2025
371
13032024
25092024
 Ana Caroline Damasceno Souza de Sá carolsouza.geo@gmail.com
Ana Caroline Damasceno Souza de Sá carolsouza.geo@gmail.com
Universidade Estadual do Ceará - UECE, Brasil
Lidriana Pinheiro lidriana@ufc.br
Universidade Federal do Ceará - UFC, Brasil
Resumo: Esta pesquisa tem como objetivo espacializar os manguezais e correlacioná-los com variáveis ambientais, a fim de verificar se existe variação na distribuição dos bosques de mangue e apicum em áreas com diferentes padrões de precipitação e períodos de seca dentro de uma mesma região brasileira. Foram utilizadas imagens do satélite CBERS-4A e vetorização manual dos polígonos de bosques de mangues e apicuns no software QGIS v. 3.10.14, além de trabalhos de campo. A análise de Correlação de Pearson, realizada no software RStudio, considerou variáveis como precipitação, meses secos, espécies de mangue, fragmentação e tamanho da área. Os resultados indicam que a variação na precipitação e o número de meses secos têm uma forte influência na distribuição dos manguezais. Por exemplo, no estuário hipersalino Piranhas-Açu (RN), observou-se uma grande fragmentação dos bosques de mangue, com 1.379 polígonos mapeados em uma área de 2.396 ha, recebendo 500 mm de precipitação anual e 8 meses secos. Já nos estuários de Timonha (CE) e Itarema (CE), com 1.000 mm de chuva e 6 meses secos, foram identificados 131 e 118 polígonos, respectivamente, em 2.482 e 1.093 ha. Em contrapartida, o estuário do rio Mamanguape (PB), com a maior precipitação anual de 1.750 mm e apenas 3 meses secos, tem uma distribuição de 4.256 ha em 24 polígonos. Em relação aos apicuns, o Piranhas-Açu (RN) apresenta uma fragmentação de 674 polígonos em uma área de 4.858 ha e o estuário do rio Mamanguape (PB) seis polígonos mapeados em uma área de 2,4 ha. A pesquisa destaca os impactos de variáveis climáticas na distribuição dos manguezais, bem como a interferência antrópica no uso do solo devido a atividades econômicas e os efeitos negativos da fragmentação, ressaltando a importância da conservação e manejo adequado desses ecossistemas vitais.
Palavras-chave: Bosque de mangue, Apicum, Costa semiárida do Brasil, Biogeografia de manguezais.
Abstract: This research aims to spatialize mangroves and correlate them with environmental variables, exploring potential variation in the distribution of mangrove and salt flat forests in relation to areas with different rainfall patterns and drought periods within the same Brazilian region. CBERS-4A satellite images and manual vectorization of mangrove and apicum forest polygons in QGIS v. 3.10.14 software were used, as well as fieldwork. Pearson's correlation analysis, carried out using RStudio software, considered variables such as rainfall, dry months, mangrove species, fragmentation, and area size. The results indicate that variations in rainfall and the number of dry months have a strong influence on the distribution of mangroves. For example, in the Piranhas-Açu hypersaline estuary (RN), a large fragmentation of mangrove forests was observed, with 1.379 polygons mapped in an area of 2.396 ha, receiving 500 mm of annual rainfall and having 8 dry months. In the Timonha (CE) and Itarema (CE) estuaries, with 1000 mm of rain and 6 dry months, 131 and 118 polygons were identified, respectively, on 2482 and 1093 ha. In contrast, the Mamanguape River estuary (PB), with the highest annual rainfall of 1750 mm and only 3 dry months, has a distribution of 4256 ha in 24 polygons. As for the salt flats, the Piranhas-Açu (RN) has a fragmentation of 674 polygons in an area of 4858 ha and the Mamanguape River estuary (PB) has six polygons mapped in an area of 2.4 ha. The research highlights the impacts of climatic variables on the distribution of mangroves, anthropogenic interference in land use due to economic activities and the negative effects of fragmentation, underscoring the importance of conservation and proper management of these vital ecosystems.
Keywords: Mangrove forests, Salt flats, Brazilian semi-arid coast Mangrove biogeography.
