Artículos

Esta obra está bajo unaLicencia Creative Commons Atribución-NoComercial-CompartirIgual 4.0 Internacional.
Received: 14 June 2023
Accepted: 18 October 2023
DOI: https://doi.org/10.37176/iea.25.1.2024.837
Resumen: Durante las primeras excavaciones realizadas en el Parador QB (Parque Nacional Islote Lobos, costa Rionegrina) se recuperaron huesos de aves en los conjuntos zooarqueológicos. Al igual que en otras localidades arqueológicas del litoral Nordpatagónico, la representación de este taxón es escasa, aunque los huesos presentan buen estado de preservación. Con el objetivo de explorar el papel de las aves en la subsistencia de las sociedades cazadoras-recolectoras que ocuparon este litoral durante el Holoceno tardío, desarrollamos un estudio tafonómico para establecer si la depositación de sus huesos se debe a la acción antrópica o a causas naturales. La evaluación tafonómica de los restos avifaunísticos es fundamental dado que actualmente existen áreas de nidificación de diversas aves marino-costeras próximas a los sitios arqueológicos, por lo que sería factible la “contaminación ósea” -o mezcla de huesos naturales y materiales arqueológicos. Se concluye que los huesos de aves de los tres loci tienen trayectorias tafonómicas similares, mientras que la mezcla de restos óseos de diferente origen se registró principalmente en el caso de los pingüinos de Magallanes. Por último, señalar que, aunque las aves no son tan abundantes, tuvieron un lugar importante en el repertorio de presas de los cazadores-recolectores de la región.
Palabras clave: Tafonomía de aves, Restos avifaunísticos, Costa nordpatagónica, Cazadores-recolectores.
Abstract: During the first excavations carried out at the Parador QB (Islote Lobos National Park, Rio Negro coast), bird bones were recovered in the zooarchaeological assemblages. As in other archaeological localities on the North Patagonian coast, the representation of this taxon is scarce, although the bones present a good state of preservation. With the aim of exploring the role of birds in the subsistence of the hunter-gatherer societies that occupied this coastline during the Late Holocene, a taphonomic study was developed to establish whether the deposition of bird bones was due to anthropogenic action or to natural causes. The taphonomic evaluation of the avifauna remains is essential given that currently there are nesting areas of various marine-coastal birds close to the archaeological sites, so that "bone contamination," or the mixing of natural bones and archaeological materials, can result. It is concluded that bird bones from the three loci have similar taphonomic trajectories, while modern bones mixed with the archaeological materials were mainly of Magellanic penguins. Finally, it should be noted that, although bird bones are not abundant, birds had an important place in the prey repertoire of the hunter-gatherers of the region.
Keywords: Avian taphonomy, Bird bone remains, Northpatagonian coast, Hunter-gatherers.
INTRODUCCIÓN
En las últimas décadas, con el aumento de las investigaciones zooarqueológicas en la costa atlántica de Patagonia continental, se comenzaron a desarrollar estudios sistemáticos sobre restos de aves recuperados en contextos arqueológicos. El objetivo general de estos estudios fue explorar su papel en la subsistencia de las sociedades cazadoras-recolectoras durante el Holoceno tardío (Cruz, 2003, 2006; Moreno, 2008; Borella y Cruz, 2012; Marani y Borella, 2014; Zubimendi y Beretta, 2018; Beretta y Zubimendi, 2019; Cruz et al., 2021, entre otros). Como han señalado varios investigadores, en términos generales la representación de las aves en el registro zooarqueológico a nivel mundial es comparativamente escasa y fragmentaria con respecto a la de otros vertebrados, lo que sin duda dificultó su interpretación y, en algunos casos, desalentó el desarrollo de estudios específicos (ver Serjeantson, 2009 para una síntesis al respecto). No obstante, si se quiere establecer por qué los cazadores-recolectores las eligieron como presas o los escenarios en los cuales fueron explotadas, es necesario conocer las características de los conjuntos avifaunísticos, aunque sean numéricamente pequeños.
La situación presentada por Serjeantson (2009) se repite en el caso de los registros arqueológicos de la Patagonia continental Argentina, ya que el análisis generalmente se ha restringido a la evaluación de los restos de unas pocas especies particulares (de las Familias Rheidae, Phalacrocoracidae y Spheniscidae, entre las más abordadas) (Fernández, 2000; Cruz, 2006; Moreno, 2008; Borella y Cruz, 2012, entre otros). Además, en muy pocos casos se efectuó una evaluación tafonómica destinada a dilucidar las condiciones de formación del registro óseo de aves (e.g., Cruz, 2007; Borella y Cruz, 2012, entre otros). El esqueleto de las aves difiere del de otros vertebrados en varios aspectos, especialmente los relacionados con la principal adaptación locomotriz del taxón, el vuelo (Bellairs y Jenkin, 1960; Gilbert et al., 1996, entre otros), lo cual puede tener consecuencias tafonómicas (e.g., sobre representación taxonómica y anatómica, Cruz, 2005, 2009a, 2009b, 2011, 2015).
Un aspecto metodológico reiteradamente mencionado por varios investigadores al abordar este tema, tanto a nivel mundial como regional, es que para indagar el papel de las aves en la subsistencia humana es necesario evaluar la integridad (sensu Binford, 1981) de los conjuntos recuperados en sitios arqueológicos (Ericson, 1987; Livingston, 1989; Higgins, 1999; Bovy, 2002; Cruz, 2003, 2006, 2011; Broughton et al., 2007; Borella y Cruz, 2012, entre otros). En este sentido, durante las últimas décadas se han efectuado estudios de tafonomía naturalista (sensu Marean, 1995) sobre huesos de aves en la costa de Patagonia continental, dedicados a generar herramientas interpretativas (Cruz, 2003, 2009a, 2011, 2015, entre otros) y, de esta manera, avanzar en el conocimiento sobre estos temas. En este marco, nuestro objetivo es presentar el análisis de los restos de aves de tres muestras arqueológicas procedentes de Parador QB, Parque Nacional Islote Lobos (PNIL), en la costa oeste del Golfo San Matías (GSM), provincia de Río Negro (Figura 1). En la actualidad, aquí, como en toda Patagonia, hay una gran abundancia de aves costero-marinas. El mar Patagónico es una de las áreas de mayor productividad biológica del hemisferio sur, que permite sustentar una notable diversidad de especies de mamíferos y aves marinas (Foro para la Conservación del Mar Patagónico y Áreas de Influencia, 2008; Yorio et al., 2015). En tal sentido, a lo largo de su costa nidifican numerosas especies que forman agrupaciones reproductivas en áreas relativamente acotadas (Frere et al., 2005; Schiavinni et al., 2005; Bertellotti et al., 2015; Reyes y García Borboroglu, 2020, entre otros). Esta abundancia y diversidad actual de las aves y la escasez de sus restos en los registros arqueológicos costeros de la región generan preguntas sobre las posibilidades de preservación de sus huesos y acerca de su disponibilidad pasada. Por lo tanto, nuestro análisis se centrará en los aspectos tafonómicos, para luego discutir algunos puntos relativos a las interacciones entre aves y poblaciones humanas durante el Holoceno tardío en este sector costero de Nordpatagonia.
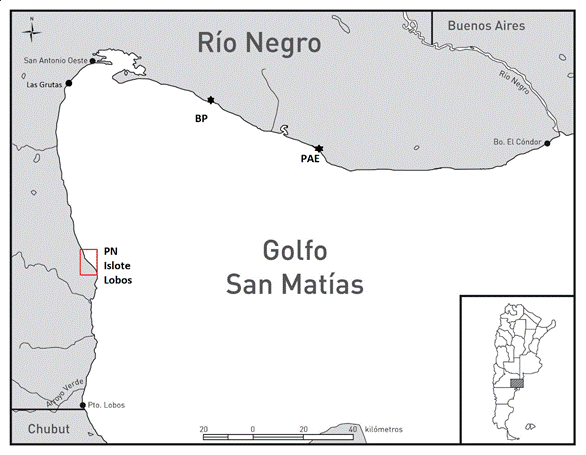
Figura 1
Mapa con ubicación del Parque Nacional Islote Lobos y de las localidades arqueológicas Bajada de los Pescadores (BP) y Paesani (PAE) mencionadas en este trabajo.
UBICACIÓN Y ASPECTOS AMBIENTALES
El Parque Nacional Islote Lobos se ubica en el tramo central de la costa oeste del golfo San Matías, en la provincia de Río Negro, Argentina. Este sector del GSM corre de norte a sur y sus principales rasgos geomorfológicos están constituidos por acantilados y serranías, particularmente en su tramo meridional (Figura 1).
Aquí se destaca la presencia de la meseta basáltica de Somuncurá, una planicie estructural lávica que desciende hacia el mar gradualmente en forma de pedimentos de flanco (González Díaz y Malagnino, 1984). El área Islote Lobos se caracteriza por presentar seis islotes graníticos que constituyen una prolongación de los afloramientos del Complejo Plutónico Sierra Grande que irrumpe en la costa y que han resistido la acción erosiva del mar. Por detrás de estos islotes se han desarrollado tómbolos (Gelós et al., 1992; Kokot y Pisani, 2011) que los conectan con el continente solo en bajamar. Estos islotes son los únicos de todo el litoral rionegrino, en ellos actualmente nidifican numerosas aves marinas (Vinci, 2004; Reyes y García Borboroglu, 2020).
Al igual que toda la costa rionegrina, esta región posee un clima templado semiárido, con una temperatura media de 12º C, vientos que soplan principalmente del cuadrante oeste, un promedio anual de precipitaciones menor a 250 mm y una vegetación de tipo xerófila (Favier Dubois y Borella, 2011).
