Abstract: The objective of this study was to evaluate the use of cocoa shell as an alternative substrate for the production of endoglucanase and xylanase by Aspergillus orizae ATCC 10124 in solid-state fermentation. The fermentation assays were performed using cocoa shells humidified with distilled water. The parameters incubation temperature, water activity and fermentation time were optimized by using a Box-Behnken design. The maximum activities achieved for endoglucanases and xylanases were 0.846 and 0.945 U g-1 at 35°C, with a water activity equal to 0.9 and an incubation time of 96 hours.
Keywords:wastewaste,solid-state fermentationsolid-state fermentation,Box-Behnken planningBox-Behnken planning.
Science, Food Technology and Food Engineering
Cocoa shell as a substrate for obtaining endoglucanase and xylanase from Aspergillus oryzae ATCC 10124
Universidade Estadual de Maringá

Esta obra está bajo una Licencia Creative Commons Atribución 4.0 Internacional.
Recepción: 03 Junio 2019
Aprobación: 05 Agosto 2019
Due to their catalytic efficiency, enzymes are used in industrial processes, among them, in the food industry to clarify beverages (Sharma, Patel, & Sugandha, 2017), in the pharmaceutical industry to produce chiral therapeutic drugs (Li, Wang, & Reetz, 2018), in the textile industry in the process of bleaching and in biopreparation, during which non-cellulosic impurities are removed (Easson, Condon, Villalpando, & Chang, 2018).
Solid-state fermentation (SSF) uses agro-industrial residues as a source of nutrients for the growth of filamentous fungi and the secretion of enzymes (Soccol et al., 2017). The application of these residues occurs due to its composition of lignin (10 to 30%), hemicellulose (15 to 35%) and cellulose (30 to 50%), the concentration of each of these elements being variable according to the type of residue (Santos et al., 2013).
For the production of enzymes, several substrates are used: cane bagasse (Biz et al., 2016), caja bran (Ferraz et al., 2018), wheat bran (Zhao, Guo, & Zhu, 2017), corn cobs (Shi, Zhang, Lu, & Wang, 2017), rice husk bran (Marques et al., 2017), beet peel (Joana et al., 2018), potato peel (Santos, Gomes, Bonomo, & Franco, 2012), and forage palm bran (Ferreira et al., 2017). In Brazilian agriculture, many crops are considered waste generators. Among these, cocoa may be included. Cocoa is a much-appreciated fruit and valued for its sweet, slightly acidic taste. After fermentation and drying, the almonds are toasted, and the shell that surrounds them is easily peeled (Lessa et al., 2018). The chocolate industry produces high amounts of cocoa shell. It is estimated that chocolate processing produces about 54% of its weight in residues, which have no specific destination and are, in most cases, incinerated or discarded in nature (Okiyama, Navarro, & Rodrigues, 2017). Our research group used cocoa shell in the production of lipases in the extraction of bioactive compounds and fatty acids (Lessa et al., 2018). Other researchers used cocoa shell for the production of activated carbon (Kalaivani et al., 2014), biogas production (Mancini, Papirio, Lens, & Esposito, 2016), and enrichment of animal feed (Amorim, Oliveira, Gutarra, Godoy, & Freire, 2017).
The cocoa shell contains high levels of lignin (30.7%), cellulose (9.7%), and hemicellulose (15.4%; Lessa et al., 2018), which can be used in SSF for the production of enzymes, among which cellulases can be mentioned.
Cellulases are enzymes that act synergistically in the hydrolysis of cellulose to generate monomeric units. Endoglucanases, or Endo-1,4-β-glucanase (EG) cleave intramolecular β-1,4-glucosidic bonds in the amorphous regions of the cellulose fibre and produce new chain ends and larger oligosaccharides (Santos et al., 2016).
The xylanase enzymes constitute the xylanolytic system and play a key role in the depolymerization of xylan present in the plant cell wall; xylanolytic activity consists of the synergy between the endoxylanases (β-1,4-D-xylanaxylanohydrolase, EC 3.2.1.8) and the β -xylosidases (β-D-xyloside xylohydrolase, EC 3.2.1.37) (Santos, Cavalcanti, Bonomo, Santana, & Franco, 2011). The degradation of hemicellulose occurs through xylanolytic action (Sousa et al., 2018).