Artigos
A influência de variáveis ambientais na distribuição de manguezais no Nordeste do Brasil
The Influence of Environmental Variables on the Distribution of Mangroves in Northeast Brazil
Ana Caroline Damasceno Souza de Sá carolsouza.geo@gmail.com
Lidriana Pinheiro lidriana@ufc.br
Received: 13 March 2024
Accepted: 25 September 2024
Published: 23 January 2025
Os manguezais são ecossistemas localizados na interface continental e marinha, com dinâmica diária da ação das marés, condicionando características inerentes às espécies que se desenvolvem neste ambiente salobro e com altas concentrações de matéria orgânica (Tomlinson, 1986).
Segundo Schaeffer-Novelli, Vale e Cintrón (2015), os manguezais detém feições caracterizadas como continuum: lavado, mangue (bosque ou floresta de mangue) e apicum. A primeira são os bancos de sedimentos lamosos ou areno-lamosos, sem vegetação, inundados constantemente pelos regimes de marés. O bosque de mangue apresenta vegetação lenhosa típica arbórea que coloniza os extensos substratos halomórficos. O apicum, também denominado de planície hipersalina ou salgados, são zonas de supramaré, onde a água do mar ou do estuário atinge nas preamares de sizígia, podendo limitar-se na transição entre áreas de manguezal e terra firme.
Estes ambientes através de suas funções ecológicas geram diversos serviços ecossistêmicos à sociedade (Costanza et al., 1997; Barbier et al., 2011; Vo et al., 2012; Meireles, 2012; Lee et al., 2014; Duke, 2014; Mukherjee et al., 2014; Costa et al., 2022), em detrimento das pressões humanas que acarretam a degradação ambiental (Lacerda et al., 2021).
As modificações ocorridas no litoral semiárido podem ser elencadas destacando a redução das taxas de precipitação e consequente hipersalinidade em estuários; diminuição dos fluxos terra-oceano; erosão costeira; elevação do nível do mar; aquecimento global e eventos climáticos extremos; eutrofização e perda de biodiversidade devido às atividades humanas (Soares et al., 2021).
Essas alterações têm consequências significativas para a prestação de serviços ecossistêmicos, como a segurança alimentar, demandando estudos para analisar o potencial impacto das mudanças na dinâmica costeira sobre a estrutura e funcionamento dos ecossistemas costeiros e marinhos (Soares et al., 2021). É de importante relevância a compreensão da estrutura e funcionamento dos manguezais para possibilitar conhecimento e a geração de ações estratégicas de planejamento e conservação (ICMBio, 2018; Pinheiro et al., 2023).
Considerando os manguezais em diferentes contextos climáticos, isto é, em áreas com distintos padrões de precipitação e períodos de seca dentro de uma mesma região brasileira, a pesquisa tem como principal questão norteadora: existe diferença na distribuição de feições de bosque de mangue e apicum localizados em ambientes com diferenças climáticas na região Nordeste do Brasil? Portanto, o objetivo desta pesquisa consiste em espacializar os manguezais e correlacioná-los com fatores ambientais, a fim de verificar se existe variação na distribuição da arquitetura dos bosques de mangue e apicum em diferentes condições climáticas.
A área de estudo compreende quatro áreas, os manguezais do rio Timonha (CE) - Área 1, Itarema (CE) - Área 2, Piranhas-Açu (RN) - Área 3 e Mamanguape (PB) - Área 4 (Figura 1).
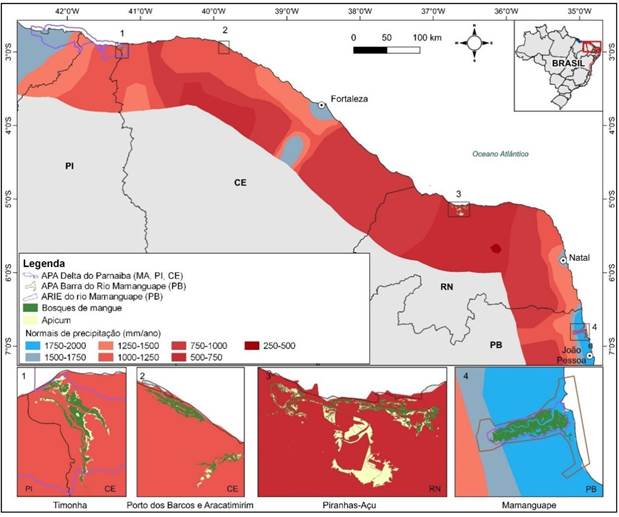
A área de estudos no Litoral do Nordeste é caracterizado por dois tipos climáticos distintos: Clima Tropical de Zona Equatorial (Costa Semiárida Brasileira) e o tropical úmido (Costa dos Recifes) (Diniz et al., 2016). O estuário do rio Timonha (área 1), situa-se nos municípios semiáridos de Chaval e Barroquinha, Oeste do estado do Ceará, na Área de Proteção Ambiental (APA) Delta do Parnaíba (Brasil, 1996).
