Biotecnologia
Characterization of different microalgae cultivated in open ponds
Characterization of different microalgae cultivated in open ponds
Acta Scientiarum. Technology, vol. 41, 2019
Universidade Estadual de Maringá
Received: 17 June 2017
Accepted: 24 August 2017
Abstract: Microalga is one of the main sources of biomasses for the production of biofuels in the 21st century. Many labs and industries around the world are seeking for more productive strains and better cultivation models. The aim of this work was to evaluate the growth and kinetic parameters of six microalgae, and the moisture and lipid contents and fatty acids profiles of the biomasses harvested at the end of each cultivation. The microalgae were isolated in the Mid-Western Region of Brazil cultivated in open pond systems. It was observed that among the six microalgae used in this work, Pseudokirchneriella subcapitata presented the higher maximum specific growth rate (μmax) (0.12 day-1). The microalgae Coelastrum sp. and P. subcapitata were the ones with the highest lipid contents, with approximately 20% of dry mass. The main fatty acid accumulated in these conditions was palmitic acid, with percentages of 59 to 69% for all microalgae evaluated. Other factors that had influenced results from cultivations in open ponds were discussed here.
Keywords: growth kinetic, composition, lipid profile.
Introduction
Microalgae are photosynthetic microorganisms with high capacity of CO2 absorption for the transformation into chemical energy. Using some nutrients such as potassium, phosphorus and nitrogen, they can be cultivated in open ponds or closed photobioreactors (Cuellar-Bermudez, Garcia-Perez, Rittmann, & Parra-Saldivar, 2015). Due to their low nutrient requirements, they can be grown in different agroindustrial and household wastes or low value-added fertilizers (Sipaúba-Tavares & Rocha, 2003; Dragone, Fernandes, Vicente, & Teixeira, 2010).
There are several applications of microalgae in the agro-industry and in the pharmaceutical and cosmetic industries, as well as great potential for the production of biofuels. The production of biodiesel from microalgae is viable due to their capacity to synthetize lipids. For this reason, the optimization of lipid accumulation by microalgae becomes important (Yen et al., 2013). The cultivation of microalgae in open ponds is considered the best alternative for biomass production in large scale due to its easy construction, maintenance and operation (Park, Craggs, & Shilton, 2011).
In order to obtain increased productivities in sustainable production systems, it should be taken into account some factors, beyond the microalgae species utilized, e.g. geographic region, environment temperature, relative humidity of air, frequency of rainfalls, presence of sand and other pollutants, addition of carbon dioxide, price of the culture medium, pH, and the level of homogenization (Chisti, 2016; Silva & Fonseca, 2018).
In addition to the accumulation of lipids, the characterization of the saturated and unsaturated fatty acids synthetized is essential for the determination of biodiesel quality. This information can be used for a more accurate selection of a microalgae species for biodiesel production (Nascimento et al., 2013). The analysis of fatty acids is also important for the characterization of three important attributes of microbial communities: viable microbial biomass, community structure, and nutritional/physiological status (Piotrowska-Seget & Mrozik, 2003).
The aim of this work was to evaluate the physiology of 6 strains of microalgae (Chlorella sp., Coelastrum sp., Coleastrum sphaericum, Scenedesmus spinosos, Scenedesmus acuminatus and Pseudokirchneriella subcapitata) grew in open ponds in terms of kinetic parameters of growth and composition (moisture and lipid contents and fatty acids profile).
Material and method
The studied microalgae strains were Chlorella sp., Coleastrum sphaericum, Scenedesmus acuminatus and Scenedesmus spinosos and Coleastrum sphaericum and Pseudokirchneriella subcapitata, all previously isolated in the Brazilian Mid-Western region (Minillo, Godoy, & Fonseca, 2013). They were maintained in solid medium under refrigeration (4°C) in the absence of light at the Laboratory of Bioengineering from the Federal University of Grande Dourados.
