Biotecnologia
Production of tacrolimus using coconut oil as an alternative to glucose
Production of tacrolimus using coconut oil as an alternative to glucose
Acta Scientiarum. Technology, vol. 43, e55721, 2021
Universidade Estadual de Maringá
Recepción: 09 Septiembre 2020
Aprobación: 30 Marzo 2021
Abstract: Tacrolimus is an immunosuppressant used in the treatment of people who have undergone organ and tissue transplants, as well as in the treatment of dermatoses and eye diseases. Strategies have been carried out to increase the productivity of tacrolimus, since many compounds have in their composition tacrolimus precursors, and bacteria Streptomyces depend on a carbon source for their growth. One strategy is to change the carbon source of the fermentation medium. The present study aims to evaluate the use of coconut oil for the production of tacrolimus via Streptomyces tsukubaensis, as an alternative in the face of glucose, a traditional source of carbon. The batch fermentation process was done in an orbital shaker at 28°C and 130 rpm. Quantification of tacrolimus was performed by High Performance Liquid Chromatography (HPLC). The sugars and proteins, present in the medium, were measured from Somogyi-Nelson and Bradford methods, respectively. According to the results, coconut oil achieved the production of tacrolimus higher than glucose. This is due to the presence of a portion of monounsaturated and polyunsaturated fatty acids that act as precursors to tacrolimus. On the other hand, glucose is a quick carbon source for the bacteria and helps in its growth, but in high amounts it inhibits the production of the drug.
Keywords: Tacrolimus, immunosuppressant, vegetable oils, coconut oil, batch fermentation.
Introduction
Tacrolimus (FK-506) is a lipophilic compound consisting of 23-membered macrolide lactone, with molecular weight of 822 Daltons (Zhang et al., 2020). It was discovered in Japan and isolated in 1984 by researchers of Fujisawa Pharmaceutical Co., during a selection of immunosuppressive activity in compounds isolated from fermentation broths using the bacteria Streptomyces tsukubaensis (Nakamura, 2019). In 1987, the first description of this immunosuppressant was published, and it was intended for the treatment of transplants (Kino et al., 1987; Liu et al., 2019). It was found that this drug is also indicated in other clinical cases such as in the treatment of dermatological disorders, atopic dermatitis (Dähnhardt, Bastian, Dähnhardt-Pfeiffer, Buchner, & Fölster-Holst, 2019), and ophthalmic diseases such as conjunctivitis (Erdinest, Ben-Eli, & Solomon, 2019).
The immunosuppression protocols recommend the use of FK-506 and cyclosporine (CsA) accompanied by anti-inflammatory drugs in the treatment of transplanted people (Fan, Chen, Lu, Li, & Zhu, 2019). Prasad et al. (2018) pointed that tacrolimus (TAC) had fewer adverse effects than CsA when used to treat patients with idiopathic nephrotic syndrome. Tacrolimus binds to proteins, and this complex produces the same action and effects described for cyclosporine. However, TAC is approximately 100 times more potent than CsA (Demain & Sanchez, 2009; Dowd, Johnson, & Mariotti, 2017). Despite the large application of FK-506, its production via fermentation is low (Li, Liang, Wang, Ma, & Wen, 2019). Some strategies are used to increase the productivity of this drug, such as the use of mutant bacteria (Li et al., 2019) or the insertion of substances in the fermentation medium that act as precursors of the TAC molecule (Wang et al., 2017; Silva, Ferrari, Moreira, Franco, & Cremasco, 2019; Moreira, Silva, & Cremasco, 2020).
Tacrolimus (Figure 1) is a secondary metabolite produced from the fermentation of various bacteria of the genus Streptomyces, in particular S. tsukubaensis (Omura, 2002). Therefore, the definition of the sources of carbon, nitrogen, and phosphate used in the preparation of the culture media for fermentation is essential to enable bacterial growth and, as a consequence, to potentiate the production of TAC. The carbon sources normally used are glucose, glycerol, and vegetable oils (Wang et al., 2017). In the case of nitrogen sources, soy bran or soy peptone is often used (Mishra & Verma, 2012; Silva et al., 2019; Moreira et al., 2020), while the sources of phosphates are monopotassium phosphate (KH2PO4) and dipotassium phosphate (K2HPO4) (Turlo et al., 2012; Wang et al., 2017;Silva et al., 2019; Moreira et al., 2020).
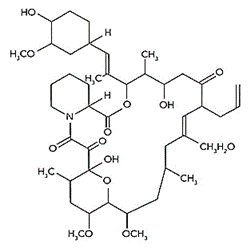
Figure 1. Structural formula of FK-506 (White, Lawson, Masters, & McLaughlin, 2016).
The capacity of Streptomyces tsukubaensis to metabolize different sources of carbon, including vegetable oils, is reported in the Literature (Martínez-Castro et al., 2013). Coconut (Cocos nucifera) is one of the most important foods in tropical and subtropical countries. In these places, milk and oil from coconut are used in daily life for various purposes (Lima & Block, 2019). In recent years, blogs, videos on the internet, and articles encourage the consumption of coconut oil (CO), claiming that this product can bring health benefits such as reducing the risk of cardiovascular disease, weight loss, improving cognitive functions (Lima & Block, 2019; Santos, Howell, Earnest, & Teixeira, 2019). Coconut oil is mainly composed of lauric fatty acid (C12: 0), in addition to other compounds such as caproic acid (C6: 0), caprylic acid (C8: 0), myristic (C14: 0), palmitic (C16: 0), and oleic (C18: 1) (Lima & Block, 2019). It is mentioned in the Literature that oils with the presence of unsaturated fatty acids assist in the production of FK-506 (Mishra & Verma, 2012; Órdóñes-Robles, Santos-Beneit, & Martín, 2018).