Ainda na Costa Semiárida, foram estudados os sistemas estuarinos-lagunares de Porto dos Barcos e do rio Aracatimirim, no município de Itarema/Ceará (área 2). No estado do Rio Grande do Norte, localiza-se o estuário do rio Piranhas-Açu entre os municípios de Porto do Mangue e Macau (área 3), região popularmente conhecida como Pólo Costa Branca, devido a elevada produção de sal marinho (Diniz et al., 2020).
O estuário do rio Mamanguape está localizado no estado da Paraíba, nos municípios de Marcação e Rio Tinto (área 4), na Área de Relevante Interesse Ecológico (ARIE) Manguezais da Foz do Rio Mamanguape (Brasil, 1985), que tem seus limites sobrepostos pela Área de Proteção Ambiental (APA) da Barra do Rio Mamanguape, e incluindo parte dos municípios de Lucena e Baía da Traição (Brasil, 1993).
A identificação da feição do manguezal “bosque de mangue” foi realizada com base na estrutura caracterizada por Schaeffer-Novelli, Vale e Cintrón (2015). Para a espacialização foi aplicada a metodologia de Interpretação Visual de Imagens (Jensen, 2000; Florenzano, 2011), elaborando-se uma chave de interpretação para análise visual e vetorização dos polígonos (Figura 2), na escala de 1:10.000, com auxílio do software QGIS v. 3.10.14 (Essen/GNU - General PublicLicense©) (QGIS TEAM, 2021).
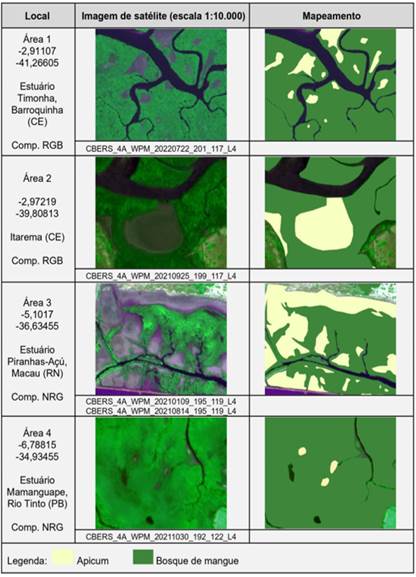
Foram considerados os critérios utilizados por Souza (2022) e Sá (2024), em que no mapeamento da feição “Bosque de Mangue” consideraram-se a tonalidade e coloração verde-escura; com textura rugosa e forma irregular. Posteriormente os dados mapeados foram validados em campo com auxílio de imagens aéreas obtidas por drone, modelo DJI Mavic Air 2.
Em específico para a espacialização dos bosques de mangue do rio Timonha (CE), que está localizado muito próximo do rio Ubatuba, devido à mistura de águas dos rios, não há como delimitar precisamente até onde chega a influência de ambos, foi definido como critério um buffer na margem esquerda a partir da legislação vigente, lei nº 12.651/2012 (Novo Código Florestal) que delimita as Áreas de Preservação Permanente (APP) de acordo com a faixa do curso do rio (Brasil, 2012).
Foram consideradas três faixas ao longo do estuário, sendo a foz o local de maior área de faixa (buffer), delimitando 500m, pois a largura do rio é superior a 600m, em seguida, onde a largura do estuário encontra-se entre 600 e 200m, a margem esquerda foi de 200m e por fim, onde a largura encontrava-se entre 200 e 50m, a margem foi delimitada em 100m.
Para a análise de variáveis ambientais (Quadro 1), foi realizada a Correlação de Pearson, conforme Silva et al. (2022), buscando analisar se existe uma possível relação entre elas:

Para a análise dos dados, foi utilizado o software R (R CORE TEAM, 2022), RStudio (R STUDIO TEAM, 2022), com o Rtools 4.3.3 (5863-5818).