Pre-inoculum was prepared in Chu-12 medium as described elsewhere (Sipaúba-Tavares & Rocha, 2003) in gallons of 9 L started with microalgae density of approximately 8 - 25 x 104 cells mL-1 at 25°C, light intensity of 10 Klux, 12 hours light photoperiods and aeration maintained by aquarium pumps, to obtain a higher cell density to later inoculate the open ponds. After 60 days, the 9L precultures were transferred to rectangular open ponds (3.5 x 1.0 x 0.6 m) filled with 2000L of NPK medium each, prepared by diluting 1 g of the chemical fertilizer N:P:K (20:5:20) for each liter of water (Sipaúba-Tavares & Rocha, 2003). The water was pumped into ponds from a local water well. In the ponds, the microalgae were exposed to environmental conditions (sunlight, temperature and rainfall) without control. For monitoring the growth of microalgae, samples (5 mL) were collected every 7 days to measure the absorbance at 600 nm (ABS600) in duplicate (Demirel, Demirkaya, Imamoglu, & Dalay, 2016). This wave-length was chosen to avoid the interference caused by the production of pigments (Griffiths, Garcin, van Hille, & Harrison, 2011) to evaluate microbial cell growth in terms of optical density (OD) during 49 days of cultivation.
At the end of the cultivations, samples were collected from each pond, centrifuged (5 g, 10 min.) and maintained in a drying oven with air circulation at 70°C up to constant weight to determine moisture. The dried biomasses were utilized to the quantification of lipids by the cold method (Bligh & Dyer, 1959) and fatty acids. All determinations were performed in triplicate.
For the characterization of fatty acids, the HCl transesterification was performed in methanol, in which the lipid extracts also extracted by the cold method were added of 500 µL of 5% HCl in methanol and the mixture was incubated for 2 hours at 100°C. After the reaction, 1.25 mL was added and the fatty acid methyl esters (Fame) were extracted with 1.25 mL of hexane (Martins, Yokoya, & Colepicolo, 2012). The Fame were determined by gas chromatography coupled to mass spectrometry (GC-MS), using a chromatograph (Agilent, model 7890A) equipped with a 30m x 0.25mm x 0.25m HP-5MS capillary column with helium entrapped as entrainment gas at a split ratio of 1:10. The analyses were performed at starting temperature of 180°C, applying heating rate of 10°C min-1 up to 300°C. The detector and injector temperatures were 325 and 300°C, respectively. The analyses were performed in duplicate. The Fame were identified by comparing the retention time of sample’s constituents with a mixture of 19 fatty acid methyl ester standards, and by comparison with mass spectra of the NIST MS Search 2.0 library, included in the chromatograph’s software. Quantification was performed relative to an internal standard of methyl tricosanoate (23:0), following the method described elsewhere and maintaining a ratio of about 200:1 between the mass of oil and mass of internal standard (Menegazzo, Petenuci, & Fonseca, 2014).
Meteorological data of solar radiation (KWh m-2 day-1), temperature (ºC), precipitation (mm), and relative humidity (RH) (%) of Dourados were obtained from Embrapa Centro-Oeste, Dourados, state Mato Grosso do Sol for the days corresponding to the cultivations in the open ponds. Temperature (ºC) and pH were also obtained in situ.
The exponential growth phase was identified as the linear region on an ln ABS600 vs. time plot for batch cultivation data. The maximum specific growth rate (µmax) was determined as the slope of this linear region. The doubling time (DT) was calculated by the quotient of the ln (2) by the µmax. The maximum biomass concentration was indicated by the maximum ABS600 observed in each experiment (Nascimento, Silva, Gomez, & Fonseca, 2016). Measured absorbance values were converted into mass values using a linear relationship of optical density units per gram dry cell mass to calculate biomass and volumetric lipid productivities.
The composition data were subjected to analysis of variance (ANOVA) at 5% probability by Tukey test, using the software Statistica version 6.0 (Statsoft, Tulsa, USA).