The goal of the present study is to evaluate the production of FK-506 from fermentation via Streptomyces tsukubaensis using coconut oil as an alternative to glucose as a carbon source in fermentation medium. In addition, to evaluate the generation of biomass and proteins, the consumption of sugar, and tacrolimus specific production.
Material and methods
Bacterial culture
The S. tsukubaensis strain (DSM 42081) was acquired by the Process and Mass Transfer Laboratory (LPMP), Department of Process Engineering (DEPro), School of Chemical Engineering (FEQ), University of Campinas (UNICAMP) from Leibniz DSMZ Institute - German Collection of Microorganisms and Cell Cultures. For the correct storage of the strain, it was sent to the Pluridisciplinary Center for Chemical, Biological and Agricultural Research (CPQBA) of UNICAMP, being found under the acronym CBMAI - 1832. The microorganism is the same strain from the bacteria used by Kino et al. (1987).
The bacteria were cultivated at the Laboratory of Biochemical Engineering, Biorefining and Products of Renewable Origin (LEBBPOR/DEPro/FEQ/UNICAMP). The seed culture used for bacterial growth in Petri dishes was recommended by DSMZ. The compounds of the seed culture are described in (Table 1).
All solid reagents, except agar, were solubilized in 1000 mL of distilled water, and pH adjusted to 7.2 (Silva et al., 2019; Moreira et al., 2020). The agar was added to the flask, and the mixture was heated to ensure the solubilization of all compounds. The mixture was sterilized in autoclave at 120°C for 20 minutes and placed in Petri dishes until 1 cm high. After cooling and the medium solidified, the bacteria were inoculated with the aid of an inoculation loop. Finally, the dishes were sealed and kept in an orbital shaker for 10 days at 28°C (Silva et al., 2019; Moreira et al., 2020), to promote bacterial growth in the solid medium.

Conditions for pre-inoculum and inoculum
The pre-inoculation, inoculation, and fermentation steps were also developed at LEBBPOR. The composition of the liquid medium used for pre-inoculation and inoculation was described by Turlo et al. (2012), Silva et al. (2019), and Moreira et al. (2020), which is listed in (Table 2).

The reagents described in (Table 2) were solubilized in 1000 mL of distilled water. The pH was corrected to 7.2, and the medium was sterilized for 20 minutes at 120°C. To prepare the pre-inoculum, 100 mL of the liquid medium was transferred to three 250 mL flasks, with the bacterial loop aseptically placed in only 2 flasks, and the third was used as control of the process. The flasks were sent to the orbital shaker (Shel Lab USA), and stored for 24 hours at 28°C and 130 rpm (Moreira et al., 2020). After 24 hours, 1 mL aliquots were collected from each pre-inoculated flask to check for bacterial growth. The verification of bacterial growth was performed through the absorbance (OD600> 0.6) of the samples in the UV-Vis spectrophotometer of the IMPLEN brand at 600 nm wavelength (Turlo et al., 2012). Once the absorbance was reached, the inoculation process was continued.
For the inoculation process, three flasks with 500 mL capacity were used, one of them was used for control. The two other flasks had 180 mL of the liquid medium and 20 mL of the pre-inoculum. The flasks were kept in the orbital shaker under the same conditions of temperature, rotation, and time in the pre-inoculation stage. The absorbance measurement was performed, keeping the same criteria described during the preparation of the pre-inoculum. The highest absorbance flask in the inoculation stage was used for fermentation.
Conditions for fermentation
The composition of the fermentation medium was based on the works of Turlo et al. (2012), Silva et al. (2019), and Moreira et al. (2020). Turlo et al. (2012) used the disaccharide maltose as carbon source and picolinic acid as additive. In the present study, two culture media were used. The difference is the carbon source, glucose or coconut oil, and no additive was used. Since there was a change in the carbon source, it was necessary to adapt the concentration used, which was based on Singh and Behera (2009). The composition of the culture media is described in (Table 3).
The solid reagents in (Table 3) were solubilized in 1000 mL of distilled water, and the pH adjusted to 7.2. Then, the culture media were sterilized in the autoclave for 20 minutes and 120°C. The volume of 225 mL of sterilized liquid medium and 25 mL of the inoculum was added in 2 flasks. The batch fermentation process was carried out in a shaker at 28°C and 130 rpm for 7 days. The definition of fermentation temperature was based on the work of Mishra and Verma (2012), which used 28°C. According to Moreira et al. (2020), FK-506 production via Streptomyces tsukubaensis is favored at 28°C. The fermentation time was based on the study of Silva et al. (2019). Every 24 hours, 10 mL samples were collected from each flask to quantify the biomass and the drug produced during the process. The fermentation was carried out in duplicate.
Preparation of samples and quantification of biomass
In each sample collected, 10 mL of acetone was added to interrupt the batch fermentation process and to extract the intracellular FK-506 (Turlo et al., 2012; Zhang et al., 2020). After 12 hours of rest, the samples were filtered on INLAB type 10 filter paper, pore diameter equal to 3 μm, with the aid of a vacuum pump (EOS, model VE18010) (Silva et al., 2019; Moreira et al., 2020). After filtration, filter papers with biomass were washed with deionized water and kept at 80°C for 24 hours for drying (drying and sterilization oven model 315 SE). This temperature was based on Silva et al. (2019) and Moreira et al. (2020), which does not alter the properties of the material generated during fermentation. The filter papers were weighed before filtration and after the drying process of biomass. The total dry cell mass was defined as biomass, which was determined by mass difference before and after filtration. The test was performed in duplicate.
