Artículos
Aprendizaje de reversión con y sin claves sociales humanas en perros domésticos: efectos del entrenamiento en detección de olores
Reversal learning with and without human social cues: effects of scent detection training
Aprendizaje de reversión con y sin claves sociales humanas en perros domésticos: efectos del entrenamiento en detección de olores
Revista Interamericana de Psicología/Interamerican Journal of Psychology, vol. 56, núm. 3, pp. 1-26, 2022
Sociedad Interamericana de Psicología

Esta obra está bajo una Licencia Creative Commons Atribución 4.0 Internacional.
Recepción: 29/06/2021
Aprobación: 01/09/2022
Resumen: El aprendizaje de reversión requiere que los sujetos inhiban una respuesta preponderante a un estímulo previamente reforzado y que cambien su conducta hacia una nueva contingencia de estímulo-refuerzo. Este permite medir el control inhibitorio, la capacidad de detectar cambios en las contingencias de reforzamiento y la flexibilidad, habilidades cruciales para los perros altamente entrenados. El objetivo de este estudio fue determinar si el entrenamiento en detección de olores modula el aprendizaje de reversión, comparando perros entrenados y perros de familia no entrenados en un paradigma de aprendizaje espacial. A su vez, se buscó evaluar si había diferencias en el desempeño de los perros comparando dos tareas: una no social y una social. En la primera la discriminación era únicamente entre claves no sociales (posición de la comida a la derecha vs a la izquierda) mientras que en la tarea social, a la clave de lugar que era reforzada en la discriminación inicial se le agregaba un estímulo social humano (posición del cuerpo del experimentador detrás del lugar reforzado). Los resultados muestran que no hubo diferencias entre grupos en ninguna de las dos tareas. Esto podría relacionarse con las características particulares de dicho entrenamiento. Asimismo, el desempeño fue mejor en la tarea social que en la no social durante la discriminación inicial, pero en la fase de reversión se observó el patrón inverso. Estas discrepancias pondrían en evidencia la importancia de considerar el efecto de las claves sociales humanas en los distintos tipos de aprendizaje. Asimismo, los presentes hallazgos contribuyen al conocimiento de las características cognitivas de los perros de detección de olores, una población escasamente estudiada hasta el momento.
Palabras clave: aprendizaje de reversión, entrenamiento, perros de detección, claves sociales humanas.
Abstract: Reversal learning requires that subjects inhibit a preponderant response towards a previously reinforced stimulus and shift their response toward a new stimulus-reinforcer contingency. This allows to measure inhibitory control, the ability to detect changes in reinforcement contingencies and flexibility, which are crucial skills for highly trained dogs. The aim of this study was to determine whether odor detection training modulates reversal learning, comparing trained dogs with untrained pets on a spatial learning paradigm. At the same time, the dogs’ performance was compared in two tasks: one non-social and one social. In the first one, the discrimination was only between non-social cues (position of the food on the right vs. on the left). In the social task, a human social stimulus was added to the spatial cue that was reinforced in the initial discrimination (position of the experimenter's body behind the reinforced side). The results show that there were no differences between trained and untrained groups in neither of the two tasks. This could be related to the particular characteristics of the training these dogs had received. Moreover, performance was better on the social task than on the non-social task during the initial discrimination, but the opposite pattern was observed in the reversal phase. These discrepancies highlight the importance of considering the effects of human social cues on different types of learning. The present findings contribute to the knowledge of the cognitive characteristics of scent detection dogs, a population scarcely studied until now.
Keywords: reversal learning, training, scent detection dogs, human social cues.
Introducción
Con el fin de adaptarse exitosamente a su entorno la mayoría de los organismos deben modificar sus conductas ante los cambios en sus circunstancias. Desde el punto de vista del aprendizaje, esta flexibilidad cognitiva suele ser evaluada a través de paradigmas experimentales como el de reversión (Wallis et al., 2011).
Generalmente, los protocolos de reversión consisten de dos fases. En una primera fase los sujetos aprenden una discriminación entre un estímulo que se asocia al refuerzo (ej. comida) y otro que se asocia al no-refuerzo. En una segunda fase, se produce una reversión de las contingencias, de modo que el estímulo previamente asociado al no-refuerzo es ahora el que predice la aparición de la comida y el inicialmente reforzado se asocia con su ausencia. Usualmente se observa que el aprendizaje de la reversión es más lento que el de la discriminación inicial (Izquierdo y Jentsch, 2012); dado que requiere un cambio cognitivo de la contingencia estimulo-refuerzo (Voytko, 1999). Asimismo, los déficits en el aprendizaje de reversión se manifiestan como errores perseverativos (Boutet et al., 2005; Tapp, 2003).
La reversión es un complejo proceso de aprendizaje que da cuenta de la habilidad de un organismo de suprimir una conducta preponderante previamente reforzada y ha sido utilizado exitosamente como una medida de control inhibitorio en diversas especies (Izquierdo et al., 2017; Tapp, 2003). Esta clase de aprendizaje dependería fundamentalmente de las funciones ejecutivas (Izquierdo y Jentsch, 2012) y permitiría medir la flexibilidad cognitiva (Wallis et al., 2011).
Sin embargo, según Izquierdo et al. (2017), esta visión clásica centrada en el control inhibitorio fue revisada considerando estudios más recientes en diversas especies. De este modo, el control inhibitorio se combina con un aprendizaje que requiere representaciones del refuerzo así como una expectativa de la posibilidad de un cambio (Costa et al., 2015; Izquierdo et al., 2017). Desde esta perspectiva, el aprendizaje de reversión implica codificar las señales de error de las predicciones basadas en los reforzamientos previos y realizar los ajustes conductuales pertinentes (e.g., Chase et al., 2011).
Adicionalmente, en el aprendizaje de reversión intervienen procesos atencionales. Tal como plantea el modelo de Pearce y Hall (1980), los resultados sorpresivos e inesperados que llevan a un error de las predicciones están asociados con un incremento de la atención. Este incremento podría ser clave para el aprendizaje de la nueva contingencia de refuerzo, combinándose así procesos atencionales con procesos de aprendizaje. En este sentido, además de los procesamientos de arriba hacia abajo (top-down) y de abajo hacia arriba (bottom-up) de la atención selectiva (Posner y Petersen, 1990), recientemente se ha postulado una tercera categoría denominada de atención motivada por la experiencia (experience-driven) (Oemisch et al., 2017). Esta última incluye la historia de selección reciente de un individuo y el aprendizaje basado en el refuerzo (Anderson et al., 2013; Awh et al., 2012). Por ejemplo, el valor previamente aprendido de una recompensa es un modulador de qué estímulos se priorizan ante una determinada situación. Por lo tanto, la atención motivada por la experiencia estaría involucrada en el aprendizaje de reversión (e.g., Anderson et al, 2013).
En el presente trabajo nos focalizaremos en el aprendizaje de reversión en perros domésticos. Existe una extensa tradición en el estudio de los procesos de aprendizaje en esta especie (Pavlov, 1927; Thorndike, 1898) que ejemplifican cómo estos modulan las conductas de los perros domésticos (e.g., Barrera et al., 2011; Elgier et al., 2012) e incluso influyen en el desarrollo de los vínculos con las personas (Barrera et al., 2010). Sin embargo, hasta el momento, la mayor parte de los estudios de reversión en esta especie se han centrado en su valor como indicador temprano de deterioro cognitivo (Piotti et al., 2018). En este línea, diversas evidencias muestran una fuerte asociación entre la edad y el desempeño en esta tarea, hallándose un deterioro en los perros mayores (e.g., Milgram et al., 1994; Mongillo et al., 2013; Mongillo et al., 2017; Piotti et al., 2018; Tapp, 2003; Van Bourg et al., 2021). Dada esta escasez de antecedentes, el presente estudio se propone indagar en la importancia de dos factores que podrían incidir en el aprendizaje de reversión: el entrenamiento en detección de olores y la presencia o ausencia de una clave social humana junto a una clave no social como estímulos discriminativos.