Results and discussion
During the cultivations, the average meteorological data were 26.2 ± 2.0ºC, 76.6 ± 8.0% of relative humidity of air, 12.6 ± 3.2 KWh m-2 day-1 of solar radiation, and precipitation ranging from 0 up to 44.6 mm (29 without any precipitation) (Figure 1). The temperature of the media in the open ponds was 30 ± 5 ºC and the pH ranged from 8.9 to 9.9 in the cultures (Table 1). This information is important to characterize the strains, e.g. in terms of productivities and lipid accumulation, and it is important to compare the obtained data with the literature at similar conditions.
Chisti (2016) numerated several recommendations for optimal cultivation in open ponds. Among the items are, the temperature between the 24 and 40ºC, cheap medium, and strains isolated in the region, which were according the suggested. However, there was no homogenization and control of level of pollutants and contaminants
These two recommendations in fact influenced microalgae growth, e.g., it was noticed that part of the microalgal mud was composed by sand, brought by the wind. This material had to be removed by decanting before algae centrifugation. The same had to be done to obtain the correct absorbance measure along the samplings. The maximal absorbance obtained are shown in Table 1.
The growth kinetics for the six microalgae stains cultivated in open ponds with NPK medium during 49 days are shown in Figure 2. The growth profiles suggest a slow cell development. Growth rates are usually lower in open ponds compared to cultures in closed photobioreactors (Mata, Martins, & Caetano, 2010). In order to optimize growth in open ponds, a good homogenization is important. Control of the growth parameters, e.g. pH, light and nutrients is also desired (Park et al., 2011).
It was observed that Pseudokirchneriella subcapitata presented the highest maximum specific growth rate (μmax) and the shortest doubling time (DT), with values of 0.12 day-1 and 5.40 days, respectively (Table 1). It was reported elsewhere an exponential growth rate of 0.11 to 0.13 day-1 for P. subcapitata cultivated in NPK (Carvalho et al., 2012).
Regarding cell composition, moisture remained in the range of 90.33 to 94.77% for the different microalgae while the lipid content reached the higher content for Coelastrum