Resumen: El trabajo se diseñó con el propósito de evaluar los efectos de variar la proporción de entregas de agua dependientes e independientes de la respuesta sobre la tasa y distribución temporal del responder en programas temporales señalados. Tres grupos de ratas se distinguieron por la proporción de reforzamiento dependiente-independiente de la respuesta (30-70, 50-50 y 70-30, respectivamente). Se utilizó un ciclo T de 60 s dividido en subciclos tD y t∆ de 30 s cada uno. En los ciclos programados con agua dependiente se presentó un tono durante tD, mientras que en los ciclos con agua independiente se presentó una luz. La tasa de respuesta fue mayor en los ciclos con agua dependiente de la respuesta, mientras que la distribución temporal del responder varió en función de la proporción, mostrando un decremento pronunciado en el responder al pasar del primero al segundo subintervalo del ciclo. Los hallazgos se discuten atendiendo al control diferencial desarrollado por los estímulos correlacionados a cada tipo de ciclo.
Palabras clave: reforzamiento dependiente e independiente de la respuesta, programas temporales, distribución temporal, tasa de respuesta, ratas.
Abstract: The study was designed to evaluate the effects of varying the proportion of response-dependent or response-independent water deliveries on rate and temporal distribution of responding in temporally defined schedules. Three groups of rats were distinguished by the ratio of response-dependent or response-independent reinforcement (30-70, 50-50, and 70-30, respectively). A Temporal (T) schedule 60 s was used, T cycle was divided in tD and t∆ periods 30 s each. Cycles programmed with response-dependent reinforcement were correlated with a tone and a light during the cycles with response-independent reinforcement. Temporal distribution of responding was as a function of response-independent reinforcement proportions identifying an abrupted decrement in responding from the first to the second sub-interval. Findings are discussed based on the control developed by the stimuli correlated to each type of cycle.
Keywords: response-dependent and response-independent reinforcement, temporal schedules, temporal distribution, response rate, rats.
Proporción de agua dependiente e independiente de la respuesta en programas temporales señalados
Proportion of dependent and independent-response water deliveries under temporal signaled schedules
Universidad Veracruzana

Esta obra está bajo una Licencia Creative Commons Atribución-NoComercial-CompartirIgual 4.0 Internacional.
Recepción: 06 Abril 2021
Aprobación: 01 Julio 2021
Con base en la relación de contingencia estímulo-estímulo y respuesta-estímulo se distinguieron dos tipos de condicionamiento: el respondiente y el operante, respectivamente (Skinner, 1938; Keller & Schoenfeld, 1950). También fue posible reconocer lo que técnicamente se denominó reforzamiento dependiente de la respuesta (contingente) y el reforzamiento independiente (no contingente) de la respuesta (Zeiler, 1968).
El interés por el estudio de la dependencia e independencia del reforzador ha sido motivado tanto teórica como experimentalmente (e.g., Cabrer et al., 1975). En el caso de la indagación empírica, diversos trabajos se han interesado por explorar los efectos del reforzamiento independiente sobre el mantenimiento de la respuesta (e.g., Bruner et al., 1994; Lachter et al., 1971; Lattal, 1974; Lattal & Bryan, 1976; Ribes et al., 1978). También se han estudiado los efectos de la secuencia de exposición a condiciones de reforzamiento dependiente y posteriormente a reforzamiento independiente y viceversa, permitiendo identificar los efectos que tiene la historia de reforzamiento (e.g., Freeman & Lattal, 1992; Polanco et al., 1979; Ribes & López, 1979; Serrano et al., 2015).
El estudio de los efectos del reforzamiento dependiente e independiente se puede identificar en varias áreas de investigación, extendiéndose desde el campo de la adquisición del responder bajo condiciones de reforzamiento demorado (e.g., Bruner et al., 1996) hasta estudios sobre discriminación condicional (e. g., Lattal, 1979).
El resultado consistente en la mayoría de los estudios es que bajo condiciones de reforzamiento dependiente se observa una tasa de respuesta más elevada que bajo condiciones de reforzamiento independiente de la respuesta. Por ejemplo, Lattal y Bryan (1976) manteniendo constante la tasa de reforzamiento dependiente por medio de un programa de intervalo fijo (IF 300 s), variaron entre condiciones la tasa de reforzamiento independiente mediante un programa de tiempo variable (TV 25, 45 y 120 s). Los autores reportaron que la tasa de respuesta fue una función monotónica decreciente que resultó de incrementar la duración del programa de TV.
En otro trabajo, Lattal y Maxey (1971) utilizando un programa múltiple de dos componentes, variaron entre fases las condiciones de reforzamiento dependiente e independiente de la respuesta. En una primera fase se programó el reforzador de manera dependiente en ambos componentes (IV 60 s – IV 60 s), en la segunda fase uno de los componentes cambió a reforzamiento independiente (IV 60 s – TV 60 s), mientras que durante la tercera fase ambos componentes entregaron el reforzador de manera independiente de la respuesta (TV 60 s – TV 60 s). Los autores identificaron que, al pasar de la Fase 1 a la Fase 2, la tasa de respuesta decrementó en el componente que cambió de reforzamiento dependiente (IV) a reforzamiento independiente (TV). Cuando en ambos componentes se programó el reforzador independiente de la respuesta (Fase 3) se observaron decrementos en la tasa de respuesta hasta llegar a niveles cercanos a cero.
