Morfofisiologia Vegetal
Received: 26 February 2016
Accepted: 08 July 2016
Abstract: Different species of plants can act as environmental filters affecting the composition, structure and spatial distribution of seeds and seedlings of forest species. This study aimed to examine whether the presence of the fern stratum acts as a biotic environmental filter on the woody species seedling bank in a semi-deciduous forest remnant (Atlantic biome), Southern Brazil. Two experiments were developed in areas with ferns (F) and ferns-free (NF): a) richness and abundance of woody seedling bank were determined in 10 plots (25 m2) in each area; b) seedlings abundance of Actinostemon concolor (Spreng.) Müll. Arg., Aspidosperma polyneuron Müll. Arg., Euterpe edulis Mart., Guarea kunthiana A. Juss. and Inga marginata Kunth were monitored for a year in three areas F, NF and an area with ferns whose fronds were cut (CF). In this experiment, it was used five plots (4 m2) in each area. The abundance of seedlings (considering all species) was higher in F areas and ferns positively and negatively affected the abundance of A. polyneuron and G. kunthiana, respectively. It is possible to conclude that ferns worked as a selective environmental filter for some woody species.
Keywords: Atlantic forest, environmental filter, mortality, recruitment, regeneration.
Resumo: Diferentes espécies de plantas alteram a composição, a estrutura e a distribuição espacial de sementes e plântulas de espécies florestais, pois atuam como filtros ambientais. Objetivou-se testar se manchas de samambaias funcionam como filtro biótico sobre o banco de plântulas de espécies lenhosas de floresta semidecidual do bioma Atlântico no sul do Brasil. Dois experimentos foram desenvolvidos em áreas com (F) ou sem samambaias (NF): a) a riqueza e a abundância de plântulas de espécies lenhosas foram avaliadas em dez parcelas de 25 m2, em cada área; b) a abundância de plântulas de Actinostemon concolor (Spreng.) Müll. Arg., Aspidosperma polyneuron Müll. Arg., Euterpe edulis Mart., Guarea kunthiana A. Juss. e Inga marginata Kunth foi monitorada, por um ano, em áreas F, NF e em uma terceira área com samambaias, porém com as frondes cortadas (CF). Neste experimento foram utilizadas cinco parcelas de 4 m2, em cada área. A abundância de plântulas, somando todas as espécies, foi maior nas áreas F e as samambaias afetaram positivamente a abundância de A. polyneuron e, negativamente a de G. kunthiana. Concluiu-se que as samambaias funcionam como filtro ambiental seletivo para algumas espécies lenhosas.
Palavras-chave: Filtro ambiental, floresta Atlântica, mortalidade, recrutamento, regeneração.
Introduction
Tropical forest regeneration is influenced by a complex interaction of biotic and abiotic forces, such as the actions of dispersers, seed predators, pathogens, herbivores, light, water, nutrients, soil and landscape structure (Hobbs & Norton, 2004). Both abiotic and biotic forces function as environmental filters, which favor or exclude species in a plant community and define its structure (Lortie et al., 2004). The concept of environmental filters can help to understand the forces that maintain biological diversity, both taxonomic and functional,
as well as explain abundance and distribution patterns of species in landscapes, which are critical problems in ecological studies (Elith & Leathwick, 2009). They can also explain the regeneration of tree species that live under the canopy, making it possible to relate the maintenance of biodiversity of tree species to the composition and structure of tropical and subtropical forest canopies (Gandolfi, Joly, & Rodrigues, 2007).
Forest species of different life forms, such as ferns, can act as environmental filters affecting the composition, structure and spatial distribution of seedlings or seeds from canopy and understory species. Rosettes of Nidularium procerum Lindm. (Bromeliaceae) formed a safe substrate for germination and/or establishment of some tree species in a Brazilian freshwater swamp forest, especially those with vegetative propagation (Scarano, Ribeiro, Moraes, & Lima, 1997). Abundance of palm trees and cyclanths negatively affected the density of seedlings of Inga fagifolia (L.) Benth in a tropical rainforest in Costa Rica (Denslow, Newell, & Ellison, 1991). Species diversity and sapling height in natural regeneration vary under the three studied canopy tree species in a restored Brazilian semi-deciduous forest (Vieira & Gandolfi, 2006).