Respecto al efecto del entrenamiento en detección de olores, este se enmarca en el estudio de la importancia de las experiencias previas en el comportamiento de los perros. Fagnani et al. (2016) proporcionaron una evidencia acerca de la relevancia de dichas experiencias. En su trabajo compararon a perros de refugio, quienes tienen menos contacto cotidiano con las personas, y perros de familia en su desempeño en la tarea de reversión llamada “A no B”. Esta tarea consta de dos fases, la primera o entrenamiento en la que una persona muestra el refuerzo al sujeto y lo esconde debajo de un recipiente (A) y le permite al perro obtenerlo. En la segunda fase o prueba, la persona coloca el refuerzo en la primera ubicación (A) y luego, llamando la atención del perro y a la vista del mismo, quita el refuerzo de la ubicación A y lo traslada a otro recipiente (B). Los resultados de este estudio mostraron que los perros de refugio cometieron una mayor cantidad de errores perseverativos, eligiendo el lugar A en la prueba, que los de familia. Sin embargo, Cavalli et al. (2018) no hallaron diferencias entre estos últimos y perros que participaban en Intervenciones Asistidas por Animales, caracterizados por un mayor contacto cotidiano con las personas.
Respecto al efecto especifico de los entrenamientos avanzados, se postula que estos podrían modular el aprendizaje de reversión dado que involucran la inhibición de respuestas inapropiadas en contextos de trabajo (Wright et al., 2011). En favor de esta hipótesis, Barrera et al. (2018) hallaron que los perros entrenados en rescate acuático tenían un rendimiento superior al de perros de familia en la tarea A no B. Contrariamente, Kelly et al. (2019) mostraron que los perros de trineo tuvieron un peor desempeño que los de familia en la misma tarea. Sin embargo, cabe destacar que los perros de trineo vivían alojados en caniles, con bajo contacto con las personas y que ciertas características de los perros como la raza, el sexo y la edad no fueron balanceadas entre los grupos. Estos factores no permiten arribar a conclusiones sólidas acerca de la capacidad de ambos grupos de aprender la tarea de reversión. En síntesis, las evidencias tanto acerca del efecto de las experiencias previas de los perros como de los entrenamientos sistemáticos avanzados sobre el aprendizaje de reversión son controversiales.
Respecto al segundo factor, la presencia de claves sociales humanas ha mostrado modular distintos aprendizajes y comportamientos de los perros. Por ejemplo, ante dos opciones que contenían la misma cantidad y calidad de comida, los perros prefirieron aquella que la persona elegía y ante la cual emitía claves ostensivas positivas tales como decir “que rico” (Prato Previde et al., 2008). Más aún, en una situación de elección similar, los perros prefirieron la opción de comida en la que había un video con la cara del dueño visible y orientada hacia ellos, en comparación con la otra fuente de comida en la que el video mostraba al dueño de espaldas al perro (Bolló et al., 2021). Estas y otras evidencias darían cuenta del valor reforzante de los estímulos sociales humanos (e.g., Cook et al., 2016). Por otro lado, en algunos casos, las claves sociales pueden funcionar como distractores, deteriorando el desempeño. Por ejemplo, en una tarea de elección de objeto los perros optaban por el recipiente que era señalado por la persona, aun cuando se encontraba vacío, en lugar del otro recipiente que contenía el olor a la comida (Szetei et al., 2003). Asimismo, las claves ostensivas produjeron un detrimento del aprendizaje social en una tarea de manipulación (Range et al., 2009).
En relación al aprendizaje de reversión, las evidencias son también contrapuestas. Por un lado, se ha observado que en la tarea A no B los perros cometieren más errores en los contextos comunicativos en los que un humano emitía claves ostensivas, que en un contexto no comunicativo en el cual la persona estaba presente pero no emitía ninguna clave (e.g., Topál et al., 2009). Resultados similares se hallaron comparando con una versión no social de la misma tarea (Gergely et al., 2016; Topál et al., 2009). Estos hallazgos indicarían que la mera presencia de una persona en la tarea de reversión podría deteriorar el desempeño de los animales. No obstante, existen evidencias que sugieren que la naturaleza social o no social de las claves no influye en el desempeño en una tarea de reversión (Wobber y Hare, 2009). Más aún, Fagnani et al. (2020) mostraron que al final de la fase de reversión de la tarea A no B los perros hicieron más respuestas correctas en la tarea social que en la no social, revelando un efecto facilitador de la presencia de las personas.
En suma, los resultados respecto al rol de los estímulos sociales en el desempeño de los perros en el aprendizaje en general y durante una tarea de reversión en particular son, hasta el momento, controversiales y no permiten determinar claramente la influencia de los mismos. Sin embargo, uno de los factores que podría ser común a los distintos trabajos es que los perros prestarían una elevada atención a los estímulos sociales humanos y eso podría modular su comportamiento en diferentes situaciones. Esto sería consistente con la hipótesis de Gácsi et al. (2009) según la cual la atención sostenida a las personas habría sido uno de los factores de selección más importantes durante la domesticación de los perros. Dado que, como antes mencionáramos, la reversión incluye un componente atencional, es posible que cuando la clave humana es la que predice el refuerzo este aprendizaje se vea facilitado. En cambio, cuando deben guiarse por una clave no social para acceder al mismo, o incluso deben ignorar un estímulo social, el desempeño se dificulte.
Teniendo en cuenta ambos factores, en el presente trabajo nos proponemos indagar en la influencia del entrenamiento avanzado en una tarea de reversión en la que todos los perros serán expuestos a dos condiciones, una sin y otra con claves sociales. Para ello, en la condición no social se utilizará el lugar (izquierda - derecha) como clave espacial, ya que se ha observado que los perros aprenden más fácilmente una tarea de reversión cuando la clave es la ubicación espacial de los estímulos, en vez de las características físicas de los mismos (Piotti et al., 2018). En cambio, en la condición con claves sociales se agregará a la clave de lugar un estímulo social que consistirá en la posición del cuerpo de un experimentador (este se parará durante toda la tarea detrás del lugar que será reforzado durante la fase de adquisición – ver métodos).
Respecto al entrenamiento, nos focalizaremos particularmente en el estudio de los perros entrenados para la detección de olores, dada su relevancia social y su uso extendido. Estos perros han mostrado ser capaces de detectar una amplia variedad de sustancias, desde aquellas más tradicionales como narcóticos, explosivos, el olor de una persona o de restos humanos, a sustancias menos comunes, tales como personas con determinadas enfermedades, plagas o animales salvajes (Simon et al., 2020).
Si bien actualmente no hay trabajos que comparen el desempeño de los perros de detección con el de los perros de familia en tareas de reversión, algunos estudios han comparado estos grupos en otras tareas inhibitorias. Una de las más utilizadas es la prueba de cilindro en la que el perro debe evitar la respuesta preponderante de acercarse directo a la comida (y tocar el cilindro que es transparente) y en su lugar, hacer un desvío y tomar la comida de uno de sus extremos. En esta línea, Tiira et al. (2020) encontraron que aquellos perros que habían mostrado mayor control inhibitorio en la prueba del cilindro fueron más exitosos en una prueba posterior de detección del olor a explosivos. Interesantemente, aquellos perros que cometían más errores en la tarea del cilindro abandonaban antes la búsqueda del explosivo y persistían menos intentando abrir un recipiente que contenía comida. Contrariamente, MacLean y Hare (2018) no hallaron asociaciones entre el desempeño en la tarea de cilindro y el éxito en graduarse como perro de detección. Tampoco se hallaron asociaciones entre las habilidades de detección de olores y el rendimiento en una tarea de cerca en forma de V en la que los perros deben hacer un rodeo para acceder a la comida que esta visible detrás del vértice de la cerca (Tiira et al., 2020). En conclusión, si bien se postula que los perros de trabajo que realizan tareas prolongadas deben tener altos niveles de autocontrol (Tiira et al., 2020), las evidencias hasta el momento distan de ser concluyentes.
