ABSTRACT: This research aimed to explore the reproductive characteristics of three species of Psidium (P. guajava L., P. cattleyanum Sabine, and P. guineense Sw.) and estimate a probable reproduction strategy based on the pollen:ovule (P:O) methodology. The number of pollen grains per floral bud (NGPB), number of pollen grains per anther (NGPA), number of anthers per flower bud (NAB), number of ovules per flower bud (NOB), and the P:O ratio of each species were estimated. All species had a P:O ratio over 2,000 and were classified as xenogamous. P.guajava presented the highest values for all characteristics evaluated, with the NGPB at 3,777,519, the NOB at 584.50 and a P:O ratio of 6,462.82. Similarly, P. cattleyanum had a P:O ratio of 5,649.89 (NGPB 762,736 and NOB 135). However, P. guineense was considered facultative xenogamous, with P:O of 2,085.75, the NGPB at 741,484 and the NOB at 355.50. Thus, it was concluded that the studied species have a preference for allogamy and require many pollen grains to fertilize each ovule, demonstrating that the transfer of pollen to the stigma is not very specialized.
Keywords: allogamyallogamy,autogamyautogamy,Psidium cattleyanumPsidium cattleyanum,Psidium guajavaPsidium guajava,Psidium guineensePsidium guineense.
GENETICS AND PLANT BREEDING
Analysis of the reproduction mode in Psidium spp. using the pollen:ovule ratio
Editora da Universidade Estadual de Maringá - EDUEM
Received: 09 August 2018
Accepted: 24 December 2018
The genus Psidium contains approximately 150 species, including P. guajava (guava) (diploid with 22 chromosomes in somatic cells, 2n = 2x = 22), P. guineense (araçazeiro) (tetraploid with 44 chromosomes in somatic cells, 2n = 4x = 44), and P. cattleyanum [ploidy, from triploid (2n = 3x = 33) to octaploid with chromosome numbers varying from 33 to 82 in somatic cells (2n = 8x = 82)] (Pereira, 1995; Souza, Resende, Lima, Santos, & Chalfun, 2014; Souza, Resende, Lima, Martins, & Techio, 2015). The annual production of guava in Brazil is 414,960 tons, and the average yield is 24,240 kg ha-1 (SEBRAE, 2015; IBGE, 2016). Brazil is the third largest producer of guava and other fruit trees worldwide behind China and India (SEBRAE, 2015).
Guava production is being jeopardized by a complex disease known as guava decline,which is caused by the synergistic association of the nematode Meloidogyne enterolobii with the fungus Fusarium solani. Guava decline causes the progressive rotting of the root system, burning of the edges, leaf yellowing and fall, and ultimately plant death (Gomes, Souza, Mussi-Dias, Silveira, & Dolinski, 2011). This disease has been reported in several Brazilian states (Maranhão, de Moura, & Pedrosa, 2001; Silva, Pereira, Araújo, & Carneiro, 2008; Biazatti et al., 2016) and has a high economic impact. To date, the control of this disease has not been effective (Pereira, de Souza, Souza, Dolinski, & Santos, 2009; Almeida, Wicker, Santos, & Martins, 2012).
The use of resistant cultivars is the best alternative for disease control. However, P. guajava genotypes resistant to guava decline are not available to date, although disease resistance has been reported in accessions of P. cattleyanum and P. guineense (Miranda, Souza, & Viana, 2011; Costa, Santos, & Castro, 2016). Moreover, the use of rootstocks for guava is unfeasible (Robaina, Campos, Marinho, Souza, & Bremenkanp, 2015). Therefore, for guava crops, plant breeding programs are important for developing cultivars or rootstocks that are genetically resistant and compatible for grafting (Carneiro, Cirotto, Quintanilha, Silva, & Carneiro, 2007; Almeida, Martins, & Santos, 2009; Gomes et al., 2011; Miranda et al., 2011; Miranda, de Souza, Gomes, Ferreira, & Almeida, 2012).
Allard (1960) reported that the effective execution of an improvement program depends on the knowledge of the reproductive mode of the species because the choice of the breeding method is based on this characteristic, i.e., whether the species is autogamous or allogamous. In guava, this information is still controversial because different studies have reported that guava is allogamous and has a high rate of autogamy or vice versa (Dasarathy, 1951; Soubihe Sobrinho & Gurgel, 1962; Alves & Freitas, 2007). This information is still scarce for P. guineense. Therefore, the reproduction mode of some species in the Psidium genus needs to be better understood.