Actualmente en el área nidifican numerosas aves costeras, marinas y terrestres, pero también es posible hallar numerosos mamíferos silvestres característicos de la región (Vinci, 2004; Reyes y García Borboroglu, 2020).
El sector Parador QB (P-QB), de donde provienen las muestras que presentamos y analizamos en este trabajo, se encuentra en el continente, por detrás de la línea de médanos litorales parcialmente edafizados, a la altura del islote La Pastosa (Figura 2A), el mayor de los islotes del Parque (Borella et al., 2020).

Figura 2
A: sector de Parador-QB dentro del Parque Nacional Islote Lobos y B: detalle de la localización de los tres loci P-QB.
EN REGISTRO AVIFAUNÍSTICOS DE LA COSTA RIONEGRINA
El registro zooarqueológico recuperado hasta el momento en el golfo San Matías (provincia de Río Negro) se caracteriza por presentar abundantes restos de especies marinas, acompañadas, en menor medida, por otras terrestres (Borella et al., 2011; Marani, 2018; Scartascini, 2020, entre otros). Esta tendencia general se cumple también con los restos de aves. Sin embargo, en la excavación de dos sitios concheros o conchales denominados Bajada de los Pescadores 2 y Paesani 5 (localizados en la costa norte del GSM; Figura 1), las aves tienen una representación relevante. En el primer caso se registraron huesos correspondientes a cinco pingüinos patagónicos asociados a restos óseos de otáridos y peces con una antigüedad de 2200 años AP (Borella y Cruz, 2012), mientras que en el segundo se recuperaron restos de doce tinámidos en un conjunto en el que predominan los huesos de peces y de pequeños mamíferos, en un contexto fechado en 1100 años AP (Marani y Borella, 2014). En Bajada de los Pescadores 2 se realizó una exhaustiva evaluación del origen de los huesos de pingüinos allí recuperados mientras que el fechado taxón confirmó la contemporaneidad de los huesos de pingüinos y los demás depósitos del conchero, lo cual permitió concluir que el descarte respondía a actividades antrópicas. En Paesani 5, la notable cantidad de huellas de corte registrada en los restos de tinámidos fue concluyente respecto al origen antrópico de los huesos de aves recuperados en ese depósito. Sin embargo, sobre más de 70 sondeos excavados en toda la costa del Golfo San Matías -donde principalmente predominan los restos de recursos marinos como peces, lobos y moluscos-, el registro de aves resulta escaso.
Las investigaciones arqueológicas realizadas hasta la fecha en el sector denominado Parador-QB (Figura 2A) en el Parque Nacional Islote Lobos (PNIL), señalaron la presencia de abundantes restos de aves en el registro zooarqueológico (Borella et al., 2020). A diferencia de la costa norte -donde en general los materiales zooarqueológicos fueron recuperados durante la excavación de concheros expuestos, semideflacionados-, los depósitos arqueológicos del sector P-QB -que analizamos en este trabajo- se encontraban sepultados y fueron detectados mediante pruebas de pala. Estos materiales incluían delgadas acumulaciones de moluscos, carbones, restos óseos en buen estado de preservación, material lítico y escasos tiestos cerámicos. La obtención de abundantes carbones en estratigrafía permitió realizar dataciones radiocarbónicas en los tres loci (Figura 2B) que señalaron en P-QB C1 una edad de 1280 ± 60 años AP; en P-QB C2 de 1460 ± 90 años AP y en P-QB C3 de 1000 ± 50 años AP (Borella et al., 2020).
Este conjunto bien preservado permite entonces explorar el papel de las aves entre los grupos humanos que ocuparon la costa oeste del Golfo San Matías, aunque requiere de una adecuada evaluación del origen de los conjuntos, previa a dar cualquier interpretación acerca de la forma en que fueron aprovechados en el pasado (¿consumidos? ¿empleados como materia prima?).
MATERIALES Y METODOS
Características de las muestras estudiadas
Las muestras aquí analizadas provienen de la excavación de tres cuadrículas de similar superficie (1 m.), emplazadas en diferentes loci espacialmente próximos y no contiguos, denominados P-QB C1, P-QB C2 y P-QB C3 (Figura 2B). En la cuadrícula 1 (0,45 m.) se recuperó un total de 630 especímenes óseos; 2405 en la cuadrícula 2 (0,35 m.) y 1425 en la cuadrícula 3 (0,40 m.). En estos conjuntos zooarqueológicos predominan los restos de peces, seguidos por los de otáridos, aves y guanacos, mientras que en muy escasa proporción se registraron restos de vertebrados de tamaños mediano y pequeño (Borella et al., 2020). En función del buen estado de preservación ósea en general (Borella y L’Heureux, 2021) se decidió plantear un estudio particular para los restos de aves.
Aunque la representación de las aves es variable en cada uno de los depósitos (9% en los conjuntos de P-QB C1 y P-QB C3, y 5% en P-QB C2), resulta mayor que la del guanaco en las cuadrículas 2 y 3, y que mamíferos como armadillos y roedores en las tres cuadrículas (Borella et al., 2020). Esto tiene implicaciones con respecto a la subsistencia humana, ya que da cuenta de la importancia que pudieron tener las aves como presas de los cazadores-recolectores. Asimismo, tiene alcances desde el punto de vista de la formación de los depósitos debido a que, en general, el potencial de preservación de los huesos de aves es menor que el de los mamíferos (Cruz, 2003, 2015, entre otros).
Aspectos metodológicos
El análisis de los huesos de aves en el laboratorio se efectuó a partir de los lineamientos usualmente utilizados en el estudio de los conjuntos zooarqueológicos (Lyman, 1994; Serjeantson, 2009; Gifford-González, 2018). Las determinaciones taxonómicas se efectuaron a través del empleo de las colecciones osteológicas comparativas del Instituto de Investigaciones Arqueológicas y Paleontológicas del Cuaternario Pampeano - Consejo Nacional de Investigaciones Científicas y Tecnológicas. Además, se utilizaron atlas osteológicos como el de Gilbert y colaboradores (1996) y France (2017). En el caso de los restos de Rheidae se consultó el atlas osteológico de Giardina (2010), la anatomía económica propuesta por ese mismo autor (Giardina 2006), mientras que para la identificación de fragmentos óseos se aplicaron los rasgos cualitativos particulares descriptos por León (2010) para la cavidad medular. Para el análisis de las variables tafonómicas, los huesos de aves se agruparon en las siguientes categorías basadas en las propiedades intrínsecas de los huesos y la estructura anatómica: Phalacrocoracidae (o cormoranes, incluye Leucocarbo atriceps), Spheniscidae (o Spheniscus magellanicus), Rheidae, aves voladoras y aves indeterminadas. Entre las aves voladoras se incluyeron aquellos restos de aves que no pudieron ser identificados a nivel específico, pero cuyas características permiten establecer que comparten este modo de locomoción. Por lo tanto, esta categoría podría incluir especies frecuentes de Patagonia continental (Familias Anatidae, Tinamidae, Laridae, Procellariidae, Podicipedidae, Ardeidae, entre otras), excepto Spheniscidae y Rheidae. Se caracterizó como restos de aves indeterminadas a los que pueden corresponder a cualquiera de los grupos (incluyendo Spheniscidae), pero no Rheidae. Sin embargo, en todos los casos en que fue posible, se afinaron las determinaciones taxonómicas al nivel de especie, género o familia, ya que se considera que es lo más adecuado y útil para el análisis zooarqueológico.
En la cuantificación de los especímenes se utilizaron el NISP (Number of Identified Specimens) y el MNI (Minimal Number of Individuals), en ambos casos se siguió la propuesta de Lyman (1994). Estas dos medidas de abundancia taxonómica son las apropiadas para el análisis tafonómico y zooarqueológico. Se relevaron las siguientes modificaciones óseas en función de los criterios morfológicos indicados en la literatura: estadios de meteorización, abrasión sedimentaria, tipos de fracturas, daños por carnívoros y roedores, trazas de raíces, huellas de corte y alteraciones térmicas (Binford, 1981; David, 1990; Lyman, 1994; Behrensmeyer et al., 2003; Serjeantson, 2009; Gifford-González, 2018). Por último, mencionar que denominamos marcas indeterminadas a aquellas modificaciones superficiales cuyas morfologías no permiten la asignación no ambigua a un determinado agente o proceso específico.
Para estimar la representación de partes esqueletarias de aves se calculó el MNE, para lo cual se consideró el grado de integridad de los elementos, las porciones anatómicas presentes y la posibilidad de ensamblaje. Luego los elementos fueron agrupados en las siguientes unidades anatómicas (Cruz, 2005): a) esqueleto axial: cráneo, mandíbula, maxilar, vértebras, costillas y esternón; b) cintura escapular: escápula, coracoides, fúrcula; c) extremidades anteriores: húmero, radio, ulna, carpometacarpo, falanges anteriores; d) cintura pélvica: pelvis y sinsacro; y e) extremidades posteriores: fémur, tibiatarso, fíbula, rótula, tarsometatarso, falanges posteriores.