Preparation of samples and quantification of FK-506
The filtrate from vacuum filtration was evaporated in rotary evaporator (FISATON 803) until the formation of an oily compound, as observed by Silva et al. (2019) and Moreira et al. (2020). In this material, 3.5 mL of mobile phase, composed of acetonitrile and water (proportion 70:30, v v-1), were added (Silva et al., 2019). Finally, the samples were centrifuged (Hettich 420R Centrifuge) and the supernatant was collected and used for quantification of FK-506 based on Li, Liang, Wang, Ma, and Wen (2019).
Quantification of FK-506 was performed from High Performance Liquid Chromatography (HPLC). The Shimatzu chromatograph was adjusted at 210 nm wavelength (Singh & Behera, 2009; Silva et al., 2019; Moreira et al., 2020). Injections of 20 uL were done in chromatography, while the process temperature was maintained at 60°C by thermostatic bath (QUIMIS - Q214M2), and flow rate equal to 1 mL min-1. The mobile phase was composed of acetonitrile and water in 70:30 (v v-1) ratio, and the column used as a stationary phase was C18 column (ThermoQuest Hyperbond 300 x 3.9 mm). The test was performed in duplicate.
The analytical curve for the quantification of FK-506 was prepared by adding FK-506 standard in 3.5 mL of the mobile phase. The FK-506 standard was purchased from Sigma-Aldrich (CAS 109581-93-3, purity ≥ 98%). The previously defined concentrations were 30, 60, 90, 120, and 150 mg L-1. The analytical curve showed a coefficient of determination equal to 0.9999.
Reducing sugars quantification
For the determination of reducing sugars, the Somogyi-Nelson method was applied according to Nelson (1944) and Somogyi (1945). To perform this method, Somogyi-Nelson I and II reagents (SN-I and SN-II) must be done. The compounds used to prepare the reagents SN-I and SN-II and their respective concentrations are shown in (Table 4) and (Table 5).

*mL
With SN-I and SN-II reagents ready, the Somogyi-Nelson method was started. First, 1 mL of the sample was collected during fermentation, and after added 2 mL of SN-I reagent. The samples were agitated in a vortex mixer laboratory, heated in water bath for 6 minutes, and cooled in ice bath for 5 minutes. 2 mL of the SN-II reagent were added to each test tube, which was agitated and left to stand for 5 minutes. Finally, 25 mL of distilled water were added to the samples, and the absorbances were read on a UV-Vis spectrophotometer with wavelength of 540 nm. The test was performed in duplicate. The analytical curve of the Somogyi- Nelson method was done from a standard glucose solution. The previously defined concentrations were 50, 100, 200, 300, 400, and 500 mg L-1.
Protein quantification
In order to quantify the proteins, the Bradford method was applied according to Bradford (1976). First, 100 μL of the sample was pipetted into test tubes. Subsequently, 100 μL of NaOH (1N) and 2.5 mL of Bradford's reagent were added to the samples. The mixtures were agitated in a vortex mixer laboratory and left for 20 minutes. Finally, the absorbances of each sample were read on the UV-Vis spectrophotometer at 595 nm wavelength. The analytical curve was done from standard bovine serum albumin (BSA) solution. The previously defined concentrations were 6.25, 12.5, 25, 50 and 100 mg L-1. The test was performed in duplicate.
Results and discussion
FK-506, biomass and specific production of tacrolimus
The aim of the present work was to evaluate two primary sources of carbon and to find which was the most suitable for the production of FK-506 via Streptomyces tsukubaensis. (Figure 2) and (Figure 3) show the production of FK-506 (mg L-1), the concentration of biomass (g L-1), and the specific production of the drug (mg g-1), that is, the relation between the production of the drug (mg L-1) by the concentration of biomass (g L-1), using coconut oil and glucose as a carbon source in the fermentation media, respectively. The highest concentration of the drug using coconut oil occurred at 96 hours and presented 31.35 mg L-1 (Figure 2). In the case of glucose as a carbon source, the FK-506 highest concentration occurred at 120 hours, with 22.88 mg L-1 (Figure 3). The maximum FK-506 production from Streptomyces tsukubaensis fermentation, found in Literature, reports time between 96 and 192 hours (Singh & Behera, 2009; Wang et al., 2017; Li et al., 2019; Moreira et al., 2020).