Por otro lado, el aprendizaje de reversión requiere que los animales dejen de persistir en su respuesta previamente aprendida para realizar la nueva conducta. Algunos trabajos indagaron en la persistencia de respuestas aprendidas de los perros de detección aunque utilizando otros paradigmas. Por ejemplo, Lazarowski et al. (2020) observaron que los perros de detección de olores seguían más frecuentemente el señalamiento para encontrar comida cuando este era realizado por una persona desconocida en comparación con los perros de familia. A su vez, en una tarea irresoluble los perros de detección persistieron más tiempo interactuando con el problema que los de familia (Lazarowski et al., 2020). Más aun, los perros que persistían menos en resolver el problema eran aquellos que abandonaban más rápidamente la búsquedas de explosivos (Tiira et al., 2020). De este modo, los perros de detección parecerían ser más persistentes que los de familia, tanto en respuestas sociales como no sociales. En este sentido, una mayor persistencia de las respuestas aprendidas podría ir en detrimento del aprendizaje de reversión.
En conclusión, dada la escasez de datos de la literatura y la naturaleza controversial de los hallazgos antes mencionados, no es posible establecer predicciones claras acerca de los resultados del presente estudio. Esto pone de relevancia la necesidad de llevar a cabo más investigaciones en el área. El estudio de la influencia del aprendizaje en la conducta de los perros aporta información relevante para el desarrollo de técnicas de entrenamiento eficaces así como para mejorar la calidad de vida de los animales de compañía. Este aporte será de gran importancia específicamente para los perros de detección de olores dado su extendido uso, las dificultades para completar exitosamente el entrenamiento (Wilsson y Sundgren, 1997) y el retiro prematuro del servicio de muchos perros costosamente entrenados (Evans et al., 2007).
Método
Sujetos
Se evaluaron 40 perros domésticos de los cuales 6 debieron ser retirados de la muestra por mostrar desinterés en la comida o la tarea, distracción o intenso apego al guía.
Los perros fueron seleccionados según la variable entrenamiento, conformándose dos grupos: el primero era el de perros entrenados en detección de olores (PD, N=17), ocho de los cuales pertenecían a la Base Naval Puerto Belgrano, Argentina y 9 al grupo de K9 de los bomberos voluntarios de la Ciudad de Punta Alta y del Departamento de Búsqueda y Rescate con Canes de la Federación de Bomberos Voluntarios de la Provincia de Córdoba. Si bien la mayoría vivía en caniles (N = 13), todos tenían un frecuente contacto con su guía. El entrenamiento en general consistía en buscar personas u objetos desaparecidos en un campo abierto donde había obstáculos. Cuando los perros lo hallaban debían señalar la posición ladrando y permaneciendo en el lugar indicado.
El segundo grupo era el de los perros de familia (PF, N=17). Todos ellos vivían en casas de familia como mascotas y no poseían entrenamiento previo. Estos perros fueron convocados a través de contactos personales apareando lo máximo posible los factores edad, sexo, raza y estado de castración con los del grupo PD. La Tabla 1 muestra las características de los perros de ambos grupos.
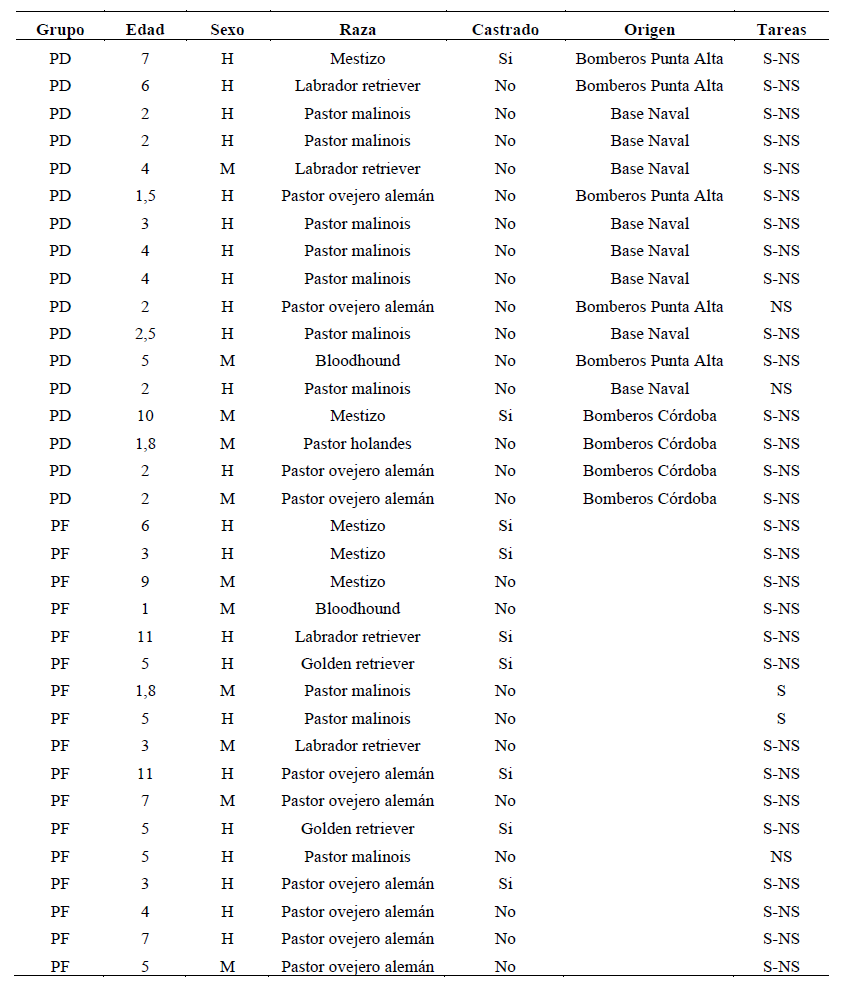
Nota. PD: perros de detección, PF: perros de familia, H: hembra, M: macho, S: tarea social, NS: tarea no social
Algunos perros no pudieron ser evaluados en ambas tareas, de modo que el tamaño final de la muestra fue de 15 PD y 16 PF en la tarea social y 17 PD y 14 PF en la tarea no social.
Se les solicitaba a los dueños que los perros no hayan comido 4 horas antes de la prueba de modo de aumentar su motivación.
Materiales
Se utilizaron dos recipientes opacos idénticos de 20 cm de diámetro y 8 cm de altura. Los mismos estaban untados con hígado y tenían un doble fondo en el que se colocaban cinco pedazos de hígado para controlar las claves de olor. Este procedimiento se utiliza típicamente como control, ya que si sólo hay un trozo de comida en el recipiente correcto y el otro se encuentra vacío, los perros pueden guiarse por la clave olfativa para encontrar el refuerzo. Los dos recipientes se ubicaban en el piso, sobre unas marcas realizadas con cinta a 1,5 m de distancia uno de otro. El perro se hallaba a 2 m junto a un experimentador (E1) que lo sostenía con correa de modo que formaban un triángulo con los recipientes. Un segundo E (E2) se mantenía alejado por detrás en la habitación y se encargaba de rellenar uno de los dos recipientes (ver Figura 1). La evaluación fue filmada con una cámara SONY DCR 308, colocada en un trípode detrás y hacia el costado de donde se ubicaban el perro y E1. Los perros fueron evaluados en un lugar familiar para ellos. Se utilizó como refuerzo trozos de aproximadamente 2 cm3 de hígado cocido.

Figura 1
Imagen de la situación experimental de la tarea no social (A) y de la social (B)
Procedimiento
Se utilizó una tarea de elección de objeto con una versión no social donde la clave espacial era la posición del recipiente con comida (derecha o izquierda) y una versión social dónde a la clave espacial se le sumaba la clave de posición del cuerpo del E2. Todos los animales fueron evaluados en ambas tareas con un intervalo de entre 30 y 45 días entre ellas. El orden de comienzo de las tareas fue contrabalanceado entre los perros.