The cellulases and xylanases can be produced by various types of microorganisms, such as filamentous fungi (Martínez-Ruiz, Tovar-Castro, García, Saucedo-Castañeda, & Favela-Torres, 2018) and bacteria (Cerda, Mejías, Gea, & Sánches, 2017). These microorganisms are used to utilize lignocellulosic residues, feed on the present nutrients and secrete metabolites, among them, enzymes (Pathak, Bhardwaj, & Singh, 2014). Therefore, the objective of the present work was to evaluate the use of the cocoa shell as a substrate and Aspergillus oryzae as a fermentation agent. The response surface methodology (MSR) was applied as a statistical tool, for it allows prediction of the best condition of each variable and the effects of interactions between independent variables.
Samples of cocoa shells were supplied by a chocolate factory located in the city of Ilhéus (State Bahia, Brazil). The cocoa shell residue was ground in a Willye-type mill to a particle size of 2 mm.
The Aspergillus oryzae ATCC 10124 was obtained from the collection of microorganisms of the National Institute of Quality Control in Health (INCQS) of the Osvaldo Cruz Foundation (Fiocruz, Manguinhos, State Rio de Janeiro, Brazil) under registration number 01771. This fungus was preserved in silica and glycerol medium at 80ºC in an ultra-freezer. The strains were harvested in agar-dextrose-potato (PDA) culture medium in Petri dishes, previously sterilized in a vertical autoclave at 121ºC for 15 min. The inoculated plates were incubated in a BOD germination chamber at a controlled temperature of 25°C for 7 days or until abundant sporulation (observed visibly due to colony green coloration) and then stored at 4°C for later use.
Preparation of the spore suspension was carried out using the microorganism cultured in PDA medium for 7 days at 25°C. After the incubation period, the sporulated culture was scraped with glass beads and suspended in Tween 80 (0.01%, v v-1) solution, both sterilized in a vertical autoclave at 121°C for 15 min. The suspension was collected in an Erlenmeyer flask, and a 0.1 mL aliquot was taken and diluted in a test tube to count the number of spores in a binocular microscope using a Neubauer chamber.
The fermentations were carried out in an incubator environment using 125 mL Erlenmeyer flasks as a bioreactor and cocoa shell as the substrate. Each flask containing 10 g of residue was sterilized in a vertical autoclave at 121°C for 15 min. The material was then placed in a biological safety cabinet under ultraviolet light until it cooled. An amount of 108 spores per gram of substrate and sterilized distilled water was added to the medium until the desired moisture content was reached. In order to evaluate the influence of the independent variables of water activity, incubation temperature (°C) and fermentation time (hour) on the production of endoglucanase and xylanase, the Box-Behnken design was applied (Table 1). For this planning, a matrix with the description of 15 experiments was used, three of them being the triplicate of the central point to estimate the experimental error, to analyse the quadratic, linear effects and interactions among the variables to fit a second-order model with quadratic terms. The activities of endoglucanases (UI g-1) and xylanases (UI g-1) were applied as response values, and the water activity factors (Aw) incubation temperature (°C) and fermentation time (hour), were analysed as independent variables.
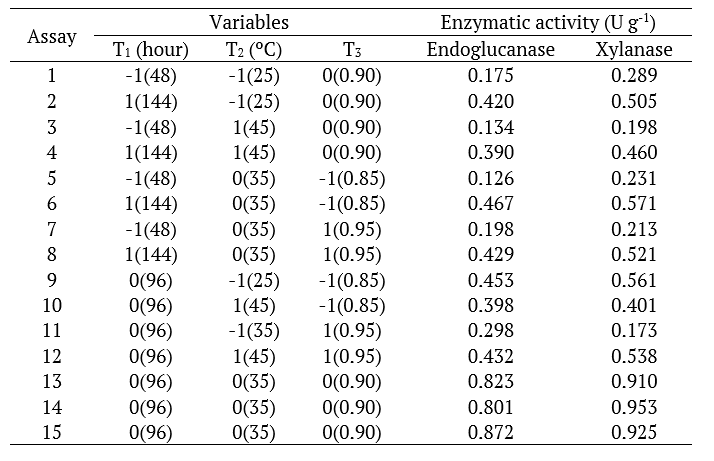
Table 1.
Box-Behnken type matrix for the description of the time factors (T1, hour), temperature (T2, °C) and water activity (T3) and on the activity of endoglucanase and xylanase, produced by A. oryzae ATCC 10124 cultivated in the shell of the almond of cocoa.
by A. oryzae ATCC 10124 cultivated in the shell of the almond of cocoa.