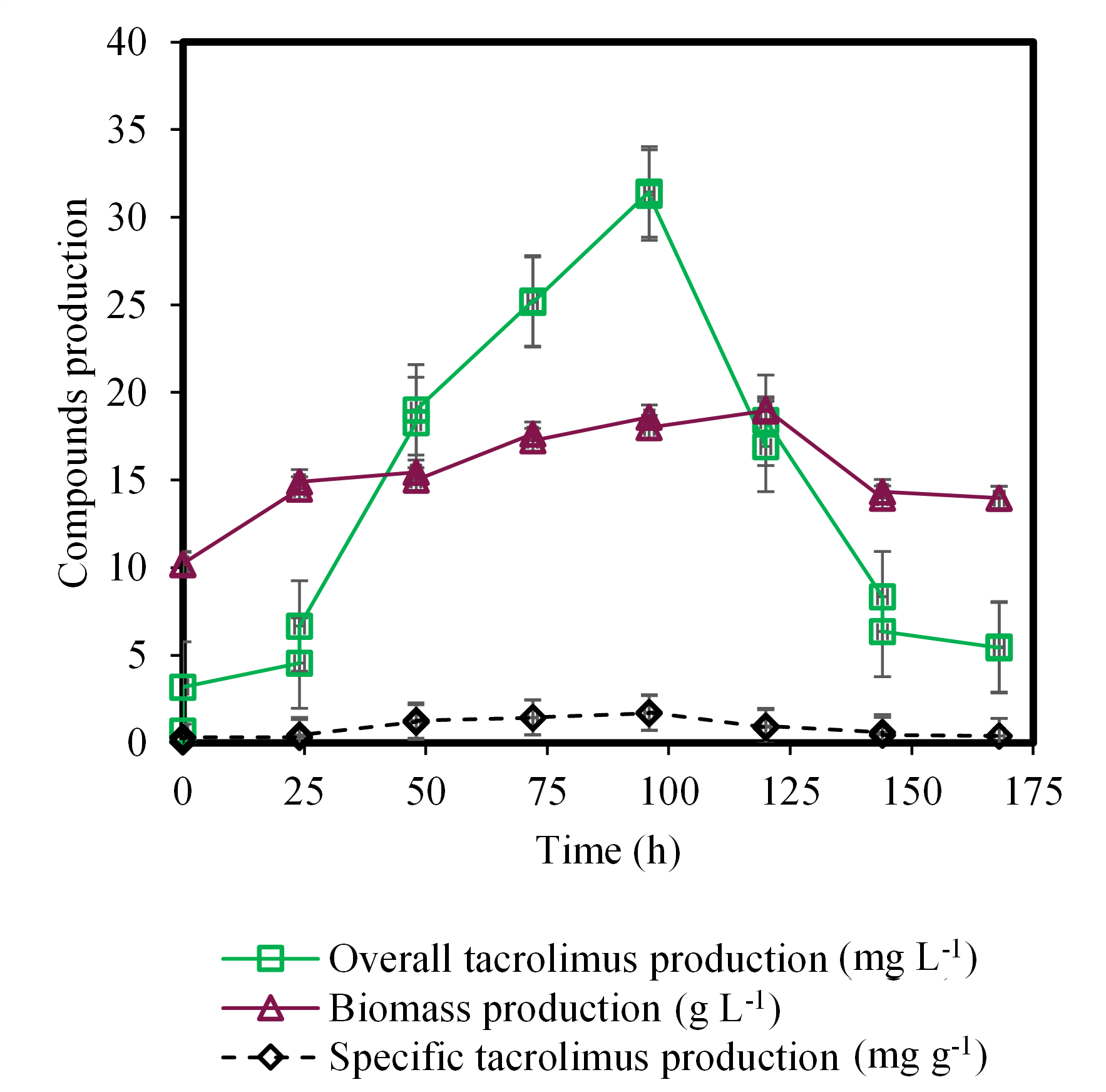
Figure 2. Fermentation process for production of tacrolimus using coconut oil as a carbon source.
The biomass production from fermentation with coconut oil in 96 hours was equal to 18.60 g L-1, while using glucose, in 120 hours, was 12.14 g L-1. Considering the specific production of FK-506, it is possible to return to (Figure 2) and (Figure 3). (Figure 2) shows the FK-506 specific production equal to 1.88 mg/g, while (Figure 3), this value is 1.69 mg g-1, showing the best performance of coconut oil, as a carbon source, compared with glucose. Martínez-Castro et al. (2013), using the combination of starch and glucose as a carbon source, obtained a specific production of FK-506 around 2.8 mg g-1, but the C:N ratio was not disclosed. In the present study, it was not possible to determine the C:N ratio for coconut oil, because both chemical elements are inserted in a complex medium. Mishra and Verma (2012) tested several sources of carbon and nitrogen, and the fermentation medium that had glucose as the primary carbon source resulted in a specific production of 8.70 mg g-1. However, the rotation of the shaker in these studies are different from the present study (130 rpm). Martínez-Castro et al. (2013) used 220 rpm, and Mishra and Verma (2012), 200 rpm. Moreira et al. (2020) shows that agitation is an important variable in the batch fermentation process and may be the cause of the difference between specific productivities among researchers.
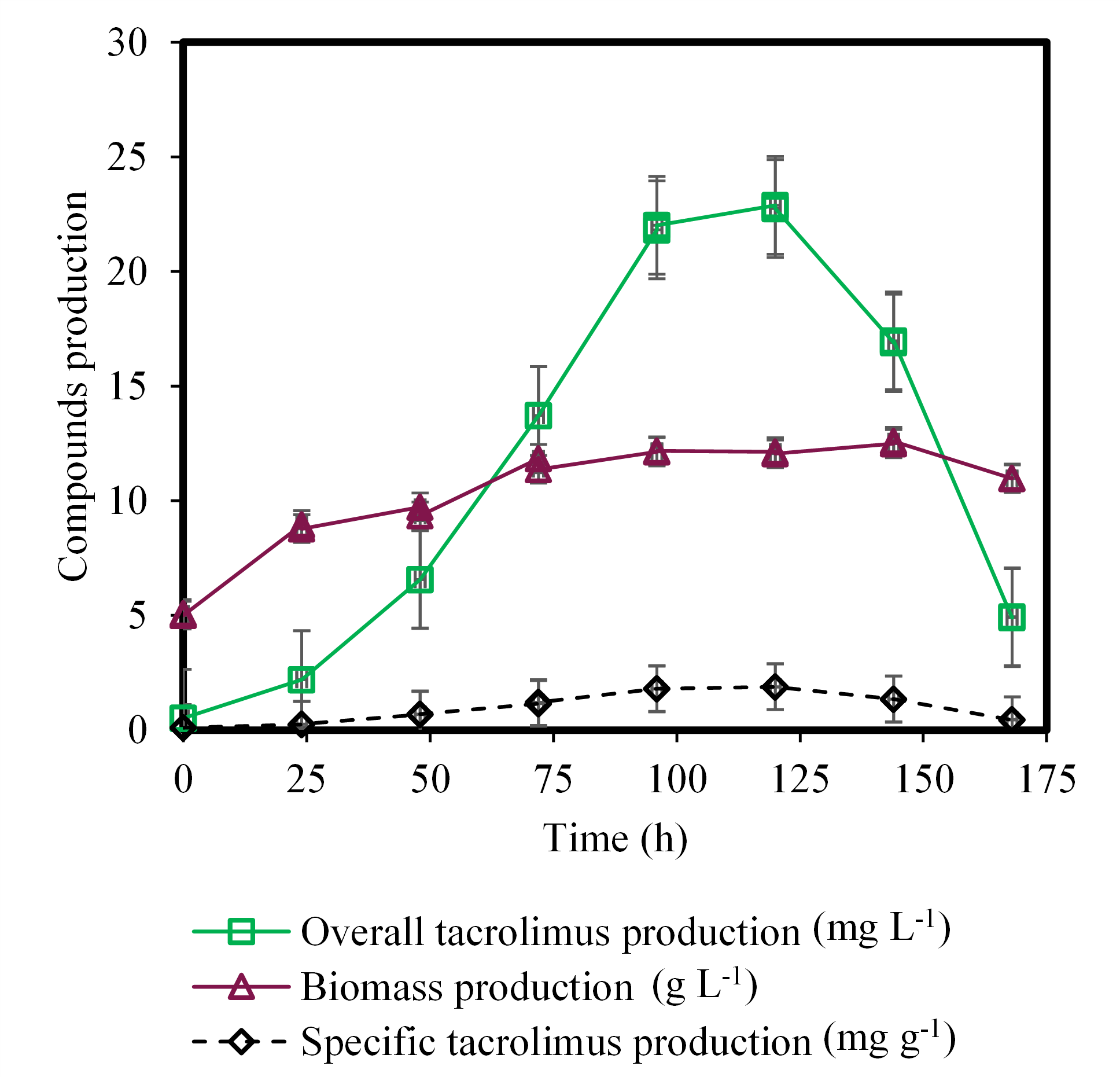
Figure 3. Fermentation process for production of tacrolimus using glucose as a carbon source.
Coconut oil is mainly composed of saturated fatty acids (92%), such as lauric acid, which about 64% correspond to medium chain fatty acids (MCFA) with several carbons ranging from 8 up to 12. They are used by cells as an energy source. Coconut oil comprises lesser amounts of monounsaturated and polyunsaturated fatty acids (Lima & Block, 2019). Glucose is a great source of carbon for bacterial growth, however, large amounts can negatively affect the production of secondary metabolites such as FK-506 (Demain, 1976; Poshekhontseva, Fokina, Sukhodolskaya, Shutov, & Donova, 2019). This effect is characteristic of a mechanism known as carbon catabolic repression (CCR), which consists in systems that suppress genes present in alternative sugars, when they are in a medium that has sugars of preferential use such as glucose (Li et al., 2020). According to Moreira et al. (2020), the glucose in the fermentation media for C:N ratio from 3 to 4 % did not interfere in the cell growth of Streptomyces tsukubaensis.