No Semiárido do Nordeste, localizado no clima tropical de Zona Equatorial (Diniz et al., 2016), o estuário com os mais baixos índices é o complexo Piranhas-Açu (RN) (área 3), com normais de precipitação médias anuais em torno de 500 mm/ano e oito meses secos (INMET, 2021). As áreas do rio Timonha e Itarema (CE) possuem precipitações de 1.000 mm/ano e seis meses secos (FUNCEME, 2023a, 2023b). No Clima Tropical do Nordeste Oriental, de subdomínio úmido moderado e três meses secos, a área do rio Mamanguape possui Precipitação Média Anual de 1.750 mm (Diniz et al., 2016) (Tabela 1).
Foram registradas quatro espécies de mangue verdadeiro nas áreas 1, 2 e 4 (Figura 3): I) Rhizophora mangle Linnaeus (1753); II. Laguncularia racemosa C. F. Gaert (1807); III. Avicennia germinans Linnaeus (1764); IV. Avicennia schaueriana Stapf & Leechm (1939). Na área 3 foram identificadas três espécies de mangue verdadeiro R. mangle, L. racemosa e A. schaueriana. Nas quatro áreas em estudo, foram identificadas uma espécie associada ao mangue, a Conocarpus erectus Linnaeus (1753), sendo contabilizada em cinco e quatro espécies, conforme a tabela 1.
A área 4 do Mamanguape apresenta a maior extensão de bosques de mangue, totalizando 4.256 hectares distribuídos em 24 polígonos (Figura 4). O manguezal do rio Timonha (área 1) abrange uma área de 2.482 hectares, fragmentado em 131 polígonos. Por sua vez, o bosque de mangue do Piranhas-Açu (área 3) exibe a maior fragmentação, com 1.379 polígonos e uma área de 2.396 hectares. A menor extensão é encontrada em Itarema (área 2), com 1.093 hectares distribuídos em 118 fragmentos.


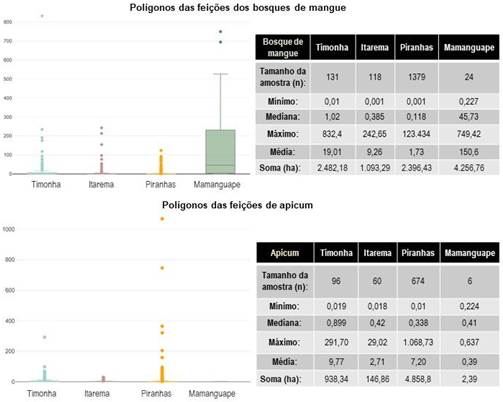
A feição de apicum foi espacializada de forma bastante significativa no estuário do rio Piranhas-Açu (RN) em detrimento dos demais, com uma área de 4.858 ha com 674 polígonos, enquanto a menor extensão foi identificada no estuário do rio Mamanguape (PB) em uma área de aproximadamente 2,4 ha distribuídos em 6 polígonos.
Os pares de variáveis apresentaram configurações especiais, indicando algum tipo de relação entre elas, sendo possível perceber a relação linear entre alguns pares (Tabela 2).
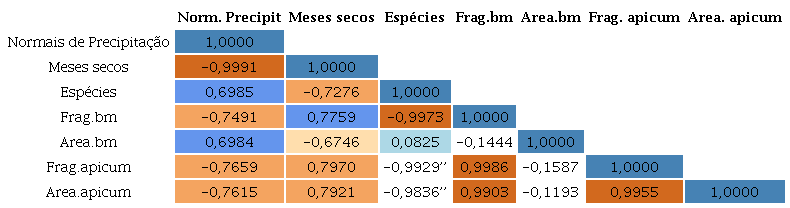
De acordo com a análise dos dados, existe uma correlação negativa muito forte (-0,99) entre os meses secos e as normais de precipitações médias anuais, sugerindo que uma menor precipitação está relacionada a um aumento no número de meses secos ao longo do ano, uma característica das precipitações no sertão nordestino, no qual as precipitações são baixas e concentradas em poucos meses durante o ano.
Nesta perspectiva, a distribuição das espécies de mangue e a fragmentação do manguezal também apresentam uma correlação negativa muito forte (-0,99), onde uma maior fragmentação está associada a uma menor distribuição das espécies.