En otro estudio, Weisman y Ramsden (1973) reportaron menores tasas de respuesta en un componente con reforzamiento independiente de la respuesta (TV) comparada con uno que entregaba el reforzador de manera dependiente (IV). Una característica en común de estos dos estudios es que, al utilizar programas múltiples, en la misma sesión se encontraban presentes ambas condiciones de reforzamiento (dependiente e independiente), a diferencia de otros estudios en los que se han evaluado los efectos de las secuencias de exposición a cada una de las condiciones de reforzamiento.
Otra característica de los estudios de Lattal y Maxey (1971) y Weisman y Ramsden (1973) es que además de que la tasa de reforzamiento dependiente e independiente se mantuvo igualada en ambos componentes, también la densidad o proporción de reforzamiento para cada caso se mantuvo constante, dado que se programó que en la sesión se presentara el mismo número de veces cada uno de los componentes de IV y de TV.
Un estudio que varió la proporción de reforzamiento independiente de la respuesta fue reportado por Kuroda et al. (2013) quienes manipularon entre condiciones la proporción de reforzadores que podían ser entregados de manera independiente de la respuesta (0, 10, 30, 70 y 100%). Al finalizar la duración del intervalo se determinó probabilísticamente si el reforzador se entregaba de manera dependiente con base en programas de intervalo (IV o IF) o independiente de la respuesta según programas de tiempo (TV o TF). Reportaron que la tasa de respuesta fue una función positiva de incrementar la proporción de reforzamiento dependiente.
Ribes et al. (2018) evaluaron los efectos de variar entre fases el momento en el que se programaba el reforzamiento independiente de la respuesta (al inicio y final, de manera aleatoria o en tercios de la sesión). Utilizando un programa definido temporalmente (Schoenfeld & Cole, 1972), programaron ciclos temporales (T) de 60 s divididos en subciclos tD y t∆ de 30 s cada uno. La primera respuesta de presión a la palanca en tD procuraba una gota de agua como reforzador, mientras que las respuestas en t∆ no tuvieron consecuencias programadas. Se programaron 20 entregas de agua independientes de la respuesta en caso de que los sujetos no presionaran la palanca durante el subciclo tD.
De manera general, encontraron que sin importar la manera en la que se programaran las entregas de agua independientes de la respuesta (al inicio y final, de manera aleatoria o en tercios de la sesión), la frecuencia de respuesta se mantuvo baja y relativamente estable tanto entre fases, como entre tD y t∆. Otro resultado fue que los sujetos mostraron elevados porcentajes de aguas perdidas, es decir, que no emitieron respuesta en tD en aquellos ciclos en los que no se programó la entrega independiente de la respuesta. Estos resultados son consistentes con otros reportados por Ribes et al. (2007, 2012) al no observar diferencias en el responder como resultado de variar el momento de programar la entrega de agua independiente de la respuesta dentro de la fase.
Ribes et al. (2018) mencionaron que: “En el presente estudio no manipulamos la proporción de entregas de agua dependientes o independientes de la respuesta (solo su ubicación), sin embargo, en estudios posteriores podría variarse la proporción de ciclos con entrega de agua C/NC con el fin de determinar su efecto sobre la frecuencia de respuestas en programas temporales señalados.” (p. 281).
Una manipulación de esta naturaleza sería importante porque permitiría dar generalidad a su resultado y extender los hallazgos sobre los efectos de variar la proporción de entregas de agua dependientes e independientes de la respuesta. Adicionalmente, en la mayoría de los estudios la principal variable dependiente es la frecuencia o la tasa de respuesta. Sin embargo, las propiedades del comportamiento no se restringen a la frecuencia de ocurrencia de una respuesta, también la distribución temporal del responder es una propiedad que puede ser sensible a las variaciones en la proporción de entregas dependientes e independientes de la respuesta. El presente estudio se diseñó con el propósito de evaluar los efectos de variar la proporción de entrega de agua dependiente e independiente de la respuesta sobre la tasa y la distribución temporal del responder en programas temporales señalados.
Nueve ratas macho Wistar de tres meses de edad al inicio del estudio e ingenuas experimentalmente. Los animales se mantuvieron con acceso libre al alimento (Purina Rodent chow 5001) y bajo un régimen de privación de agua por un periodo de h, con 30 minutos de acceso libre en su caja habitación después de cada sesión experimental. Los sujetos se mantuvieron alojados en el bioterio en un ciclo de luz: obscuridad de 12:12 horas iniciando el periodo de luz a las 6:00 am y con una temperatura entre 22° y 25°C. El cuidado y mantenimiento de los sujetos se realizó con base en el protocolo establecido en las “Especificaciones técnicas para la producción, cuidado y uso de los animales de laboratorio” (NOM-062-ZOO-1999) y con base en las regulaciones institucionales.