After the fermentation step, 5 mL of distilled water was added to the fermented media for each 1 g of residue. The blends were maintained in an orbital shaker incubator (Shaker) for 20 min. at 170 rpm and 35°C. The entire contents of each flask were pressed through the gauze to separate the solid from the enzyme extract, and the filtrate was collected in a Falcon tube and centrifuged at 3000 rpm for 10 min. The enzymatic assays were performed using the supernatant, which was called the crude multienzyme extract (CME).
Enzyme activity assays were conducted based on standard procedures recommended by IUPAC (Ghose & Bisaria, 1987). The activity of xylanase was determined by the amount of reducing sugars released from the incubation of 1 mL of beechwood xylan (Sigma®) at a concentration of 1% (w v-1) in sodium citrate buffer (0.05 M, pH 4.8) with 1 mL of the crude enzyme extract (previously diluted) in a water bath at 50°C for 10 min. To control the reaction, 1 mL of sodium citrate buffer was incubated with 1 mL of the crude enzyme extract, while the reaction blank consisted of 1 mL of sodium citrate buffer and 1 mL of the xylan solution. After the incubation period, the reducing sugars were dosed based on the method of Miller (1959), adding 2 mL of 3,5 dinitrosalicylic acid (DNS) to the reaction media followed by incubation in boiling water for 5 minutes and subsequent cooling in running tap water. After dilution with 6 mL of distilled water, the absorbance of the samples was read in a spectrophotometer at 540 nm, and the amount of reducing sugar released was quantified using xylose as standard (1 to 10 μmol mL-1). An international unit of enzyme activity (IU) was defined as the amount of enzyme capable of releasing 1 μmol of reducing sugars per minute at 50°C. The results of these analyses were expressed in international units per gram of substrate (UI g-1).
Enzyme activity assays were conducted based on standard procedures recommended by IUPAC (Ghose & Bisaria, 1987). The endoglucanase activity was determined by the amount of reducing sugars released from the incubation of 0.1 mL of carboxymethylcellulose (CMC) at a concentration of 1% (w v-1), with 0.5 mL of the crude enzyme extract in a water bath at 50°C for 15 min. To control the reaction, 0.1 mL and sodium acetate buffer were incubated with 0.5 mL crude (previously diluted) enzyme extract. After the incubation period, the reducing sugars were dosed based on the method of Miller (1959), adding 0.2 mL of 3,5-dinitrosalicylic acid (DNS) to the reaction media, followed by incubation in boiling water for 5 minutes and subsequent cooling. After dilution with 6 mL of distilled water, the absorbance of the samples was read in a spectrophotometer at wavelength at 540 nm and the amount of reducing sugar released was quantified using standard glucose curve. An international unit of enzyme activity (IU) was defined as the amount of enzyme capable of releasing 1 μmol of reducing sugars per minute at 50°C. The results of these analyses were expressed in international units per gram of substrate (UI g-1).
Scanning electron microscopy analysis was performed at the Electronic Microscopy Center (CME) of the State University of Santa Cruz (UESC-Bahia) using a scanning electron microscope Quanta 250, FEI Companye. Cocoa shell, fermented cocoa shell and Aspergillus oryzae colonies were fixed on ‘Stub’ stands. The readings were performed in an ambient mode that is used to investigate samples incompatible with high vacuum and therefore impossible to be analysed by other traditional methods of scanning electron microscopy. Electromicrographs were performed using magnification of 1000 and 3000 x.
The statistical significance level of the generated model was evaluated by analysis of variance (ANOVA, p < 0.05), the regression coefficients were evaluated from the standardized effects (based on Student's t-test, p < 0.05) and the quality of the model evaluated by the coefficient of determination (R2), adjusted coefficient of determination (R2-adj) and lack of fit test. Statistica version 10 software (Statsoft) was used for data analysis and graphing.
The matrix of the experimental design and the corresponding results of the enzymatic activities (U g-1) are shown in Table 1. The solid-state fermentation performed by Aspergillus orizae ATCC 10124 using residue of cocoa shell and water as the culture medium resulted in the production of a crude enzymatic extract with significant activities of endoglucanases and xylanases. According to Gomi (2014), Aspergillus oryzae develops in a solid matrix with Aw greater than 0.8 under different temperature conditions (between 32 and 36ºC).
