PLANT BREEDING
Multi-trait selection of tomato introgression lines under drought-induced conditions at germination and seedling stages

Multi-trait selection of tomato introgression lines under drought-induced conditions at germination and seedling stages
Acta Scientiarum. Agronomy, vol. 44, e55876, 2022
Editora da Universidade Estadual de Maringá - EDUEM
Received: 17 September 2020
Accepted: 27 January 2021
Funding
Funding source: Department of Plant Science at Universidade Federal de Viçosa (UFV
Funding source: Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES
Funding source: Fundação de Amparo a Pesquisa do Estado de Minas Gerais (FAPEMIG
Funding source: Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq
Contract number: (Finance Code 001
ABSTRACT: To be considered drought-tolerant, a tomato cultivar is required to present some level of tolerance at all developmental stages of plant growth. Since drought tolerance is a stage-specific phenomenon, genotype assessment must be performed separately at all developmental stages. In this study, we used a multi-trait index based on factor analysis and genotype-ideotype distance (FAI-BLUP index) to properly rank 49 tomato genotypes according to their tolerance to drought stress conditions at germination and seedling stages. Seeds of 47 introgression lines, which cultivar M82 is considered drought-sensitive, and the Solanum pennellii wild accession LA 716, which is considered drought-tolerant, were subjected to a control condition, where seeds were soaked in distilled water, and a drought condition, where seeds were soaked in a polyethylene glycol (PEG) solution (-0.3 MPa). Drought stress, induced by PEG, had a significant impact on all nine germination and growth performance-related traits; there was a reduction in shoot length (SL), total length (TL), initial germination percentage (IGP), final germination percentage (FGP), germination velocity index (GVI), and germination rate (GR). In contrast, the root-to-shoot ratio (R/S) and time to reach 50% germination (T50) increased under drought stress. Root length (RL) was less affected by drought, and in some genotypes, it was even increased. As expected, LA 716 ranked closest to the drought tolerance ideotype. IL 1-4-18, IL 2-3, IL 1-2, IL 9-2, and IL 10-1 were the most drought-tolerant at the germination stage. These results will serve as guidance for breeders who are aiming at developing drought-resistant tomato cultivars.
Keywords: Solanum lycopersicum L., Solanum pennellii Corr., genotype-ideotype distance, drought tolerance, water stress.
Introduction
Drought is one of the most significant environmental constraints that limits crop growth and yield in agriculture worldwide (Thabet, Moursi, Karam, Graner, & Alqudah, 2018). It is already known that a growing world population requires the production of more food by using less area and inputs such as water and fertilizers. The development of cultivars with tolerance to water deficit is, therefore, a sustainable and economically viable approach to enhance yield and ensure food security for the human population (Merchuk-Ovnat et al., 2016). Thus, plant breeding for drought tolerance is a promising strategy since it allows crop yield in water-limited environments and also makes it possible to grow crops using less water (Cattivelli et al., 2008).
Tomato (Solanum lycopersicum L.) is one of the most important vegetables in the world owing to its socioeconomic and nutritional importance (Rigano et al., 2016). Cultivated tomato is sensitive to drought stress from seed germination to harvest (Foolad, Zhang, & Subbiah, 2003). Tomato plants require approximately 0.89 L to 2.3 L of water daily during their growing cycle (Ayankojo, Morgan, Ozores-Hampton, & Migliaccio, 2018; Goyal & Sharma, 2018), with qualitative and quantitative losses during a drought period (George, Ahmad Jatoi, & Uddin Siddiqui, 2013; Zhou et al., 2019).
The development of drought-tolerant tomato plants faces several challenges. First, the trait is controlled by many genes with different effects and is highly influenced by environmental variation (Florido Bacallao & Bao Fundora, 2014; Foolad & Foolad, 2004). Thus, the precision and repeatability of field trials to assess drought tolerance are reduced because variations in environmental conditions such as rainfall, temperature, salinity, and nutrient availability cannot be fully controlled (Mohammadi, 2016). Another problem is that drought tolerance is a developmentally regulated stage-specific phenomenon. Therefore, specific stages throughout the ontogeny of plants must be evaluated separately for the assessment of drought tolerance and identification of its genetic components (Foolad, 2007).
Most of the recent tomato cultivars are drought-sensitive at all stages of plant development; the stage of seed germination and early seedling growth are the most sensitive (Mishra, Rai, Kumar, Singh, & Pandey, 2016). Genetic variability among the modern cultivated tomato is limited owing to natural and artificial selection during domestication and evolution (Steinhauser et al., 2011), which makes tomato wild relatives a good source of drought tolerance genes. Solanum pennellii Corr. is one of the crossable wild relatives of the cultivated tomato that displays drought tolerance (Bolger et al., 2014). Eshed and Zamir (1995) generated a collection of introgression lines (ILs) in which defined genomic segments of wild species replaced homologous regions in the background of the cultivated tomato variety M82 (S. lycopersicum). The IL population provides complete coverage of the wild species genome, aiding the investigation of the wild gene reservoir. Moreover, any phenotypic difference found between an IL and the recurrent parent M82 subjected to a drought condition can be attributed solely to the introgressed chromosomal segment of the drought-tolerant S. pennellii (Lippman, Semel, & Zamir, 2007). Thus, they are great sources for identifying genomic regions associated with drought tolerance and interactions between genomic regions (Schauer et al., 2006).
