del fruto conlleva cambios anatómicos, los cuales fueron afectados por la estación del año en la que crecen. Así, frutos de tamaño comercial E2 cosechados en Otoño presentaron una cutícula más delgada, un menor número y tamaño de las células del epicarpio, y pérdida de la integridad y mayor desorganización celular tanto en el tejido endocárpico externo como interno.
Mientras que, tanto el tamaño como número de semillas fue significativamente mayor en frutos de tamaño comercial E2 cosechados en la estación Verano, y con un número intermedio de semillas aquellos cosechados en Otoño.
Referencias
Bargel,H., Koch, K., Cerman, Z. y Neinhuis, C.2006. Structure–function relationships of the plant cuticle and cuticular waxes—a smart material? Funct. Plant Biol. 33:893- 910.
Jenks, M., Eigenbrode, S. y Lemieux, B. 2002. Cuticular waxes of Arabidopsis. J. Cell Biol. 17: 208-212.
Jetter , R., Kunst, L. y Samuels, L. 2006. Composition of plant cuticular waxes. In: Biology of the Plant Cuticle. M Riederer, C Müller (eds). Julius-von-Sachs-Institut, für Biowissenschaften Universität Würzburg, Germany. pp:145-175.
Johansen, D. A. 1940. Plant microtechnique. McGraw-Hill Book Company, Inc: London; 530p.
Ju, Z., Duan, Y., Ju, Z. y Guo, A. 2001. Different responses of ‘Snow Giant’ and ‘Elegant Lady’ peaches to fruit maturity and storage temperature. Journal of Horticultural Science e Biotechnology, v. 76, n. 5,p. 575- 580.
Kowalska, G. 2008. Flowering biology of eggplant and procedures intensifying fruit set. Review. Acta Sci. Pol., Hortorum Cultus 7(4), 63-76
Lawande, K. E. y. Chavan, J.K. 1998. Eggplant (Brinjal). Salunkhe D. K., Kadam S. S., (red.), Handbook of vegetable science and technology. Production, composition, storage and processing. New York, 225– 244.
Luza, J.G., Van Gorsel, R., Polito, V.S. y Kader, A.A. 1992. Chilling injury in peaches: a cytochemical and ultrastructural cell Wall study. J Am Soc Hortc Sc. 117, 114–118.
Polverente M. R., Fontes D. C. Cardoso A. I. I., 2005. Produção e qualidade de sementes de berinjela em função do horãrio de polinização manual. Bragantia, Campinas 64, 3, 467–472.
Stark, R. y Tian, S. 2006. Thecutin biopolymer matrix: In: Biology of the Plant Cuticle. M Riederer, C Müller (eds). Julius-von-SachsInstitut, für Biowissenschaften Universität Würzburg, Germany. pp: 126-141.
Tafolla-Arellano, J. C., González-León, A., Tiznado-Hernández, M.E., Zacarías García, L. y Báez-Sañudo, R. 2013. Composición, fisiología y biosíntesis de la cutícula en plantas. Revista Fitotecnia Mexicana, 36(1), 3-12.
Yeats, T.H., Howe, K.J., Matas, A.J., Buda, G.J., Thannhauser, T.W. y Rose J.K.C. 2010. Mining the surface proteome of tomato (Solanum lycopersicum) fruit for proteins associated with cuticle biogenesis. J. Exp. Bot. 61:3759-3771.
Zaro, M.J., Keunchkarian, S., Chaves, A.R., Vicente, A.R. y Concellón A. 2014. Changes in bioactive compounds and response to postharvest storage conditions in purple eggplants as affected by fruit developmental stage.Postharvest Biology and Technology, 96, 110-117.
Zhang, M., Chen, D.W., 2006. Effects of low temperature soaking on color andtexture of green eggplants. Journal of Food Engineering 74 (1), 54–59