sp. and P. subcapitata, which presented an average of 20% of lipids of the total dry weight (Table 1). In cultivation in open ponds, isolated microalgae Graesiella sp. WBG-1 accumulated up to 33.4% of lipids in its dry mass (Wen et al., 2016).
Although the growth rate is one of the crucial factors in the production of microalgae, it is important that the biomass present a high lipid yield (Stemmler, Massimi, & Kirkwood, 2016). Hannon, Gimpel, Tran, Rasala, and Mayfield (2010) suggested that strains with increased growth rates and high oil accumulation are better obtained by the selection of new species and genetic improvement.
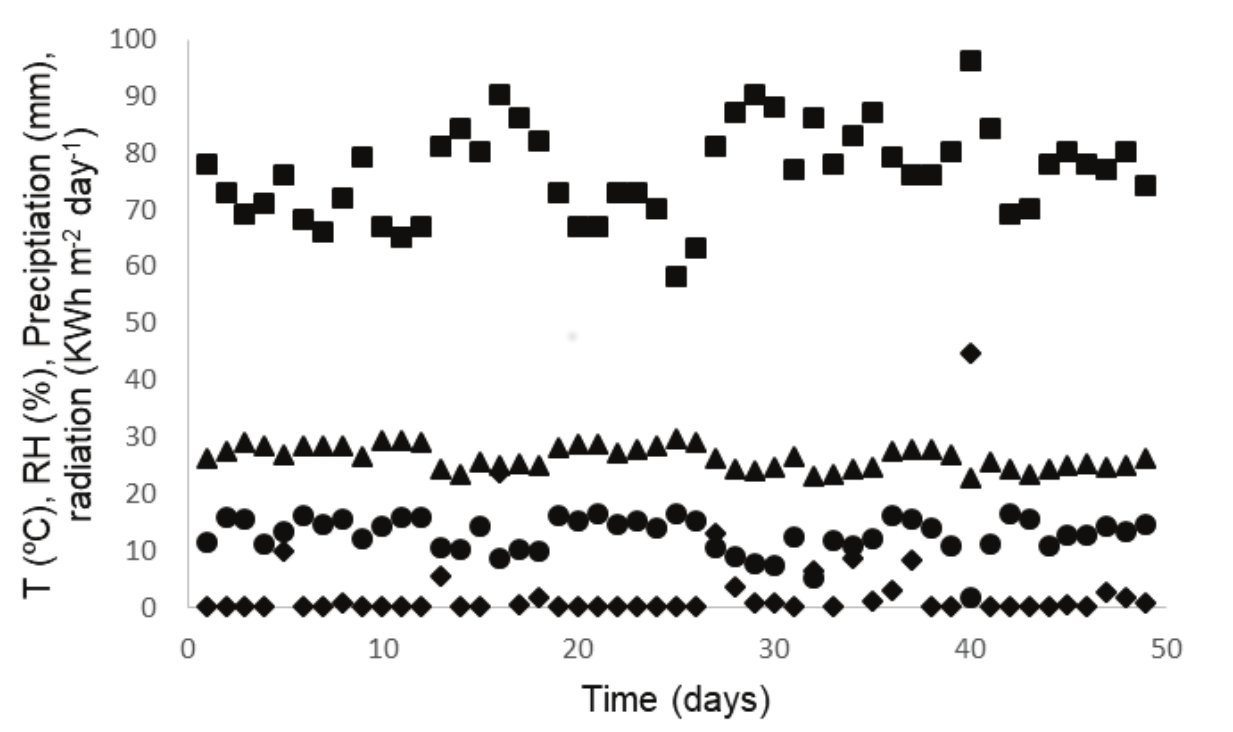
Figure 1.
Meteorological data of (●) radiation (KWh m-2 day-1), (▲) temperature (ºC), (◆) precipitation (mm) and (■) relative humidity (RH) (%) for the days corresponding to the cultivations in the open ponds.
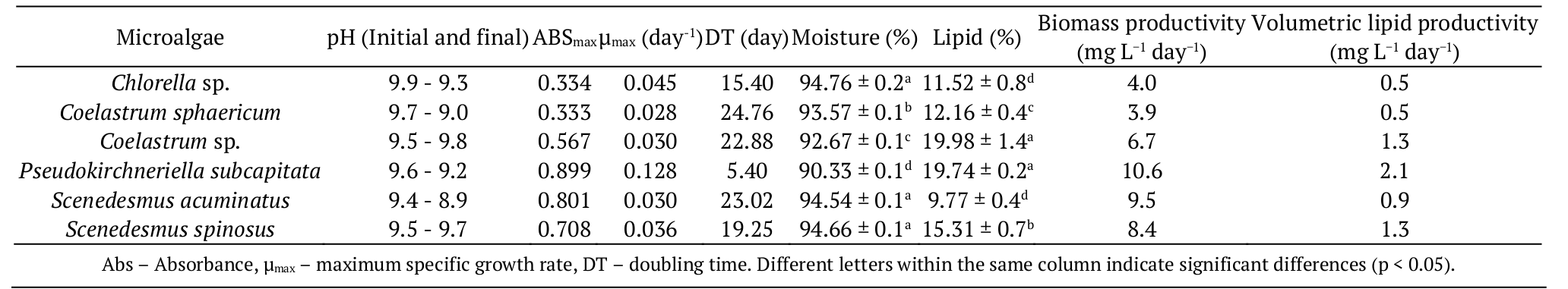
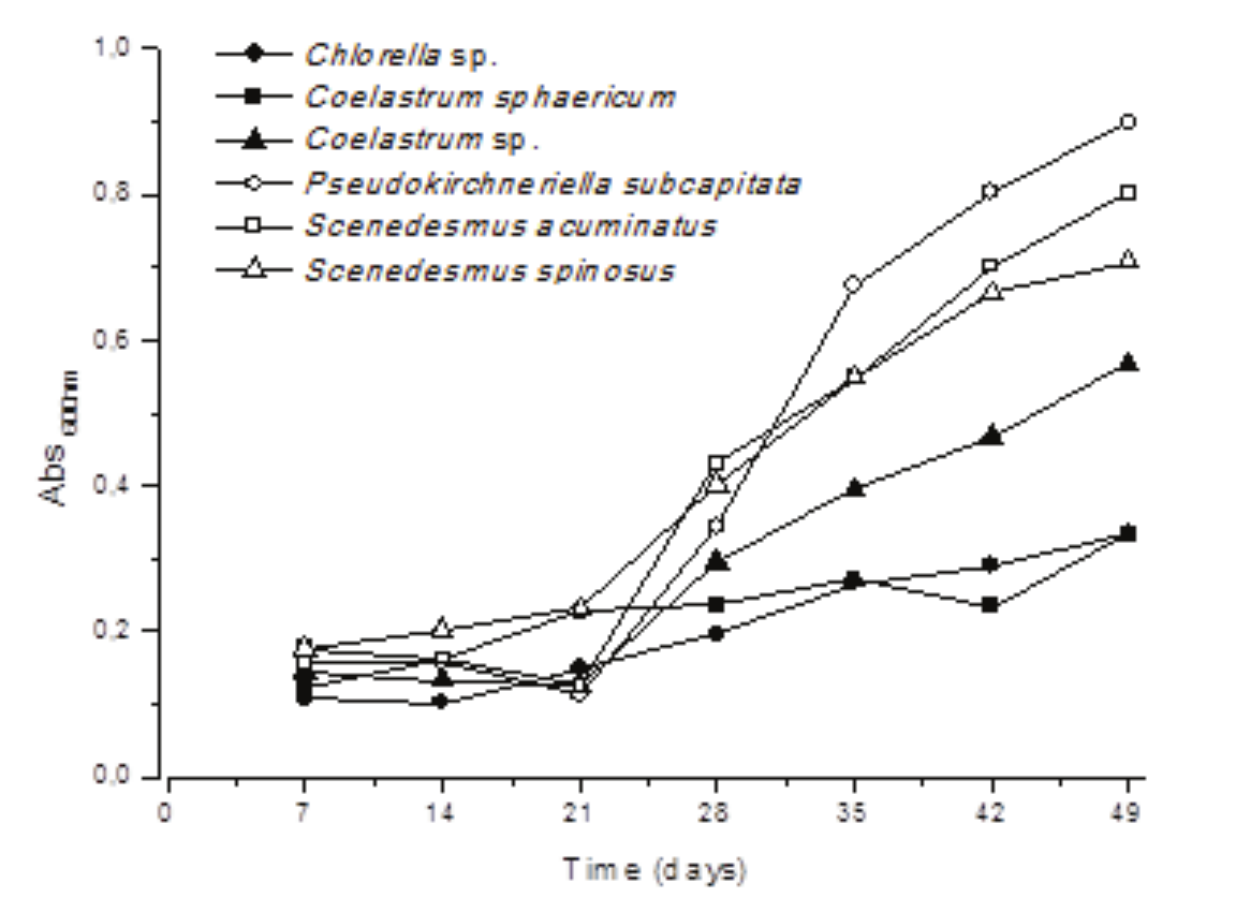
Figure 2.
Growth kinetics of Chlorella sp., Pseudokirchneriella subcaptata, Coelastrum sp., Coelastrum sphaericum, Scenedesmus spinosus and Scenedesmus acuminatus in open ponds during 49 days.