It is important to mention that coconut oil also has a high concentration of bioactive compounds as vitamin E, sterols, and polyphenols (Gao et al., 2020). Vitamin E comprises eight compounds found in nature, four of them are tocopherols (α-, β-, γ-, δ-tocopherol) and four are tocotrineols (α-, β-, γ-, δ-tocotrienol). The most common and biologically active of this vitamin E isomers is α-tocopherol (C29H50O2). In α-tocopherol’s structure, there is a saturated chain, a phenol, and ether group (Fairus, Cheng, & Sundram, 2020). Vitamins are one of the most important micronutrients in biological metabolism, including growth, development, and maintenance. The biological role of vitamin E is principally based on its antioxidant properties, as prevention of lipid peroxidation. The cells are constantly exposed to free radicals that can cause oxidative stress and induce damage to biological structures (Price & Preedy, 2020).
Reducing sugars and proteins
The quantification of proteins and reducing sugars was performed during both fermentations. The evaluation of these parameters is important because these substances are present in the fermented broth, and influence the future purification of the drug. It is reported in the Literature that the amount of proteins increases during FK-506 production, and they are produced and consumed during fermentation (Wang et al., 2017; Silva et al., 2019). The concentration profile of proteins produced in both fermentation media is shown in (Figure 4). In the present study, the rate of protein production was higher than the rate of its consumption. At the beginning of the fermentation with coconut oil, the protein concentration was 91.07 mg L-1 in the culture medium, and 105.17 mg L-1 for glucose. This is explained by the physicochemical characteristics of the corn steep liquor used in the fermentation media. According to Achal, Mukherjee, and Reddy (2010), corn steep liquor is composed of 24% protein and several amino acids, such as arginine (0.4%), cysteine (0.5%), glycine (1.1%), histidine (0.3%), isoleucine (0.3%), isoleucine (0.9%), methionine (0.5%) and minor compounds. These amino acids may have formed more proteins with bacteria aid, and this explains the differences in initial protein concentrations in both fermentation media.