The data available in Table 1 were submitted to regression analysis for the construction of a mathematical prediction model. The statistical significance of the quadratic, linear and interaction terms (linear x linear) was plotted as a Pareto plot. From the Pareto graphs (Figure 1) it is possible to note that when admitted to a 95% confidence level the relations between the variables are represented and demonstrated through the vertical line drawn.
The results demonstrate that the quadratic terms had a significant effect, indicating the influence of the variables investigated in the production of each enzyme, being the time the independent variable of greater influence in the enzymatic production.
Using the Pareto graphs, it was possible to observe that the models presented non-significant terms, and the regression coefficients were removed for each non-significant term observed, and new reduced models were constructed by regression analysis and submitted to ANOVA (p < 0.05) for endoglucanase and xylanase (Table 2).
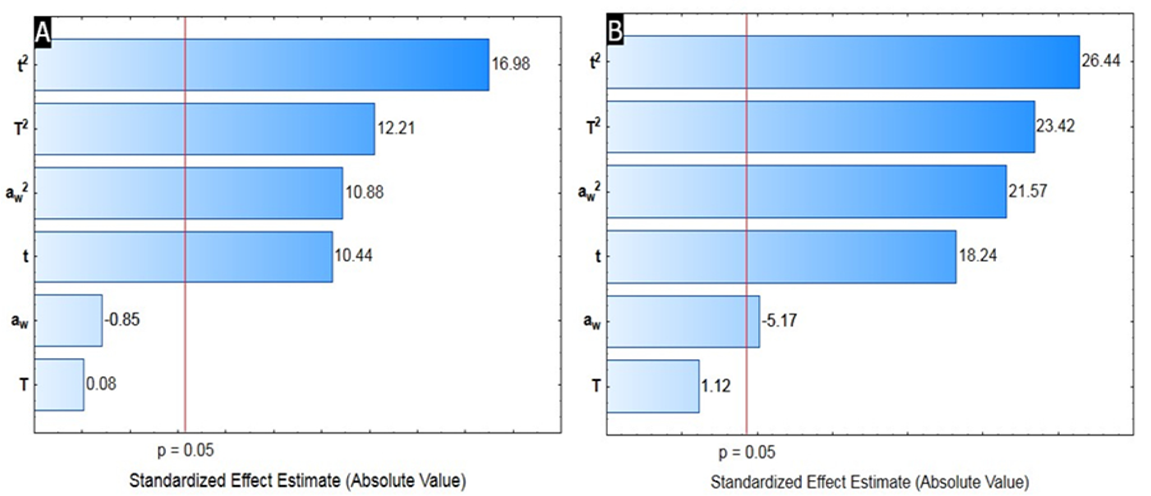
Figure 1.
Pareto chart showing the significance of the terms of the variables: time of fermentation (t), water activity (Aw) and incubation temperature (T), in the model adjusted for production of endoglucanase (A) and xylanase (B) by A. oryzae ATCC 10124.
by A. oryzae ATCC 10124.
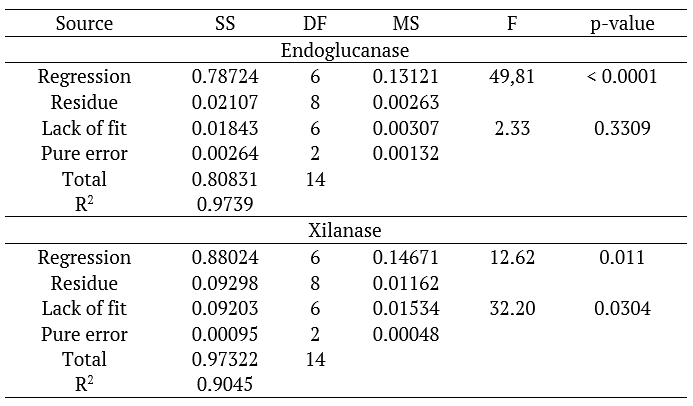
Table 2.
ANOVA for the adjusted reduced model.
According to Ferraz et al. (2018), from ANOVA one can affirm the significance of the parameters through evaluation of the P value, or descriptive level of the test. At the 95% confidence level, a P value for the regression of less than 0.05 suggests that the model is statistically significant. The quality of the models can be verified through the coefficient of determination (R2) and the significance of the lack of fit. The value of R2 is interpreted as the ratio of total variation of the dependent variable (enzymatic activities) that is explained by the variation of the independent variables (temperature, Aw and time) (Santos et al., 2017).