Many studies indicate that ferns can block the regeneration of certain species, and thus, ‘filter’ which species are capable of regenerating (e.g. George & Bazzaz, 1999; Walker, Landau, Velázquez, Shiels, & Sparrow, 2010; Gallegos, Hensen, Saavedra, & Schleuning, 2015). Fern understory decreased emergence of Betula spp., Pinus strobus L., and Quercus rubra L. but did not affect the emergence of Acer rubrum L. or Fraxinus americana L. in a deciduous forest (George & Bazzaz, 1999). Ferns also inhibited the establishment of late succession woody plants and regeneration of woody plants increased after the removal of the fern clumps (Walker et al., 2010).
Several studies on the impact of ferns on plant communities have been carried out in the world (George & Bazzaz, 1999; Walker et al., 2010; Gallegos et al., 2015), but almost nothing was published about this subject in Brazil (Silva & Silva Matos, 2006; Silva Matos & Belinato, 2010; Guerin & Durigan, 2015), specially in seasonal semi-deciduous forest (SSF). Knowledge about the influence of herbaceous stratum on tree regeneration is essential for conservation and reforestation programmes of these forests.
The present study reports the results of experiments designed to test the hypothesis in which fern stratum acts as an ecological filter on the woody species seedlings bank in a Brazilian SSF remnant. Ferns are considered an environmental filter when they promote the exclusion of the species or affect positively or negatively their abundance.
Material and methods
Study area
The climate is classified as Köppen’s Cfa type with an average rainfall of 1,600 mm distributed unevenly throughout the year. The rainfall from December to January (summer) is three times higher than from June to August (winter). The average annual temperature is around 21oC, and January (23.9°C) and June (16.3°C) are the hottest and the coldest month, respectively. The climate data were obtained from the Instituto Agronômico do Paraná.
Mata dos Godoy State Park (MGSP), located in Londrina Municipality (23°26’33” S, 51°14’34” W), is one of the largest (about 680 ha) and best preserved forest (Silveira, 2006) in the North of Paraná State, southern Brazil. The study was developed on flat area with good conservation condition and did not present gaps (Silveira, 2006) in the northern portion of the remnant. In this portion, the tree species with the highest values of importance are Aspidosperma polyneuron Müll. Arg., Euterpe edulis Mart., Croton floribundus Spreng., Trichilia claussenii C. DC. and Nectandra megapotamica (Spreng.) Mez (L. H. Soares-Silva, personal communication). The canopy ranges from 16 to 20 m in height and has high canopy cover (Bianchini, Pimenta, & Santos, 2001). Several species of ferns occur in this area, especially Ctenitis cf. falciculata (Raddi) Ching, Didymochlaena truncatula (Sw.) J. Sm Diplazium cristatum (Desr.) Alston and Lastreopsis effusa (Sw.) Tindale. Patches of ferns are observed in the understory separated by large ferns-free areas. Further studies should be conducted to assess the biotic and abiotic factors involved in the cluster distribution of ferns, as well as clonal reproduction. In the patches, it was observed 0.5 ± 0.04 (mean ± standard errors) individuals per m² and fronds averaged 87.8 ± 3.1 cm in height.
Data collection
In the first experiment, it was quantified the natural regeneration of woody species in two plots divided into 10 contiguous sub-plots of 5 m x 5 m, totaling 250 m2 in each plot. The sub-plots were contiguous due to difficulty of finding multiple locations with the same characteristics (no gaps, soil, slope, distance of edges, etc). The first plot was designated in an area with ferns (F) and the other in a ferns-free area and any other dominating herbaceous species (NF).
In this study, it was considered as seedlings all woody individuals up to 50 cm in height. The seedlings were identified as species or morpho-species (hereafter species) due to the difficulty of differentiation between some species in this stage of development (Scarano et al., 1997). Species identification was performed by comparison with the seedlings produced by the Universidade Estadual de Londrina nursery.
To characterize the environmental features, it was quantified the index of canopy openness (ICO), dry mass of litter, moisture and pH of the soil. Four ICO measurements were taken at ground level using a spherical densiometer in each sub-plot. The litter samples were collected in wooden squares of 0.5 x 0.5 m in the center of each sub-plot. Then, the litter was stored in paper bags, ovendried at 80oC and weighed on a semi-analytical scale. Moisture (Fieldscout TDR 100 soil moisture portable meter) and pH (E. W. System Soil Tester) of the soil were measured in the center of each sub-plot.