The screening of drought-tolerant genotypes during germination and seedling stages is often performed using polyethylene glycol (PEG) to simulate drought stress conditions (Ali et al., 2018; Feng, Wang, Shao, Zhang, & Tang, 2018; Ghebremariam, Liang, Li, Li, & Qin, 2013; Toscano, Romano, Tribulato, & Patanè, 2017). Drought tolerance in these trials was assessed by evaluating the genotype performance for several traits by using data from germination and seedling growth tests on a daily basis (Foolad et al., 2003), and genotype ranking for all the evaluated traits is performed simultaneously using selection indexes (Baker, 1986). Most of the available selection indexes require orthogonality among traits. Germination and seedling traits, however, are usually multicollinear. The FAI-BLUP, a multi-trait index based on factor analysis and ideotype design, proposed by Rocha, Machado, and Carneiro (2018), is used in this study; it combines factor analysis and genotype-ideotype design by using a multi-trait approach free from multicollinearity and has been successfully used for multi-trait selection of genotypes for several purposes (Oliveira et al., 2019; Rocha et al., 2019; Silva et al., 2018; Woyann et al., 2019).
Therefore, the purpose of the present study was to use the FAI-BLUP index to select drought-tolerant ILs and then identify the tomato genome regions associated with drought tolerance at germination and seedling stages. To be considered a drought-resistant, a tomato genotype must exhibit a fair level of drought resistance at all developmental stages of plant growth. Our findings will serve as guidance for breeders aiming at developing drought-resistant tomato cultivars.
Material and methods
Seed materials
Forty-seven ILs from the collection developed by Eshed and Zamir (1995) covering twelve tomato chromosomes (Figure 1) were chosen and tested for drought tolerance at germination and seedling stages. The cultivar M82, considered to be drought-sensitive, and S. pennellii accession LA716, considered to be drought-tolerant, were included in the experiment as testers.
Seeds of all genotypes were produced under uniform and optimum environmental conditions at the Research and Extension Farm Unit Horta Velha in Universidade Federal de Viçosa (UFV), Minas Gerais State, Brazil (20°45’14’ S; 42°52’53’ W). Seeds were collected from healthy mature fruits and subjected to natural fermentation for two days to remove the mucilaginous material surrounding the seed coat. After fermentation, the solution was passed through a fine-mesh sieve and washed with tap water to remove the mucilage. The cleaned seeds were then dried on filter paper at ambient temperature and stored in a storage room (13°C and 30% RH) inside paper bags until further use.
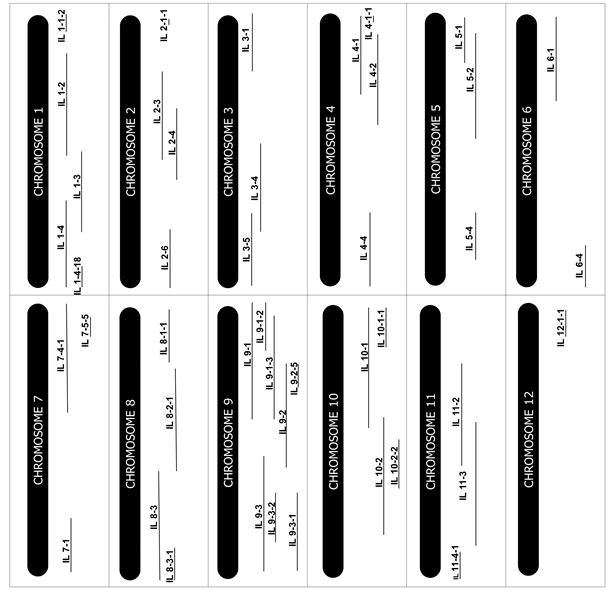
Figure 1
Scheme presenting the chromosomes and introgression sites of the evaluated ILs.
Seed germination and seedling growth
The study was performed in the Seed Laboratory at UFV. To assess drought tolerance at the germination and seedling stages, seeds of all genotypes were subjected to two different conditions: a control condition, where seeds were soaked in distilled water, and a drought condition, where seeds were soaked in a PEG solution (-0.3 MPa) to induce drought stress. The osmotic potential of -0.3 MPa was chosen here because it was reported to be the most suitable for selecting drought-tolerant tomato genotypes during germination and seedling stages (Borba, Maciel, Marquez, Junior, & Nogueira, 2017).
The experiment was arranged in a completely randomized design with four replicates. Each replicate consisted of an acrylic square box (11 x 11 x 3 cm) containing 25 seeds. The sterilized acrylic boxes were lined up with two layers of paper towels, moistened with distilled water in the control condition and a PEG-6,000 solution (-0.3 MPa) in the drought condition. The boxes were then placed inside plastic bags to avoid water loss and incubated in a growth chamber at a constant temperature of 25°C. Distilled water was sprayed on the paper to replace the evaporated water whenever it was noticed that they were dry. After incubation, the boxes were examined daily for 14 days to determine the number of germinated seeds (normal seedlings). On the 14th day, seedling length was measured. Both daily germination and seedling length data were analyzed using the SeedCalc R package (Silva, Medeiros, & Oliveira, 2019) to estimate nine germination and growth performance-related traits, as described in Table 1.

Statistical analysis
To assess genotype response to drought, the means of each parameter of drought-stressed seeds were divided by the means of their respective controls, as described by Thabet et al. (2018). Data were subjected to analysis of variance (ANOVA) to confirm significant differences between genotypes for each trait by using the GENES software (Cruz, 2013).
The univariate statistical model used to predict the genotypic values was denoted by:
where: y is the data vector, m is the vector of the effects of the measurement- combined replicates (assumed as fixed) added to the overall mean; g = vector of genotype effects (assumed as random); ɛ = vector of residue (random); X and Z represent the incidence matrices for these effects.