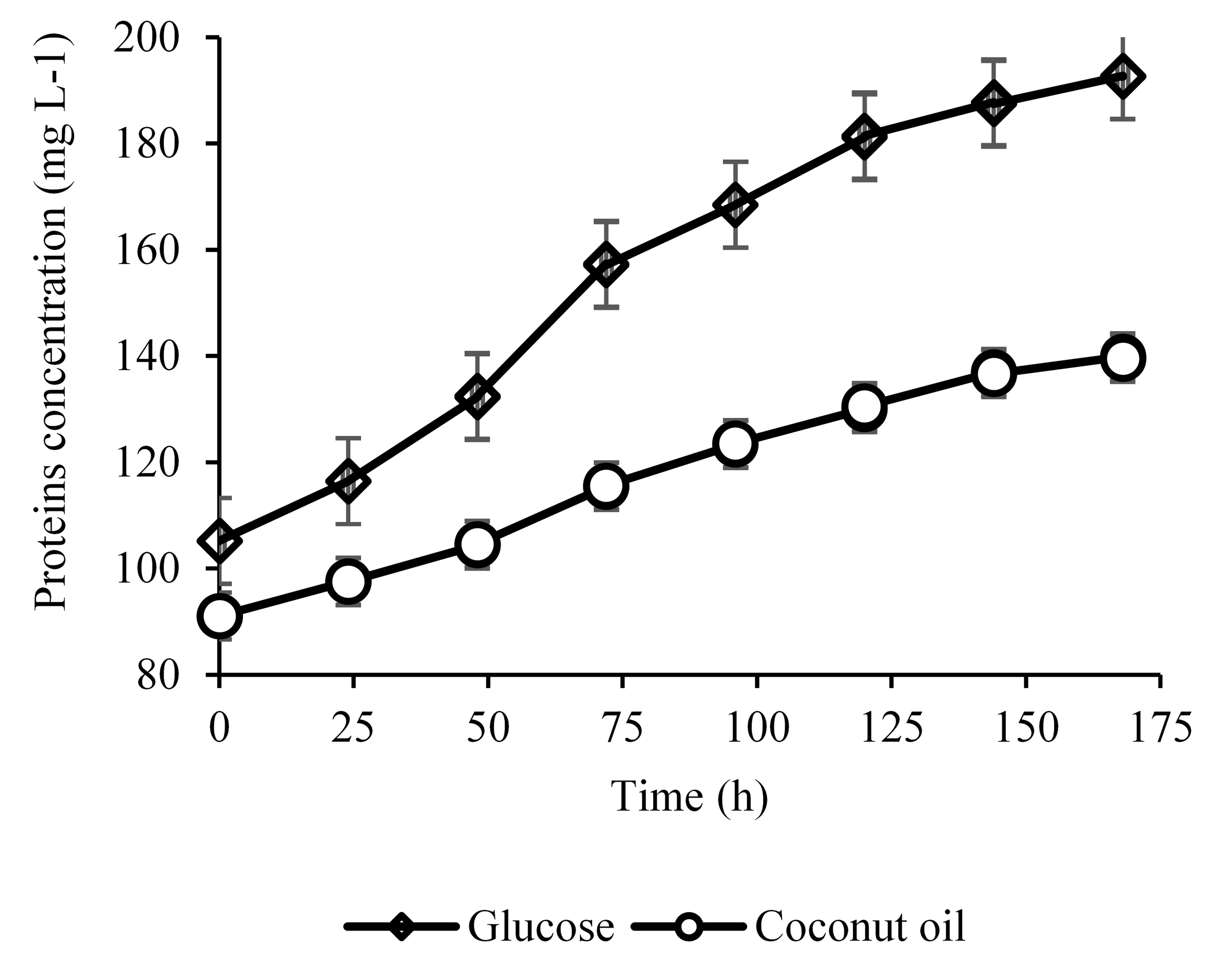
Figure 4. Proteins concentration from glucose and coconut oil as carbon sources.
(Figure 5) shows the profile of the reducing sugars concentration consumed in both fermentation media. The initial concentration of sugars in the fermentation conducted with coconut oil and glucose were respectively 567.59 mg L-1 and 507.33 mg L-1. The sugar concentration in 120 hours for the medium with glucose was 181.16 mg L-1, and for coconut oil, in 96 hours, equal to 123.33 mg L-1. The most pronounced increase in protein production in both fermentations was between 72 and 144 hours. The fermentation with glucose consumed 308.69 mg L-1 of sugar and with coconut oil, this value is 271.23 mg L-1 during the fermentation process. Glucose was consumed more easily compared to coconut oil because this monosaccharide is a fast source of assimilation by the bacteria and is used for the production of cells (Poshekhontseva et al., 2019).
According to (Figure 5), the medium using coconut oils always presents a higher concentration of reducing sugars than medium with glucose during the fermentation process. This is due to the complexity of coconut oil, which makes it difficult for the bacteria to assimilate the nutrients. In general, a large amount of sugar was consumed in the first 72 hours in both fermentation media, corresponding to the exponential phase of bacterial growth. According to Bertrand (2019), during the exponential phase all the cells enter cell division, and they are dependent on the amount of nutrients present in the medium. It is the period of best cell growth being influenced by the culture conditions and genetic characteristics. Hwang, Kim, Charusanti, Palsson, and Lee (2014) mention that the production of secondary metabolites is the response to the stress to which the bacteria was subjected, such as the lack of nutrients.
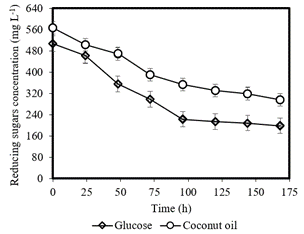
Figure 5. Reducing sugars concentration from glucose and coconut oil as carbon sources.