Several methodologies can be used to determine the reproductive mode. Cruden (1977) reported that determining the pollen:ovule (P:O) ratio is a good method to predict the type of reproduction because it is fast and costs little compared with molecular marker or cross-breeding techniques (Cruden, 2000). Many studies have used the P:O ratio to determine the probable pollination strategy (Galloni, Podda, Vivarelli, & Cristofolini, 2007) or pollination efficiency (Chouteau, Barabé, & Gibernau, 2006). Additionally, this method was used to determine the mechanisms of pollination in species with unknown reproduction modes (López, Rodriguez-Riaño, Ortega-Olivencia, Devesa, & Ruiz, 1999).
The P:O ratio reflects the probability of pollen grains reaching the stigma, resulting in the maximum production of seeds (Cruden, 1977). A lower P:O ratio indicates a more efficient transfer of pollen grains. Therefore, plants considered cleistogamic should have a lower P:O ratio. Moreover, the P:O ratio of autogamous plants should be lower than that of allogamous plants. The P:O ratio is directly correlated with the pollination and reproduction mode of the species (Cruden, 1977; Lenzi & Orth, 2004).
The objective of this study was to study the reproductive characteristics of three species of Psidium (P. guajava L., P. cattleyanum Sabine, and P. guineense Sw.), determine how these characteristics allow estimation of the P:O ratio, and identify the likely reproductive strategy based on this ratio.
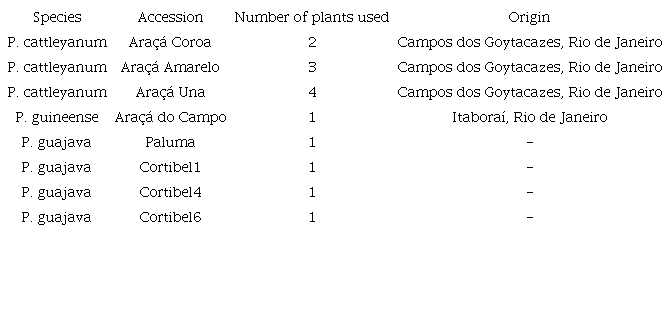
The accessions used in this study belonged to the germplasm collection of the Genetic Improvement Program of Guava Trees of the State University of Northern Rio de Janeiro Darcy Ribeiro (Universidade Estadual do Norte Fluminense Darcy Ribeiro -UENF). The plant collection was located in the Antônio Sarlo Agricultural School in Campos dos Goytacazes, Rio de Janeiro State, Brazil.
For this study, flower buds (FB) of four araçá accessions were collected: Araçá Amarelo (P. cattleyanum), Araçá Coroa (P. cattleyanum), Araçá Una (P. cattleyanum), along with Araçá do Campo (P. guineense), and four guava varieties: Cortibel 1, Cortibel 4, Cortibel 6, and Paluma (Table 1).
The guava trees were propagated by cutting, whereas P. cattleyanum varieties were propagated using vegetative techniques. The spacing was 5 m within each row and 7 m between rows, and cover crop treatment was applied as needed.
During flowering, four FBs per accession in preanthesis were randomly collected in 70% ethanol and kept at 4°C until slides preparation.
Accessions of Psidium spp. maintained at the Antônio Sarlo Agricultural School, Campos dos Goytacazes, Rio de Janeiro State, Brazil.
The NPGA was counted using the protocol of Dafni (1992) and Damasceno Junior et al. (2009). For this purpose, one anther per FB was macerated in a solution containing four drops of Triton-X, three drops of 0.5% methylene blue, and 900 µL of 70% ethanol. Then, the anther was placed in a centrifuge tube containing the same solution, and the solution was shaken manually for 1 min. A 10-µL aliquot of the total suspension (1,000 µL) was placed on a histological slide, the slide was covered with a cover slip, and the number of pollen grains was counted under an Olympus BX60 optical microscope.
Four slides were prepared per anther (with a 10-µL sample on each slide), and the total number of pollen grains was counted. This procedure was performed with five anthers per FB, and four FBs were used per accession.
The number of pollen grains present in one anther (macerated in 1,000 µL) was estimated considering the mean number of pollen grains in the four tissue slides. Subsequently, the mean number of pollen grains present in five anthers from the same FB and the number of pollen grains in one anther of each FB were estimated.
The NAB was obtained by counting the number of anthers per flower. NPGB was estimated by multiplying the NPGA by the NAB of each FB.