The transformed means were then used to rank the ILs according to their level of tolerance to drought using a multi-trait index based on factor analysis and genotype-ideotype distance (FAI-BLUP index), using the R script provided by Rocha et al. (2018) and R software (R Core Team, 2017). This methodology was adopted because it deals very well with multicollinearity problems, considers the correlation structure observed, and drives genotype ranking closer to the one hypothesized by researchers as the ideotype.
Principal component analysis was used to extract factorial loads of the phenotypic correlation matrix, obtained from the predicted genotypic values. Varimax criterion described by Kaiser (1958) was used for the analytic rotation, and the calculation of the factor scores of the weighted least squares was performed using the method proposed by Bartlett (1938).
The number of ideotypes was defined based on the combination of desirable and undesirable factors according to the selection purpose. The following algorithm gives the number of ideotypes: NI = 2n, where: NI = number of ideotypes and n = number of factors.
The drought-tolerant ideotype was built considering the maximum transformed mean values for IGP, GVI FGP, SL, RL, TL, and R/S and minimum mean values for T50 and GR; it is expected that a drought-tolerant genotype will show this performance. In the present study, besides the drought-tolerant ideotype, we further analyzed the ideotype for each of the four factors individually to better understand the performance of the genotype under drought.
After ideotype determination, genotype-ideotype distances were estimated and converted into spatial probability, enabling genotype ranking. The following algorithm was used:
where: Pij is the probability of the ith genotype (i = 1, 2, ... , n) to be similar to the jth ideotype (j = 1, 2,. . ., m); dij = Genotype-ideotype distance from the ith genotype to the jth ideotype, based on standardized mean Euclidean distance.
Results
The ANOVA detected significant genotype effect (p < 0.05) for all the evaluated traits in the two conditions. Therefore, all the nine traits were used in the following analyses.
Drought stress induced by the PEG 6,000 solution (-0.3 MPa) had a considerable impact on all evaluated traits (Figure 2). Under drought conditions, SL was reduced for most genotypes, which probably led to reductions in TL. RL was less affected by drought, and for some genotypes, it was even increased. The R/S increased in almost all the genotypes (Figure 2), but decrease in SL was more evident. IGP, FGP, GVI, and GR were also severely reduced under drought-induced conditions for all genotypes, as shown in Figure 2. IGP showed the highest reduction under drought condition compared to the other traits (Figure 2). Consistent with this result, the T50 of all genotypes increased under drought condition (Figure 2). Even though different performances were observed between ILs, in general, a decrease in germination percentage and germination velocity, followed by a reduction in seedling growth occurred under drought stress (Figure 2).

Figure 2
Heat map showing the effect of PEG-induced drought for all evaluated traits and genotypes. White or pale colors indicate no difference between the control and the drought-stress condition. The redder, the bigger the trait reduction; the bluer, the bigger the trait increase by drought exposure. SL = Shoot length, RL= Root length, TL = Total length, R/S= Root-to-shoot ratio, IGP = Initial germination percentage, GVI = Germination velocity index, FGP = Final germination percentage, T50 = Time to reach 50% germination, GR = Germination rate.
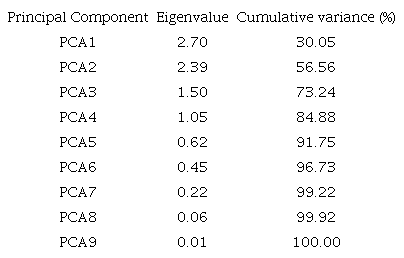
As for the principal component analysis, we found that from the eigenvalues and cumulative frequency for the nine principal components obtained via the phenotypic correlation matrix, the first four principal components had eigenvalues higher than one (Table 2). Thus, according to Kaiser’s criterion (Kaiser, 1958), the data may be condensed (dimensional reduction) into four factors. The cumulative frequency for the first four principal components or communality mean (common variance) was higher than 84%, indicating that they are sufficient to represent 84% of all data variability (Table 2).
After varimax rotation (Table 3), a high genetic correlation for the first factor was observed among the traits IGP, FGP, and GVI, and we named this factor germination. For the second factor, a high genetic correlation was observed among SL, RL, and TL, and we named it size. The third factor was named velocity of germination, to which T50 and GR were strongly correlated. The fourth factor was named root-to-shoot ratio and was formed only by R/S. The genetic correlations among traits within a factor can be given in the same and/or opposite directions.
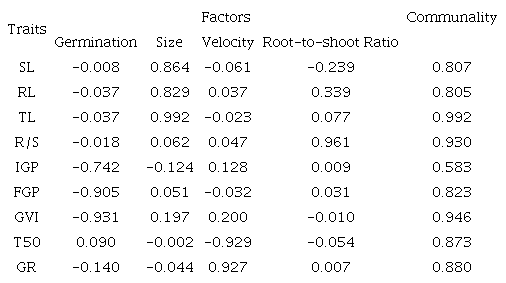
IGP = initial germination percentage; GVI = germination velocity index; FGP = final germination percentage; T50 = time to reach 50% germination; GR = germination rate; SL = shoot length; RL= root length; TL = total length; and R/S= root-to-shoot ratio.