Se utilizaron nueve cámaras experimentales para ratas (Med Associates Inc. Modelo ENV-008). Cada cámara estuvo equipada con un dispensador de agua (ENV- 202M), un detector de entradas de cabeza al bebedero (ENV-254-CB) que permitió registrar la frecuencia de entradas de cabeza y una palanca retráctil ubicada en el panel operativo (ENV-112CM). El bebedero se localizó en el centro del panel operativo, mientras que la palanca estuvo ubicada a 2.5 cm del piso y requirió de una fuerza de 0.25 N para cerrar el micro interruptor. Arriba del bebedero se colocó una tecla traslucida de 2.5 cm de diámetro. La cámara experimental estuvo equipada con una bocina y una luz general localizada en la pared opuesta al panel operativo. Como reforzador se utilizó una gota de agua de 0.01 cc que se presentó por medio de la activación del dispensador de agua. Cada cámara experimental se colocó dentro de un cubículo de aislamiento acústico (ENV-022MD) con un ventilador que sirvió como ruido blanco y facilitó la circulación del aire al interior de la cámara. La programación, registro y colección de eventos se realizó mediante un equipo de cómputo, una interfaz (Med Associates Inc. Modelo SG-6080D) y el software MED-PC IV para ambiente Windows.
Los sujetos fueron expuestos por dos sesiones a un programa conjuntivo de reforzamiento continuo (rfc) -Tiempo Variable (TV) 30 s que concluyó al entregarse 60 aguas. Al día siguiente se descontinuó el programa TV y los sujetos fueron mantenidos durante dos sesiones con reforzamiento continuo, cada sesión concluyó cuando los sujetos produjeron 60 entregas de agua.
Concluido el mantenimiento de la respuesta los sujetos fueron puestos durante 30 sesiones a un programa definido temporalmente (Schoenfeld & Cole, 1972). Cada sesión constó de 60 ciclos T de 60 s, dividido en dos subciclos de 30 s cada uno (tD y t∆, respectivamente). Se conformaron tres grupos de tres sujetos que se distinguieron por la proporción de ciclos con agua dependiente y con agua independiente de la respuesta (Grupo 1 = 30-70, Grupo 2 = 50-50 y Grupo 3 = 70-30, respectivamente). De este modo y con base en los 60 ciclos por sesión, se programaron 18 ciclos con agua dependiente y 42 con agua independiente de la respuesta para los sujetos del Grupo 1; 30 dependientes y 30 independientes para los sujetos del Grupo 2; mientras que para los sujetos del Grupo 3 fueron 42 dependientes y 18 independientes de la respuesta.
En los ciclos programados con agua dependiente de la respuesta se presentó un tono de 80 dB y 2000 Hz en el subciclo tD; mientras que en los ciclos con agua independiente de la respuesta se presentó una luz blanca mediante el encendido de un foco (28-V); durante el subciclo t∆ no hubo eventos programados. La presentación de ciclos con agua dependiente e independiente de la respuesta variaron de manera semialeatoria con la restricción de que no se presentara más de tres veces consecutivas el mismo tipo de ciclo en una sesión.
En los ciclos dependientes, la primera respuesta en tD fue seguida por 3 s de acceso al agua y el apagado del tono, las respuestas posteriores durante el resto del ciclo no tuvieron consecuencias programadas. Si los sujetos no emitían respuesta durante el subciclo tD se perdía el reforzador programado en ese ciclo. En los ciclos con agua independiente de la respuesta la luz se mantuvo encendida durante todo el subciclo tD y se activó el bebedero hasta el final de dicho subciclo.
Para ambos tipos de ciclo se estimaron las tasas de respuesta de presión a la palanca y de entrada de cabeza al bebedero para cada una de las sesiones, así como la distribución temporal del responder a lo largo del ciclo T.
En la Figura 1 se presenta la tasa de respuesta (respuestas por minuto) por sesión de las presiones a la palanca para cada uno de los sujetos durante los ciclos con agua dependiente (izquierda) e independiente de la respuesta (derecha). De manera general se observa que la tasa de respuesta tiende a decrementar durante las primeras sesiones en ambos tipos de ciclo, llegando a niveles cercanos a cero en los ciclos con agua independiente de la respuesta; mientras que en los ciclos dependientes la tasa se mantiene entre una y dos respuestas por minuto a lo largo de las sesiones. También se puede observar que la tasa de respuesta no varió sistemáticamente en función de la proporción de ciclos con agua dependiente e independiente de la respuesta.