Tarea de reversión no social
Primera fase: Pre entrenamiento
Cuando E1 ubicaba al perro en el punto de partida, E2 se dirigía al medio de las marcas, colocaba los recipientes en el suelo y luego daba dos pasos hacia atrás permaneciendo de espaldas al perro. Luego E1 lo guiaba gentilmente hacia el lado reforzado de modo de instigar la respuesta correcta. Esto se repitió por 6 ensayos de modo de que el perro aprendiera que ese recipiente contenía comida. El lugar en el que se encontraba el recipiente con comida fue contrabalanceado entre los perros.
Al finalizar los 6 ensayos se realizó un intervalo de 20 segundos antes de comenzar con la fase de adquisición.
Segunda fase: Adquisición
Al comenzar cada ensayo, E1 ubicaba al perro en el punto de partida. Luego el E2 colocaba los recipientes en el suelo y daba dos pasos hacia atrás permaneciendo de espaldas al perro (Ver Figura 1A). E1 aflojaba la correa y avanzaba dos pasos cortos hacia el centro de ambos recipientes para que el animal pudiera elegir. El lugar del recipiente que contenía comida para cada perro era el mismo que durante el pre entrenamiento. Si el perro se acercaba a dicho recipiente (e.g., derecha), se consideraba una respuesta correcta. En ese caso, este podía comer el trozo de hígado y E1 lo felicitaba diciéndole en tono positivo “muy bien y el nombre del perro”. Si se aproximaba al recipiente incorrecto (lado no reforzado, e.g., izquierda), E1 le decía “no” y volvía al punto de inicio. En el intervalo el E2 rellenaba el recipiente. El lugar en el que se colocaba la comida (derecha-izquierda) fue contrabalanceado entre los perros.
En esta fase se requería que el sujeto recuperara el refuerzo hasta alcanzar el criterio de un mínimo de 8 de 10 ensayos correctos. Se realizaba un máximo de 30 ensayos, agrupados en 3 sesiones de 10 ensayos cada una, con un intervalo de 1 minuto entre ellas. Todos los perros evaluados alcanzaron dicho criterio y por lo tanto continuaron en el estudio. Un minuto después del último ensayo se pasaba a la siguiente fase.
Tercera fase: Reversión
El procedimiento era idéntico al de la adquisición, excepto que se realizaba un cambio en la contingencia de los estímulos. El lado reforzado en la fase de adquisición pasaba a ser el no reforzado y viceversa (es decir que el lado originalmente no reforzado en la fase de adquisición, en esta nueva fase contenía comida). Se realizaron sesiones de 10 ensayos con un intervalo entre sesiones de 1 minuto. Esta fase duraba hasta que el perro alcanzara el criterio de 8 de 10 respuestas correctas o hasta un máximo de 4 sesiones.
En ambas fases una elección se codificaba como correcta si el perro se acercaba a menos de 10 cm del recipiente que contenía la comida; y como incorrecta si se acercaba a menos de 10 cm del recipiente vacío. Por último, una respuesta era codificada como no elección si el perro no elegía ningún recipiente transcurridos 15 s desde que el E1 aflojaba la correa. En todas las fases los intervalos entre ensayos fueron de 20 s. Al finalizar la totalidad de la tarea se le ofrecían al perro 5 trozos de hígado para descartar que el desempeño del mismo durante la fase de reversión haya estado afectado por un fenómeno de saciedad.
Tarea de reversión social
Esta tarea era idéntica a la anterior a excepción de que la clave fue compuesta. Junto a la clave de lugar se utilizaba la posición del cuerpo del E2. Al comienzo del ensayo el E2 se dirigía al medio de los recipientes, los apoyaba en el suelo y permanecía parado en ese lugar, hasta que el perro mirara. En ese momento se colocaba detrás del recipiente con comida (e.g., derecha). De este modo, siguiendo el ejemplo, la clave que predecía el refuerzo en esta fase era persona-lugar a la derecha (ver Figura 1B). En el test de reversión, el compuesto original ya no era reforzado (el recipiente se encontraba vacío) y el perro debía aprender que el refuerzo ahora estaba asociada a la clave estimular opuesta: ausencia de persona-lugar a la izquierda. Se consideraba como respuesta correcta que el perro fuera al lugar opuesto al de la adquisición (e.g., izquierda) detrás del cual no se hallaba el E2.
Las demás condiciones eran iguales a las de la tarea no social.
Análisis de datos
Las elecciones fueron medidas en vivo por dos experimentadores obteniéndose un acuerdo del 100% entre ambos.
Las variables registradas en cada tarea fueron:
- 1. Número de ensayos necesarios para llegar al criterio de aprendizaje en la fase de adquisición.
- 2. Número de ensayos necesarios para llegar al criterio de aprendizaje en la fase de reversión.
- 3. Número de no elecciones en la fase de reversión. Sin embargo, dado que esta variable presentó una gran cantidad de ceros, se la transformó en dicotómica. Para ello se consideró en cada perro si presentó al menos una no elección o no presentó ninguna durante toda la fase de reversión.
Para las principales variables se realizaron cálculos del tamaño muestral mínimo basados en la varianza obtenida en pruebas preliminares con 8 animales y estimando una potencia mínima de 0.80. Para ello se utilizó el módulo de cálculo de tamaño muestral de Infostat. Se observó que para la mayoría de las variables se requerían entre 12 y 14 perros para alcanzar dicha potencia.
Previo al análisis, se realizaron pruebas para determinar la distribución de los datos y establecer si los grupos tenían homocedasticidad, en caso de que su distribución fuera normal. Siendo que en todos los casos las variables no siguieron una distribución normal (Shapiro-Wilks, P < 0.01 en todos los casos) se decidió evaluar potenciales diferencias entre los grupos mediante análisis no paramétricos.
En la tarea social y la no social, se comparó entre los grupos PD y PF el número de ensayos necesarios para llegar al criterio de aprendizaje en las fases de adquisición y reversión, utilizando la prueba U de Mann Whitney. Se utilizó la prueba de comparación de proporciones de Fisher para comparar entre grupos y entre tareas la proporción de perros que presentaron al menos una vez una no elección en la fase de reversión. Se realizó un análisis considerando el total de la muestra y otro para cada grupo por separado. A su vez, se comparó el desempeño del total de los perros y de cada uno de los grupos en las variables número de ensayos necesarios para llegar al criterio de aprendizaje en las fases de adquisición y reversión en la tarea social comparada a la no social, con la prueba de Wilcoxon. Se analizó la presencia de posibles asociaciones entre las distintas variables de cada tarea así como entre la tarea social y la no social en cada variable, utilizando la correlación de Spearman. Asimismo, se evaluó el efecto del orden de las tareas utilizando la prueba U de Mann Whitney. Dada la relevancia que la edad mostró tener en el aprendizaje de reversión, se realizó un análisis de correlación utilizando la prueba de Spearman para ver posibles asociaciones entre las distintas variables de ambas tareas y la edad. Se utilizó la probabilidad de superioridad (PSest) como índice del tamaño del efecto para comparaciones de dos grupos con medidas no paramétricas (Mann-Whitney). Esta es definida como la probabilidad de que un puntaje elegido aleatoriamente de una población X sea superior a un puntaje elegido al azar de otra población Y (Caycho et al., 2016). Para la prueba de Wilcoxon se utilizó el coeficiente r basado en el z-factor como indicador del tamaño del efecto (D’Angelo, 2021). Los datos fueron analizados con Infostat (Di Rienzo et al. 2020) y IBM SPSS v23.