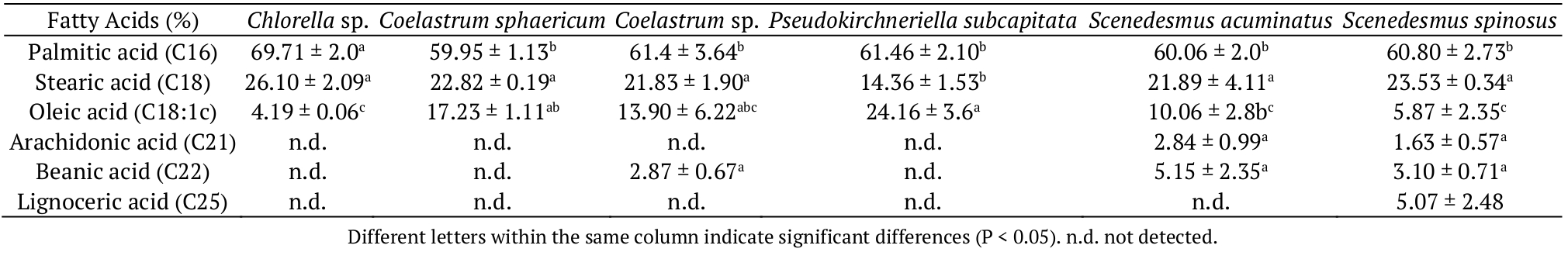
Oils were extracted from cellular content to determine the fatty acid composition. Palmitic acid (C16) was the predominant fatty acid in most microalgae lipid extracts (Table 2), which is in agreement with literature (Nascimento et al., 2013). A high accumulation of palmitic acid (69.71%) was described elsewhere for Chlorella sp. (Kalpish, Sharma, & Peer, 2012).
It was observed that the genus Scenedesmus presented a fatty acids profile different from the other species. S. spinosus synthetized both polyunsaturated fatty acids (PUFAs) C21 and C25, while S. acuminatus presented the C25 in its composition (Table 2). The PUFAs present a series of special applications mainly as functional food, helping in the physical, physiological and mental development of the human organism (Martins, Suaiden, Piotto, & Barbosa, 2008).
The fatty acids content can be altered due to their accumulation or their use as an energy source. These changes occur through enzymes that act on the β-oxidation pathway, varying the fatty acid chains in two carbon units, which explains why most fatty acid chains are pairs (Voet, Voet, & Pratt, 2008; Taiz & Zeiger, 2008). Only the microalgae S. acuminatus and S. spinosus showed odd-chain fatty acids (Table 2), which is not usually observed. This happens in some plants and marine organisms, because the last cycle of β-oxidation provides propionyl-CoA that can be converted to succinyl-CoA, which enters the cycle of citric acid. This reaction requires the involvement of three enzymes: propionyl-CoA-carboxylase, methylmalonyl-CoA-racemase and methylmalonyl-CoA mutase (Voet et al., 2008). The question is that to achieve this transformation, it is necessary the coenzyme B12, which is not found in bacteria (Voet et al., 2008; Taiz & Zeiger, 2008). It suggests that bacteria could have contaminated the open ponds, making this coenzyme available in the culture, which would have enabled the generation of fatty acids of odd chains.
Bacteria incorporate the fatty acids, mainly in phosphoslipids (Piotrowska-Seget & Mrozik, 2003). This characteristic is attributed to the ability to adapt to various extreme conditions (Sahu et al., 2013). Moreover, the chemical composition of microalgae alters when it is present in the presence of bacteria (Stemmler et al., 2016).
The evolution of the research in relation to the media, the way of cultivation, types of photobioreactor and later processes are of global interest so that the microalgae cultivation becomes an economically viable alternative for the obtaining of energy (Pereira et al., 2012).
Conclusion
Beyond microalgae species, several other factors influenced results from cultivations in open ponds, which were polluted with sand and contaminated with bacteria. However, it was possible to evaluate the kinetic parameters of six microalgae strains. The microalga species that presented the higher maximum specific growth rate in open ponds was Pseudokirchneriella subcaptata. Pseudokirchneriella subcaptata and Coelastrum sp. presented the highest lipid content, but the genus Scenedesmus presented a differentiated fatty acids profile, including C21 and C25 polyunsaturated fatty acids, despite it was not clear if these compounds were resulting from contaminant bacteria.