Marco tafonómico para el análisis de los restos de aves
Para interpretar las características de los conjuntos avifaunísticos se utilizan los resultados de observaciones tafonómicas de restos actuales de aves en el sur de Patagonia, así como de estudios efectuados en distintos lugares del mundo sobre las propiedades intrínsecas de los huesos de estos vertebrados. Los estudios actuales incluyen observaciones sobre la depositación natural de huesos de diferentes aves, su distribución y abundancia en ambientes costeros de Patagonia (Cruz, 2003, 2011, 2015), incluidas áreas de nidificación de pingüinos Patagónicos (Cruz, 2007) y de cormoranes imperiales (Cruz, 2009b). Se consideraron también diversos estudios sobre restos de ñandú patagónico en la costa y el interior de la región (Belardi, 1999; Cruz, 2003, 2011, 2015; Cruz y Muñoz, 2020). La meteorización de huesos de aves fue analizada a partir de diversos trabajos (Behrensmeyer et al., 2003; Cruz, 2003, 2007, 2014, 2015).
Los estudios sobre propiedades intrínsecas de huesos de aves son los referidos a la densidad mineral ósea de Rhea pennata (Cruz y Elkin, 2003), cormoranes y anátidos (Higgins, 1999; Broughton et al., 2007). También se tuvieron en cuenta las consideraciones de Ksepka y colaboradores (2015) sobre las características de los huesos de pingüinos, de Bellairs y Jenkins (1960) sobre los correspondientes a aves voladoras y de Drenowatz (1995) para los de ñandúes. La representación de partes esqueletarias de aves se analizó según las propuestas de varios investigadores: aves voladoras (Livingston, 1989; Bovy, 2002; Cruz, 2005, 2011; Lefèvre y Laroulandie, 2014), cormoranes (Cruz, 2009b), pingüinos (Cruz, 2005, 2007, 2015) y ñandúes (Belardi, 1999; Cruz, 2003, 2005, 2015; Cruz y Muñoz, 2020).
RESULTADOS
En los conjuntos óseos de aves de P-QB se identificaron tanto especies costero-marinas como terrestres (Tabla 1). Entre las primeras, en los tres conjuntos se recuperaron especímenes correspondientes a pingüinos Patagónicos y cormoranes. En este último caso se identificaron especímenes tanto a nivel de especie (Leucocarbo atriceps), como de Familia (Phalacrocoracidae). Los restos de estas aves son los más abundantes en P-QB C2 y P-QB C3. Además, en P-QB C2 se recuperaron dos especímenes de ostrero (Haematopus sp.) -que corresponden a un único tibiatarso- y un húmero de anátido indeterminado. Con respecto a las aves terrestres, se registraron especímenes de Rheidae, que previamente solo había sido identificada a partir de fragmentos de cáscaras de huevos (Borella et al., 2020). Además, en P-QB C1 y P-QB C3 también se determinaron huesos de martineta.
Diversidad y abundancia taxonómica (NISP, %NISP, MNI) de los restos de Aves de Parador QB (Parque Nacional Islote Lobos, Río Negro, Argentina). *los elementos correspondientes a Phalacrocoracidae son complementarios de aquellos con los que se determinó el MNI de L. atriceps, por lo que no se calculó el número de individuos para esta categoría.
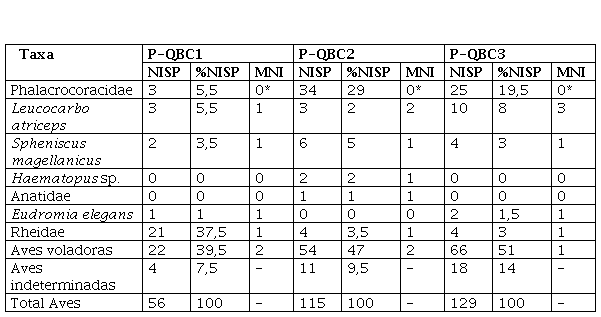
En los tres conjuntos, el grupo más representado en términos del NISP es el de las aves voladoras. Los huesos de cormoranes son los más numerosos entre aquellos determinados en algún nivel taxonómico. La mayor parte de los huesos de cormoranes están fusionados, lo cual indica la presencia de adultos: el 100% de los especímenes en P-QB C1, el 73% en P-QB C2 y el 86% en P-QB C3, mientras que no fue posible determinar las edades de los especímenes restantes en los dos últimos casos.
Si bien en términos del NISP la muestra de Rheidae correspondiente a P-QB C1 es una de las más abundantes (37,5% del conjunto avifaunístico), esto es debido a que está totalmente fragmentada, principalmente compuesta por pequeños fragmentos diafisiarios de huesos largos (NISP = 15) (Figura 3). Sin embargo, el NISP no implica una mayor cantidad de elementos (MNE) o de individuos (MNI) representados. En este sentido, si se comparan los valores de MNI de los tres conjuntos, la representación taxonómica en P-QB C1 es de un individuo por grupo o taxón, con excepción de las aves voladoras indeterminadas que presentan un MNI de dos (Tabla 1).

Figura 3
Fragmentos diafisarios de hueso largo de Rheidae con modificaciones antrópicas (fracturas) y termoalterado.
En P-QB C2 el MNI es de dos individuos para cormoranes (calculado sobre la base del húmero izquierdo) y aves voladoras, mientras que para el resto se determinó un único individuo por caso. Por último, en P-QB C3 los cormoranes están representados por tres individuos (en función del coracoides izquierdo) y el resto de los grupos o taxones por un único individuo. Entonces, dadas las características de las muestras, los valores de MNI son los que permiten una mejor aproximación a la abundancia taxonómica, mientras que, en muchos de los casos, las diferencias en la representación del NISP se relacionan con la mayor o menor fragmentación (Tabla 2). Entre los huesos de Phalacrocoracidae se registraron especímenes completos o casi completos, debido a lo cual se pudieron efectuar identificaciones taxonómicas a nivel de especie (L. atriceps). En el caso de las aves voladoras, hay varios elementos completos que, por no tratarse de huesos diagnósticos, fueron determinados a nivel de género (Haematopus sp.) o de Familia (Anatidae). En síntesis, si se consideran en conjunto las tres muestras, los cormoranes y las aves voladoras son las más representadas tanto en términos del NISP como del MNI.
Modificaciones naturales y antrópicas (%NISP) en los conjuntos óseos de P-QB (Parque Nacional Islote Lobos, Río Negro, Argentina). Referencias: Ph: Phalacrocoracidae; Sp: Spheniscidae; Rh: Rheidae; AV: aves voladoras. *En P-QBC1 y P-QBC2 la categoría aves voladoras incluye los especímenes de E. elegans.
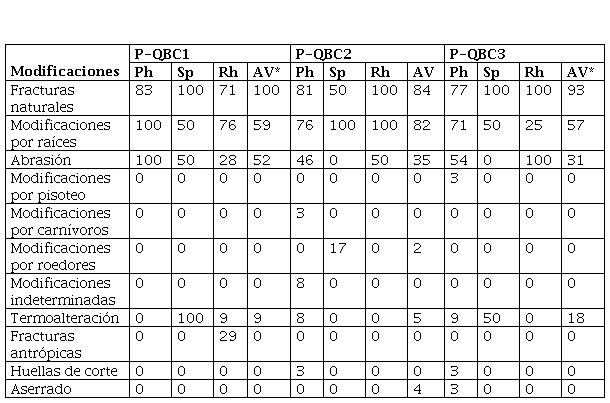
En la Tabla 2 se presentan las modificaciones naturales observadas en los huesos de cada grupo de aves para los tres loci. Se puede observar que tres variables -las modificaciones por raíces (Figura 4A y B), las fracturas naturales y la abrasión- son las que presentan los valores más altos en las tres cuadrículas. Aunque no se descarta que el ambiente de depositación -donde se recuperaron los restos óseos de aves- haya generado algún grado de abrasión (Serjeantson, 2009), se considera que las fracturas naturales y la abrasión pueden vincularse -al menos en parte- a la acción de pisoteo del ganado introducido en tiempos históricos (vacas y ovejas) (Borella et al., 2020).
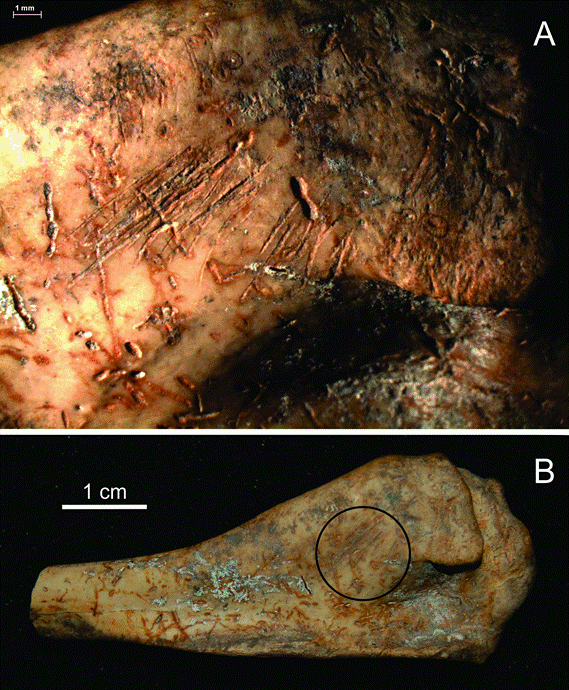
Figura 4
A) Sección del fragmento proximal de húmero de cormorán donde se registraron conspicuas huellas de corte y B) vista general del extremo proximal de húmero con la localización de las huellas de corte. En ambas fotos pueden observarse las modificaciones por raíces.