Consideraciones éticas
El presente protocolo fue aprobado por el Comité Institucional de Cuidado y Uso de Animales de Laboratorio (CICUAL) del Instituto de Investigaciones Biológicas y Tecnológicas (IIByT, CONICET-UNC) y cumple con la actual ley Argentina de protección animal (Ley 14.346). Todos los dueños y guías de los perros expresaron su consentimiento para la participación de los mismos en el estudio. Los autores declaran que no poseen ningún conflicto de interés.
Resultados
En el siguiente apartado se presentan los resultados obtenidos, dividiéndolos en función de las comparaciones realizadas. En primer lugar se analizan posibles diferencias entre los grupos en las distintas variables medidas en cada una de las tareas. En el segundo apartado se realizan las comparaciones del desempeño de los perros en cada una de las variables en la tarea no social versus la social. Para ello se realizó primero un análisis considerando las respuestas de todos los perros en su conjunto y luego otro análisis de las respuestas de cada uno de los grupos por separado. En tercer lugar se presentan las correlaciones de las respuestas de los perros en las distintas variables tanto dentro como entre tareas. Por último, se muestra el análisis del efecto del orden de las tareas así como las correlaciones del desempeño en las tareas con la edad de los perros.
Comparación entre los grupos PD y PF
La Figura 2 muestra la mediana (q1 – q3) del número de ensayos hasta criterio en las distintas fases tanto en la tarea social como en la no social en función de los grupos PD y PF. Respecto a la comparación entre los grupos, no hubo diferencias significativas en la tarea no social, ni en el número de ensayos requeridos para alcanzar el criterio de adquisición (U = 105.5 p = 0.575 psest = 0.36) ni en el de reversión (U = 101, p = 0.473 psest = 0.3.4). De modo similar, en la tarea social, no se observaron diferencias significativas entre los grupos en el número de ensayos requeridos para alcanzar el criterio de adquisición (U = 108, p = 0.583 psest = 0.37), ni de reversión (U = 85.5, p = 0.172 psest = 0.29) (ver Figura 2).
Por otro lado, en la tarea no social 1 de 17 perros del grupo PD y 1 de 14 perros del grupo PF mostraron al menos una no elección en la fase de reversión. En la tarea social, el número fue de 3 perros en PD (N 15) y 6 en PF (N 14). En ninguno de los casos la comparación de las proporciones entre grupos fue significativa (ps > 0.245).
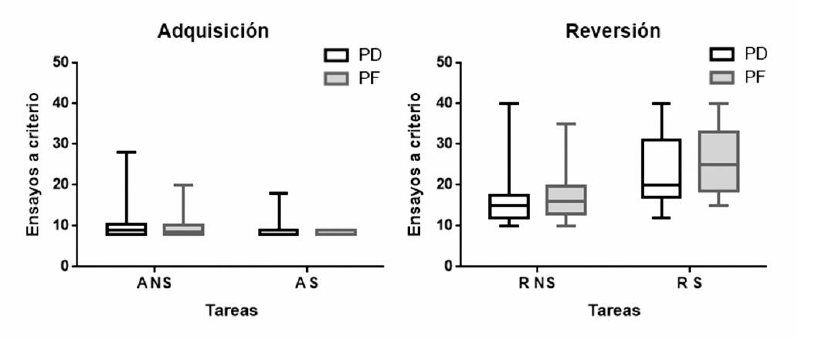
Figura 2
Mediana (q1 – q3) del número de ensayos hasta criterio en las fases de adquisición (A) y reversión (R), en las tareas no social (NS) y social (S) de los grupos PD (perros de detección) y PF (perros de familia)
Comparación entre las tareas social y no social
Tomando el conjunto total de los perros, se observa una diferencia significativa entre las tareas en el número de ensayos para llegar a criterio tanto en la fase de adquisición (Z = -2.33, p = 0.020 r = 0.53) como en la de reversión (Z = -3.13, p = 0.002 r = 0.71). De este modo, en la tarea social, los perros mostraron un mejor desempeño que en la tarea no social durante la adquisición pero un peor desempeño durante la fase de reversión.
Analizando cada grupo por separado, se encuentran diferencias significativas en el número de ensayos para llegar al criterio en la fase de reversión tanto en el grupo PD (Z = -1.98, p = 0.047 r = 0.51), como en el PF (Z = -2.41, p = 0.016 r = 0.64). De modo consistente con el total de la muestra, ambos grupos necesitaron menos ensayos para llegar al criterio en la tarea no social durante la reversión. Sin embargo, no hubo diferencias significativas en la fase de adquisición (ps > 0.091).
Respecto a las no elecciones, tomando el conjunto de la muestra, se encuentra que la proporción de perros que realizaron al menos una no elección es significativamente mayor en la tarea social que en la no social (p = 0.019). Sin embargo, al analizar a cada grupo por separado no se observan diferencias significativas (ps > 0.05) entre las tareas en ninguno de los dos grupos.
Correlaciones entre e intra tareas
No se hallaron correlaciones significativas en ninguna de las variables entre las tareas (ps > 0.34) ni entre las diferentes variables dentro de cada tarea (ps > 0.15).
Análisis de las variables orden de las tareas y edad
No se hallaron efectos del orden de comienzo de las tareas (ps > 0.21).
Por otro lado, se observó una correlación positiva significativa entre la edad de los perros y el número de respuestas necesarias para llegar al criterio de reversión tanto en la tarea no social (Rho = 0.557, p = 0.001), como en la social (Rho = 0.551, p = 0.001). No se encontraron otras asociaciones significativas (ps > 0.10).
Discusión
El primer objetivo del presente estudio fue determinar si el entrenamiento en tareas de detección de olores (narcóticos, explosivos y personas) podía modular el aprendizaje de reversión. Un segundo objetivo pretendió evaluar si el desempeño de los perros era diferente en una tarea en la que solo había claves no sociales comparado a otra en la que se agregaba a dichas claves una señal social. Para ello los perros recibieron una fase de adquisición de una clave de lugar asociada a la comida (tarea no social), o la suma de la clave de lugar más la posición de una persona desconocida ubicada detrás de la posición correcta (tarea social). En una segunda fase, de reversión, se invirtieron las contingencias de modo de que la clave previamente reforzada ya no lo era, y la comida estaba en el lugar opuesto. Los resultados mostraron que no hubo diferencias entre los grupos de perros entrenados y los de familia en la cantidad de ensayos hasta alcanzar el criterio de aprendizaje tanto en la fase de adquisición como de reversión. Más aún, tampoco se observaron diferencias entre ambos grupos comparando la proporción de perros de cada grupo que presentaron al menos una no elección durante la fase de reversión. Estos resultados fueron observados en ambas tareas, no social y social.
La ausencia de diferencias entre grupos resulta llamativa ya que según los antecedentes revisados (Lazarowski et al., 2020; Tiira et al., 2020) diversos factores vinculados al entrenamiento de los perros de detección de olores podrían haber modulado el aprendizaje de reversión. Sin embargo, considerando la escasez y ambigüedad de los hallazgos previos, no estaba claro en qué dirección se producirían las diferencias de grupos. En este sentido, algunos factores predecirían un peor desempeño de los sujetos entrenados. Por ejemplo, los perros de detección deben trabajar en ambientes extremos y por períodos largos de tiempo, siendo escasamente reforzados (Lazarowski et al., 2020). Por esta razón, algunos autores han sugerido incluir esquemas de reforzamiento frecuentes pero variables durante los entrenamientos (Troisi et al., 2019). Estos esquemas podrían llevar a una mayor resistencia a la disrupción y a la extinción (Hall, 2017). En este caso, esta mayor resistencia podría traducirse en un mayor número de errores perseverativos y por lo tanto en un menor aprendizaje en la fase de reversión comparado a los perros de familia. Esto iría en línea con la mayor persistencia observada en los perros de detección en la resolución de un problema irresoluble (Lazarowski et al., 2020).