NOB was estimated by counting the number of ovules. For this purpose, the ovaries were cut, and the ovules were placed on a tissue slide with three drops of a lactophenol cotton-blue solution. Then, a coverslip was placed on the slide, and the ovules were counted under an Olympus BX60 optical microscope.
The values for each accession were estimated by determining the mean counts in the four FBs analyzed.
The P:O ratio was estimated for each accession and species by dividing the NPGB by the NOB. The reproduction mode was determined by dividing the number of pollen grains by the number of ovules according to Cruden (1977).
NGPA, NGPB, NAB, and NOB were subjected to analysis of variance using a statistical model based on a completely randomized design:
where: ( = the constant; Gi = the effect of the ith fixed genotype (i = 1,2 ..., 8); and (ijk = the experimental error associated with each observation, assuming an NID (0, (²).
The accessions were classified by comparing the means using Tukey’s test at a level of significance of 0.05. All statistical analyses were conducted using Statistical Analysis System [SAS] (1996).
The results of the analysis of variance, the estimates of the means and coefficient of variation for NPGA, NPGB, NAB, NOB, and the P:O ratio are shown in Table 2.
Analysis of variance for the number of pollen grains per flower bud (NPGB), number of pollen grains per anther (NPGA), number of anthers per flower bud (NAB), number of ovules per flower bud (NOB), and pollen:ovule ratio (P:O) in eight accessions of Psidium ssp.
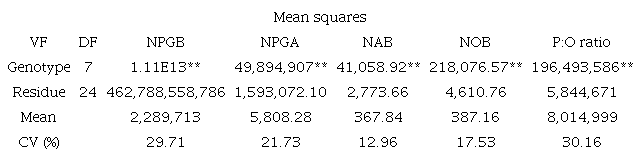
VF, variation factor; DF, degree of freedom; **different using the F-test at a level of significance of 0.01; CV(%), coefficient of variation.
All evaluated variables presented significant differences using the F-test at a level of significance of 1% (p < 0.01). Gomes (1990) analyzed the coefficients of variation (CV) and observed that the estimated values obtained in the field were considered moderate for the NAB (12.96%) and the NOB (17.53%) and high for the NPGB (29.71%), the NPGA (21.73%), and the P:O ratio (30.16%) using CVs of < 10%, 10-20%, 20-30%, and > 30% as low, moderate, high, and very high, respectively.
However, although the CVs were classified as moderate to high, these values considered the variability in three species of the genus Psidium. In addition, the CVs for the NPGB and P:O ratio were highest (29.71 and 30.16%, respectively) because these characteristics were estimated using two variables-NAB and NPGA for NPGB and for P:O ratio, NPGB and NOB-and these correlations may have increased the CV.
The mean values for NPGA, NPGB, NAB, and NOB were analyzed using Tukey's test at a level of significance of 0.05 are presented in Table 3.
Number of pollen grains per flower bud (NPGB), number of pollen grains per anther (NPGA), number of anthers per flower bud (NAB), and number of ovules per flower bud (NOB) in eight accesses of Psidium ssp.
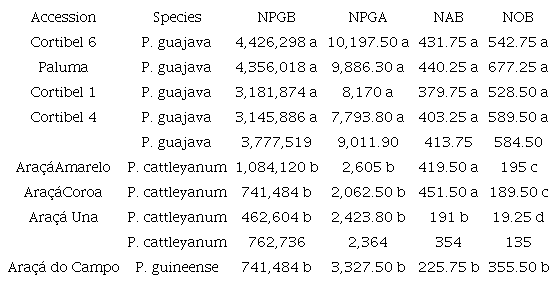
The means followed by the same letters in the column do not differ using Tukey’s test at a level of significance of 0.05.
The accessions with the highest mean NPGB were the same as those with the highest mean NPGA, and these values were higher in all P. guajava varieties than in all accessions of Araçá (both P. guineense and P. cattleyanum). The variety Cortibel 6 had the highest mean NPGB (4,426,298) and NPGA (10,197.50). Araçá Una presented the lowest mean NPGB (462,604), whereas Araçá Coroa had the lowest mean NPGA (2,062.50).
Araçá Una presented the lowest mean NAB (191), whereas Araçá Coroa had the highest mean NAB (451.50). In addition to Araçá Coroa, Araçá Amarelo and all P. cattleyanum varieties had the highest mean NAB, indicating that only Araçá do Campo and Araçá Una presented the lowest mean NAB.
The guava trees presented the highest mean NOB, whereas the Paluma variety had the highest mean NOB, with 677.25 ovules per FB. The araçá trees were divided into three groups, and the accession with the lowest mean NOB was Araçá Una, with 19.25 ovules.