In the second experiment, the effects of ferns on seedlings of five shade-tolerant tree species were investigated. The species were Actinostemon concolor (Spreng.) Müll. Arg. (Euphorbiaceae), Aspidosperma polyneuron (Apocynaceae), Euterpe edulis (Arecaeae), Guarea kunthiana A. Juss (Meliaceae) and Inga marginata Kunth (Fabaceae). These shade-tolerant tree species are among the most abundant tree species in the MGSP (field observations). Five contiguous sub-plots of 4 m² were allocated in different areas of the first experiment for the following treatments: sub-plots with ferns (F), sub-plots with ferns whose fronds were cut with the aid of pruning shears at the beginning of the study and monthly throughout the experiment (CF) and sub-plots naturally without ferns (NF). In these sub-plots, the seedlings of the species were counted and marked with aluminum plates. Subsequently, these sub-plots were monitored every three months for a year, and new individuals or deaths of these five species were recorded.
Data analysis
The rarefaction curves were computed in order to compare the species richness between F and NF. These rarefaction curves were compared with two estimates of total species richness: Chao2 and Jackknife2. Rarefaction curves were generated by EstimateS 9.1.0 software (Colwell, 2013), after 100 randomizations on the sampling order with 95% confidence intervals. The Mann-Whitney test (a£0.05) was used to compare the total seedling abundance between F and NF. Environmental characteristics of F and NF were compared by Kruskal Wallis test, with significance level of 0.05. The Friedman's variance analysis was used to compare the abundance, recruitment and mortality of the five species among treatments.
Results
First experiment
It was sampled a total of 4629 seedlings of 62 species. The abundance of seedlings in F plot (2749) was higher than in NF plot (1880), with a difference of 869 individuals (p < 0.05). In F plot, it was recorded 274.9 ± 94.6 (mean ± standard errors) individuals per sub-plot, with 11 individuals per m2 belonging to 54 species and 188 ± 57.8 individuals per sub-plot, with 7.5 individuals per m2 belonging to 50 species in NF plot (Table 1).
Number of individuals per species or morpho-species in with (F) and without ferns (NF) plots in the Mata dos Godoy State Park, Londrina, PR, Brazil. Species were numbered in decreasing order of abundance in the F plot.

Number of individuals per species or morpho-species in with (F) and without ferns (NF) plots in the Mata dos Godoy State Park, Londrina, PR, Brazil. Species were numbered in decreasing order of abundance in the F plot.
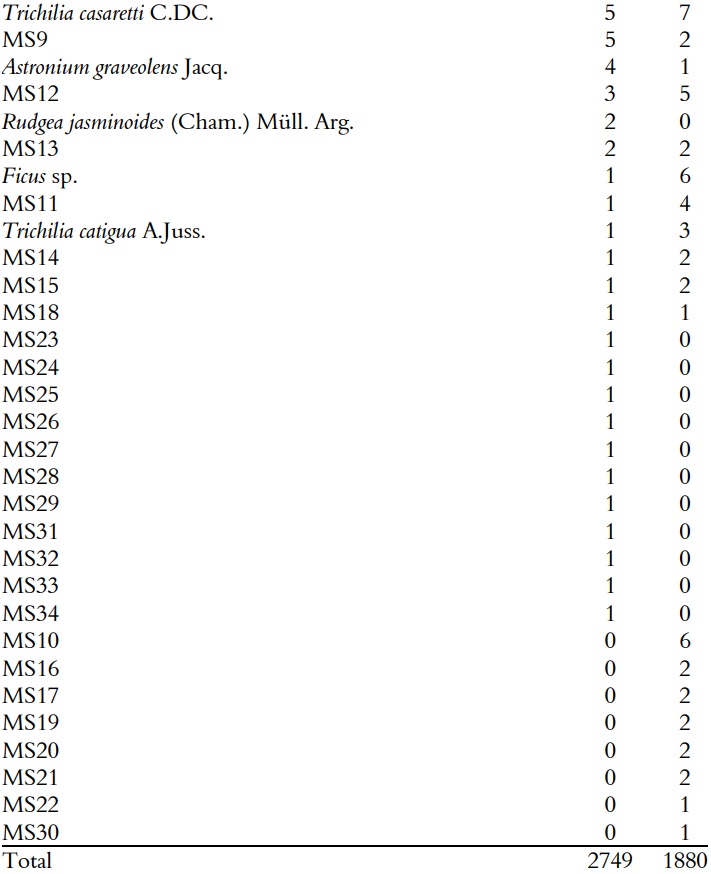
From 62 species sampled in the study, 42 occurred in both plots and 12 and 8 were exclusive to the F and the NF respectively (Table 1). However, the exclusive species had low abundance.