En cuanto a otras modificaciones naturales (Tabla 2), se registraron en muy bajos porcentajes y no afectaron a todos los conjuntos. En este sentido, en P-QB C2 se observaron las únicas modificaciones por carnívoros (perforaciones en un 3%), en huesos de Phalacrocoracidae. En esta misma cuadrícula también se observaron las producidas por roedores en huesos de aves voladoras (2%) y de Spheniscidae (17%), así como modificaciones indeterminadas en huesos de Phalacrocoracidae (8%). La presencia de estas trazas da cuenta de una historia de formación levemente más compleja de esta cuadrícula. Por su parte, en P-QB C3 el registro de modificaciones por pisoteo en huesos de Phalacrocoracidae (3%), apoyan lo afirmado con respecto al origen de las fracturas y la abrasión en los tres conjuntos.
Respecto de los perfiles de meteorización de los grupos de aves de cada locus de P-QB (Tabla 3), todos muestran el predominio de los huesos no meteorizados y levemente meteorizados (estadio 1), mientras que en ningún caso se registraron los estadios más altos (4 y 5). En este sentido, las numerosas fracturas naturales no están asociadas al desarrollo de este proceso, lo que estaría acorde con que el principal proceso responsable de la fragmentación de los huesos de aves sería el pisoteo del ganado.
Perfiles de meteorización (%NISP) en los conjuntos óseos de P-QB (Parque Nacional Islote Lobos, Río Negro, Argentina). Referencias: Ph: Phalacrocoracidae; Sp: Spheniscidae; Rh: Rheidae; AV: aves voladoras. *En P-QBC1 y P-QBC2 la categoría aves voladoras incluye los especímenes de E. elegans.
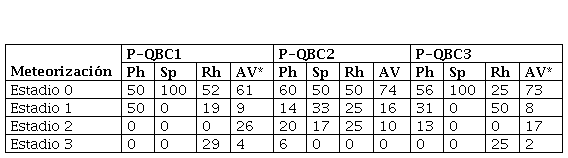
El perfil de meteorización de los especímenes de Rheidae en P-QB C1 y C3 incluye más de un 25% en estadio 3. Aunque algunos huesos de ñandú pueden meteorizar tan rápidamente como los de otras aves, los correspondientes a las extremidades posteriores y cintura pélvica meteorizan más lentamente y pueden alcanzar los estadios más altos en algunos casos (Cruz, 2003, 2014, 2015; Cruz y Muñoz, 2020). Al comparar los perfiles de meteorización de Rheidae y aves voladoras, se hacen visibles algunas incongruencias con lo esperado en función de las observaciones actuales. Si los huesos de ambos grupos de aves fueron depositados simultáneamente, entonces los huesos de Rheidae deberían presentar un perfil más orientado a una mayor representación de los estadios más bajos. Como esto no ocurre, puede proponerse que los huesos de aves voladoras se depositaron tiempo después que los de ñandúes o, alternativamente, que la gran fragmentación de los huesos de Rheidae promovió un mayor avance de la meteorización. Dada la buena preservación de los conjuntos y los depósitos (Borella et al., 2020), consideramos que esta última alternativa es la más plausible. En la Tabla 3 también se puede observar que, aunque con algunas diferencias, los perfiles de meteorización de huesos de aves voladoras y de Phalacrocoracidae tienen una tendencia similar. Lo mismo ocurre con los huesos de aves voladoras.
En cuanto a la termoalteración, se trata de una modificación que aparece en escasa proporción en algunos especímenes de aves voladoras, Spheniscidae y de Rheidae de todas las cuadrículas (Tabla 2). En general se trata de fragmentos total (Figura 3) o parcialmente quemados (Figura 5A y C), con tonalidades marrón y negro, pero en ningún caso se registró carbonización -gris- o calcinación -blanco-azulado- de los fragmentos óseos (David, 1990; Serjeantson, 2009).
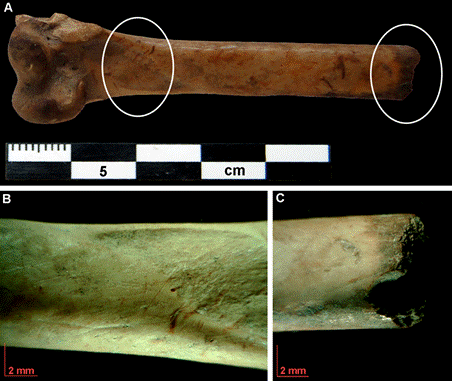
Figura 5
Esta modificación es ambigua con respecto al agente de depositación, por lo que, en ausencia de otras trazas asignables a humanos en los restos de estas aves, es difícil sostener su origen antrópico (ver en Discusión).
Con respecto a las trazas de origen cultural, en P-QB C1 no se registraron modificaciones de este tipo, con excepción de las fracturas antrópicas en huesos de Rheidae (Figura 3), mientras que en las otras dos cuadrículas las modificaciones incluyen huellas de corte y especímenes con aserrado perimetral (sensu Acosta, 2000).
En P-QB C2 dos especímenes de aves voladoras -una ulna y una diáfisis indeterminada- presentan aserrado perimetral (Figura 6), y además se registraron numerosas huellas de corte en un húmero de Leucocarbo atriceps (Figura 4A) que indican procesamiento. Entre los restos óseos de cormoranes de P-QB C3 se destaca un radio con huellas de corte y en un fragmento de húmero distal en cuya diáfisis (porción medial) se registraron huellas de corte (Figura 5A y B). Este espécimen además presenta una fractura transversal donde se observan dos modificaciones en forma de muescas opuestas, asociadas a termoalteración, que podrían indicar que se trata de un remanente de la manufactura de algún artefacto (Figura 5C). En síntesis, las modificaciones claramente antrópicas de los tres loci son escasas pero significativas, corresponden a unos pocos especímenes de cormoranes de P-QB C2 y C3, así como de Rheidae en P-QB C1 y de aves voladoras en P-QB C2.

Figura 6
Fragmento diafisario de hueso largo de ave indeterminada con aserrado perimetral.
En P-QB C1, los cormoranes tienen una mayor frecuencia de la porción axial y la cintura escapular, con menor presencia de las extremidades anteriores (un húmero) y posteriores (un tarsometatarso). Los pingüinos están representados por dos especímenes, uno correspondiente a las extremidades anteriores (radio) y otro de las posteriores (tibiatarso). Muy pocos elementos fueron identificados como Rheidae, y estos corresponden al esqueleto axial (dos vértebras, un fragmento de cráneo) y al esqueleto apendicular (al menos dos huesos largos indeterminados). Por último, los restos de aves voladoras presentan un alto porcentaje de fragmentos de diáfisis de huesos largos indeterminados, en igual frecuencia que los elementos de las extremidades posteriores, que son los más frecuentes. El esqueleto axial es el que sigue en abundancia y, por último, la cintura escapular (coracoides) y las extremidades anteriores en igual proporción, mientras que la cintura pélvica está ausente (Tabla 4). Estos perfiles no son acordes con lo registrado en las observaciones actuales. Como la muestra avifaunística de este locus es pequeña y está muy fracturada (Tabla 2), la representación esqueletaria seguramente se encuentra sesgada por estas características, que impiden una determinación más fina de los especímenes, especialmente del esqueleto apendicular.
Representación anatómica (MNE) en los conjuntos óseos de P-QB (Parque Nacional Islote Lobos, Río Negro, Argentina). Referencias: Ph= Phalacrocoracidae; Sp= Spheniscidae; Rh= Rheidae; AV= aves voladoras. *En P-QBC1 y P-QBC2 la categoría aves voladoras incluye los especímenes de E. elegans.
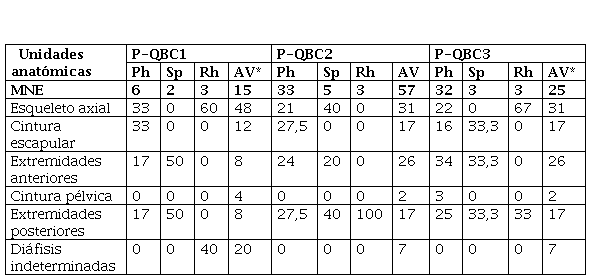
En P-QB C2, entre los huesos de cormoranes predominan la cintura escapular y las extremidades posteriores, seguidas por las extremidades anteriores y el esqueleto axial. Las muestras de pingüinos y Rheidae son muy pequeñas. En el caso de los primeros, su perfil anatómico muestra el predominio de las extremidades posteriores (dos falanges) y el esqueleto axial (esternón y maxilar), con un solo espécimen (una falange) que corresponde a las extremidades anteriores. Los Rheidae, por su parte, están representados por dos tibiatarsos y un fémur (extremidades posteriores). En la muestra de aves voladoras predomina el esqueleto axial por la presencia de varias costillas y vértebras, seguido por las extremidades anteriores, la cintura escapular y las extremidades posteriores en igual proporción. En el conjunto, dado el mayor tamaño de la muestra, se observa una amplia diversidad de elementos. Los perfiles de aves voladoras y cormoranes indican una representación más importante del esqueleto axial y una menor de las extremidades posteriores que en los conjuntos naturales.
Por último, en P-QB C3 los restos de cormoranes muestran el predominio de las extremidades anteriores, seguidas por las extremidades posteriores, el esqueleto axial, la cintura escapular y un único espécimen correspondiente a la cintura pélvica (Tabla 4). En el caso de los huesos de pingüinos hay una frecuencia similar de la cintura escapular, las extremidades anteriores y las posteriores, con un espécimen de cada unidad anatómica. En el caso de los Rheidae, la muestra solo incluye dos elementos del esqueleto axial y uno de las extremidades posteriores. Los restos de aves voladoras muestran el predominio del esqueleto axial (principalmente vértebras), una alta representación relativa de las diáfisis de huesos largos indeterminados, seguida de la cintura escapular y, por último, igual proporción de las extremidades anteriores y las posteriores. Las pequeñas muestras de Spheniscidae y Rheidae presentan perfiles anatómicos que se asemejan bastante a los perfiles naturales, mientras que no ocurre lo mismo con el de cormoranes ni el de aves voladoras.