Acknowledgements
The authors gratefully acknowledge the Brazilian research funding agencies CNPq and Fundect for their financial support.
References
Bligh, E. G., & Dyer, W. J. (1959). A rapid method of total lipid extraction and purification. Canadian Journal of Biochemistry and Physiology, 37(8), 911-917. doi: 10.1139/o59-099
Carvalho, E. M., Ottonelli, F., Ansilago, M., Godoy, H. C., Nakagaki, J. M., & Ramires I. (2012). Growth kinetics of the microalga Pseudokirchneriella subcapitata (Korshikov) Hindak (Chlorophyceae) in natural water enriched with NPK fertilizer. Biochemistry and Biotechnology Reports, 1(2), 14-18. doi: 10.5433/2316-5200.2012v1n2p14
Chisti, Y. (2016). Large-scale production of algal biomass: raceway ponds. In F. Bux, & Y. Chisti (Eds.), Algae biotechnology: Products and processes, VI (p. 21-40). Basel, SW: Springer Nature. doi: 10.1007/978-3-319-12334-92
Cuellar-Bermudez, S. P., Garcia-Perez, J. S., Rittmann, B. E., & Parra-Saldivar, R. (2015). Photosynthetic bioenergy utilizing CO2: an approach on flue gases utilization for third generation biofuels. Journal of Cleaner Production, 98, 53-65. doi: 10.1016/j.jclepro.2014.03.034
Demirel, Z., Demirkaya, C., Imamoglu, E., & Dalay, M. C. (2016). Diatom cultivation and lipid productivity for non-cryopreserved and cryopreserved cells. Agronomy Research, 14(4), 1266-1273.
Dragone, G., Fernandes, B., Vicente, A., & Teixeira, J. (2010). Third generation biofuels from microalgae. In A. Méndez-Vilas, (Ed.), Current research, technology and education topics in applied microbiology and microbial biotechnology (p. 1355-1366). Madrid, ES: Formatex.
Griffiths, M. J., Garcin, C., van Hille, R. P., & Harrison, S. T. L. (2011). Interference by pigment in the estimation of microalgal biomass concentration by optical density. Journal of Microbiological Methods, 85(2), 119-123. doi: 10.1016/j.mimet.2011.02.005
Hannon, M., Gimpel, J., Tran, M., Rasala, B., & Mayfield, S. (2010). Biofuels from algae: challenges and potential. Biofuels, 1(5), 763-784.
Kalpish, K., Sharma, H. S., & Peer, M. S. (2012). High lipid induction in microalgae for biodiesel production. Energies, 5(5), 1532-1553. doi: 10.3390/en5051532
Martins, A. P., Yokoya, N. S., & Colepicolo, P. (2012). Comparison of extraction and transesterification methods on the determination of the fatty acid contents of three Brazilian seaweed species. Brazilian Journal of Pharmacognosis, 22(4), 854-860. doi: 10.1590/S0102-695X2012005000088
Martins, M. B., Suaiden, A. S., Piotto, R. F., & Barbosa, M. (2008). Propriedades dos ácidos graxos poliinsaturados - Omega 3 obtidos de óleo de peixe e óleo de linhaça. Revista do Instituto de Ciências da Saúde, 26(2), 153-159.