There was no difference in species richness between F and NF plots (Figure 1). The richness observed in NF plots was close to that expected by Chao2 and Jacknife2 estimators, whereas the richness observed in F plots was considerably lower than the estimated richness (Figure 1). The analysis of the rarefaction curves indicated that the F area was subsampled in the MGSP.
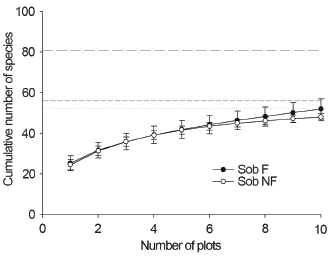
Figure 1
Richness of species observed (Sob) in plots covered by ferns (F) and ferns-free (NF) in the Mata dos Godoy State Park, Londrina, PR, Brazil, with 95% confidence intervals (errors bars). Species richness estimated by Chao2 (solid line to F and short dash line to NF) and Jacknife2 (long dash line to F and dotted line to NF)
Abundance of some species differed between F and NF plots (Figure 2). Among the species, it can be highlighted that Aspidosperma polyneuron (8x), Campomanesia xanthocarpa (Mart.) O. Ber (5x), Myrtaceae (4x), Holocalyx balansae Micheli (3x) and Chrysophyllum gonocarpum (Mart. & Eichler ex Miq.) Engl. (3x) were more abundant in F plot, whereas Ficus sp. (6x), Sorocea bonplandii (Baill.) W.C.Burger, Lanj. & de Boer (3x), Guarea kunthiana A. Juss. (2.5x) and Trichilia claussenii C. DC. (2.5x) were more abundant in NF plot (Figure 2).
The ICO was higher in NF than F plots indicating more open canopy in NF plots. No differences were found in other environmental parameters analyzed (Table 2).
Second experiment
For A. concolor (Figure 3A), more seedlings were observed in F than in NF and CF treatments, both at the beginning and end of the experiment. Among the species analyzed in this experiment, A. concolor showed the lowest recruitment and mortality rates (null in CF and NF areas).
Unlike the first experiment (p < 0.01), no differences for recruitment, mortality and abundance (Figure 3B) were observed for A. polyneuron among treatments. For E. edulis, only the recruitment of CF treatment was higher than in the F treatment (Figure 3C).
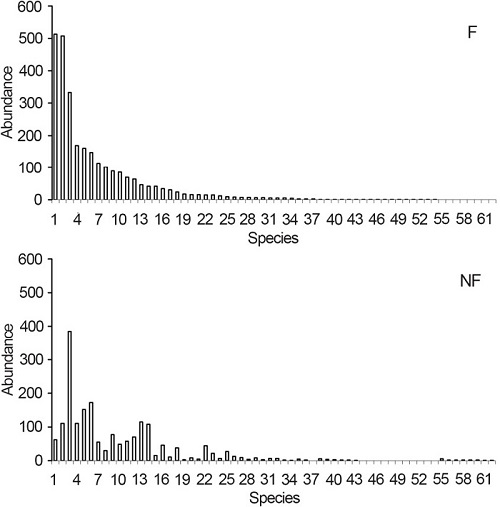
Figure 2
Abundance of species in plots covered by ferns (F) and ferns-free (NF) in Mata dos Godoy State Park, Londrina, PR, Brazil. Species were numbered in decreasing order of abundance in the plot F (see Table 1).