For the models described in Table 2, R2 values for endoglucanase (0.9739) and xylanase (0.9045), indicated that the models fitted well to the experimental results. In mathematical models adjusted for the endoglucanase and xylanase enzymes, with R2 0.9739 and 0.9045, respectively, it means that 97.39 and 90.45% of the activity responses of these enzymes were explained by the adopted model.
The values of lack of fit were insignificant for the endoglucanase model (p = 0.3309), but in the mathematical model adjusted for xylanase production, it can be observed that the lack of fit was significant (p = 0.0304). The lack of adjustment observed at first would render mathematical models invalid for prediction. However, Coutinho et al. (2015) stated that these values can be treated as ‘false positives’ when very low variations are observed in the central point of the study variables, resulting in a small ‘pure error’ value.
The model adopted for xylanase was considered valid, since similar conditions have already been described in the literature, where mathematical models for optimization were used, even with a lack of significant adjustment (Ballus, Meinhart, Bruns, & Godoy, 2011).
Equation 1 and 2 express the second-order quadratic reduced models constructed for endoglucanase and xylanase.
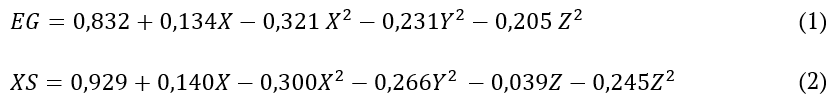
Equation 1 and 2
where:
(EG) represents the endoglucanase activity and (XS) xylanase activity. The independent variables temperature, T (Z), water activity, Aw (Y) and time, t (X) are in coded values, respectively.
The graphics of response surfaces and contour arranged in Figure 2 and 3 of each model adopted enable illustration of the effects of the variables and their interactions.
The response surface graphs of endoglucanase indicated that the variable time of fermentation was the independent variable with the greatest influence. In the study of optimization of the fermentation time for enzymatic production, it can be observed that for the endoglucanases and xylanases, a period of 96 hours in the fermentative process made possible a better enzymatic synthesis (greater production), being identified after this time a decrease in the activities of these enzymes.
Reis et al. (2019) using Aspergillus niger fungus ATCC 1004 under SSF in forage palm bran found a maximum xylanase production at the 96 hour (35.23 U mL-1) fermentation time and endoglucanase time of 72 hours (3.35 U mL-1). Silva et al. (2018) using Botrytis ricini sob SSF in sugarcane bagasse observed the highest enzymatic production (1289.83U mL-1) in 144 hours of fermentation. Ferraz et al. (2018) employing Penicillium roqueforti ATCC 10110 in the SSF process in caja peels recorded a xylanolytic activity value equal to 14 IU g-1 in 120 hours. Brito et al. (2017) using Aspergillus oryzae as a fermentation agent in rice peels and peanut peels obtained 141.5 hours the highest production of endoglucanase in rice peels (17.7 U g-1), while in peanut peels in 196 hours, the activity value was equal to 41.8 U g-1.
Santos et al. (2016) affirm that the fermentation time is a significant variable, for in the beginning of the fermentative process, the solid matrix presents a great amount of nutrients that is being reduced as the fungus develops, promoting nutrient depletion and a consequent decrease in the production of enzymes.
By means of micromorphological analysis (Figure 4), it can be observed how Aspergillus oryzae developed under the cocoa shell during the fermentation time of 96 hours.
The electromicrographs (Figure 4A) show the cocoa shell meal (substrate), the colonies of Aspergillus oryzae which was the bioconversion agent (Figure 4B) and the growth of Aspergillus oryzae ATCC 10124 under bran of the cocoa shell (Figure 4C). Species belonging to the genus Aspergillus typically produce ‘aspegillum’ or ‘conidial head’, as seen on electromyography (Figure 4D), that have a stem that ends in a rounded vesicle containing one or two layers of specialized cells and metastasis), demonstrating that Aspergillus oryzae grew under the analysed solid matrix, during the fermentation time statistically indicated as ideal for development, and it can be affirmed that possibly during the fermentation process, the filamentous fungus excreted the enzymes quantified in the study.