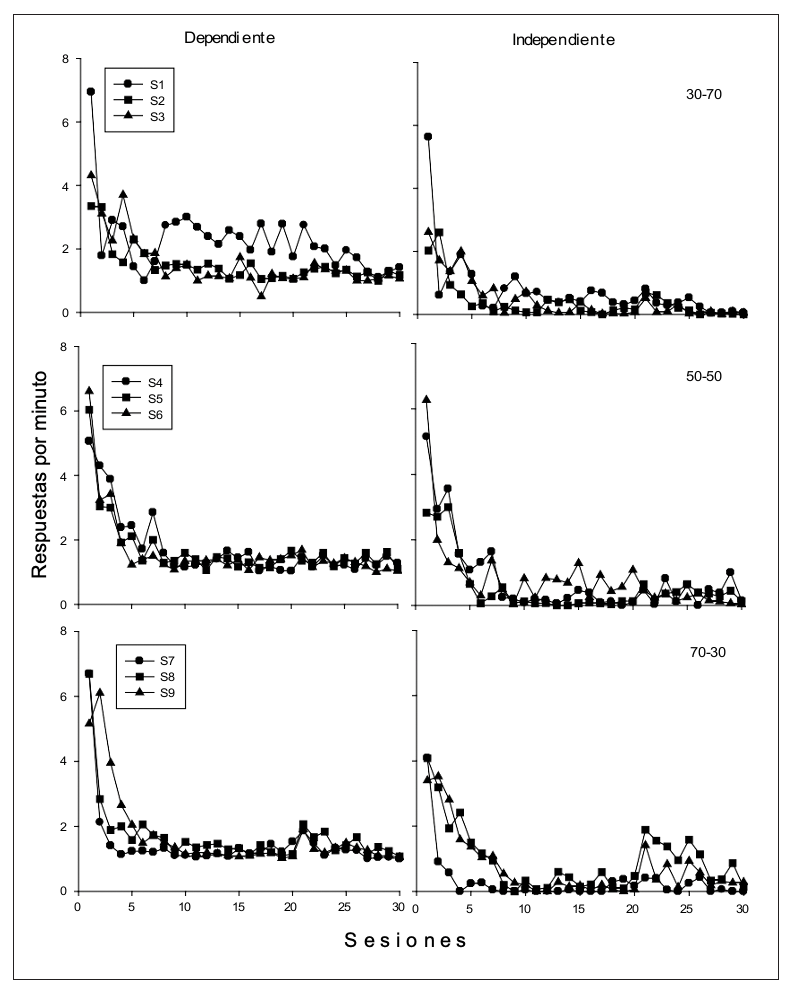
Figura 1
Tasa de respuesta (presiones a la palanca) por sesión para cada uno de los sujetos durante los ciclos con agua dependiente de la respuesta (izquierda) y con agua independiente de la respuesta (derecha)
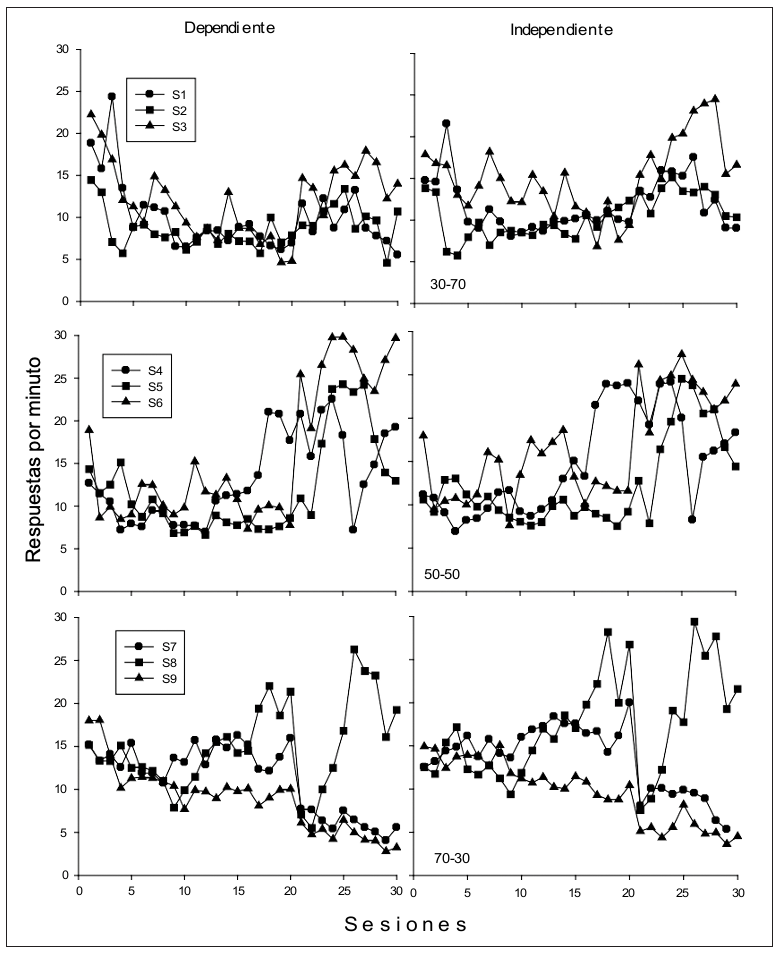
Figura 2
Tasa de respuesta (entradas de cabeza) por sesión para cada uno de los sujetos durante los ciclos con agua dependiente de la respuesta (izquierda) y con agua independiente de la respuesta (derecha).
La Figura 2 muestra la tasa de respuesta por sesión de las entradas de cabeza al bebedero para cada uno de los sujetos durante los ciclos con agua dependiente e independiente de la respuesta. A diferencia de las presiones a la palanca, en el caso de las entradas de cabeza se encontró mayor variabilidad en el responder, tanto entre sujetos como entre sesiones. No obstante, se puede observar que las tasas de respuesta son ligeramente superiores en los ciclos con agua independiente de la respuesta.
En la Tabla 1 se presenta la tasa de respuesta por sujeto, promedio de las últimas cinco sesiones, tanto de las presiones a la palanca como de las entradas de cabeza para cada tipo de ciclo. De manera consistente se encontró que la tasa de respuesta de presión a la palanca fue mayor en los ciclos con agua dependiente de la respuesta, no se identifican diferencias sistemáticas por variar la proporción de ciclos. En el caso de las entradas de cabeza se identifica un efecto inverso, es decir, la tasa de respuesta en los ciclos con agua independiente de la respuesta es consistentemente superior a la tasa en los ciclos con agua dependiente, excepto el Sujeto S6.