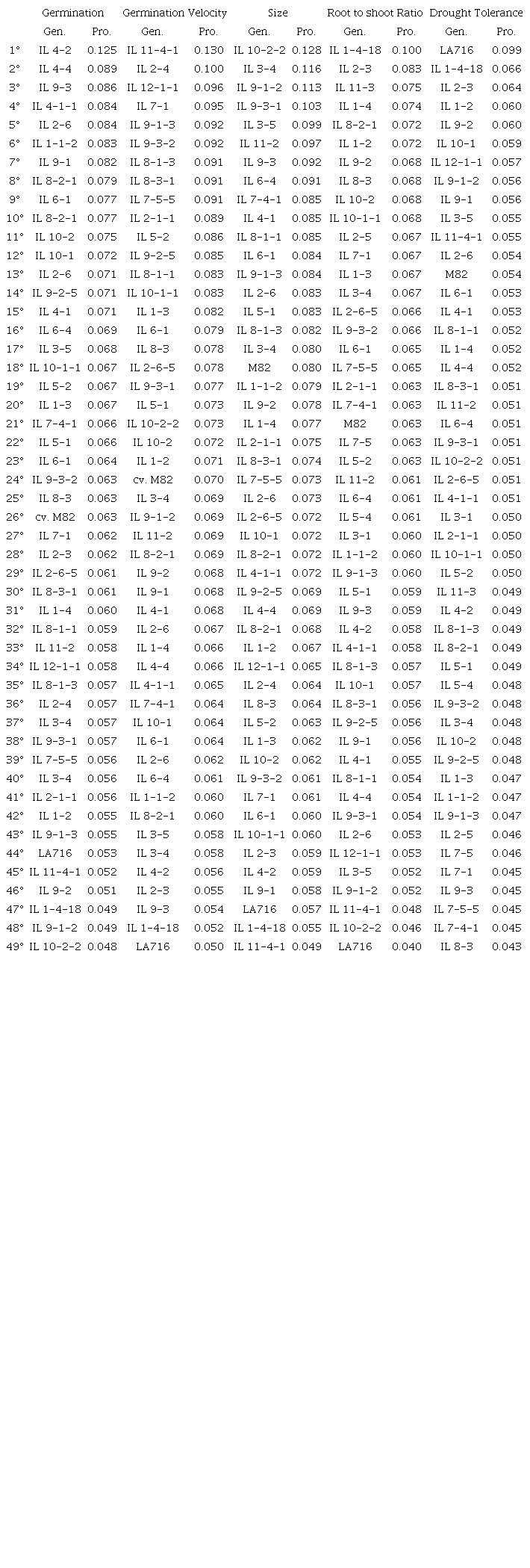
Table 4 shows genotype ranking according to the FAI-BLUP index and the probability associated with the distance from a given genotype to the ideotype. Besides the drought-tolerant ideotype, which shows desirable traits for all the five factors simultaneously, we decided to analyze the ideotype for each factor individually in order to better understand genotype performance under drought.
The accession LA716 (S. pennellii) was ranked closest to the drought tolerance ideotype during germination and seedling stages, followed by the introgression lines IL 1-4-18, IL 2-3, IL 1-2, IL 9-2, and IL 10-1. For the germination ideotype, the closest genotype was IL 4-2, whereas M82 was ranked 26th and LA716 44th. For velocity of germination, IL 11-4-1 was ranked closest to the ideotype, cultivar M82 was ranked 24th, and LA716 was the furthest genotype from the germination ideotype. The IL 10-2-2 was ranked closest to the length ideotype, and the cultivar M82 ranked 18th and LA716 47th. IL 1-4-18 was ranked closest to the root-to-shoot ratio ideotype.
Discussion
Water scarcity for agricultural production is a rising concern, especially for high water-demanding crops such as tomato, and plant breeding appears to be an economically viable approach to ensure food production in such arid areas. Since genetic tolerance to drought is a stage-specific phenomenon, the first step of a breeding program for drought tolerance is to identify drought-tolerant genotypes during the initial stages of plant growth. In this study, we used a multicollinearity free multi-trait index to properly rank 50 tomato genotypes according to their level of drought tolerance during germination and early seedling stages.
As expected, we found a significant genotype effect for all germination and initial seedling-growth traits, revealing genetic variability within the IL population. This genetic variability among ILs is associated with the small genomic segments of S. pennellii that replaced the homologous regions in the M82 background (Eshed & Zamir, 1995).
The low water potential of the external environment, promoted by PEG, made water less available to the seeds; hence, germination became either delayed or occurred at a reduced rate (Partheeban, Chandrasekhar, Jeyakumar, Ravikesavan, & Gnanam, 2017), which explains why drought imposed by PEG had a considerable impact on all evaluated traits (Figure 2). These drought effects on germination parameters were also recurrent for several other crops (Khodarahmpour, 2011). Low water potentials also result in reduced seedling growth owing to restricted cell enlargement and cell division (Borba et al., 2017). Consistent with other studies, shoot and root growths were inhibited by drought, and because shoot growth is more sensitive than root growth, the root-to-shoot ratio typically increases (Li, Mu, & Lin, 2014; Naveed, Mitter, Reichenauer, Wieczorek, & Sessitsch, 2014). This means that under drought stress, plants allocate more resources for root growth than for shoot growth in order to enhance water acquisition and limit evaporation (Lynch, 2007; Sharp, 2004).
Since genotype performance under drought conditions was very diverse, as shown in Figure 2, a given genotype can show an increase for some traits and a decrease for other traits. Therefore, we decided to utilize a multi-trait approach to properly rank and select the most drought-resistant genotypes. The FAI-BLUP index was chosen over other multivariate approaches because it performs the selection without multicollinearity problems. Moreover, it allows breeders to design an ideotype based on purpose and according to their dataset. In this way, the genotype ranking, associated simultaneously with spatial probability for multiple traits, is performed without the need to assign weights to each of the evaluated traits (Rocha et al., 2018).
This method was first proposed and applied for multi-trait selection of elephant grass (Pennisetum purpureum Schum.) for bioenergy purposes (Rocha et al., 2018). It was also recently used for multi-trait selection in several other studies. Woyann et al. (2019) used the FAI-BLUP index to select soybean (Glycine max [L.] Merrill) genotypes that best fit biodiesel production. Silva et al. (2018) evaluated lines and hybrids of biomass sorghum (Sorghum bicolor (L.) Moench) by using the same index. Oliveira et al. (2019) also used this index for the selection of photosensitive sorghum hybrids for energy cogeneration. Rocha et al. (2019) used the FAI-BLUP index for the selection of superior inbred progenies of common beans (Phaseolus vulgaris L.).