The estimated P:O ratio for P. guajava was 6,462.82; therefore, this species was considered xenogamous. As expected, the range for the P:O ratio did not differ significantly between the guava cultivars (Table 4).
Pollen:ovule ratio (P:O) and reproduction mode of eight accessions of Psidium spp. according to the methodology of Cruden (1977).
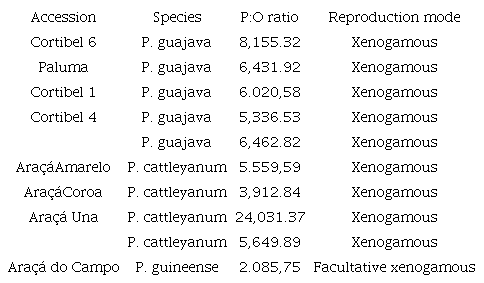
P. cattleyanum was classified as xenogamous because the obtained P:O ratio was 5,649.89. All accessions of this species were classified as xenogamous (Table 4), except for P. guineense, which was considered facultative xenogamous, with a P:O ratio of 2,085.75 (Table 4).
Raseira and Raseira (1996) studied the number of ovules, number of pollen grains, and number of anthers in four varieties of P. cattleyanum (“roxo planalto", “amarelo planalto", “amarelo litoral”, and “roxo sudeste”) and found that the mean number of ovules ranged from 94 to 165, which differs from our results in which the number of ovules varied from 19.25 to 677.25. However, the range was higher in the present study because three Psidium species were evaluated. It was lower (19.25 to 195.00) when only P. cattleyanum was analyzed.
The mean number of anthers found by Raseira and Raseira (1996) was similar to that observed in the three Psidium species evaluated. Both in the current study and in the Raseira and Raseira (1996) study, yellow araçá presented the highest number of anthers (Araçá Coroa, 451; “amarelo planalto", 480), and purple araçá presented the lowest number of anthers (Araçá Una, 191; “roxo sudeste”, 256). The most contradictory characteristic in the two studies was NPGA, in which the values in the three Psidium species analyzed in this study were higher than those in the four varieties of araçá evaluated by Raseira and Raseira (1996). However, the methodology used by these authors (Tuite, 1969) was different from that used in the present study (Dafni, 1992).
According to the classification of Cruden (1977), all evaluated species, P. guineense, P. cattleyanum, and P. guajava, were considered xenogamous, i.e., species that reproduce by cross-pollination (allogamous). P. guineense was considered facultative xenogamous, i.e., it may have some degree of self-pollination, where as P. guajava and P. cattleyanum presented the highest scores in Cruden’s classification (1977) and thus were considered xenogamous, i.e., some factors may limit self-pollination in these species. In these cases, pollination is performed primarily by pollinators (Cruden, 1977; Lenzi & Orth, 2004).
The probable reproduction mode of P. guajava is xenogamy, which agrees with previous studies that found cross-pollination was more common in guava (Dasarathy, 1951; Balasubrahmanyan, 1959).This conclusion is reinforced by the fact that the flower has a pleasant aroma and releases most of the pollen, attracting many visitors (Alves & Freitas, 2006). In addition, the Paluma variety has a modified spoon-shaped petal, which protects the stigma while the flower is closed, and this characteristic also favors allogamy (Alves & Freitas, 2007).
Corroborating this, studies report that honey bees (Apis mellifera), jandaíra bees (Melipona subnitida), and mamangava bees (Xylocopa frontalis) visit guava flowers and have grazing behavior consistent with that of pollinators (Alves & Freitas 2006; Alves & Freitas, 2007). This behavior explains why the comparison of different types of pollination in guava, including self-pollination, indicated that cross-pollination produced significantly more fruits and seeds (Alves & Freitas, 2007).
Raseira and Raseira (1996) suggested that P. cattleyanum, which was considered xenogamous, presented self-incompatibility because the results of the pollination test indicated that the penetration of the pollen tube into the ovule occurred in few pistils, demonstrating that the pollen tube stopped growing before reaching the ovary of the flower.
In addition, in the same study, Raseira and Raseira (1996) found that the percentage of fructification from self-pollination was lower than that from free pollination. However, fructification was also observed when the flowers were emasculated, protected, and not pollinated, indicating the formation of unfertilized fruits, probably originating from parthenocarpy or apomixis.