Environmental characterization (mean ± standard error) of sub-plots with ferns (F) and without ferns (NF) in the Mata dos Godoy State Park, Londrina, PR, Brazil. ICO = index of canopy openness
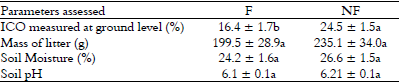
Different letters in rows indicate significant differences between treatments in the Kruskal Wallis test (p ≤ 0.05)
For G. kunthiana, there were no differences in recruitment and mortality among treatments. In the beginning of the experiment, fewer individuals were recorded in the F treatment than in the NF treatment. In the end of the experiment, the abundance in the F treatment was lower than both CF and NF treatment. Mortality in F treatment together with the recruitment in CF treatment resulted in a significant difference in the mean number of individuals between F and CF treatments at the end of the experiment (Figure 3D).
Despite equivalent numbers of individuals sampled in the first experiment, I. marginata showed fewer individuals in the F treatment than in CF treatment in the end of the experiment (Figure 3E).
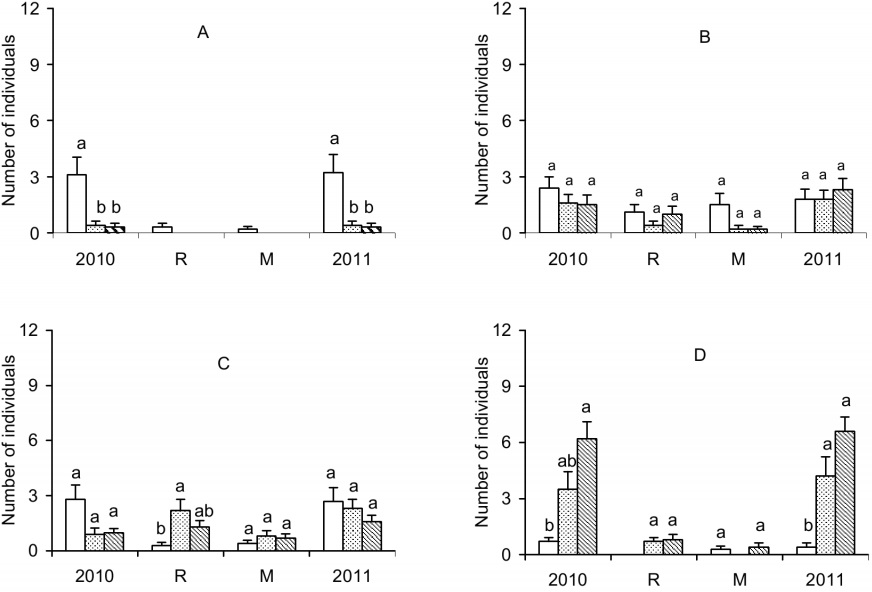
Figure 3
Average number of individuals in the beginning of the experiment (2010), recruited (R), mortality (M) and the end of the experiment (2011) per sub-plot in areas with ferns (white bars), with the fronds of ferns cut (dotted bars) and without ferns (bars with diagonal lines) in the Mata dos Godoy State Park, Londrina, PR, Brazil. A - Actinostemon concolor; B - Aspidosperma polyneuron; C - Euterpe edulis; D - Guarea kunthiana; E - Inga marginata. The vertical lines on the bars indicate the standard error. Different letters above the bars indicate differences between treatments by Friedman's variance analysis (a £ 0.05)
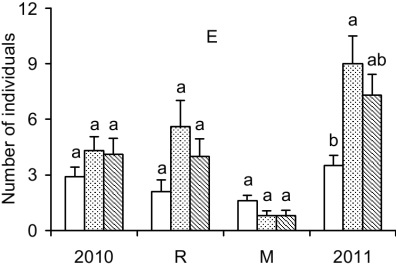
Figure 3 (Cont.)
Average number of individuals in the beginning of the experiment (2010), recruited (R), mortality (M) and the end of the experiment (2011) per sub-plot in areas with ferns (white bars), with the fronds of ferns cut (dotted bars) and without ferns (bars with diagonal lines) in the Mata dos Godoy State Park, Londrina, PR, Brazil. A - Actinostemon concolor; B - Aspidosperma polyneuron; C - Euterpe edulis; D - Guarea kunthiana; E - Inga marginata. The vertical lines on the bars indicate the standard error. Different letters above the bars indicate differences between treatments by Friedman's variance analysis (a £ 0.05).