Multicollinearity is a common problem when handling multiple traits, even more, when traits are not orthogonal, as in the case of our study. It provides inflated errors, leading to inaccurate results regarding significance tests of predictors, and hence affects the selection process (Dormann et al., 2013; Prunier, Colyn, Legendre, Nimon, & Flamand, 2015). The use of factor analysis, as in the FAIBLUP index, produces uncorrelated or orthogonal axes among the final factors, generating scores without multicollinearity (Dormann et al., 2013). In addition, the approach, based on genotype-to-ideotype distance, focuses on multiple traits simultaneously, allowing breeders to consider all the evaluated traits and to underly plant morphological and physiological nuances (Van Oijen & Höglind, 2016). In fact, by analyzing drought tolerance ranking, germination, velocity of germination, size, and root-to-shoot ideotypes, we could better understand the behavior of ILs under drought conditions.
IL 4-2 was ranked closest to the germination ideotype, whereas M82 was ranked 26th and LA716 44th. Introgressed fragments of S. pennellii in the M82 background promoted an increase in the germination of 25 out of the 47 ILs, and 23 out of 47 ILs in the velocity of germination. However, these fragments alone did not result in the desired germination traits, for example, LA716 was one of the worst genotypes concerning germination and velocity of germination (Table 4). One possible hypothesis is that these traits are controlled by more than one gene and, hence, the increased germination and velocity of germination observed is probably due to the interaction between S. pennellii introgressed fragments and the rest of the M82 genome.
IL 10-2-2 was ranked closest to the length ideotype, M82 was ranked 18th and LA716 47th (Table 4). Therefore, we may infer that the S. pennellii introgressed fragments in M82 promoted an increase in seedling length-related traits for 17 out of the 47 evaluated ILs. However, as previously discussed, the M82 genome seems to be essential for its expression. Another explanation for why LA716 was found between the worst genotypes for seedling growth is that LA 716 seeds are smaller than the seeds from the other evaluated genotypes. Larger seeds tend to have a higher amount of nutritional reserves, which enable greater initial seedling development even under stress conditions such as drought (Pereira, Pereira, & Dias, 2013).
IL 1-4-18 was ranked closest to the root-to-shoot ratio ideotype (Table 4). The same IL was also closest to the drought tolerance ideotype. Some studies have shown that tolerant genotypes tend to sustain root-to-shoot ratios, whereas sensitive genotypes show a decrease in root-to-shoot ratios (Shamim, Saqlan, Habib-Ur-Rehman, & Waheed, 2014; Rahman, Nawata, & Sakuratani, 1999). In other crops, plant growth performance was positively associated with well-developed root systems and early seedling vigor, both of which can help to improve stress tolerance (Chloupek, Dostál, Středa, Psota, & Dvořáčková, 2010). Thus, focusing on such traits would be an efficient approach for phenotyping a large number of genotypes under drought conditions in a short time. Hence, selection based on shoot and root traits would be useful to identify genotypes for better performance under drought stress conditions. An important finding from this study is that the relationship between shoot and root, which have already been successfully implemented in existing high-throughput screening platforms, appears to be the most effective way to screen drought-tolerant genotypes prior to more laborious field trials. Similar findings were also observed for cotton and maize (Avramova et al., 2016; Riaz et al., 2013).
Although LA 716 did not perform well in terms of seedling germination and growth, it was ranked closest to the drought tolerance ideotype during germination (Table 4). As previously discussed, LA 716 seeds are smaller than those of the other genotypes. Due to the degree of contact between seed and water, sensitivity to drought is inversely related to seed size. This means that small seeds tend to be more efficient at absorbing water owing to an increased contact surface area between seed and substrate (Jiang & Su, 2018). Lima, Santiago, Araújo, and Teixeira (2005) and Pereira et al. (2013) observed that smaller seeds were less affected by drought because in smaller seeds, the amount of water required to reach a water content is smaller than that required by larger seeds.
Considering a 15% selection intensity, we selected ILs 1-4-18, IL 2-3, IL 1-2, IL 9-2, and IL 10-1 as drought-tolerant genotypes. The name of the ILs is related to the position and chromosome where the introgression was made. For example, IL1-1 means that one fragment of chromosome 1 of S. pennellii was introduced into the background genotype of M82 (Eshed & Zamir, 1995). Thus, in our findings, fragments of different S. pennellii chromosomes (1, 2, 9, and 10) and different regions within a chromosome are involved in determining drought tolerance during germination stage. Our findings are consistent because it is known that plants respond to drought by expressing quantitative traits, which involve multiple genes and physiological mechanisms. In addition, according to Foolad and Foolad (2004), drought tolerance during germination is genetically controlled, and additivity is the main genetic component. As M82 was ranked far from the drought tolerance ideotype, the improved performance of the selected ILs is related to the introgressed chromosomal segments of S. pennellii. The introgressions contain genes that contribute to the increased drought of these ILs.