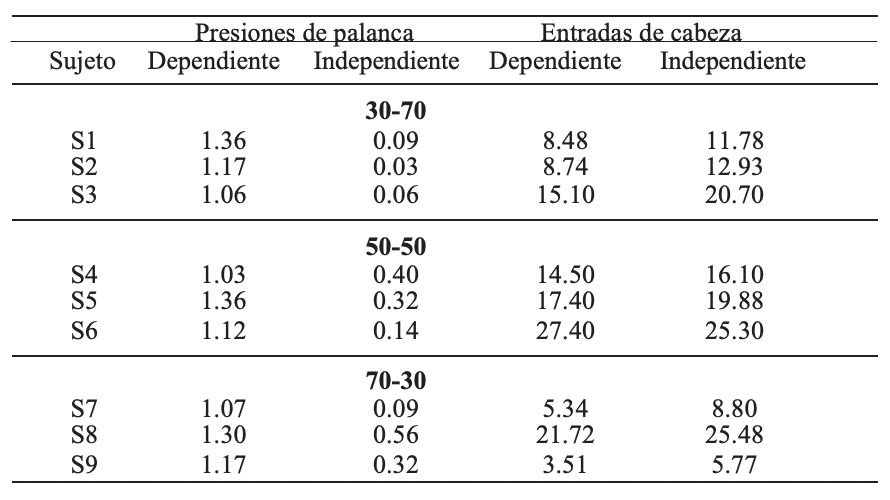
Promedio de las últimas cinco sesiones de la tasa de respuesta (respuestas por minuto) de presión a la palanca y de entrada de cabeza durante los ciclos con agua dependiente e independiente de la respuesta para cada uno de los sujetos
En la Figura 3 se presenta la tasa de respuesta de las presiones de palanca (promedio de las últimas cinco sesiones) para cada uno de los sujetos durante los subintervalos (cada subintervalo de 3 s) de los ciclos con agua dependiente e independiente de la respuesta. En los ciclos con agua dependiente se puede observar de manera consistente, para los sujetos de los tres grupos, un decremento pronunciado en el responder al pasar del primero al segundo subintervalo, también se puede identificar que la tasa de respuesta fue mas alta en función de un mayor número de ciclos con agua dependiente de la respuesta. En el caso de los ciclos con agua independiente de la respuesta se observa que la tasa de respuesta se mantiene prácticamente sin cambios en niveles cercanos a cero y sin observarse diferencias por la proporción en el tipo de ciclo.
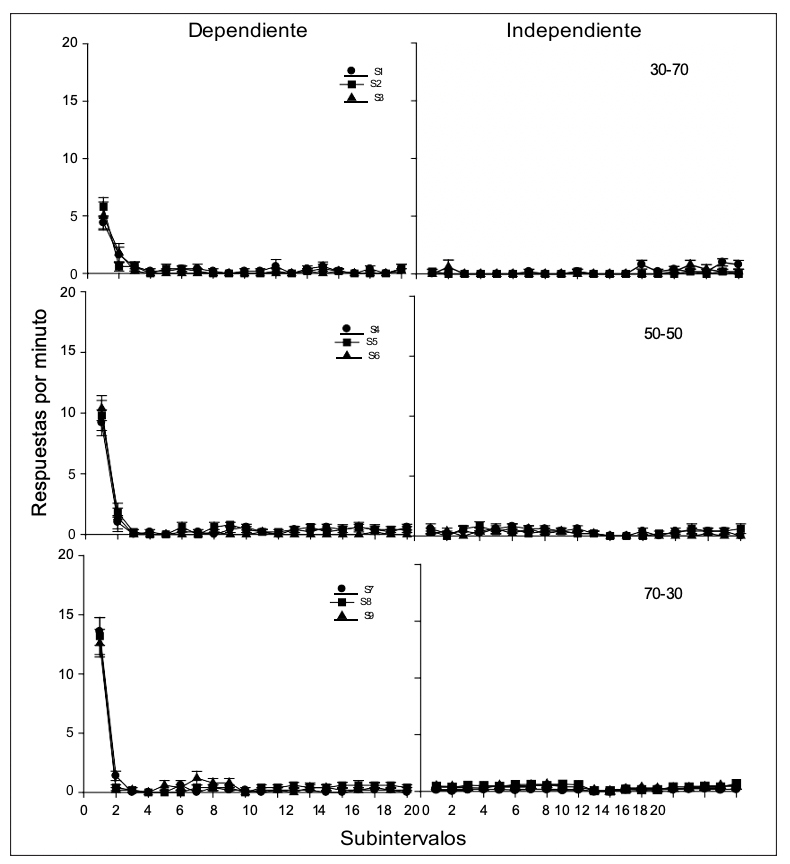
Figura 3
Tasa de respuesta (presiones de palanca) para cada uno de los sujetos en los subintervalos del ciclo temporal durante los ciclos con agua dependiente de la respuesta (izquierda) y con agua independiente de la respuesta (derecha).
La Figura 4 muestra la distribución temporal del responder (entradas de cabeza) para cada uno de los sujetos durante los subintervalos de los ciclos con agua dependiente e independiente de la respuesta (promedio de las últimas cinco sesiones). De manera general, en los ciclos dependientes se puede observar un decremento en el responder al pasar del primero al segundo subintervalo, mostrando una tendencia a incrementar gradualmente conforme transcurren los subintervalos. Este incremento es mucho más acentuado en los sujetos del Grupo 70-30 y menos marcado en los del Grupo 30-70. A diferencia de los ciclos dependientes, en el caso de los
ciclos con agua independiente de la respuesta se observa un incremento gradual en el responder durante los primeros subintervalos, siendo mayor para los sujetos del Grupo 30-70 y menos para los del Grupo 70-30. También se observa, de manera consistente en los sujetos de los tres grupos, un decremento abrupto en el responder al momento en el que se entregó el agua y que tiende a incrementar ligeramente durante el resto de los subintervalos.