Conclusion
The use of coconut oil as the carbon source in fermentation via Streptomyces tsukubaensis increased the production of FK-506 (31.35 mg L-1 in 96 hours) compared with glucose (22.88 mg L-1 in 120 hours). In addition, the specific FK-506 production using coconut oil was superior to the fermentation using glucose, being equal to 1.88 mg g-1 and 1.69 mg g-1, respectively. Thus, coconut oil is an option to replace the glucose. This fact can be explained because glucose is a fast source of carbon, that is, the bacteria use this compound for their growth during exponential phase. However, in high concentrations, it acts as inhibitor of secondary metabolites such as FK-506. On the other hand, coconut oil has unsaturated fatty acids in its composition, another factor that contributes to this is the degrees of unsaturation in the structure molecule of the drug.
As expected throughout the batch fermentation process, sugars were consumed and proteins were produced. In the present study, the rate of protein production was higher than the rate of consumption. The amount of proteins secreted by bacteria in the fermented broth interferes in the purification process. These results are very interesting, especially for countries with tropical and subtropical climates, where coconut oil is widely produced.
Acknowledgements
This project was supported by CAPES and CNPQ. The authors would like to thank these Institutions for all their support and financing.
References
Achal, V., Mukherjee, A., & Reddy, M. S. (2010). Biocalcification by Sporosarcina pasteurii using corn steep liquor as nutrient source. Industrial Biotechnology, 6(3), 170-174. DOI: http://doi.org/10.1089/ind.2010.6.170
Bertrand, R. L. (2019). Lag phase is a dynamic, organized, adaptive, and evolvable period that prepares bacteria for cell division. Journal of Bacteriology, 201(7), 1-21. DOI: http://doi.org/10.1128/JB.00697-18
Bradford, M. M. (1976). A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Analytical Biochemistry, 72, 248-254. DOI: http://doi.org/10.1006/abio.1976.9999
Dähnhardt, D., Bastian, M., Dähnhardt-Pfeiffer, S., Buchner, M., & Fölster-Holst, R. (2019). Comparing the effects of proactive treatment with tacrolimus ointment and mometasone furoate on the epidermal barrier structure and ceramide levels of patients with atopic dermatitis. Journal of Dermatological Treatment, 9, 1-9. DOI: http://doi.org/10.1080/09546634.2019.1708240
Demain, A. L. (1976). Genetic regulation of fermentation organisms: fermentation, regulation, antibiotics. Stadler Genetics Symposium Series, 8, 41-55.
Demain, A. L., & Sanches, S. (2009). Microbial drug discovery: 80 years of progress. The Journal of Antibiotics, 62, 5-16. DOI: http://doi.org/10.1038/ja.2008.16
Dowd, F. J., Johnson, B. S., & Mariotti, A. J. (2017). Pharmacology and therapeutics for dentistry [eBook]. St. Louis, MO: Elsevier.
Erdinest, N., Ben-Eli, H., & Solomon, A. (2019). Topical tacrolimus for allergic eye diseases. Current Opinion in Allergy and Clinical Immunology, 19(5), 535-543. DOI: http://doi.org/10.1097/ACI.0000000000000560
Fairus, S., Cheng, H. M., & Sundram, K. (2020). Antioxidant status following postprandial challenge of two different doses of tocopherols and tocotrienols. Journal of Integrative Medicine, 18(1), 68-79. DOI: http://doi.org/10.1016/j.joim.2019.11.005
Fan, J., Chen, L., Lu, X., Li, M., & Zhu, L. (2019). The pharmacokinetic prediction of cyclosporin A after coadministration with wuzhi capsule. AAPS PharmSciTech, 20(6), 2-7. DOI: http://doi.org/10.1208/s12249-019-1444-6
Gao, W., Jiang, Z., Du, X., Zhang, F., Liu, Y., Bai, X., & Sun, G. (2020). Impact of surfactants on nanoemulsions based on fractionated coconut oil: emulsification stability and in vitro digestion. Journal of Oleo Science, 69(3), 227-239. DOI: http://doi.org/10.5650/jos.ess19264
Hwang, K.-S., Kim, H. U., Charusanti, P., Palsson, B. Ø., & Lee, S. Y. (2014). Systems biology and biotechnology of Streptomyces species for the production of secondary metabolites. Biotechnology Advances, 32(2), 255-268. DOI: http://doi.org/10.1016/j.biotechadv.2013.10.008
Kino, T., Hatanaka, H., Hashimoto, M., Nishiyama, M., Goto, T., Okuhara, M., ... Imanaka, H. (1987). FK506, a novel immunosuppressant isolated from a Streptomyces. I. Fermentation, isolation, and physic-chemical and biological characteristics. The Journal of Antibiotics, 40(9), 1249-1255. DOI: http://doi.org/10.7164/antibiotics.40.1249
Li, C., Chen, X., Wen, L., Cheng, Y., An, X., Li, T., … Hou, N. (2020). An enhancement strategy for the biodegradation of high-concentration aliphatic nitriles: utilizing the glucose-mediated carbon catabolite repression mechanism. Environmental Pollution, 265(Part A), 114302. DOI: http://doi.org/10.1016/j.envpol.2020.114302
Li, Y., Liang, S., Wang, J., Ma, D., & Wen, J. (2019). Enhancing the production of tacrolimus by engineering target genes identified in important primary and secondary metabolic pathways and feeding exogenous precursors. Bioprocess and Biosystems Engineering, 42(7), 1081-1098. DOI: http://doi.org/10.1007/s00449-019-02106-9
Lima, R. d. S., & Block, J. M. (2019). Coconut oil: what do we really know about it so far? Food Quality and Safety, 3(2), 61-72. DOI: http://doi.org/10.1093/fqsafe/fyz004
Liu, Y., Zhang, C., Li, L., Ou, B., Yuan, L., Zhang, T., … Peng, Z. (2019). Genome-wide association study of tacrolimus pharmacokinetics identifies novel single nucleotide polymorphisms in the convalescence and stabilization periods of post-transplant liver function. Frontiers in Genetics, 10(528), 1-9. DOI: http://doi.org/10.3389/fgene.2019.00528
Martínez-Castro, M., Salehi-Najafabadi, Z., Romero, F., Pérez-Sanchiz, R., Fernández-Chimeno, R. I, Martín J. F., & Barreiro, C. (2013). Taxonomy and chemically semi-defined media for the analysis of the tacrolimus producer 'Streptomyces tsukubaensis'. Applied Microbial and Cell Physiology, 97, 2139-2152. DOI: http://doi.org/10.1007/s00253-012-4364-x
Mishra, A., & Verma, S. (2012). Optimization of process parameters for tacrolimus production by new isolate of Streptomyces sp. using response surface methodology. Journal of Biochemical Technology, 3(4), 419-425.