Conclusion
In summary, the genotypes IL 1-4-18, IL 2-3, IL 1-2, IL 9-2, and IL 10-1 were selected as drought-tolerant during germination and seedling stages. They will be evaluated at other developmental stages of plant growth and be used as a source of drought tolerance genes in tomato breeding programs. The fragment 4-18 on chromosome 1, the fragment 3 on chromosome 2, the fragment 2 on chromosomes 1 and 9, and the fragment 1 on chromosome 10 can be used to identify more accurately the portions of S. pennellii genome that enhance tolerance to drought in tomato plants. Given the information of our study and the genome sequencing of S. lycopersicum and S. pennellii, the identification of candidate genes that control morphological and physiological traits related to drought tolerance at seed level will be made possible by exploring the map of the introgressed region. The identification of polymorphisms in the key genes controlling these traits can also be used to guide tomato breeding programs for drought tolerance.
Acknowledgements
The authors would like to acknowledge the Department of Plant Science at Universidade Federal de Viçosa (UFV). This work was supported by the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES) (Finance Code 001), Fundação de Amparo a Pesquisa do Estado de Minas Gerais (FAPEMIG) and Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq).
References
Ali, Q., Haider, M. Z., Iftikhar, W., Jamil, S., Tariq Javed, M., Noman, A., … Perveen, R. (2018). Drought tolerance potential of Vigna mungo L. lines as deciphered by modulated growth, antioxidant defense, and nutrient acquisition patterns. Revista Brasileira de Botânica, 39(3), 801-812. DOI: https://doi.org/10.1007/s40415-016-0282-y
Avramova, V., Nagel, K. A., Abdelgawad, H., Bustos, D., Duplessis, M., Fiorani, F., & Beemster, G. T. S. (2016). Screening for drought tolerance of maize hybrids by multi-scale analysis of root and shoot traits at the seedling stage. Journal of Experimental Botany, 67(8), 2453-2466. DOI: https://doi.org/10.1093/jxb/erw055
Ayankojo, I. T., Morgan, K. T., Ozores-Hampton, M., & Migliaccio, K. W. (2018). Effects of real-time location-specific drip irrigation scheduling on water use, plant growth, nutrient accumulation, and yield of Florida fresh-market tomato. HortScience, 53(9), 1372-1378. DOI: https://doi.org/10.21273/HORTSCI13183-18
Baker, R. J. (1986). Selection indices in plant breeding. Selection Indices in Plant Breeding.
Bartlett, M. S. (1938). Methods of estimating mental factors. Nature, 141, 609-610.
Bolger, A., Scossa, F., Bolger, M. E., Lanz, C., Maumus, F., Tohge, T., … Fernie, A. R. (2014). The genome of the stress-tolerant wild tomato species Solanum pennellii. Nature Genetics, 46(9), 1034-1038. DOI: https://doi.org/10.1038/ng.3046
Borba, M. E. A., Maciel, G. M., Marquez, G. R., Júnior, E. F. F., & Nogueira, G. G. S. (2017). Genetic diversity and selection in tomato genotypes under water stress induced by mannitol diversidade genética e seleção em genótipos de tomateiro sob estresse hídrico induzido por manitol. Bioscience Journal, 33(3), 592-600.
Cattivelli, L., Rizza, F., Badeck, F. W., Mazzucotelli, E., Mastrangelo, A. M., Francia, E., … Stanca, A. M. (2008). Drought tolerance improvement in crop plants: An integrated view from breeding to genomics. Field Crops Research, 105(1-2), 1-14. DOI: https://doi.org/10.1016/j.fcr.2007.07.004
Chloupek, O., Dostál, V., Středa, T., Psota, V., & Dvořáčková, O. (2010). Drought tolerance of barley varieties in relation to their root system size. Plant Breeding, 129(6), 630-636. DOI: https://doi.org/10.1111/j.1439-0523.2010.01801.x
Cruz, C. D. (2013). GENES - Software para análise de dados em estatística experimental e em genética quantitativa. Acta Scientiarum. Agronomy, 35(3), 271-276. DOI: https://doi.org/10.4025/actasciagron.v35i3.21251
Dormann, C. F., Elith, J., Bacher, S., Buchmann, C., Carl, G., Carré, G., … Lautenbach, S. (2013). Collinearity: a review of methods to deal with it and a simulation study evaluating their performance. Ecography, 36(1), 27-46. DOI: https://doi.org/10.1111/j.1600-0587.2012.07348.x
Eshed, Y., & Zamir, D. (1995). An introgression line population of Lycopersicon pennellii in the cultivated tomato enables the identification and fine mapping of yield-associated QTL. Genetics, 141(3), 1147-1162.
Farooq, M., Basra, S.M.A., Ahmad, N., & Hafeez, K. (2005). Thermal Hardening: A New Seed Vigor Enhancement Tool in Rice. Journal of Integrative Plant Biology, 47(2), 187-193. DOI: http://dx.doi.org/10.1111/j.1744-7909.2005.00031.x.
Feng, J., Wang, D., Shao, C., Zhang, L., & Tang, X. (2018). Effects of cold plasma treatment on alfalfa seed growth under simulated drought stress. Plasma Science and Technology, 20(3). DOI: https://doi.org/10.1088/2058-6272/aa9b27
Florido Bacallao, M., & Bao Fundora, L. (2014). Revisión bibliográfica.Tolerancia a estrés por déficit hídrico en tomate (Solanum lycopersicum L.). Cultivos Tropicales, 35(3), 70-88.
Foolad, M. R., & Foolad, M. R. (2004). Recent advances in genetics of salt tolerance in Tomato. Plant Cell, Tissue and Organ Culture, 76(2), 101-119. DOI: https://doi.org/10.1023/B:TICU.0000007308.47608.88
Foolad, M. R., Zhang, L. P., & Subbiah, P. (2003). Genetics of drought tolerance during seed germination in tomato: Inheritance and QTL mapping. Genome, 46(4), 536-545. DOI: https://doi.org/10.1139/g03-035
Foolad, M. R. (2007). Genome mapping and molecular breeding of tomato. International Journal of Plant Genomics, 2007, 1-53. DOI: https://doi.org/10.1155/2007/64358
George, S., Ahmad Jatoi, S., & Uddin Siddiqui, S. (2013). Genotypic differences against PEG simulated drought stress in tomato. Pakistan Journal of Botany, 45(5), 1551-1556.