A correlação entre a fragmentação dos bosques de mangue e apicum se mostrou muito forte (0,99), quanto maior a área de fragmentação da feição de bosque de mangue, maior a fragmentação e área do apicum.
A fragmentação dos manguezais associadas com as normais climatológicas de precipitações médias anuais, resultou em uma correlação negativa (-0,74). O que pode indicar que uma menor quantidade de precipitações contribui para uma maior fragmentação dos manguezais e área do apicum.
Os dados das correlações apresentam uma relação linear entre a fragmentação dos bosques de mangue e as normais de precipitação, as quais são inversamente proporcionais aos meses secos durante o ano e à distribuição de espécies. Pode-se observar no estuário hipersalino Piranhas-Açu (RN) a fragmentação dos bosques de mangue em 1.379 polígonos e a normal climatológica com precipitações de aproximadamente 500 mm/ano contendo 8 meses secos, já nos estuários das áreas 1 (Timonha) e 2 (Itarema), onde a precipitação anual é de 1.000mm com 6 meses secos, foram mapeados 131 e 118 polígonos, respectivamente, enquanto o estuário do rio Mamanguape (PB) tem uma distribuição dos bosques de mangue em 24 polígonos e precipitações médias anuais de 1.750 mm/ano e 3 meses secos.
Durante os trabalhos de campo foram identificadas quatro espécies de mangue verdadeiro (R. mangle, L. racemosa, A. Schaueriana, A. germinans) e uma espécie associada (C. erectus) nas áreas 1, 2 e 4, exceto no estuário Piranhas-Açu (RN) - Área 3 a qual não foi identificada a A. germinans, dado reforçado por Souza, Silva e Costa (2023) que identificaram no referido estuário as quatro espécies citadas (R. mangle, L. racemosa, A. Schaueriana e C. erectus).
Observa-se em estuários do Nordeste brasileiro a ocorrência de A. germinans (Maia; Coutinho, 2012; Maia, 2016; Ximenes et al., 2016) pela sua capacidade de sobrevivência em alta salinidade. Esta resposta foi encontrada por Costa, Rocha e Cestaro (2014), onde a espécie se distribui desde a foz até a montante no estuário do rio Apodi-Mossoró, aproximadamente 40km a oeste do Piranhas-Açu, mas não foi encontrada no referido estuário durante os trabalhos de campo, necessitando de estudos mais específicos para compreender os elementos que levaram a ausência da sua distribuição na área 3, no Piranhas-Açu (RN).
A fragmentação do manguezal apresentou uma relação negativa com a distribuição das espécies (-0,99), mas isto não indica que uma variável influencia a outra ou que existe uma relação de causa e efeito entre elas como Zar (2009) e Silva et al. (2022) destacam na aplicação de métodos estatísticos como a Correlação de Pearson.
A distribuição de espécies de mangue pode ocorrer por diversos motivos, como resposta aos próprios limites fisiológicos das espécies, com variáveis relacionados à salinidade, temperatura do ar, temperatura da superfície do mar, precipitação, aridez, amplitude das marés, correntes oceânicas, a própria dispersão dos propágulos (sementes) através da hidrocoria e viviparidade, além das ações antrópicas (Tomlinson, 1986; Schaeffer-Novelli et al., 1990; Osland et al, 2007; Lima; Galvani, 2010; Schaeffer-Novelli et al., 2015; Ximenes et al., 2016; Lacerda et al., 2021; Soares et al., 2021; Adame et al., 2021; Madeira et al., 2023).
Esse conjunto de condicionantes ambientais resulta na formação dos manguezais. As características de topografia aplainada e clima semiárido das planícies costeiras do litoral setentrional do Rio Grande do Norte fazem com que durante as dinâmicas das marés, com amplitude variando de 0 a aproximadamente 2.8 m nas marés de sizígia, alcancem vastas áreas. Onde há o contato diário com as marés, ocorre a colonização da planície por mangue, pois nos locais mais elevados onde apenas as águas das marés de sizígia alcançam, a salinidade se torna um fator limitante para a presença da flora.