P. guineense was classified as facultative xenogamous considering the P:O ratio, which corroborates the results of da Silva, Araujo, Martins, Martins, and Veasey (2016), who found that there was an absence of endogamy and excess heterozygosity in four natural populations of P. guineense. This result suggests that the primary reproduction mode in this species is allogamy (da Silva et al., 2016) because a good indication of allogamy is the low frequency of loci in homozygotes (Allard, 1960).
Studies have reported that the P:O ratio of perennial species, including the three Psidium species studied, is higher than that of annual species, which are xenogamous in most cases (Cruden, 1977; Aarssen, 2000; Jurgens & Gottsberger, 2002; Galloni et al., 2007). This is because annual species have a short amount of time available to produce progeny, and thus self-pollination is advantageous by ensuring efficient reproduction in a limited time.
Other studies (Lopez et al., 1999; Jurgens & Gottsberger, 2002; Wang, Zhang, Renner, & Chen, 2005; Michalski & Durka, 2009) have found that the P:O ratio corresponds to the intervals estimated by Cruden (1977) or follows an expected pattern in which the P:O ratio of autogamous species is lower than that of allogamous species (Lehnebach & Riveiros, 2003; Medan, 2003). However, in some studies, the results of the P:O ratio disagree with the reproduction mode identified (Chouteau et al., 2006; Hokched & Ramírez, 2008).
This was the case in the study by Garcia, Miguez, and Gottsberguer (2014), who examined four species of Passiflora, two self-incompatible (P. caerulea and P. misera) and two self-compatible (P. foetida and P. suberosa) species and observed that although the P:O ratio was higher in self-incompatible species, the ratio classified P. caerulea between facultative autogamous and facultative xenogamous and P. misera as facultative xenogamous. This result indicates that the P:O ratio was lower than expected because these species are self-incompatible and self-pollination is not feasible.
Because of these disagreements, the estimates used for determining the reproduction mode have come under criticism. However, Cruden (1977) reported that the P:O ratio was one of the factors to be analyzed to predict the type of reproduction of the species. Therefore, further studies are necessary to confirm whether the P:O ratio in the three Psidium species corresponds to the type of reproduction of these species in the field.
Cruden (2000) noted that the P:O ratio may be confusing, especially for allogamous species with a low P:O ratio. However, other authors reported that species with precise or highly specialized pollen transfer mechanisms presented a P:O ratio lower than expected compared to species with a similar reproduction system but a less efficient pollen flow (Cruden & Miller-Ward, 1981; Chouteau et al., 2006).
Few studies have evaluated the reproductive biology of native species of the Myrtaceae family, especially those of the genus Psidium. With the exception of guava, no genetic improvement programs are available for wild Psidium species (Franzon, Campos, Proença, & Sousa-Silva, 2009). Therefore, studies seeking basic knowledge on the type of reproduction of native species are fundamental (Franzon et al., 2009) require many pollen grains to fertilize each ovule, demonstrating that the transfer of pollen to the stigma is not very specialized and corresponds to the intervals proposed by Cruden (1977) because genetic improvement depends primarily on the type of reproduction of these species.
The results indicate that the evaluated species; all species were classified as xenogamous. According to the P:O ratio, P. guajava and P. cattleyanum were classified as xenogamous, where as P. guineense was classified as facultative xenogamous.
However, considering that the P:O ratio is indicative of only the reproduction mode, additional studies are necessary to determine the presence of self-incompatibility and identify the factors that limit self-pollination in these species.
We thank CAPES and FAPERJ for financial support
*Author for correspondence. E-mail: fonsecaloliveira@gmail.com
Accessions of Psidium spp. maintained at the Antônio Sarlo Agricultural School, Campos dos Goytacazes, Rio de Janeiro State, Brazil.
Analysis of variance for the number of pollen grains per flower bud (NPGB), number of pollen grains per anther (NPGA), number of anthers per flower bud (NAB), number of ovules per flower bud (NOB), and pollen:ovule ratio (P:O) in eight accessions of Psidium ssp.
VF, variation factor; DF, degree of freedom; **different using the F-test at a level of significance of 0.01; CV(%), coefficient of variation.
Number of pollen grains per flower bud (NPGB), number of pollen grains per anther (NPGA), number of anthers per flower bud (NAB), and number of ovules per flower bud (NOB) in eight accesses of Psidium ssp.
The means followed by the same letters in the column do not differ using Tukey’s test at a level of significance of 0.05.
Pollen:ovule ratio (P:O) and reproduction mode of eight accessions of Psidium spp. according to the methodology of Cruden (1977).