DISCUSIÓN
Las trayectorias tafonómicas
Los conjuntos avifaunísticos de los tres loci excavados en P-QB presentan características comunes y también diferencias. Estas características permiten conocer al menos parte de la historia de formación de los depósitos y, por lo tanto, evaluar su potencial como fuente de conocimiento sobre las relaciones de los cazadores-recolectores con las aves en la región.
Entre las características comunes se destaca que los tres conjuntos muestran perfiles en los que predominan los huesos no meteorizados o levemente meteorizados. Dado que los huesos de aves en Patagonia continental meteorizan muy rápidamente (Cruz, 2003, 2008, 2009a, 2014, 2015, entre otros), estos perfiles indican que fueron sepultados poco después de la depositación. Esto fue propuesto para P-QB en función de las características de otros taxa identificados en el registro zooarqueológico (Borella et al., 2020; Borella y L’Heureux, 2021). Los resultados referidos a las aves refuerzan esta apreciación previa. Además, es consistente con la presencia de modificaciones por raíces, que indican que los huesos de aves fueron sepultados en entornos de crecimiento de vegetación, en ambientes estables, al menos durante algunos períodos.
Por otra parte, las observaciones actuales indican que los huesos de aves perduran poco en superficie y que su destrucción es rápida debido a procesos como la meteorización, la acción de carnívoros o el pisoteo de animales (Cruz 2003, 2007, 2009a, 2015; Serjeantson, 2009; Massigoge et al., 2015, entre otros). En P-QB, la fragmentación que afecta a gran parte de los huesos de todos los grupos de aves es otra de las características compartidas, propia de la mayoría de los conjuntos avifaunísticos (Serjeantson, 2009). A través de investigaciones actuales se observó que, en líneas generales, la meteorización presenta tasas más altas en los huesos de aves que en los de mamíferos (Behrensmeyer et al., 2003; Cruz, 2003, 2008; Serjeantson, 2009). Sin embargo, también se registró que su impacto no es homogéneo y, aunque se expresa de la misma manera en los huesos de aves voladoras, cormoranes y pingüinos, es diferente con respecto a los Rheidae (Cruz, 2003, 2014, 2015, entre otros). Estas diferencias en las tasas de meteorización se deben al mayor tamaño corporal de los Rheidae y a su particular estructura esqueletaria (Cruz y Elkin, 2003).
Dados los perfiles de meteorización registrados en P-QB, las fracturas no pueden vincularse a este proceso y es muy factible que respondan al pisoteo por el ganado que, en mayor o menor medida, afecta a los depósitos arqueológicos de la zona. Aunque escasas, se registraron trazas que evidencian el pisoteo. Afortunadamente, este proceso no afectó a todos los restos, ya que se recuperaron varios elementos completos que permitieron efectuar determinaciones taxonómicas, incluso a nivel de especie, en todos los conjuntos.
En síntesis, aunque las muestras presentan diferencias cronológicas y se encuentran espacialmente distantes, el análisis de las variables tafonómicas permite plantear que tienen trayectorias tafonómicas similares.
La representación taxonómica y la composición anatómica de las aves
La evaluación tafonómica de los huesos de aves permite comprender si la representación taxonómica registrada en P-QB es fruto de los procesos de formación naturales o culturales. A partir de las propiedades intrínsecas de los huesos de cada grupo de aves y en función de los procesos actuantes en este sector, exploramos la composición de los conjuntos. Como se vio en los resultados, la representación faunística es parecida en los tres loci, ya que en todos ellos aparecen los mismos taxones -con leves variaciones en la composición taxonómica- en cantidades semejantes.
Los cormoranes, aunque son aves voladoras, también nadan y bucean, por lo que su anatomía y estructura ósea responden a los requerimientos de estas formas de locomoción (Bellairs y Jenkin, 1960; Higgins, 1999; Bovy, 2011; entre otros). Causey y Lefèvre (2006), señalaron que algunos elementos son diagnósticos para la determinación taxonómica de los cormoranes patagónicos (coracoides, húmero, fémur, tibiatarso). Estos elementos tienen valores altos de densidad mineral. Lo mismo ocurre con el esqueleto de las aves voladoras, en el que los valores relativos de densidad mineral entre elementos son similares a los de los cormoranes (Broughton et al., 2007; Bovy, 2011). Además, los huesos de cormoranes son más densos que los de otras aves voladoras (Broughton et al., 2007; Bovy, 2011). Si bien no se cuenta con mediciones de densidad mineral para los Spheniscidae, sus huesos son más compactos y densos que los de otras aves, incluso que los de cormoranes, por lo que es esperable que sean menos afectados por los procesos tafonómicos en un mismo contexto de depositación (Cruz, 2009a; Bovy, 2011; Lefèvre y Laroulandie, 2014; entre otros). Esta expectativa, basada en la biomecánica de la locomoción, explica por qué se recuperaron varios elementos completos o casi completos de cormoranes en P-QB.
Por otro lado, algunos de los elementos registrados para cormoranes y aves voladoras -extremidades anteriores y posteriores- corresponden a los huesos más densos. Sin embargo, en los dos casos hay una alta representación del esqueleto axial, para el que no se cuenta con mediciones de densidad mineral ósea, pero que se estima que son menos densos. En tal sentido entendemos que en P-QB el origen de los restos de cormoranes, el de aves voladoras y el de los Rheidae es principalmente producto del descarte antrópico.
En cuanto a los pingüinos, son aves pelágicas no voladoras que poseen una anatomía altamente especializada para nadar y bucear (Bellairs y Jenkin, 1960; García Borboroglu y Boersma, 2015). En P-QB la muestra de estas aves es muy escasa e incluye unos pocos elementos de ambas extremidades y vértebras, que además no presentan modificaciones antrópicas. Por lo tanto, consideramos que muy posiblemente respondan a procesos de mezcla entre huesos actuales y restos arqueológicos, vinculado con la reciente instalación del área de nidificación de pingüinos Patagónicos en el Parque Nacional (Cardillo et al., 2016). Este proceso fue recurrentemente señalado a partir de observaciones tafonómicas actuales efectuadas en otros sectores costeros de Patagonia continental (Cruz, 2003, 2007).
Por último, en el esqueleto de los Rheidae los miembros anteriores y posteriores son marcadamente diferentes, con un importante desarrollo de las extremidades posteriores en función de su adaptación corredora. De tal forma, tibiatarso, tarsometatarso y falanges posteriores presentan paredes gruesas, con un canal central que posee médula, mientras que el fémur es un hueso neumatizado, corto y con un gran diámetro. En cambio, el esqueleto de la cintura escapular y el miembro anterior están muy reducidos (Drenowatz, 1995). Estas diferencias se reflejan en los valores de densidad mineral determinados para la especie (Cruz y Elkin, 2003), que muestran que los elementos de las extremidades posteriores y pelvis presentan valores más altos que los de las extremidades anteriores y el esqueleto axial. En P-QB el perfil anatómico incluye escasos fragmentos tanto del esqueleto axial (vértebras) como de las extremidades posteriores (numerosos fragmentos de diáfisis indeterminadas, mientras que solo un fragmento de fémur, tibiatarso y tarsometarso), que incluyen tantos elementos de alta como de baja densidad mineral. Por otra parte, sólo se registraron fracturas de origen cultural en una de las cuadrículas (Figura 3). Sin embargo, aunque las fracturas observadas en las otras cuadrículas no poseen rasgos diagnósticos que permitan caracterizarlas como antrópicas, es posible plantear que responden a la extracción de médula ósea en función de la representación de partes esqueletarias (ver también Giardina, 2010; Salemme y Frontini, 2011).
Si la depositación de los restos de todas las aves en P-QB respondiera únicamente a causas naturales, sería esperable que los patrones de representación anatómica de los conjuntos tuvieran la forma detectada a partir de observaciones tafonómicas actuales y las estimaciones derivadas de las propiedades intrínsecas de sus huesos. En este sentido, el perfil anatómico característico de las aves voladoras incluye el predominio de elementos correspondientes a las extremidades anteriores y la cintura escapular (Ericson, 1987; Livingston, 1989; Bovy, 2002; Cruz, 2003, 2005), seguidas en abundancia por las extremidades posteriores, el esqueleto axial y, por último, la cintura pélvica. El correspondiente a los Phalacrocoracidae incluye una alta representación de ambas extremidades, generalmente con mayor proporción de las posteriores (Cruz, 2009b; Bovy, 2011; Lefevre y Laroulandie, 2014). La información actual correspondiente a los Spheniscidae muestra que también en este caso predominan ambas extremidades, pero con leve predominio de las anteriores, seguidas de la cintura escapular, el esqueleto axial y la cintura pélvica (Cruz, 2003, 2005, 2007). Por último, los perfiles anatómicos correspondientes a los Rheidae señalan también una mejor representación de las extremidades posteriores, seguidas por el esqueleto axial, la cintura pélvica, la cintura escapular y las extremidades anteriores (Belardi, 1999; Cruz, 2003, 2005; Cruz y Muñoz, 2020). Entonces, en líneas generales, los conjuntos de P-QB no se ajustan a estas expectativas, por lo que no puede afirmarse que los patrones registrados resulten directamente de procesos naturales. Por otro lado, si la representación anatómica registrada fuese similar en los especímenes de un mismo grupo de aves, sería posible detectar la existencia de una explotación antrópica estandarizada, lo cual no ocurre en estos conjuntos. Por ejemplo, en el caso del análisis de Cabo Blanco 1 (Santa Cruz, Argentina), sitio arqueológico costero en la Patagonia Austral (Moreno et al., 1998; Moreno, 2008), se propuso un patrón estandarizado de explotación de los cormoranes patagónicos. A partir de una muestra de 77 individuos, se determinó que en el modo de aprovechamiento de estas aves predominaban las partes anatómicas caracterizadas como “centrales” o de mayor rendimiento económico. Otro ejemplo similar sería el de los restos de las 12 martinetas patagónicas recuperadas en el sitio Paesani 5 en el litoral nordpatagónico (Marani y Borella, 2014), donde también se observó el predominio de partes anatómicas similar al descripto por Moreno y colaboradores (1998). Dado que en P-QB la cantidad de individuos representada es menor que la de los casos mencionados, no descartamos que el tamaño de las muestras esté sesgando el patrón de aprovechamiento de las aves. Sin embargo, como ha planteado Bovy (2002) para el caso de las aves voladoras, también hay que considerar que puede existir una gran diversidad de actividades humanas vinculadas a los restos de aves con diferentes morfoanatomías: consumo de carne y grasa, empleo de huesos, tendones y plumas para la confección de artefactos. Por lo tanto, los patrones de representación anatómica de sus huesos serían diferentes (Cruz, 2003, 2005, 2009b, 2011, 2015, entre otros).