Ghebremariam, K. M., Liang, Y., Li, C., Li, Y., & Qin, L. (2013). Screening of tomato inbred-lines for drought tolerance at germination and seedling stage. Journal of Agricultural Science, 5(11), 93-101. DOI: https://doi.org/10.5539/jas.v5n11p93
Goyal, M. R., & Sharma, P. (2018). Water requirement of drip-irrigated tomato inside a shade net house. In Engineering Interventions in Sustainable Trickle Irrigation (p. 35-42). DOI: https://doi.org/10.1201/9781315184241-2
International Seed Testing Association [ISTA]. (2015). The germination test. In International rules for seed testing (p. i-5-56). Zurich, SW: ISTA. DOI: http://dx.doi.org/10.15258/istarules.2015.05.
Jiang, Y., & Su, D. (2018). Models of turfgrass seed germination related to water content. PLoS ONE, 13(10), 1-13. DOI: https://doi.org/10.1371/journal.pone.0204983
Kaiser, H. F. (1958). The varimax criterion for analytic rotation in factor analysis. Psychometrika, 23(3), 187-200. DOI: https://doi.org/10.1007/BF02289233
Khodarahmpour, Z. (2011). Effect of drought stress induced by polyethylene glycol (PEG) on germination indices in corn (Zea mays L.) hybrids. African Journal of Biotechnology, 10(79), 18222-18227. DOI: https://doi.org/10.5897/AJB11.2639
Labouriau, L. G., & Valadares, M. B. (1976). On the germination of seeds of Calotropis procera. Anais da Academia Brasileira de Ciências, 48, 174-186.
Li, X., Mu, C., & Lin, J. (2014). The germination and seedlings growth response of wheat and corn to drought and low temperature in spring of Northeast China. Journal of Animal &Plant Sciences, 21(1), 3212-3222.
Lima, E. R., Santiago, A. S., Araújo, A. P., & Teixeira, M. G. (2005). Effects of the size of sown seed on growth and yield of common bean cultivars of different seed sizes. Brazilian Journal of Plant Physiology, 17(3), 273-281. DOI: https://doi.org/10.1590/s1677-04202005000300001
Lippman, Z. B., Semel, Y., & Zamir, D. (2007). An integrated view of quantitative trait variation using tomato interspecific introgression lines. Current Opinion in Genetics and Development , 17(6), 545-552. DOI: https://doi.org/10.1016/j.gde.2007.07.007
Lynch, J. P. (2007). Rhizoeconomics: The roots of shoot growth limitations. HortScience , 42(5), 1107-1109. DOI: https://doi.org/10.21273/HORTSCI.42.5.1107
Merchuk-Ovnat, L., Barak, V., Fahima, T., Ordon, F., Lidzbarsky, G. A., Krugman, T., & Saranga, Y. (2016). Ancestral qtl alleles from wild emmer wheat improve drought resistance and productivity in modern wheat cultivars. Frontiers in Plant Science, 2018(7), 1-14. DOI: https://doi.org/10.3389/fpls.2016.00452
Mishra, U., Rai, A., Kumar, R., Singh, M., & Pandey, H. P. (2016). Gene expression analysis of Solanum lycopersicum and Solanum habrochaites under drought conditions. Genomics Data, 9, 40-41. DOI: https://doi.org/10.1016/j.gdata.2016.04.001
Mohammadi, R. (2016). Efficiency of yield-based drought tolerance indices to identify tolerant genotypes in durum wheat. Euphytica, 211, 71-89. DOI: https://doi.org/10.1007/s10681-016-1727-x
Nakagawa, J., Krzyzanowski, F. C., Vieira, R. D., França-Neto, J. B. (1999). Testes de vigor baseados no desempenho das plântulas. In F.C. Krzyananowski, R. D.Vieira, & J. B. França-Neto (Eds.), Vigor de sementes: conceitos e testes. (p. 9-13). Londrina, PR: ABRATES.
Naveed, M., Mitter, B., Reichenauer, T. G., Wieczorek, K., & Sessitsch, A. (2014). Increased drought stress resilience of maize through endophytic colonization by Burkholderia phytofirmans PsJN and Enterobacter sp. FD17. Environmental and Experimental Botany, 97, 30-39. DOI: https://doi.org/10.1016/j.envexpbot.2013.09.014
Oliveira, I. C. M., Marçal, T. D. S., Bernardino, K. D. C., Ribeiro, P. C. D. O., Parrella, R. A. D. C., Carneiro, P. C. S., … Carneiro, J. E. D. S. (2019). Combining ability of biomass sorghum lines for agroindustrial characters and multitrait selection of photosensitive hybrids for energy cogeneration. Crop Science, 59(4), 1554-1566. DOI: https://doi.org/10.2135/cropsci2018.11.0693
Partheeban, C., Chandrasekhar, C. N., Jeyakumar, P., Ravikesavan., R., & Gnanam, R. (2017). Effect of PEG induced drought stress on seed germination and seedling characters of maize (Zea mays L.) genotypes. International Journal of Current Microbiology and Applied Sciences, 6(5), 1095-1104. DOI: https://doi.org/10.20546/ijcmas.2017.605.119
Pereira, W. A., Pereira, S. M. A., & Dias, D. C. F. S. (2013). Infuência do tamanho de semente e da restrição hídrica na germinação de sementes de soja e no desenvolvimento inicial das plântulas. Journal of Seed Science, 35(3), 316-322. DOI: https://doi.org/10.1590/S2317-15372013000300007
Prunier, J. G., Colyn, M., Legendre, X., Nimon, K. F., & Flamand, M. C. (2015). Multicollinearity in spatial genetics: separating the wheat from the chaff using commonality analyses. Molecular Ecology, 24(2), 263-283. DOI: https://doi.org/10.1111/mec.13029
R Core Team. (2017). R: A language and environment for statistical computing. Viena, AT: R Foundation for Statistical Computing.