Discussion
The understory filter may be nonspecific and regulate only the density of the seedling bank, or it may differentially influence emergence and survival of tree species, thereby controlling species composition and spatial structure of the seedling bank.
The higher estimated richness and higher abundance of woody seedlings were observed in area with ferns, suggesting that ferns could favor the regeneration of woody species in the MGSP. Shading promoted by ferns could produce suitable microsites for regeneration of shade-tolerant species, like A. polyneuron. These microsites would be especially important from April to September, a period of less rainfall and more open forest canopy (Bianchini et al., 2001). In this period, the increase in soil irradiance could promote higher evaporation and lower soil moisture and air humidity. Seedlings located in areas without ferns may be subject to water stress. Desiccation sensitivity is common in late-successional tropical forest trees (Vázquez-Yanes & Orozco-Segovia, 1984).
Studies have shown that Pteridium arachnoideum (Kaulf.) Maxon. (Silva Matos & Belinato, 2010; Gallegos et al., 2015) and P. aquilinum (L.) Kuhn (Marrs & Watt, 2006) are aggressive competitor species. This dominance is due to the allelopathic substances that inhibit germination and growth of other plants (Dolling, Zackrisson, & Nilsson, 1994) and reduces the number of propagules that reach the ground, holding them in their biomass (Marrs & Watt, 2006). It can be inferred that other species of ferns that have similar morphology, as the ferns of this study, also could inhibit the regeneration of woody species. However, the results presented here contradict this expectation. Gallegos et al. (2015) also observed higher recruitment and survival of Clusia spp. under P. arachnoideum.
Some environmental characteristic of ferns area favored the seedlings clumping of certain species. Environmental heterogeneity, vegetative propagation and limited dispersion contribute to the aggregation of individuals of some species (Hutchings, 1997). It is also known that environmental filters select species, which are functionally similar to their tolerance to abiotic and biotic conditions (Fukami, Bezemer, Mortimer, & van der Putten, 2005). This would explain the seedlings clumping, mainly of shade-tolerant species such as A. polyneuron, with abundance about eight times higher in ferns than in ferns-free areas.
The results analyses indicated that the abiotic factors analyzed did not contribute to the clumping of the sub-plots. However, the shading promoted by ferns can be important. Tsvuura, Griffiths, Gunton, Franks and Lawes (2010) observed that the shading promoted by herbaceous Isoglossa woodii C. B. Clarke (Acanthaceae) acts as a positive environmental filter on shade-tolerant tree species in a subtropical forest, for example: Eugenia woodii Dummer (Myrtaceae) and Drypetes natalensis (Harv.) Hutch. (Putranjivaceae). Therefore, seeds and seedlings of shade-tolerant species, such as the most abundant species in this study, can germinate and grow well in environments with low light levels, like microhabitat beneath ferns. Gallegos et al. (2015) recorded that Pteridium arachnoideum can facilitate seedling recruitment of Clusia spp. and potentially other late-successional species.
Although several studies suggested that removing fronds is key factor for ecological restoration of fern-infested sites (e.g. Silva Matos & Belinato, 2010; Gallegos et al., 2015), in this study, removing of fronds only affected the vital rates of Guarea kunthiana and Inga marginata and the recruitment of Euterpe edulis.
Ferns did not act as an environmental filter for Actinostemon concolor because the number of individuals in the first experiment was equivalent in F and NF, and cutting the fronds did not affect the results in the second experiment.
Ferns can act as a positive environmental filter for Aspidosperma polyneuron. The highest abundance of this species in F observed in the first experiment may be related to the formation of a seedling bank (characteristic of this species), since their seeds are considered indifferent to light (Sakita, Silva, & Paula, 2007) and could benefit from a wetter microclimate beneath the ferns. However, the results of second experiment make it impossible to define which process (recruitment or mortality) explains the greater abundance in F.
The results analyses for Euterpe edulis suggest that ferns did not act as an environmental filter for this species. No difference in the percentage of propagule germination was found for this species at various levels of shading (Tavares, Ramos, Aguiar, & Kanashiro, 2008). Although the survival of E. edulis seedling was higher in areas with greater canopy openness and brighter, the light levels no influenced the height growth of this species (Ribeiro, Martins, Lana, & Silva, 2011). Fruit predation, because of its attractiveness, especially for avifauna (Cavalheiro, 2006), would result in reduced fruit number and therefore less seedling. Ferns could reduce the view of fruit by predators, which could result in a greater number of seedlings in these areas. However, the largest recruitment in CF over F areas, especially in the January and April samples in the second experiment, suggests that recruitment could be higher in the NF areas, if there were no limitations on the number of propagules.