En síntesis, no parece que la representación taxonómica de P-QB estuviera sesgada por procesos tafonómicos que hayan actuado diferencialmente sobre cada grupo de aves. Además, la representación esqueletaria de las aves de P-QB no se adecua a lo observado en conjuntos naturales actuales ni a lo esperado para un patrón de procesamiento común. En tal sentido, el patrón registrado en P-QB respondería tanto a factores naturales como antrópicos.
Acerca del uso antrópico de las aves en P-QB
Otro aspecto a considerar, tanto desde el punto de vista tafonómico como del de la explotación humana de estas presas, es la presencia de trazas antrópicas, que muestra una señal diferente en los tres conjuntos.
Como se mencionó antes, se registraron algunos restos óseos quemados en los tres loci, pero en ningún caso se trató de fragmentos carbonizados o calcinados (Figuras 3 y 5). Esta modificación es ambigua con respecto al agente de depositación, por lo que, en ausencia de otras trazas asignables a humanos en los restos de aves, es difícil sostener su origen antrópico. Aunque es factible que la termoalteración sea producto de la acción antrópica en el pasado -por el descarte de huesos en los fogones o su cocción sobre piedras calientes o sobre las brasas de un fuego abierto (Serjeantson, 2009)-, también existen otras posibilidades como los incendios naturales (Álvarez et al., 2017 y referencias allí mencionadas). En este sentido, -en nuestra región- debido a las escasas precipitaciones, elevadas temperaturas y bajas humedades relativas que imperan, se generan condiciones propicias para los incendios naturales en temporadas estivales (Zacconi y Toppazzini, 2018). Recientes estudios de materiales líticos recuperados en superficie en sectores afectados por fuegos actuales no controlados señalaron la presencia de guijarros termoalterados y pseudoartefactos producto de dichos eventos (Cardillo et al., 2022). En consecuencia, si bien no descartamos que en algunos casos la termoalterción pueda resultar de este tipo de fenómenos naturales acontecidos azarosamente en el pasado, es posible que otras alteraciones térmicas respondan a manipulación intencional o exposición de ciertas partes de las carcasas al fuego. Aun así, la evidencia de Rheidae en P-QB es escasa, por lo que podría señalar el aprovechamiento ocasional de uno o unos pocos individuos. El registro arqueológico de estas aves en Patagonia indica que previamente a la introducción del caballo (a partir del siglo XVIII) su captura era muy limitada y ocasional, dadas las dificultades que involucraba su caza por las altas velocidades de carrera de los ñandúes (Fernández, 2000; Cruz y Elkin, 2003; Salemme y Frontini, 2011; entre otros).
En cambio, en P-QB C2 y C3 además de varios especímenes con alteración térmica, se registraron huellas de corte y otras modificaciones antrópicas que, aunque no son abundantes, indican que al menos parte de los conjuntos fue depositada y modificada por los cazadores-recolectores. La observación de huellas de corte en húmeros en ambos loci es consistente con la información proveniente de registros avifaunísticos de sitios del Paleolítico Superior en Francia. En esos casos se señaló que independientemente de las especies, es en este elemento donde se registran la mayor frecuencia de marcas de corte, epifenómeno que puede ser indicador tanto de desarticulación como de fileteado (Serjeantson, 2009). Por otra parte, esta autora señaló que la información etnográfica de distintos lugares del mundo ha mostrado que la manipulación de las aves no necesariamente deja trazas en sus huesos (Serjeantson, 2009 y citas allí incluidas), por lo que la ausencia de modificaciones de este origen no es del todo concluyente.
También se registraron otras evidencias de manipulación antrópica, como son las marcas de aserrado perimetral en las diáfisis de huesos largos de aves voladores recuperados en la cuadrícula 2 y 3 (Figura 6). Estas modificaciones pueden ser interpretadas como desechos de manufactura de cuentas óseas o la obtención de cilindros (formas base) para confeccionar artefactos varios (pendientes, adornos corporales, u otros). Si bien la evidencia actual de P-QB es escasa, se suma a otros hallazgos realizados en otras localidades arqueológicas de la costa del GSM, como en Paesani y en Bajo de la Quinta donde también se recuperaron diáfisis de huesos largos de ave con dicha modificación antrópica (Marani, 2018).
La abundancia taxonómica (MNI) es otra característica que, juntamente con las modificaciones antrópicas, permite abordar el tema de la explotación humana de manera más robusta. Las observaciones tafonómicas fuera de áreas de nidificación de diferentes aves patagónicas mostraron que, en general, la depositación natural de restos de aves se caracteriza por corresponder a uno o unos pocos individuos aislados (Cruz, 2003, 2007, 2009a, 2011, 2015, entre otros). En los conjuntos de P-QB la abundancia taxonómica es baja, ya que corresponde a uno o dos individuos para cada grupo o taxón. Por otro lado, la presencia de modificaciones antrópicas -que no es homogénea entre cuadrículas y entre grupos de aves- es suficiente para garantizar que hubo alguna forma de aprovechamiento humano.
Dado que los cormoranes son las aves que están relativamente mejor representadas y que además presentan evidencia de manipulación antrópica (Figuras 4 y 5), es posible considerar los distintos escenarios propuestos para su captura en el pasado: a) la captura de individuos provenientes de las colonias durante el período reproductivo, b) la obtención de aves en sus áreas de alimentación o descanso y c) la colecta de carcasas de animales muertos naturalmente (Cruz et al., 2021).
En el Parque Nacional se registran actualmente áreas de nidificación de biguás (Phalacrocorax brasilianus) y es común el avistaje de L. atriceps, que no nidifica en el área (Reyes y García Borboroglu, 2020). Sin embargo, a mediados del siglo XX se explotaba el guano de un área de nidificación de cormoranes imperiales que habría desaparecido en ese momento (Godoy, 1963; Punta, 1996). Si en el pasado también existían colonias reproductivas en áreas próximas, los huesos en los conjuntos de P-QB podrían evidenciar su explotación, ya que habrían sido recursos predecibles espacial y temporalmente. Un indicador importante para discutir la explotación de áreas de nidificación es la presencia de pichones o juveniles de las aves marinas (Bovy, 2011; Cruz et al., 2021; y referencias allí citadas). La mayor parte de los huesos de cormoranes de los tres loci de P-QB corresponden a individuos adultos. Dado este perfil etario, la evidencia podría indicar cualquiera de los tres escenarios propuestos, en los que la captura pudo haber implicado tanto el aprovechamiento de carne, como de sus huesos, tendones y plumas para la confección de artefactos como lo atestiguan las huellas de corte registradas.
CONCLUSIONES
La información aportada por el análisis de los huesos de aves de P-QB sustenta las interpretaciones sobre el estado general de preservación de los depósitos en este sector de la costa rionegrina, efectuadas previamente sobre la base de otras líneas de evidencia. A la vez, alerta sobre las particularidades que presenta el registro avifaunístico, ya que, aunque las muestras son pequeñas, se pudo efectuar la evaluación tafonómica de los restos de aves y generar inferencias sobre los procesos que actuaron en la formación de los conjuntos. Consideramos que era necesario y pertinente indagar acerca del origen de los conjuntos avifaunísticos dado que, en un área donde actualmente anidan varias especies, no podía descartarse a priori que pudiera existir “contaminación” de los depósitos zooarqueológicos con restos de aves muertas naturalmente. Por otro lado, a partir de esta valoración tafonómica fue posible comenzar a explorar las tendencias en el uso de las aves como presas de los cazadores-recolectores en este sector costero durante el Holoceno tardío final. Esta información no solo permitirá profundizar el conocimiento sobre las poblaciones humanas que habitaron la zona, sino que también permitirá conocer parte de la historia natural de la fauna, lo cual es de sumo interés, dada la reciente creación del Parque Nacional Islote Lobos.
Por otra parte, la identificación de huellas de cortes y otras modificaciones antrópicas es consistente con otros registros de procesamiento humano de las aves, y permite comenzar a explorar las propiedades culturales emergentes en conjuntos avifaunísticos de la región, aun cuando las muestras son pequeñas.