Moreira, J. V., Silva, S. C. M., & Cremasco, M. A. (2020). Evaluation of carbon: nitrogen ratio in semi-defined culture medium to tacrolimus biosynthesis by Streptomyces tsukubaensis and the effect on bacterial growth. Biotechnology Reports, 26, e00440. DOI: http://doi.org/10.1016/j.btre.2020.e00440
Nakamura, K. (2019). Tacrolimus (Prograf). In S. Nagaoka. (Ed.), Drug Discovery in Japan (p. 147-167). New York, NY: Springer.
Nelson, N. (1944). A photometric adaptation of the Somogyi method for the determination of glucose. Journal of Biological Chemistry, 153(2), 375-380. DOI: http://doi.org/10.1017/S0009840X00101660
Omura, S. (2002). Macrolide Antibiotics. Cambridge, MA: Academic Press.
Órdóñez-Robles, M., Santos-Beneit, F., & Martín, J. F. (2018). Unraveling nutrional regulation of Tacrolimus biosynthesis in Streptomyces tsukubaensis through omic approaches. Antibiotics, 7(2), 1-19. DOI: http://doi.org/10.3390/antibiotics7020039
Poshekhontseva, V. Y., Fokina, V. V., Sukhodolskaya, G., Shutov., A., & Donova, M. (2019). Effect of starch composition on the biosynthesis of immunosuppressant tacrolimus (FK-506) by Streptomyces tsukubaensis VKM Ac-2618D strain. Applied Biochemistry and Microbiology, 55, 534-543. DOI: http://doi.org/10.1134/S0003683819040148
Prasad, N., Manjunath, R., Rangaswamy, D., Jaiswal, A., Agarwal, V., Bhadauria, D., ... Gupta, A. (2018). Efficacy and safety of cyclosporine versus tacrolimus in steroid and cyclophosphamide resistant nephrotic syndrome: a prospective study. Indian Journal of Nephrology, 28(1), 46-52. DOI: http://doi.org/10.4103/ijn.IJN_240_16
Price, M. Y., & Preedy, V. R. (2020). Reference dietary requirements of vitamins in different stages of life. In V. B. Patel (Ed.), Molecular nutrition vitamins (p. 3-32). Cambridge, MA: Academic Press.
Santos, H. O., Howell, S., Earnest, C. P., & Teixeira, F. J. (2019). Coconut oil intake and its effects on the cardiometabolic profile – a structure literature review. Progress in Cardiovascular Diseases, 62(5), 436-443. DOI: http://doi.org/10.1016/j.pcad.2019.11.001
Silva, S. C. M., Ferrari, W. M., Moreira, J. V., Franco, T. T, & Cremasco, M. A. (2019). Streptomyces tsukubaensis fermentation using Brazil nut oil to enhance tacrolimus production. Journal of Applied Biotechnology Reports, 6(3), 109-112. DOI: http://doi.org/10.29252/JABR.06.03.05
Singh, B. P., & Behera, B. K. (2009). Regulation of tacrolimus production by altering primary source of carbons and amino acids. Letters in Applied Microbiology, 49(2), 254-259. DOI: http://doi.org/10.1111/j.1472-765X.2009.02652.x
Somogyi, M. (1945). A new reagent for the determination of sugar. The Journal of Biological Chemistry, 160, 61-68. DOI: http://doi.org/10.1369/0022155415575032
Turlo, J., Gajzlerska, W., Klimaszewska, M., Król, M., Dawidowski, M., & Gutkowska, B. (2012). Enhancement of tacrolimus productivity in Streptomyces tsukubaensis by the use of novel precursors for biosynthesis. Enzime and Microbial Technology, 51(6-7), 388-395. DOI: http://doi.org/10.1016/j.enzmictec.2012.08.008
Wang, J., Liu, H., Huang, D., Jin, L., Wang, C., & Wen, J. (2017). Comparative proteomic and metabolomic analysis of Streptomyces tsukubaensis reveals the metabolic mechanism of FK506 overproduction by feeding soybean oil. Applied Microbiology and Biotechnology, 101, 2447-2465. DOI: http://doi.org/10.1007/s00253-017-8136-5.
White, D., Lawson, N., Masters, P., & McLaughlin, D. (2016). Clinical Chemistry. New York, NY: Garland Science.
Zhang, J., Liu, Z., Tao, C., Lin, X., Zhang, M., Zeng, L, … Song, H. (2020). Cationic nanoemulsions with prolonged retention time as promising carriers for ophthalmic delivery of tacrolimus. European Journal of Pharmaceutical Sciences, 144, 105229. DOI: http://doi.org/10.1016/j.ejps.2020.105229
Notas de autor
alessandra.suzin.bertan@gmail.com