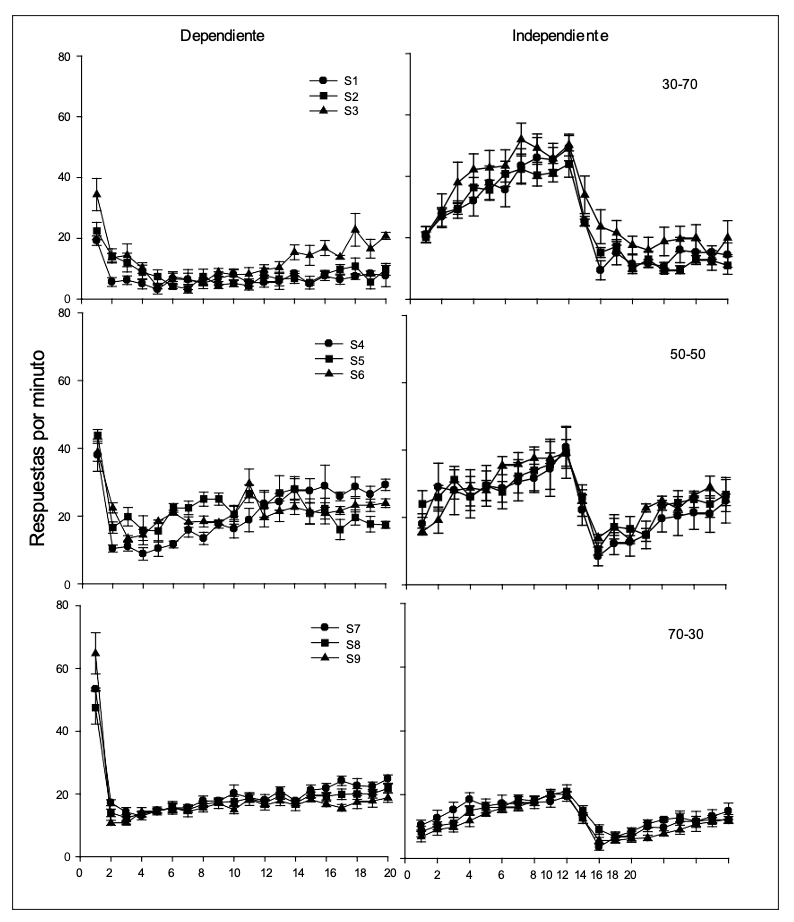
Figura 4
Tasa de respuesta (entradas de cabeza) para cada uno de los sujetos en los subintervalos del ciclo temporal durante los ciclos con agua dependiente de la respuesta (izquierda) y con agua independiente de la respuesta (derecha).
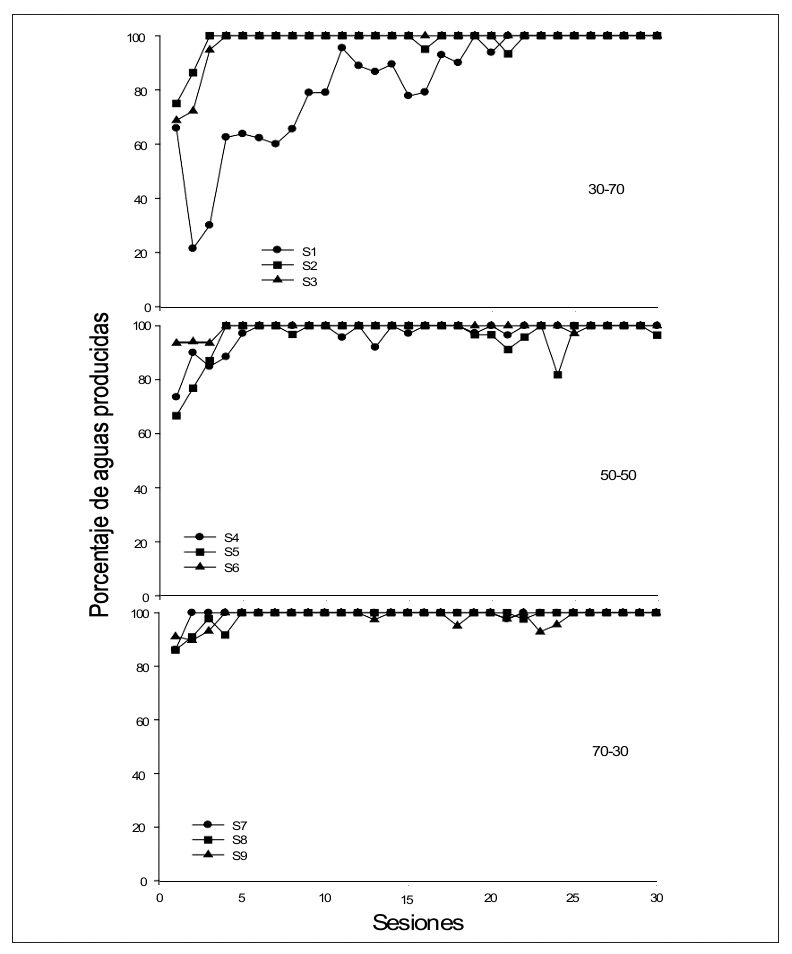
Figura 5
Porcentaje de aguas producidas por sesión de los sujetos de cada uno de los grupos.