En el futuro, los estudios en escala local como el presente, que contemplen ambas perspectivas (discusión tafonómica y zooarqueológica), podrán entregar resultados sólidamente generados que serán la base para las discusiones en escalas regionales. Esto es fundamental para avanzar en un estudio más amplio sobre el papel de las aves en la subsistencia de las sociedades del pasado, al comparar distintos momentos y formas de uso en diferentes espacios costeros en Patagonia.
Agradecimientos
A la Secretaría de Cultura de la Provincia de Río Negro y a las autoridades de PN por otorgar los pemisos de trabajo en el área. Los subsidios de investigación fueron otorgados por CONICET y ANPCyT. A los miembros del equipo del Golfo San Matías que colaboraron en la excavación durante los diferentes trabajos de campo. A la Dra. Jimena Alberti por la traducción del resumen y a la Dra. Eugenia Carranza por la diagramación de las figuras. A los evaluadores anónimos cuyas sugerencias permitieron mejorar la comprensión del manuscrito. A la Dra. Mariela González y a la Dra. Ana Paula Alcaraz del equipo editorial de la revista por su valiosa labor.
REFERENCIAS
Acosta, A. (2000). Huellas de corte relacionadas con la manufactura de artefactos óseos en el nordeste de la provincia de Buenos Aires. Relaciones de la Sociedad Argentina de Antropología, 25, 159-178.
Álvarez, M. C., Massigoge, A., Scheifler, N., Gonzalez, M. E., Kaufmann, C. A., Gutiérrez, M. A. y Rafuse, D. J. (2017). Taphonomic Effects of a Grassland Fire on a Modern Faunal Sample and its Implications for the Archaeological Record. Journal of Taphonomy, 15(1-3), 77-90.
Behrensmeyer, A.K., Stayton, C.T. y Chapman, R.E. (2003). Taphonomy and ecology of modern avifaunal remains from Amboseli Park, Kenya. Paleobiology, 29, 52-70. http://doi:10.1666/0094-8373(2003)0292.0.CO
Belardi, J.B. (1999). Hay choiques en la terraza. Información tafonómica y primeras implicaciones arqueofaunísticas para Patagonia. Arqueología. Revista de la Sección Prehistoria, 9, 163-185.
Bellairs, A. D. y Jenkin, C. R. (1960). The Skeleton of Birds. En A. J. Marshall (Ed.), Biology and Comparative Physiology of Birds 1 (pp. 241-300). J. Wiley & Sons.
Beretta, J. M. y Zubimendi, M. (2019). Explotación de avifauna en el sitio Cueva del Negro (costa norte de Santa Cruz). Mundo de Antes, 13(1), 85-112. http://www.mundodeantes.org.ar//pdf/revista13-1/04-beretta.pdf
Bertellotti, M., Yorio, P. M. y García Borboroglu, P. (2015). Las aves como recurso en la zona costera patagónica. En H. E. Zaixso y A. L. Boraso (Eds.), La Zona Costera Patagónica Argentina. (Vol. III, pp. 57-76). Editorial Universitaria de la Patagonia.
Binford, L. R. (1981). Bones. Ancient Men and Modern Myths. Academic Press.
Borella, F. y Cruz, I. (2012). Taphonomic evaluation of penguin (Spheniscidae) remains at a shell-midden on the northern coast of Patagonia (San Matías Gulf, Río Negro, Argentina). Quaternary International, 278, 45-50. http://doi.org/10.1016/j.quaint.2012.04.023
Borella, F. y L´Heureux, G. L. (2021). Exploring the zooarchaeological evidence of otariids exploitation by terrestrial hunter-gatherers along the western coast of San Matías Gulf, Argentina. En E. Piskin (Ed.), 13th International Council of Archaeozoology Conference: Archaeological, biological and historical approaches in archaeozoological research (pp. 75-83). BAR Series 3028.
Borella, F., Cardillo, M., Alberti, J., Scartascini, F. L., Carranza, E., Favier Dubois, C. M., Steffan, P. G. y Guichón-Fernández, R. (2020). Resultados preliminares de las investigaciones arqueológicas en el Área Natural Complejo Islote Lobos, costa oeste del golfo San Matías (provincia de Río Negro). Revista del Museo de Antropología, 13(2), 69-78. https://doi.org/10.31048/1852.4826.v13.n2.25382
Borella, F., Scartascini, F. L. y Marani, H. A. (2011). Explorando la subsistencia humana a partir de la evidencia faunística en la costa rionegrina. En F. Borella y M. Cardillo (Eds.), Arqueología de pescadores y marisqueadores en Nordpatagonia. Descifrando un registro de más de 6000 años (pp. 87-110). Dunken.
Bovy, K. M. (2002). Differential avian skeletal part distribution: Explaining the abundance of wings. Journal of Archaeological Science, 29, 965-978. doi.org/10.1006/jasc.2001.0795
Bovy, K. M. (2011). Archaeological Evidence for a Double-Crested Cormorant (Phalacrocorax auritus) Colony in the Pacific Northwest, USA. Waterbirds, 34(1), 89-95. https://doi.org/10.1675/063.034.0111
Broughton, J. M., Mullins, D. y Ekker, T. (2007). Avian resource depression or intertaxonomic variation in bone density? A test with San Francisco Bay avifaunas. Journal of Archaeological Science, 34, 374-391. http://doi.org/10.1016/j.jas.2006.05.013
Cardillo, M., Carranza, E., Alberti, J. y Borella, F. (2022). Alteraciones térmicas en guijarros costeros en la localidad de Las Grutas (Río Negro). Discutiendo sus implicancias para la interpretación del registro arqueológico lítico. Revista del Museo de Antropología, 15(3), 273-288. http://doi.org/10.31048/1852.4826.v15.n3.38007
Cardillo, M., Carranza, M. E. y Borella, F. (2016). Tafonomía y tecnología lítica en un ambiente altamente dinámico. El caso de la pingüinera del islote La Pastosa (Provincia de Río Negro), Patagonia, Argentina. Journal of Lithic Studies, 2(2), 49-66. http://doi.org/10.2218/jls.v2i2.1305.
Causey, D. y Lefèvre, C. (2006). Diagnostic osteology and analysis of the Mid- to Late Holocene dynamics of shags and cormorants in Tierra del Fuego. International Journal of Osteoarchaeology, 17(2), 119-137. http:// doi.org/10.1002/oa.865
Cruz, I. (2003). Paisajes tafonómicos de restos de Aves en el sur de Patagonia continental. Aportes para la interpretación de conjuntos avifaunísticos en registros arqueológicos del Holoceno [Tesis doctoral, Facultad de Filosofía y Letras, Universidad de Buenos Aires]. http://repositorio.filo.uba.ar/handle/filodigital/1313
Cruz, I. (2005). La representación de partes esqueléticas de aves. Patrones naturales e interpretación arqueológica. Archaeofauna, 14, 69-81.
Cruz, I. (2006). Los huesos de pingüinos (Spheniscidae) de los sitios de Cabo Blanco (Santa Cruz, Patagonia Argentina). Análisis tafonómico y perspectivas arqueológicas. Intersecciones en Antropología, 7, 15-26.
Cruz, I. (2007). Avian Taphonomy: observations at two Magellanic penguin (Spheniscus magellanicus) breeding colonies and their implications for the fossil record. Journal of Archaeological Science, 34, 1252-1261. http://doi.org/10.1016/j.jas.2006.10.016
Cruz, I. (2008). Avian and mammalian bone taphonomy in Southern Continental Patagonia. A comparative approach. Quaternary International, 180, 30-37. http://doi.org/10.1016/j.quaint.2007.08.008
Cruz, I. (2009a). Tafonomía en escalas espaciales amplias: el registro óseo de las aves en el sur de Patagonia. En A. Acosta, D. Loponte y L. Mucciolo (Eds.), Temas de Arqueología: Estudios tafonómicos y zooarqueológicos (I) (pp.15-34). Instituto Nacional de Antropología y Pensamiento Latinoamericano.
Cruz, I. (2009b). Tafonomía de huesos de cormoranes en la costa patagónica. Primeros resultados. En M. Salemme, F. Santiago, M. Alvarez, E. Piana, M. Vázquez y E. Mansur (Eds.), Arqueología de la Patagonia. Una mirada desde el último confín (pp. 729-741). Utopía.
Cruz, I. (2011). Tafonomía de huesos de aves. Estado de la cuestión y perspectivas desde el sur del Neotrópico. Antípoda, Revista de Antropología y Arqueología, 13, 147-174.
Cruz, I. (2014). Estudios sobre meteorización de huesos en Patagonia. Revista Chilena de Antropología, 29, 89-94.
Cruz, I. (2015). Las investigaciones sobre preservación de huesos de aves y mamíferos en Patagonia (Argentina). Archaeofauna, 24, 189-204. http://doi.org/10.15366/archaeofauna2015.24.012
Cruz, I. y D. Elkin (2003). Structural Bone Density of the Lesser Rhea (Pterocnemia pennata) (Aves: Rheidae). Taphonomic and Archaeological Implications. Journal of Archaeological Science 30, 37-44. http:// doi.org/10.1006/jasc.2001.0826
Cruz, I. y Muñoz, A.S. (2020). Between Space and Time. Naturalist Taphonomic observations of Lesser Rhea (Rhea pennata pennata) bone remains in Southern Patagonia and its archaeological implications. Journal of Archaeological Science Reports, 31, 102290. http://doi.org/10.1016/j.jasrep.2020.102290
Cruz, I., Cañete Mastrángelo, D. y Ercolano, B. (2021). Interacciones entre cormoranes (Phalacrocorax spp.) y humanos en Punta Entrada y Monte León (Santa Cruz, Argentina) durante el Holoceno. Magallania, 49(1), Epub 17 de octubre de 2021 http://dx.doi.org/10.22352/magallanla202149001
David, B. (1990). How was this Bone Burnt? En S. Solomon, Davidson, I. y D. Watson (Eds.), Problem Solving in Taphonomy: Archaeological and Paleontological Studies from Europe, Africa and Oceania (pp. 65-79). Universidad de Queensland.