Rahman, S. M. L., Nawata, E., & Sakuratani, T. (1999). Effect of water stress on growth, yield and eco-physiological responses of four tomato (Lycopersicon esculentum Mill.) cultivars. Engei Gakkai Zasshi, 68(3), 499-504. DOI: https://doi.org/10.2503/jjshs.68.499
Riaz, M., Farooq, J., Sakhawat, G., Mahmood, A., Sadiq, M., & Yaseen, M. (2013). Genotypic variability for root/shoot parameters under water stress in some advanced lines of cotton (Gossypium hirsutum L.). Genetics and Molecular Research , 12(1), 552-561. DOI: https://doi.org/10.4238/2013.February.27.4
Rigano, M. M., Arena, C., Di Matteo, A., Sellitto, S., Frusciante, L., & Barone, A. (2016). Eco-physiological response to water stress of drought-tolerant and drought-sensitive tomato genotypes. Plant Biosystems, 150(4), 682-691. DOI: https://doi.org/10.1080/11263504.2014.989286
Rocha, J. R. A. S. C., Machado, J. C., & Carneiro, P. C. S. (2018). Multitrait index based on factor analysis and ideotype-design: proposal and application on elephant grass breeding for bioenergy. GCB Bioenergy, 10(1), 52-60. DOI: https://doi.org/10.1111/gcbb.12443
Rocha, J. R. A. S. C., Nunes, K. V., Carneiro, A. L. N., Marçal, T. S., Salvador, F. V., Carneiro, P. C. S., & Carneiro, J. E. S. (2019). Selection of superior inbred progenies toward the common bean ideotype. Agronomy Journal, 111(3), 1181-1189. DOI: https://doi.org/10.2134/agronj2018.12.0761
Schauer, N., Semel, Y., Roessner, U., Gur, A., Balbo, I., Carrari, F., … Fernie, A. R. (2006). Comprehensive metabolic profiling and phenotyping of interspecific introgression lines for tomato improvement. Nature Biotechnology , 24(4), 447-454. DOI: https://doi.org/10.1038/nbt1192
Shamim, F., Saqlan, S. M., Habib-UR-Rehman A., & Waheed A. (2014). Screening and selection of tomato genotypes/cultivars for drought tolerance using multivariate analysis. Pakistan Journal of Botany46(4), 1165-1178.
Sharp, R. E. (2004). Root growth maintenance during water deficits: physiology to functional genomics. Journal of Experimental Botany , 55(407), 2343-2351. DOI: https://doi.org/10.1093/jxb/erh276
Silva, L. J., Medeiros, A. D., & Oliveira, A. M. S. (2019). SeedCalc, a new automated R software tool for germination and seedling length data processing. Journal of Seed Science , 41(2), 250-257. DOI: https://doi.org/10.1590/2317-1545v42n2217267
Silva, M. J., Carneiro, P. C. S., Carneiro, J. E. S., Damasceno, C. M. B., Parrella, N. N. L. D., Pastina, M. M., … Parrella, R. A. C. (2018). Evaluation of the potential of lines and hybrids of biomass sorghum. Industrial Crops and Products, 125, 379-385. DOI: https://doi.org/10.1016/j.indcrop.2018.08.022
Steinhauser, M. C., Steinhauser, D., Gibon, Y., Bolger, M., Arrivault, S., Usadel, B., … Stitt, M. (2011). Identification of enzyme activity quantitative trait loci in a Solanum lycopersicum x Solanum pennellii introgression line population. Plant Physiology, 157(3), 998-1014. DOI: https://doi.org/10.1104/pp.111.181594
Thabet, S. G., Moursi, Y. S., Karam, M. A., Graner, A., & Alqudah, A. M. (2018). Genetic basis of drought tolerance during seed germination in barley. PLoS One, 13(11), 1-21. DOI: https://doi.org/10.1371/journal.pone.0206682
Toscano, S., Romano, D., Tribulato, A., & Patanè, C. (2017). Effects of drought stress on seed germination of ornamental sunflowers. Acta Physiologiae Plantarum, 39(8), 184. DOI: https://doi.org/10.1007/s11738-017-2484-8
Van Oijen, M., & Höglind, M. (2016). Toward a Bayesian procedure for using process-based models in plant breeding, with application to ideotype design. Euphytica , 207(3), 627-643. DOI: https://doi.org/10.1007/s10681-015-1562-5
Woyann, L. G., Meira, D., Zdziarski, A. D., Matei, G., Milioli, A. S., Rosa, A. C., … Benin, G. (2019). Multiple-trait selection of soybean for biodiesel production in Brazil. Industrial Crops and Products , 140, 111721. DOI: https://doi.org/10.1016/j.indcrop.2019.111721
Zhou, R., Kong, L., Wu, Z., Rosenqvist, E., Wang, Y., Zhao, L., … Ottosen, C. O. (2019). Physiological response of tomatoes at drought, heat and their combination followed by recovery. Physiologia Plantarum, 165(2), 144-154. DOI: https://doi.org/10.1111/ppl.12764
Author notes
* Author for Correspondence. E-mail: herikapaulapessoa@gmail.com