En la Figura 5 se presenta el porcentaje de aguas producidas (dependientes de la respuesta) por sesión para cada uno de los sujetos en los tres grupos. De manera general se puede observar que sin importar la proporción de ciclos dependientes e independientes, el porcentaje de aguas se mantiene relativamente estable entre el 80% y el 100% en la mayoría de los sujetos y desde las primeras sesiones, excepto el Sujeto S1 del Grupo 30-70 en el que es hasta después de la sesión 15 que el porcentaje de aguas obtenidas es consistentemente superior al 80 %.
El propósito del presente trabajo fue evaluar el efecto de variar la densidad de ciclos con agua dependiente e independiente de la respuesta sobre el responder en programas temporales. De manera general se observó que la tasa de respuesta de presión a la palanca fue superior en los ciclos con agua dependiente sin importar la densidad de ciclos programados de cada tipo. Sin embargo, la tasa de respuesta de entrada de cabeza fue ligera y consistentemente más alta en los ciclos programados con agua independiente de la respuesta.
Haber observado superiores tasas de respuesta de presión a la palanca en los ciclos con agua dependiente y menores en los ciclos programados con agua independiente de la respuesta es un resultado consistente con lo reportado en otros trabajos (e.g., Lattal & Maxey, 1971; Weisman & Ramsden, 1973). No obstante, el no haber observado diferencias en la tasa en función de la proporción de ciclos de cada tipo contrasta con lo reportado por Kuroda et al. (2013) quienes identificaron que la tasa de respuesta incrementó en función de un mayor número de entregas de agua dependientes de la respuesta.
Esta diferencia en los resultados pudiera ser explicada por el señalamiento diferencial de cada tipo de ciclo. En el estudio de Kuroda et al. (2013) no había estímulos correlacionados con la programación del reforzador dependiente o independiente de la respuesta, mientras que en el presente estudio los ciclos con agua dependiente estaban señalados por un tono y los programados con agua independiente señalados por una luz, ello pudo haber contribuido al desarrollo de un control del estímulo minimizando el efecto de variar la proporción de entregas de agua de cada tipo. Esta interpretación se podría ver fortalecida también por las características del programa de reforzamiento utilizado en cada uno de los estudios. En el estudio de Kuroda et al. (2013) el reforzamiento dependiente se entregó con base en programas de IF o IV y las entregas independientes mediante programas de TF o TV; mientras que en el presente trabajo se utilizó un programa definido temporalmente, los cuales generan tasas de respuesta mas bajas que las generadas por programas de reforzamiento de Ferster y Skinner (1957) y que también pudo haber contribuido a la ausencia de un efecto por la densidad de entregas de agua dependientes e independientes de la respuesta.
No encontrar diferencias en la tasa de respuesta de presión a la palanca en función de la densidad de entregas dependientes e independientes, es un resultado consistente con lo reportado por Ribes et al. (2018) quienes tampoco observaron diferencias por variar el momento en el que se programaba la entrega de agua independiente de la respuesta al interior de la sesión.
Si bien la tasa de respuesta de presión a la palanca fue baja en los ciclos con agua dependiente de la respuesta y aún mas baja en los ciclos con agua independiente de la respuesta, no fue así en el caso de la tasa de respuesta de entradas de cabeza al bebedero. En este caso las tasas fueron sustancialmente mas elevadas e incluso superiores en los ciclos programados con agua independiente de la respuesta (ver Tabla 1). La diferencia entre la tasa de respuesta de presión a la palanca y de entrada de cabeza en los ciclos con agua dependiente e independiente de la respuesta es un resultado que pudiera ser explicado atendiendo al tipo de respuesta implicada dependiendo si la respuesta es procuradora de agua (presión a la palanca) o una respuesta consumatoria (entrada de cabeza al bebedero). En este sentido, las respuestas para cada caso podrían representar las propiedades descritas como amalgabilidad y amoldabilidad, respectivamente (Ribes, 2007).
En el caso de las entradas de cabeza, estas repuestas muestran un ajuste a la regularidad temporal con la que ocurren las entregas de agua, y en este sentido una correspondencia isomórfica con la distribución temporal de las entregas, las cuales son independientes de la respuesta. Por su parte, las presiones a la palanca son respuestas necesarias para la ocurrencia del agua y en ese sentido la distribución temporal de entregas de agua es dependiente (contingente) de la distribución temporal de la respuesta, la amalgabilidad se reconoce en la medida que el responder es efectivo produciendo cambios en el ambiente (i.e., la producción de aguas).
La distribución temporal de la respuesta de presión de la palanca se mantuvo sin variaciones a lo largo de ambos tipos de ciclo sin mostrar diferencias entre grupos, excepto en el nivel de respuesta en el primer subintervalo (ver Figura 3). Sin embargo, en el caso de la distribución temporal de las entradas de cabeza se pueden identificar cambios sistemáticos entre grupos, tanto al interior del ciclo como entre los tipos de ciclo (ver Figura 4). El incremento en el responder durante el subciclo tD en los ciclos con agua independiente y el súbito decremento al momento de activar el bebedero es un caso en el que el flujo conductual, en este caso de las respuestas de sondeo (entradas de cabeza) es interrumpido por la presentación del agua. A diferencia de la distribución de la respuesta de presión a la palanca, la distribución de entradas de cabeza sí fue modulada por la proporción, siendo más alto el incremento en los sujetos con una mayor cantidad de ciclos programados con agua independiente de la respuesta.