The Guarea kunthiana results analyses for both experiments suggests that this species is intolerant or has a low degree of shade tolerance in the early stages of development. Despite the large number of fruit that accumulated beneath the reproductive individuals, it was not observed seedlings under reproductive plants in the F plot during the sample period. However, seedlings were observed under reproductive plants in NF plot. The seedling intolerance to shading was also evidenced by Silva Matos and Belinato (2010) who observed high seedling mortality in sub-plots with Pteridium arachnoideum after a year of sampling. Therefore, ferns must have acted as a negative environmental filter on the population of G. kunthiana.
For Inga marginata, ferns did not act as an environmental filter, because there was no difference between areas in the first experiment and the first sampling of the second experiment. The greater abundance in CF than in F in the second sampling of second experiment was caused by other factors or processes not evaluated.
As shown by this study, ferns acted as selective filter on the tree species seedling, promoting spatial heterogeneity in structure and composition of the seedling community. This will certainly reflect in the future local tree community. Therefore, native ferns naturally occurring in preserved fragments should be considered in the management plans of forest fragments or reforestation with older age.
Conclusion
Abundance of the woody seedling bank was higher in areas with ferns. Composition and spatial structure of this seedling bank also differed between ferns and ferns-free areas because ferns differentially influenced emergence and survival of woody species. It can be concluded that the ferns functioned as an environmental filter favoring some tree species (A. polyneuron) while reducing the abundance of others species (G. kunthiana) in a fragment of Atlantic Forest in southern Brazil. However, further studies should be conducted including news forest remnants of the region, in order to evaluate the role of ferns as environmental filters in community composition.
Acknowledgements
We thank the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES) for the encouragement and financial support through the scholarship to the first author; the Instituto Agronômico do Paraná (IAPAR), for giving the climatic data and the Instituto Ambiental do Paraná (IAP), for allowing access to Mata dos Godoy State Park.
References
Bianchini, E., Pimenta, J. A., & Santos, F. A. M. (2001). Spatial and temporal variation in the canopy cover in a tropical semi-deciduous forest. Brazilian Archives of Biology and Technology, 44(3), 269-276.
Cavalheiro, A. L. (2006). Os palmiteiros do Parque Estadual Mata dos Godoy. In J. M. D. Torezan (Org.), Ecologia do Parque Estadual Mata dos Godoy (p. 61-64). Londrina, PR: Itedes.
Colwell, R. K. (2013). EstimateS 9.1.0. Storrs, CT: University of Connecticut.
Denslow, J. S., Newell, E., & Ellison, A. M. (1991). The effect of understory palms and cyclanths on the growth and survival of Inga seedlings. Biotropica, 23(3), 225-234.
Dolling, A., Zackrisson, O., & Nilsson, M. C. (1994). Seasonal variation in phytotoxicity of bracken (Pteridium aquilinum (L.) Kuhn). Journal of Chemical Ecology, 20(12), 3163-3172.
Elith, J., & Leathwick, J. R. (2009). Species distribution models: ecological explanation and prediction across space and time. Annual Review of Ecology, Evolution, and Systematics, 40(1), 677-697.
Fukami, T., Bezemer, T. M., Mortimer, S. R., & van der Putten, W. H. (2005). Species divergence and trait convergence in experimental plant community assembly. Ecology Letters, 8(12), 1283-1290.
Gallegos, S. C., Hensen, I., Saavedra, F., & Schleuning, M. (2015). Bracken fern facilitates tree seedling recruitment in tropical fire-degraded habitats. Forest Ecology and Management, 337, 135-143.
Gandolfi, S., Joly, C. A., & Rodrigues, R. R. (2007). Permeability-Impermeability: canopy trees as biodiversity filters. Scientia Agricola, 64(4), 433-438.
George, L. O., & Bazzaz, F. A. (1999). The fern understory as an ecological filter: emergence and establishment of canopy-tree seedlings. Ecology, 80(3), 846-856.