Por otro lado, también sería posible esperar un mejor desempeño de los perros de detección. Teniendo en cuenta que los perros de trabajo deben hacer tareas largas, Tiira et al. (2020) plantearon que estos deben tener bajos niveles de activación (arousal) y alto autocontrol. Según esta afirmación, los perros de detección deberían inhibir mejor el acercarse al estímulo previamente reforzado, superando en su desempeño a los perros de familia. Esto cobra apoyo en un estudio que mostró un mejor rendimiento en una tarea de reversión (tarea A no B) en perros entrenados para rescate acuático (Barrera et al., 2018). Asimismo, algunos autores postulan que los perros de detección representan un tipo único de perros de trabajo, ya que deben poder desempeñarse en coordinación con su guía, pero a su vez mostrando un alto grado de autonomía. Es decir que deben exhibir diferentes grados de sensibilidad social e independencia según el contexto (Lazarowski et al., 2020). En apoyo de esta hipótesis, contrariamente a lo observado en perros de familia (Szetei et al., 2003), cachorros entrenados para ser perros de detección siguieron más la clave de olor cuando debían elegir entre esta clave y un gesto comunicativo humano engañoso. Más aún, la falta de sensibilidad a la clave humana fue predictora del éxito como perros de detección (Lazarowski et al., 2019). Asimismo, los perros que se desempeñaron más exitosamente en la detección y el rescate de personas durante avalanchas eran aquellos que dependían menos de su guía y pasaban más tiempo explorando el ambiente (Diverio et al., 2017). A diferencia de estos, los perros de familia suelen ser menos estimulados a resolver problemas de modo independiente (Udell, 2015). Esta mayor autonomía de los perros de detección respecto a los estímulos sociales humanos podría haber favorecido el comportamiento de inhibir el acercamiento o incluso de ignorar dichos estímulos. De este modo, los perros de detección podrían haber mostrado un mejor desempeño en la reversión de la tarea social. Sin embargo, es importante considerar que la tarea aquí llamada social, no era exclusivamente social ya que los perros contaban además con la información provista por la clave de lugar. Esto pudo haber interferido en la forma en la cual estos procesaron la información provista por la persona. En estudios futuros sería interesante evaluar a los perros de detección y los de familia en un aprendizaje de reversión en el cual haya exclusivamente claves sociales.
A pesar del conjunto de estas consideraciones, en el presente estudio no se evidenciaron diferencias entre los perros de detección y los de familia. Una posible explicación es que la ausencia de diferencias entre grupos podría deberse a que los perros entrenados en detección de olores no suelen enfrentar situaciones similares a las aquí evaluadas. Si bien deben persistir durante un largo tiempo en su búsqueda, aún sin refuerzo, rara vez son expuestos a cambios en las contingencias de refuerzo, ya que los olores objetivos son siempre los mismos y su detección resulta siempre reforzada. En este sentido, sería interesante en estudios futuros evaluar otros componentes tanto del control inhibitorio y la flexibilidad como de la selectividad de la atención en esta población de perros. Asimismo, sería interesante poder equiparar las condiciones de vida de ambos grupos y evaluar perros de detección que vivan en casas de familia, en lugar de caniles donde tienen menos interacción con las personas.
Respecto al segundo objetivo, se observó que el desempeño del total de los perros fue mejor en la adquisición de la tarea social que de la no social, pero este patrón se invirtió en la fase de reversión. Esta diferencia respecto a la reversión fue también encontrada al analizar a cada grupo por separado. De este modo, si bien la clave compuesta (lugar y posición de la persona) pudo facilitar la adquisición de la discriminación al incrementar las discrepancias entre las dos opciones (Blough et al., 2004; George et al., 2018), el efecto opuesto podría haberse observado en la reversión. Esta discrepancia podría deberse al hecho de que los perros, tal como se mencionó en la introducción, prestan elevada atención a los estímulos sociales humanos (Gácsi et al., 2009), estos les resultan reforzantes (Bolló et al., 2021; Cook et al., 2016) y son utilizados como claves para guiar su comportamiento de modo persistente, aun en situaciones desventajosas (e.g., Kundey et al., 2010; Szetei et al., 2003). Teniendo esto en cuenta, la presencia de la clave humana durante la adquisición pudo haber captado una mayor atención de los perros y haber incrementado el valor apetitivo de esa opción, sumando al valor reforzante de la comida el valor de la persona como refuerzo social. Esto pudo facilitar la discriminación inicial que realizaron los perros entre ambas claves en la tarea social. Al mismo tiempo, esto pudo incrementar la dificultad durante la reversión, en la cual los perros debían inhibir el acercamiento al humano y aprender a elegir la clave no social. Este déficit se observó durante la reversión tanto en un mayor número de ensayos necesarios para alcanzar el criterio de aprendizaje como en una mayor proporción de perros que realizaron no elecciones en la tarea social que en la no social. Posiblemente esto refleje una mayor dificultad para seguir la clave que era exclusivamente no social. Estos hallazgos son acordes a los reportados en la literatura utilizando la tarea A no B (e.g., Gergely et al., 2016; Topál et al., 2009) pero contrastan con los de Fagnani et al. (2020). Estos últimos autores mostraron un efecto facilitador de la presencia de las personas en el aprendizaje de reversión. Sin embargo, en el estudio de Fagnani et al. (2020) las dos opciones utilizadas en la discriminación eran compuestas, ya que había dos personas sentadas detrás de los recipientes en dónde se colocaba la comida y para encontrarla los perros debían seguir la clave de lugar. Teniendo esto en cuenta, ambas opciones podían captar por igual la atención de los perros y la diferencia en su valor apetitivo radicaba únicamente en la presencia o no de la comida. Es posible que los perros se guiaran únicamente por la clave de lugar, evitando hacer un proceso complejo de discriminación entre las personas desconocidas. A su vez, las mismas podrían haber funcionado como un contexto positivo que favoreciera la resolución del problema.
Por otro lado, no se hallaron asociaciones significativas en el desempeño de los perros entre las dos tareas. Estos resultados son consistentes con Fagnani et al. (2016) quienes no hallaron correlaciones significativas entre una versión social y una no social de una prueba de reversión. Una posible explicación es que ambas tareas pudieron haber involucrado diferentes demandas de aprendizaje para los perros.
Por último, se observó una correlación entre la edad de los sujetos y el desempeño en la tarea de reversión tanto social como no social. Esto indica que a mayor edad de los perros se requirieron más ensayos para llegar al criterio del aprendizaje de reversión. Este hallazgo es consistente con numerosas evidencias previas (e.g., Milgram et al., 1994; Mongillo et al., 2013; Piotti et al., 2018; Tapp, 2003; Van Bourg et al., 2021) dando cuenta de una perdida de la flexibilidad de los perros durante el envejecimiento y resaltando la utilidad de esta tarea de reversión para medir las funciones ejecutivas en esta especie.
Una limitación importante a tener en cuenta en el presente estudio, así como en la mayoría de las investigaciones en las que se evalúan perros de trabajo, refiere a las condiciones de vivienda específicas de estos perros. A diferencia de los de familia, los perros de trabajo suelen estar alojados en caniles. El grado de interacción con la gente es variable de acuerdo con la frecuencia de los entrenamientos y de sus actividades, el número de guías y cuidadores que están en contacto con ellos así como la cantidad de personas a las que son expuestos durante su trabajo. Todos estos factores vuelven a la muestra necesariamente heterogénea en lo que concierne a sus experiencias de socialización.
En conclusión, no se observaron diferencias entre perros entrenados en detección de olores y perros de familia no entrenados. A su vez, los perros tuvieron un mejor desempeño en la adquisición de la tarea social que de la no social, aunque se observó un patrón opuesto respecto a la fase de reversión. Se requieren más investigaciones para conocer las características específicas de estos perros de trabajo así como del modo en que puede repercutir la presencia de estímulos sociales durante los aprendizajes.