Mesmo que as espécies desenvolvam mecanismos de eliminação de sais, há um limite para sua colonização (Tomlinson, 1986), resultando em extensas áreas de apicum, caracterizadas por feições de solo exposto ou com vegetações herbáceas halófitas (Figura 4). Ademais, a precipitação média anual é de aproximadamente 500 mm, com chuvas concentradas em quatro meses, e os rios da bacia hidrográfica do rio Piranhas-Açu apresentam um regime intermitente ou temporário, entre outros elementos que contribuem para a formação dessas paisagens (Diniz, 2013; Medeiros; Pinheiro, 2018; Medeiros, 2020).
Medeiros e Pinheiro (2018) investigaram efeitos da insuficiência pluvial no estuário do Piranhas-Açu, no qual o clima semiárido da região é caracterizado por baixos índices pluviométricos e irregularidade de chuvas, com elevadas taxas de evapotranspiração potencial, resultando em um déficit anual de 1.829,8 mm. Combinado a outros fatores como a quantidade de barramentos na bacia hidrográfica (1.536 barramentos) e retirada da água para atividades econômicas, diminui consideravelmente a vazão dos rios para a zona estuarina, a fonte de água doce se torna quase “nula” (Godoy; Lacerda, 2015; Diniz; Vasconcelos, 2016; Medeiros, 2020). Nos estuários do semiárido brasileiro percebe-se tendência a hipersalinidade sazonal (Morais; Pinheiro, 2011; Soares et al., 2021). Essas condições afetam a vegetação local das florestas de mangue e Caatinga no entorno do Piranhas-Açu, diminuindo 41,8% entre os anos de 2013 e 2015 (Medeiros; Pinheiro, 2018).

Nos estuários de baixa energia, o acúmulo de sedimentos propiciam a formação de ilhas e bancos de areia (Morais; Pinheiro, 2011; Godoy; Lacerda, 2014; 2015). Esse processo estabelece condições favoráveis para a colonização por manguezais, como observado nas ilhas presentes nos estuários dos rios Timonha, Aracatimirim e Piranhas-Açu (Morais; Pinheiro, 2011; Godoy; Lacerda, 2014; 2015; Godoy, 2015).
Na área 1 entre os estuários do rio Timonha e Ubatuba, a Ilha Grande apresenta quase 2.000 ha, sendo uma das maiores ilhas estuarinas do Nordeste (Choi-Lima, 2017), contendo outras duas, a ilha do Coronel e ilha dos Preás, com aproximadamente 700ha e 855ha, respectivamente. Outros 11 polígonos menores de bosques de mangue foram mapeados em ilhas no curso do rio Timonha com áreas entre 54 e 0,5 ha.
No Aracatimirim, o tamanho dos polígonos de bosque de mangue mapeados em ilhas é menor, variando entre 9 e 0,03 ha dentre 14 polígonos. No Piranhas-Açu a maior ilha tem tamanho de 221ha (a oeste de Macau, se assemelha a um coração invertido, conhecida por Ilha Maracanã), os polígonos do bosque de mangue em ilhas somam-se em 145 variando entre 78 e 0,001ha. E, no Mamanguape, foram mapeadas oito ilhas contendo oito polígonos de bosques de mangue, variando entre 78 e 0,32 ha.
Como exemplo de expansão dos manguezais, ao comparar os dados fornecidos pelo MapBiomas (2022) para o ano de 1985 (Souza Júnior et al., 2020), em que os bosques de mangue no estuário do rio Aracatimirim compreendiam uma área de 24,57 ha e de acordo com os dados desta pesquisa, a área mapeada em 2021 foi de 324 ha, resultando no aumento de 1,218% das áreas de manguezal. Além da intrusão salina e baixa vazão fluvial, outros fatores de condições globais como o aumento do nível do mar e mudanças climáticas propiciam sua expansão (Godoy; Lacerda, 2015; Soares et al., 2021).
Estudos de Albuquerque et al. (2014a), Albuquerque (2015) e Medeiros (2020) apontam a correlação do desenvolvimento e extensão dos apicuns com variáveis climáticas, como o déficit hídrico, a topografia aplainada, a hidrografia de rios intermitentes, a dinâmica costeira e a oscilação das marés. Esses ambientes extremos e hipersalinos permitem o desenvolvimento de apenas algumas espécies herbáceas, que são adaptadas a condições de alta salinidade, pelos fatores citados anteriormente e pela sua topografia ser um pouco mais elevada, a qual apenas as marés mais altas, preamares de sizígia chegam nesses locais, obtendo a evaporação e consequentemente um acúmulo de sais (Schaeffer-Novelli et al., 2015; ICMBio, 2018; Medeiros, 2020).