Guerin, N., & Durigan, G. (2015). Invasion impact by Pteridium arachnoideum (Kaulf.) Maxon (Dennstaedtiaceae) on a neotropical savanna. Acta Botanica Brasilica, 29(2), 213-222.
Hobbs, R. J., & Norton, D. A. (2004). Ecological filters, thresholds and gradients in resistance to ecosystem reassembly. In V. M. Temperton, R. J. Hobbs, T. Nuttle & S. Halle (Eds.), Assembly rules and restoration ecology (p. 72-95). Washington D.C.: Island Press.
Hutchings, M. J. (1997). The structure of plant population. In M. J. Crawley (Ed.), Plant Ecology (p. 97-136). Oxford, UK: Blackwell Scientific Publishers.
Lortie, C. J., Brooker, R. W., Choler, P., Kikvidze, Z., Michalet, R., Pugnaire, F. I., & Callaway, R. M. (2004). Rethinking plant community theory. Oikos, 107(2), 433-438.
Marrs, R. H., & Watt, A. S. (2006). Biological flora of the British Isles: Pteridium aquilinum (L.) Kuhn. Journal of Ecology, 94(6), 1272-1321.
Ribeiro, T. M., Martins, S. V. Lana, V. M., & Silva, K. A. (2011). Sobrevivência e crescimento inicial de plântulas de Euterpe edulis Mart. transplantadas para clareiras e sub-bosque em uma floresta estacional semidecidual, em Viçosa, MG. Revista Árvore, 35(6), 1219-1226.
Sakita, A. E. N., Silva, A., & Paula, R. C. (2007). Germinação de sementes de Aspidosperma polyneuron M. Arg. (peroba-rosa) sob diferentes condições de qualidades de luz e temperatura. Instituto Florestal Série Registros, 31, 203-207.
Scarano, F. R., Ribeiro, K. T., Moraes, L. F. D., & Lima, H. C. (1997). Plant establishment on flooded and unflooded patches of a freshwater swamp forest of southeastern Brazil. Journal of Tropical Ecology, 13(6), 793-803.
Silva Matos, D. M., & Belinato, T. A. (2010). Interference of Pteridium arachnoideum (Kaulf.) Maxon. (Dennstaedtiaceae) on the establishment of rainforest trees. Brazilian Journal of Biology, 70(2), 311-316.
Silva, U. S. R., & Silva Matos, D. M. (2006). The invasion of Pteridium aquilinum and the impoverishment of the seed bank in fire prone areas of Brazilian Atlantic Forest. Biodiversity and Conservation, 15(9), 3035-3043.
Silveira, M. (2006). A vegetação do Parque Estadual Mata dos Godoy. In J. M. D. Torezan (Org.), Ecologia do Parque Estadual Mata dos Godoy (p. 19-27). Londrina, PR: Itedes.
Tavares, A. R., Ramos, D. P., Aguiar, F. F. A. & Kanashiro, S. (2008). Jussara palm seed germination under different shade levels. Horticultura Brasileira, 26(4), 492-494.
Tsvuura, Z., Griffiths, M. E., Gunton, R. M., Franks, P. J., & Lawes, M. J. (2010). Ecological filtering by a dominant herb selects for shade tolerance in the tree seedling community of coastal dune forest. Oecologia, 164(4), 861-870.
Vázquez-Yanes, C., & Orozco-Segovia, A. (1984). Ecophysiology of seed germination in the tropical humid forest of the World: a review. In E. Medina, H. A. Mooney, & C. Vázquez-Yanes (Eds.), Physiological ecology of plants of the wet tropics (p. 37-49). The Hague, NE: W. Junk Publishers.
Vieira, D. C. M., & Gandolfi, S. (2006). Chuva de sementes e regeneração natural sob três espécies arbóreas de uma floresta em processo de restauração. Revista Brasileira de Botânica, 29(4), 541-544.
Walker, L. R., Landau, F. H., Velázquez, E., Shiels, A. B., & Sparrow, A. D. (2010). Early successional woody plants facilitate and ferns inhibit forest development on Puerto Rican landslides. Journal of Ecology, 98(3), 625-635.
Author notes
bianchi@uel.br