Conflicts of Interest:
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Agradecimientos
Queremos expresar nuestra enorme gratitud al personal de la Base Naval Puerto Belgrano, Argentina, al grupo K9 de los bomberos voluntarios de la Ciudad de Punta Alta, especialmente al entrenador Lucas Maciel y al Departamento de Búsqueda y Rescate con Canes de la Federación de Bomberos Voluntarios de la Provincia de Córdoba, así como a todos los dueños que gentilmente colaboraron participando en el estudio. El mismo fue financiado por la Agencia Nacional de Promoción de la Investigación, el Desarrollo Tecnológico y la Innovación (PICT 2014 Nº 0883) y CONICET (PIP 2013 Nº 11220130100182).
Referencias
Anderson, B. A., Laurent, P. A., y Yantis, S. (2013). Reward predictions bias attentional selection. Frontiers in Human Neuroscience, 7, 262. https://doi.org/10.3389/fnhum.2013.00262
Awh, E., Belopolsky, A. V., y Theeuwes, J. (2012). Top-down versus bottom-up attentional control: a failed theoretical dichotomy. Trends in Cognitive Sciences, 16, 437–443. https://doi.org/10.1016/j.tics.2012.06.010
Barrera, G., Alterisio, A., Scandurra, A., Bentosela, M., y D’Aniello, B. (2018). Training improves inhibitory control in water rescue dogs. Animal Cognition, 22(1), 127–131. https://doi.org/10.1007/s10071-018-1224-9
Barrera, G., Mustaca, A. y Bentosela, M. (2011). Gaze at the human face in shelter and pet dogs. Animal Cognition, 14, 727-734. https://doi.org/10.1007/s10071-011-0407-4
Barrera, G., Jakovcevic, A., Elgier, A. M., Mustaca, A., y Bentosela, M. (2010). Responses of shelter and pet dogs to an unknown human. Journal of Veterinary Behavior, 5(6), 339-344. https://doi.org/10.1016/j.jveb.2010.08.012
Blough, D. S. (2004). Reaction time signatures of discriminative processes: Differential effects of stimulus similarity and incentive. Animal Learning & Behavior, 32(2), 157-172. https://doi.org/10.3758/bf03196017
Bolló, H., Kiss, O., Kis, A., y Topál, J. (2021). The implicit reward value of the owner's face for dogs. iScience, 24(8), 102763. https://doi.org/10.1016/j.isci.2021.102763
Boutet I., Ryan M., Kulaga V., McShane C., Christie L., Freedman M. y Milgram N. W. (2005). Age-associated cognitive deficits in humans and dogs: A comparative neuropsychological approach. Progress in Neuro Psychopharmacology and Biological Psychiatry, 29 (3), 433-441. https://doi.org/10.1016/j.pnpbp.2004.12.015
Cavalli, C., Carballo, F., Dzik, V., Underwood, S. y Bentosela, M. (2018). Are Animal Assisted Activities dogs different from pet dogs? A comparison of their sociocognitive abilities. Journal of Veterinary Behavior. https://doi.org/10.1016/j.jveb.2017.12.001
Caycho T., Ventura-León J., Castillo-Blanco R. (2016). Magnitud del efecto para la diferencia de dos grupos en ciencias de la salud. Anales del Sistema Sanitario de Navarra, 39 (3), 459. https://doi.org/10.23938/ASSN.0242
Chase, H. W., Swainson, R., Durham, L., Benham, L., y Cools, R. (2011). Feedback-related negativity codes prediction error but not behavioral adjustment during probabilistic reversal learning. Journal of Cognitive Neuroscience, 23, 936–946. https://doi.org/10.1162/jocn.2010.21456
Cook, P. F., Prichard, A., Spivak, M., y Berns, G. S. (2016). Awake canine fMRI predicts dogs’ preference for praise vs food. Social Cognitive and Affective Neuroscience, 11(12), 1853-1862. https://doi.org/10.1101/062703
Costa, V. D., Tran, V. L., Turchi, J. y Averbeck, B. B. (2015) Reversal learning and dopamine: a bayesian perspective. Journal of Neuroscience, 35, 2407-2416. https://doi.org/10.1523/JNEUROSCI.1989-14.2015
D’Angelo, L. (2021). Tamaño de efecto, potencia de la prueba, factor de Bayes y meta-análisis en el marco de la crisis de reproducibilidad de la ciencia. El caso de la diferencia de medias -con muestras independientes- (primera parte). Cuadernos del cimbage, 1 (23), 47-82. Recuperado a partir de https://ojs.econ.uba.ar/index.php/CIMBAGE/article/view/2055
Di Rienzo, J. A., Casanoves, F., Balzarini, M. G., Gonzalez, L., Tablada, M. y Robledo C.W. (2020). InfoStat versión 2020. Centro de Transferencia InfoStat, FCA, Universidad Nacional de Córdoba, Argentina. URL http://www.infostat.com.ar
Diverio, S., Menchetti, L., Riggio, G., Azzari, C., Iaboni, M., Zasso, R. Di Mari, W., y Santoro, M. M. (2017). Dogs’ coping styles and dog-handler relationships influence avalanche search team performance. Applied Animal Behaviour Science, 191, 67–77. https://doi.org/10.1016/j.applanim.2017.02.005
Elgier, A., Jakovcevic, A., Mustaca, A. y Bentosela, M. (2012). Pointing following in dogs: are simple or complex cognitive mechanisms involved? Animal Cognition. 15 (6), 1111-1119. https://doi.org/10.1007/s10071-012-0534-6
Evans, R. I., Herbold, J. R., Bradshaw, B. S., y Moore, G. E. (2007). Causes for discharge of military working dogs from service: 268 cases (2000–2004). Journal of the American Veterinary Medical Association, 231(8), 1215–1220. https://doi.org/10.2460/javma.231.8.1215
Fagnani, J., Barrera, G., Carballo, F., y Bentosela, M. (2016). Is previous experience important for inhibitory control? A comparison between shelter and pet dogs in A-not-B and cylinder tasks. Animal Cognition, 19(6), 1165–1172. https://doi.org/10.1007/s10071-016-1024-z
Fagnani, J., Bentosela, M., y Barrera, G. (2020a). Assessing dogs’ performance in a social and non-social reversal learning task. Journal of Dog Behavior, 6(2). https://doi.org/10.4454/db.v6i2.118
Gácsi, M., McGreevy, P., Kara, E., y Miklósi, Á. (2009). Effects of selection for cooperation and attention in dogs. Behavioral and Brain Functions, 5(1), 31. https://doi.org/10.1186/1744-9081-5-31
George, D. N. (2018). Stimulus similarity affects patterning discrimination learning. Journal of Experimental Psychology: Animal Learning and Cognition, 44(2), 128-148. https://doi.org/10.1037/xan0000164
Gergely, A., Compton, A. B., Newberry, R. C., y Miklósi, Á. (2016). Social Interaction with an “Unidentified Moving Object” Elicits A-Not-B Error in Domestic Dogs. PLOS ONE, 11(4), e0151600. https://doi.org/10.1371/journal.pone.0151600
Hall, N. J. (2017). Persistence and resistance to extinction in the domestic dog: Basic research and applications to canine training. Behavioural Processes, 141(1), 67-74. http://dx.doi.org/10.1016/j.beproc.2017.04.001
IBM Corp. (2015). IBM SPSS Statistics for Windows, Version 23.0. IBM Corp.