Mata, T. M., Martins, A. A., & Caetano, N. S. (2010). Microalgae for biodiesel production and other applications: A review. Renewable and Sustainable Energy Reviews, 14(1), 217-232. doi: 10.1016/j.rser.2009.07.020
Menegazzo, M. L., Petenuci, M. E., & Fonseca, G. G. (2014). Production and characterization of crude and refined oils obtained from the co-products of Nile tilapia and hybrid surubim processing. Food Chemistry, 157, 100-104. doi: 10.1016/j.foodchem.2014.01.121
Minillo, A., Godoy, H. C., & Fonseca, G. G. (2013). Growth performance of microalgae exposed to CO2. Journal of Clean Energy Technologies, 1(2), 110-114. doi: 10.7763/JOCET.2013.V1.26
Nascimento, I. A., Marques, S. S. I., Cabanelas, I. T. D., Pereira, S. A., Druzian, J. I., Souza, C. O., ... Nascimento, M. A. (2013). Screening microalgae strains for biodiesel production: lipid productivity and estimation of fuel quality based on fatty acids profiles as selective criteria. BioEnergy Research, 6(1), 1-13. doi: 10.1007/s12155-012-9222-2
Nascimento, V. M., Silva, L. F., Gomez, J. G. C., & Fonseca, G. G. (2016). Growth of Burkholderia sacchari LFM 101 cultivated in glucose, sucrose and glycerol. Scientia Agricola, 73(5), 429-433. doi: 10.1590/0103-9016-2015-0196
Park, J. B. K., Craggs, R. J., & Shilton, A. N. (2011). Wastewater treatment high rate algal ponds for biofuel production. Bioresource Technology, 102(1), 35-42. doi: 10.1016/j.biortech.2010.06.158
Pereira, C. M. P., Hobuss, C. B., Maciel, J. V., Ferreira, L. R., Del Pino, F. B., & Mesko, M. F. (2012). Biodiesel renovável derivado de microalgas: avanços e perspectivas tecnológicas. Química Nova, 35(10), 2013-2018. doi: 10.1590/S0100-40422012001000022
Piotrowska-Seget, Z., & Mrozik, A. (2003). Signature lipid biomarker (SLB) analysis in determining changes in community structure of soil microorganisms. Polish Journal of Environmental Studies, 12(6), 669-675.
Sahu, A., Pancha, I., Jain, D., Paliwal, C., Ghosh, T., Patidar, S., … Mishra, S. (2013). Fatty acids as biomarkers of microalgae. Phytochemistry, 89, 53-58. doi: 10.1016/j.phytochem.2013.02.001
Silva, A. A., & Fonseca, G. G. (2018). Bioprospection of freshwater microalgae from Bonito, MS, Brazil. International Journal of Biodiversity and Conservation, 10(3), 117-121. doi: 10.5897/IJBC2017.1102
Sipaúba-Tavares, L. H., & Rocha, O. (2003). Produção de plâncton (fitoplâncton e zooplâncton) para alimentação de organismos aquáticos (2 ed.). São Carlos, SP: Rima.
Stemmler, K., Massimi, R., & Kirkwood, A. E. (2016). Growth and fatty acid characterization of microalgae isolated from municipal waste-treatment systems and the potential role of algal-associated bacteria in feedstock production. PeerJ, 4, e1780. doi: 10.7717/peerj.1780.
Taiz, L., & Zeiger, E. (2008). Fisiologia vegetal (4a ed.). Porto Alegre, RS: Artmed.
Voet, D., Voet, J. G., & Pratt, C. W. (2008). Fundamentos de bioquímica: a vida em nível molecular (2 ed.). Porto Alegre, RS: Artmed.
Wen, X., Du, K., Wang, Z., Peng, X., Luo, L., Tao, H., … Li, Y. (2016). Effective cultivation of microalgae for biofuel production: a pilot-scale evaluation of a novel oleaginous microalga Graesiella sp. WBG-1. Biotechnology for Biofuels, 9, 123. doi: 10.1186/s13068-016-0541-y
Yen, H. W., Hu, C., Chen, C. Y., Ho, S. H., Lee, D. J., & Chang, J. S. (2013). Microalgae-based biorefinery - From biofuels to natural products. Bioresource Technology, 135, 166-174. doi: 10.1016/j.biortech.2012.10.099
Author notes
ggf@ufgd.edu.br