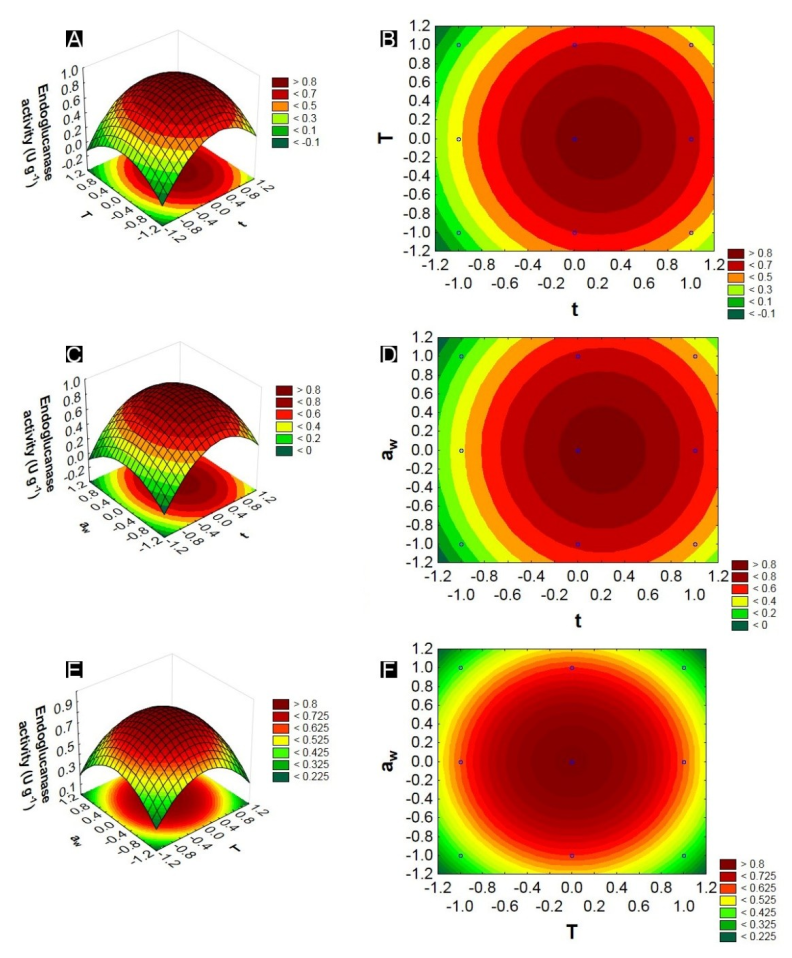
Figure 2.
Response and contour surface graphs constructed based on the model representing the interactions between the variables: water activity and fermentation time, with endoglucanase activity as a response.
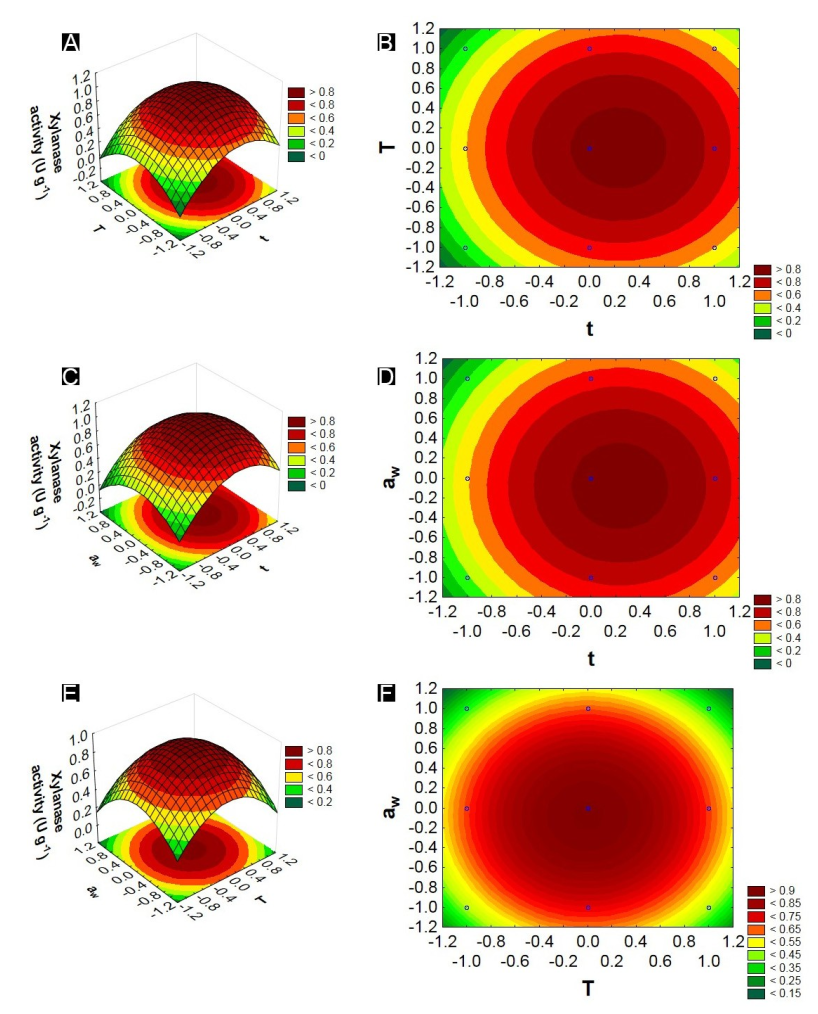
Figure 3.