Izquierdo, A., Brigman, J. L., Radke, A. K., Rudebeck, P. H., y Holmes, A. (2017). The neural basis of reversal learning: An updated perspective. Neuroscience, 345, 12-26. https://doi.org/10.1016/j.neuroscience.2016.03.021
Izquierdo, A., y Jentsch, J. D. (2012). Reversal learning as a measure of impulsive and compulsive behavior in addictions. Psychopharmacology, 219(2), 607–620. https://doi.org/10.1007/s00213-011-2579-7
Kelly, D. M., Adolphe, J. L., Vernouillet, A., McCausland, J. A., Rankovic, A., y Verbrugghe, A. (2019). Motoric self-regulation by sled dogs and pet dogs and the acute effect of carbohydrate source in sled dogs. Animal Cognition, 22(6), 931–946. https://doi.org/10.1007/s10071-019-01285-y
Kundey, S. M. A., De Los Reyes, A., Arbuthnot, J., Allen, R., Coshun, A., Molina, S., y Royer, E. (2010). Domesticated dogs’ (Canis familiaris) response to dishonest human points. International Journal of Comparative Psychology, 23(2), 201–215. https://doi.org/10.46867/ijcp.2010.23.02.02
Lazarowski, L., Strassberg, L. R., Waggoner, L. P., y Katz, J. S. (2019). Persistence and human-directed behavior in detection dogs: Ontogenetic development and relationships to working dog success. Applied Animal Behaviour Science, 220, 104860. https://doi.org/10.1016/j.applanim.2019.104860
Lazarowski, L., Thompkins, A., Krichbaum, S., Waggoner, L. P., Deshpande, G., y Katz, J. S. (2020). Comparing pet and detection dogs (Canis familiaris) on two aspects of social cognition. Learning & Behavior, 48(4), 432–443. https://doi.org/10.3758/s13420-020-00431-8
MacLean, E. L., y Hare, B. (2018). Enhanced Selection of Assistance and Explosive Detection Dogs Using Cognitive Measures. Frontiers in Veterinary Science, 5, 236. https://doi.org/10.3389/fvets.2018.00236
Milgram, N. W., Head, E., Weiner, E., y Thomas, E. (1994). Cognitive functions and aging in the dog: Acquisition of nonspatial visual tasks. Behavioral Neuroscience, 108(1), 57–68. https://doi.org/10.1037/0735-7044.108.1.57
Mongillo, P., Araujo, J. A., Pitteri, E., Carnier, P., Adamelli, S., Regolin, L., y Marinelli, L. (2013). Spatial reversal learning is impaired by age in pet dogs. AGE, 35(6), 2273–2282. https://doi.org/10.1007/s11357-013-9524-0
Mongillo, P., Scandurra, A., D’Aniello, B., y Marinelli, L. (2017). Effect of sex and gonadectomy on dogs’ spatial performance. Applied Animal Behaviour Science, 191, 84–89. https://doi.org/10.1016/j.applanim.2017.01.017
Oemisch, M., Watson, M. R., Womelsdorf, T. y Schubö, A. (2017) Changes of Attention during Value-Based Reversal Learning Are Tracked by N2pc and Feedback-Related Negativity. Frontiers in Human Neuroscience, 11, 540. https://doi.org/10.3389/fnhum.2017.00540
Pavlov, I. P. (1927). Conditioned reflexes. Oxford University Press.
Pearce, J. y Hall, G. (1980). A model for Pavlovian learning: variation in the effectiveness of conditioned but not unconditioned stimuli. Psychological Review, 87, 532-552. https://doi.org/10.1037/0033-295x.87.6.532
Piotti, P., Szabó, D., Bognár, Z., Egerer, A., Hulsbosch, P., Carson, R. S., y Kubinyi, E. (2018). Effect of age on discrimination learning, reversal learning, and cognitive bias in family dogs. Learning & Behavior, 46(4), 537–553. https://doi.org/10.3758/s13420-018-0357-7
Posner, M. y Petersen, S. E. (1990). The attention system of the human brain. Annual Review of Neuroscience, 13, 25-42. https://doi.org/10.1146/annurev.ne.13.030190.000325
Prato-Previde, E., Marshall-Pescini, S., y Valsecchi, P. (2007). Is your choice my choice? The owners’ effect on pet dogs’ (Canis lupus familiaris) performance in a food choice task. Animal Cognition, 11(1), 167–174. https://doi.org/10.1007/s10071-007-0102-7
Range, F., Heucke, S. L., Gruber, C., Konz, A., Huber, L., y Virányi, Z. (2009). The effect of ostensive cues on dogs’ performance in a manipulative social learning task. Applied Animal Behaviour Science, 120(3-4), 170-178. https://doi.org/10.1016/j.applanim.2009.05.012
Simon, A., Lazarowski, L., Singletary, M., Barrow, J., Van Arsdale, K., Angle, T., Waggoner, P., y Giles, K. (2020). A Review of the Types of Training Aids Used for Canine Detection Training. Frontiers in Veterinary Sciences, 7, 313. https://doi.org/10.3389/fvets.2020.00313
Szetei, V., Miklósi, Á., Topál, J., y Csányi, V. (2003). When dogs seem to lose their nose: an investigation on the use of visual and olfactory cues in communicative context between dog and owner. Applied Animal Behaviour Science, 83(2), 141–152. https://doi.org/10.1016/s0168-1591(03)00114-x
Tapp, P. D. (2003). Size and Reversal Learning in the Beagle Dog as a Measure of Executive Function and Inhibitory Control in Aging. Learning & Memory, 10(1), 64–73. https://doi.org/10.1101/lm.54403
Thorndike, E. L. (1898). Experiments with Dogs. In E. L. Thorndike (Ed.), Animal intelligence: An experimental study of the associative processes in animals (pp. 31–34). Columbia University Press. https://doi.org/10.1037/10780-004
Tiira, K., Tikkanen, A., y Vainio, O. (2020). Inhibitory control – Important trait for explosive detection performance in police dogs? Applied Animal Behaviour Science, 224, 104942. https://doi.org/10.1016/j.applanim.2020.104942
Topal, J., Gergely, G., Erdohegyi, A., Csibra, G., y Miklosi, A. (2009). Differential Sensitivity to Human Communication in Dogs, Wolves, and Human Infants. Science, 325(5945), 1269–1272. https://doi.org/10.1126/science.1176960
Troisi, C. A., Mills, D. S., Wilkinson, A., y Zulch, H. E. (2019). Behavioral and Cognitive Factors That Affect the Success of Scent Detection Dogs. Comparative Cognition & Behavior Reviews, 14, 51–76. https://doi.org/10.3819/ccbr.2019.140007
Udell, M. A. R. (2015). When dogs look back: inhibition of independent problem-solving behaviour in domestic dogs (Canis lupus familiaris) compared with wolves (Canis lupus). Biology Letters, 11(9), 20150489. https://doi.org/10.1098/rsbl.2015.0489
Van Bourg, J., Gunter, L. M., y Wynne, C. D. L. (2021). A rapid serial reversal learning assessment for age-related cognitive deficits in pet dogs. Behavioural Processes, 186, 104375. https://doi.org/10.1016/j.beproc.2021.104375
Voytko, M. L. (1999). Impairments in acquisition and reversals of two-choice discriminations by aged rhesus monkeys. Neurobiology of Aging, 20(6), 617–627. https://doi.org/10.1016/s0197-4580(99)00097-4
Wallis, L., Range, F., Müller, C. y Virányi, Z. (2011). Reversal learning in a social communication task: is there an effect of cue? Journal of Veterinary Behavior. http://dx.doi.org/10.1016/j.jveb.2010.09.0017
Wilsson, E., y Sundgren, P. E. (1997). The use of a behaviour test for the selection of dogs for service and breeding, I: Method of testing and evaluating test results in the adult dog, demands on different kinds of service dogs, sex and breed differences. Applied Animal Behaviour Science, 53(4), 279–295. https://doi.org/10.1016/s0168-1591(96)01174-4
Wobber, V., y Hare, B. (2009). Testing the social dog hypothesis: Are dogs also more skilled than chimpanzees in non-communicative social tasks? Behavioural Processes, 81(3), 423–428. https://doi.org/10.1016/j.beproc.2009.04.003
Wright, H. F., Mills, D. S., y Pollux, P. M. (2011). Development and validation of a psychometric tool for assessing impulsivity in the domestic dog (Canis familiaris). International Journal of Comparative Psychology, 24 (2), 210–225. https://doi.org/10.46867/ijcp.2011.24.02.03
Notas de autor
Correspondence about this article should be addressed Mariana Bentosela: marianabentosela@gmail.com