La distribución del responder durante los ciclos con agua dependiente también reflejó diferencias entre grupos. Sin embargo, el haber cancelado la señal correlacionada a tD por la ocurrencia de la primera respuesta no permitió identificar si también se pudiera haber generado una distribución temporal similar al patrón de respuesta identificado en los ciclos con agua independiente de la respuesta. Exploraciones futuras deberían considerar el mantenimiento de la señal correlacionada a tD.
Los resultados de distribución de repuestas de presión a la palanca son similares a los reportados por Serrano et al. (2015), quienes evaluando los efectos de la secuencia de exposición a entregas de agua dependientes e independientes de la respuesta también observaron un decremento en el responder justo al momento entregar el agua y de cancelar la señal correlacionada al subciclo tD por la emisión de la respuesta.
A diferencia del presente trabajo en el que también se registró la frecuencia de respuesta en los ciclos con entrega de agua independiente de la respuesta, en su estudio, Serrano et al. (2015) registraron el tiempo que los sujetos permanecían con la cabeza dentro del bebedero. Sin importar si la dimensión de la respuesta es la frecuencia o la duración, en ambos experimentos se observó la misma distribución del responder. Es decir, un incremento progresivo en el responder que se ve interrumpido y decrementa abruptamente al momento de entregar el agua al final del subciclo tD (ver Figura 4). Este resultado es interesante porque además de mostrar generalidad, permite reconocer efectos comunes en diversas propiedades de la respuesta.
Haber observado un elevado porcentaje de aguas producidas por la respuesta es un resultado consistente con otros estudios (e.g., Flores et al., 2007; Flores & Ribes, 2016; Mateos & Flores, 2009) pero inconsistente con los reportado por Ribes et al. (2018), quienes en su caso reportaron que sus sujetos recibieron muy pocas aguas producidas por la respuesta. Este efecto pudiera ser explicado nuevamente porque en el presente estudio los ciclos se encontraban diferencialmente señalados, haciéndolos mas discriminables.
En el estudio de Ribes et al. (2018) los mismos sujetos fueron expuestos a las diferentes condiciones en las que se varió el momento de entrega de agua independiente de la respuesta, mientras que en el presente estudio se utilizó un diseño de grupos procurando evitar posibles efectos de arrastre cuando se utilizan diseños intrasujeto (Sidman, 1950; Peron, 1991). Comparaciones adicionales con los presentes resultados hacen necesaria su replicación en la que los mismos sujetos son expuestos a las diferentes variaciones en la proporción de ciclos con agua dependiente e independiente de la respuesta.
Por otro lado, como se ha mencionado, en el presente estudio se correlacionó un estímulo diferente para cada tipo de ciclo, mientras que en el estudio de Ribes et al. (2018) no había un estímulo que diferenciara si el agua se entregaría de manera dependiente o independiente de la respuesta. Este aspecto es importante porque pudo haber contribuido al desarrollo de un control por parte del estímulo y con ello explicar las escasas respuestas de presión a la palanca en los ciclos programados con agua independiente. Adicionalmente, esta diferencia en el responder se pudo haber potenciado debido al uso de estímulos de diferente dimensión, que como se sabe desarrollan mayor control discriminativo que cuando los estímulos son de la misma dimensión (Harrison, 1991).
Para minimizar los efectos que pudiera tener el desarrollo de un control por parte de los estímulos correlacionados a cada tipo de ciclo, sería conveniente realizar una manipulación como la reportada en el presente trabajo, pero bajo condiciones no señaladas. Un estudio de este tipo podría contribuir a identificar los efectos de variar la densidad de entregas dependientes e independientes sobre el responder, haciendo comparables los resultados tanto en condiciones señaladas como en no señaladas.
Figura 1
Tasa de respuesta (presiones a la palanca) por sesión para cada uno de los sujetos durante los ciclos con agua dependiente de la respuesta (izquierda) y con agua independiente de la respuesta (derecha)
Figura 2
Tasa de respuesta (entradas de cabeza) por sesión para cada uno de los sujetos durante los ciclos con agua dependiente de la respuesta (izquierda) y con agua independiente de la respuesta (derecha).
Promedio de las últimas cinco sesiones de la tasa de respuesta (respuestas por minuto) de presión a la palanca y de entrada de cabeza durante los ciclos con agua dependiente e independiente de la respuesta para cada uno de los sujetos
Figura 3
Tasa de respuesta (presiones de palanca) para cada uno de los sujetos en los subintervalos del ciclo temporal durante los ciclos con agua dependiente de la respuesta (izquierda) y con agua independiente de la respuesta (derecha).
Figura 4
Tasa de respuesta (entradas de cabeza) para cada uno de los sujetos en los subintervalos del ciclo temporal durante los ciclos con agua dependiente de la respuesta (izquierda) y con agua independiente de la respuesta (derecha).
Figura 5
Porcentaje de aguas producidas por sesión de los sujetos de cada uno de los grupos.