Response and contour surface graphs constructed based on the EG model representing the interactions between the variables: incubation temperature, water activity and fermentation time, with xylanase activity as a response.
Temperature is another independent variable to be considered in the fermentation process. In this study, the temperature indicated as the best for the production of endoglucanase and xylanase was around the maximum level (35ºC). Similar temperatures for the production of xylanases were found by Marques et al. (2017) using Penicillium roqueforti and Santos et al. (2012) using Aspergillus niger. Incubation temperatures below 20ºC in the bioprocesses may lead to a reduction of the enzymatic production, which may be associated with a decrease in microbial metabolism (Santos et al., 2018). Pandey (2003) states that spore germination, cell growth and product formation are directly influenced by temperature. However, at temperatures above 45°C the microorganisms synthesize only a small amount of proteins required for their development (Pal & Khanum, 2010).
The variable Aw for the production of endoglucanase and xylanase had the centre point 0 (0.9) as the region of maximum production of these enzymes. According to Gomi (2014), Aspergillus oryzae is a fungus that develops well in an average Aw around 0.8; therefore, Aw is an important parameter to consider in the fermentation process. Ferreira et al. (2017) mentioned that the low Aw interferes with microbial growth due to the decrease in nutrient solubility of the solid matrix, the low degree of wettability and the higher water stress.

Figure 4.
Electromyrographs of the growth of Aspergillus oryzae under the cocoa shell: A) Cocoa shell meal; B) Aspergillus oryzae conidia; C) Aspergillus oryzae hyphae; D) Aspergillus oryzae conidial head.
From the study, it was possible to obtain a crude multienzymatic extract with endoglucanases and xylanases from Aspergillus oryzae ATCC 10124 under solid-state fermentation using as solid matrix the shell of the cocoas without any type of supplementation, serving as the only source of energy for fungal growth. The application of the Box-Behnken design was able to verify the best conditions of the investigated variables temperature, aw, and fermentation time for the production of endoglucanases and xylanases, optimizing the process.
The authors would like to thank Banco do Nordeste do Brasil (BNB, Brazil), the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (Capes), the Desenvolvimento Científico e Tecnológico (Bolsas de Produtividade em Pesquisa 302259/2018-0, CNPq, Brazil) and the Fundação de Amparo à Pesquisa do Estado da Bahia (Fapesb, Brazil) for their financial support.
marcelofranco@globo.com
Table 1.
Box-Behnken type matrix for the description of the time factors (T1, hour), temperature (T2, °C) and water activity (T3) and on the activity of endoglucanase and xylanase, produced by A. oryzae ATCC 10124 cultivated in the shell of the almond of cocoa.
by A. oryzae ATCC 10124 cultivated in the shell of the almond of cocoa.
Figure 1.
Pareto chart showing the significance of the terms of the variables: time of fermentation (t), water activity (Aw) and incubation temperature (T), in the model adjusted for production of endoglucanase (A) and xylanase (B) by A. oryzae ATCC 10124.
by A. oryzae ATCC 10124.
Table 2.
ANOVA for the adjusted reduced model.
Equation 1 and 2
Figure 2.
Response and contour surface graphs constructed based on the model representing the interactions between the variables: water activity and fermentation time, with endoglucanase activity as a response.
Figure 3.
Response and contour surface graphs constructed based on the EG model representing the interactions between the variables: incubation temperature, water activity and fermentation time, with xylanase activity as a response.
Figure 4.
Electromyrographs of the growth of Aspergillus oryzae under the cocoa shell: A) Cocoa shell meal; B) Aspergillus oryzae conidia; C) Aspergillus oryzae hyphae; D) Aspergillus oryzae conidial head.