Drenowatz, C. (Ed.) (1995). The Ratite Encyclopedia. Ostrich, Emu, Rhea. Ratite Records Incorporated.
Ericson, P. G. (1987). Interpretations of Archaeological Bird Remains: a Taphonomic Approach. Journal of Archaeological Science, 14, 65-75. http:// doi.org/10.1016/S0305-4403(87)80006-7
Favier Dubois C. M. y Borella, F. (2011). Contrastes en la costa del golfo: una aproximación al estudio del uso humano del litoral rionegrino en el pasado. En F. Borella y M. Cardillo (Eds.), Arqueología de pescadores y marisqueadores en Nordpatagonia. Descifrando un registro de más de 6000 años (pp. 13-42). Dunken.
Fernández, P.M. (2000). Rendido a tus pies: acerca de la composición anatómica de los conjuntos arqueofaunísticos con restos de Rheiformes de Pampa y Patagonia. Desde el país de los gigantes. Perspectivas arqueológicas en Patagonia (Tomo II, pp. 573-586). Universidad Nacional de la Patagonia Austral.
Foro para la Conservación del Mar Patagónico y Áreas de Influencia (2008). Síntesis del estado de conservación del Mar Patagónico y áreas de influencia. Edición del Foro.
France, D. L. (2017). Comparative Bone Identification. Human subadult to Nonhuman. CRC Press Taylor & Francis Group.
Frere, E., Quintana, F. y Gandini, P. (2005). Cormoranes de la costa Patagónica: estado poblacional, ecología y conservación. El Hornero, 20, 35-52. http://www.scielo.org.ar/pdf/hornero/v20n1/v20n1a04.
García Borboroglu, P. y Boersma, P. D. (2015). Pingüinos. Historia natural y conservación. Vázquez Mazzini Editores.
Gelós, E. M., Spagnuolo, J. O. y Schillizzi, R. A. (1992). Las unidades morfológicas de la costa oeste de Golfo San Matías y su evolución. Revista de la Asociación Geológica, XLIII (3), 315-326.
Giardina, M. A. (2006). Anatomía económica de Rheidae. Intersecciones en antropología, 7, 263-276.
Giardina, M. (2010). El aprovechamiento de la avifauna entre las sociedades cazadoras-recolectoras del sur de Mendoza: un enfoque arqueozoológico [Tesis doctoral, Facultad de Ciencias Naturales y Museo, Universidad Nacional de la Plata].
Gilbert, B. M., Martin, L. D., y Savage, H. G. (1996). Avian Osteology. Missouri Archaeological Society, Inc.
Gifford-González, D. (2018). An Introduction to Zooarchaeology. Springer International Publishing.
Godoy, J. C. (1963). Fauna silvestre. Tomo VIII (Vol. 1). Consejo Federal de Inversiones.
González Díaz, E. y Malagnino, E. (1984). Geomorfología de la provincia de Río Negro. Actas del IX Congreso Geológico Argentino (p. 159). Secretaría de Minería, Buenos Aires, Argentina.
Higgins, J. (1999). Túnel: A Case Study of Avian Zooarchaeology and Taphonomy. Journal of Archaeological Science, 26, 1449-1447. https://doi.org/10.1006/jasc.1999.0429
Kokot, R. y Pisani, F. (2011). Ingresión marina holocena entre Isla Lobos y Punta Pórfido, Provincia de Río Negro. Actas de XVIII Congreso Geológico Argentino (CD room) Neuquén.
Ksepka, D. T., Werning, S., Sclafani, M. y Boles, Z. M. (2015). Bone histology in extant and fossil penguins (Aves: Sphenisciformes). Journal of Anatomy, 227, 611-630. https://doi.org/10.1111/joa.12367
Lefèvre, C. y Laroulandie, V. (2014). Avian Skeletal Part Representation: A Case Study from Offing 2, A Hunter-Gatherer-Fisher Site in the Strait of Magellan (Chile). International Journal of Osteoarchaeology, 24, 256-264. https://doi.org/10.1002/oa.2361
León, D. C. (2010). Identificación taxonómica y anatómica de muestras arqueofaunísticas a partir de fragmentos diafisiarios de huesos largos. En M. A. Gutiérrez, M. De Nigris, P. M. Fernández, M. Giardina, A. Gil, A. Izeta, A. G. Neme y G. H. Yacobaccio (Eds.), Zooarqueología a principios del siglo XXI: Aportes Teóricos, Metodológicos y Casos de Estudio (pp. 145-155). Libros del Espinillo.
Livingston, S. D. (1989). The Taphonomic Interpretation of Avian Skeletal Part Frecuencies. Journal of Archaeological Science, 16, 537-547. https://doi.org/10.1016/0305-4403(89)90072-1
Lyman, R. L. (1994). Vertebrate Taphonomy. Cambridge University Press.
Moreno, E. (2008). Arqueología y etnohistoria de la Costa Patagónica Central en el Holoceno Tardío. Fondo Editorial de la Provincia del Chubut.
Moreno, E., Castro, A., Martinelli, K. y Abello, A. (1998). El material faunístico de Cabo Blanco 1. Anales del Instituto de la Patagonia (Serie Ciencias Sociales), 26, 169-180.
Marani, H. A. (2018). Aprovechamiento de vertebrados terrestres por las poblaciones humanas que habitaron la costa del Golfo San Matías (Río Negro, Argentina) durante el Holoceno tardío. Archaeopress Publishing LTD.
Marani, H. A. y Borella, F. (2014). Patrón de explotación de Tinámidos en la costa atlántica Norpatagónica (Argentina) durante el Holoceno Tardío. Revista Chilena de Antropología, 29(1), 68-73.
Marean, C. W. (1995). Of Taphonomy and Zooarchaeology. Evolutionary Anthropology, 4, 64-72.
Massigoge, A., Rafuse, D. J., Álvarez, M. C., González, M. E., Gutiérrez, M. A., Kaufmann, C. A. y Scheifler, N. A. (2015). Beached penguins on the Atlantic Coast in the Pampas region of Argentina: Taphonomic analysis and implications for the archaeological record. Palaeogeography, Palaeoclimatology, Palaeoecology, 436, 85-95. https://doi.org/10.1016/j.palaeo.2015.06.045
Punta, G. (1996). Estado de situación del recurso guanero en la República Argentina. Informes Técnicos del Plan de Manejo Integrado de la Zona Costera Patagónica, Fundación Patagonia Natural, 6, pp.1-19. https://patagonianatural.org.ar/wp-content/uploads/2021/04/248_IT06.pdf
Reyes, L. M. y García Borboroglu, P. (2020). Plan de Manejo del Área Natural Protegida Complejo Islote Lobos. Provincia de Río Negro y Consejo Federal de Inversiones. 176 págs. Boletín Oficial Provincia de Río Negro, Suplemento 5921. http://biblioteca.cfi.org.ar/wp-content/uploads/sites/2/2019/10/plan-de-manejo-complejo-islote-lobos-documento-final.pdf
Salemme, M. y Frontini, R. (2011). The exploitation of Rheidae in Pampa and Patagonia (Argentina) as recorded by chroniclers, naturalists and voyagers. Journal of Anthropological Archaeology, 30, 473-483. https://doi.org/10.1016/j.jaa.2011.08.001
Scartascini, F. L. (2020). Arqueología de la pesca en la costa rionegrina: Patagonia Argentina. IIDyPCa-Instituto de Investigaciones en Diversidad Cultural y Procesos de Cambio.
Schiavini, A., Yorio, P., Gandini, P., Raya Rey, A. y Boersma, P. D. (2005). Los pingüinos en las costas argentinas: estado poblacional y conservación. El Hornero, 20(1), 5-23 http://www.scielo.org.ar/scielo.php?script=sci_arttext&pid=S0073-34072005000100002
Serjeantson, D. (2009). Birds. Cambridge University Press.
Vinci, M. C. (2004). Los humedales de la costa patagónica de la Provincia de Río Negro. En Malvárez, A. I. y Bó, R. F. (Comps.), Documentos del curso-taller “Bases ecológicas para la clasificación e inventario de humedales en Argentina”. (Buenos Aires, 30 de septiembre - 4 de octubre 2002) FCEYN-UBA; RAMSAR; USFWS; USDS.
Yorio, P. M., Caille, G. M., Schwindt, E., Tagliorette, A., Esteves, J. L., Crespo, E. A., Arias, A. M. y Harris, G. (2015). Conservación de la Diversidad Biológica en la Zona Costera de la Patagonia Argentina. En H. E. Zaixso y A. L. Boraso (Eds.), La Zona Costera Patagónica Argentina. (Vol. III, pp. 159-217). Editorial Universitaria de la Patagonia.
Zacconi, G. y Toppazzini, M. (2018). Áreas afectadas por incendios forestales y rurales en la región pampeana y noreste de la región patagónica durante la temporada 2016-2017. Informe Técnico N° 13. Servicio Nacional de Manejo del Fuego, Ministerio de Ambiente y Desarrollo Sustentable de la Nación.
Zubimendi, M. A. y Beretta, M. (2018). La presencia de aves en el registro arqueológico de la Costa Norte de Santa Cruz. Comechingonia, 22(2),167-198. https://doi.org/10.37603/2250.7728.v22.n2.25593