O apicum é de grande importância, pois, com a tendência de elevação do nível do mar, os bosques de mangue migrarão para as áreas que hoje são apicuns. Pesquisas mostram que áreas atualmente ocupadas por apicuns, foram, no passado, ocupadas por bosques de mangue, indicando que ao longo do tempo geológico, essa dinâmica ocorre periodicamente (Albuquerque et al., 2014b; Schaeffer-Novelli et al., 2016; Barbosa et al., 2018; Soares et al., 2021).
Em ordem decrescente, o estuário do Piranhas-Açu apresenta uma fragmentação de 674 polígonos em uma área de 4.858 ha com normais de precipitação de 500mm/ano e 8 meses secos, o Timonha 96 polígonos com área 938ha com índices pluviométricos anuais de 1008 e 6 meses secos, Itarema (CE) com 60 polígonos em uma área de 146 ha que tem precipitações de 1009 mm/ano e 6 meses secos e o estuário do rio Mamanguape (PB) seis polígonos mapeados em uma área de 2,4 ha com precipitações anuais de 1.875mm e 3 meses secos.
A fragmentação nos bosques de mangue no semiárido é bastante elevada, 49% dos fragmentos são inferiores a 1ha no Timonha, 71% na área 2, e 82% da fragmentação menor que 1ha no Piranhas-Açu. Enquanto na área 4 apenas 2% dos fragmentos tem área inferior a 1ha. Dos fragmentos dos apicuns, no Timonha 51%, Itarema 63%, Piranhas-Açu 69%, Mamanguape 100% são polígonos de apicum inferiores a 1ha.
Um dos impactos negativos causados decorrentes da fragmentação consiste na perda da conectividade da paisagem, alterando a capacidade das espécies em habitar o ecossistema (Jaramillo et al., 2023). As modificações nos fragmentos concentram-se nas bordas, chamado de efeito de borda (Souza, et al., 2023), em áreas mais continentais, no ecótono do apicum para o continente, e podem ocorrer mais veemente em decorrência das atividades antrópicas como o desmatamento e mudanças no uso do solo (Bryan-Brown et al., 2020).
A ocupação do manguezal nas áreas em estudo por tanques de aquicultura/carcinicultura e salinas solares é evidente. Costa et al (2022) estudaram serviços ecossistêmicos entre as diferentes unidades de paisagem no sistema estuarino Galinhos-Guamaré no litoral semiárido do Rio Grande do Norte, sendo possível comparar a prestação de serviços em estuário/canais de maré, bosque de mangue, apicum, planícies de maré, salinas solares e carcinicultura. Como resultado, foi identificado que as áreas de bosque de mangue e apicum ofertam respectivamente 3,7 e 2,7 vezes mais serviços ecossistêmicos do que tanques de carcinicultura.
Nas áreas 1 e 3 há ocupação das planícies por salinas solares, e em todas as áreas há fazendas de camarão, a carcinicultura. Na área 1 devido ao clima, as salinas funcionam apenas durante o período dos meses secos e é pouco rentável (Araújo, 2013). Já no estado do Rio Grande do Norte, a presença de salinas solares é evidente nos estuários do litoral setentrional majoritariamente nas áreas que antes eram manguezais (apicum e bosques de mangue), chegando a 95% da produção nacional de sal marinho, devido as condicionantes ambientais locais. No município de Macau, a Salinor tem uma área de 4.540,53 ha de tanques evaporadores e cristalizadores produz aproximadamente 45% de todo sal marinho brasileiro (Costa et al., 2013; Diniz, 2013; Soares et al., 2018). Já a produção de camarão nos estados do Ceará e Rio Grande do Norte obteve em 2021, segundo a Associação Brasileira de Criadores de Camarão (ABCC, 2022) 55,6 e 26 t, respectivamente, e ocupam o 1º e o 2º lugar no ranking nacional, onde o Ceará produziu 47,1% de todo camarão nacional e o Rio Grande do Norte 21,6%.
A aquicultura e as salinas solares no município de Chaval-CE, na área 1, compreendem 323 ha, de acordo com a Secretaria Estadual de Meio Ambiente Do Ceará (Sema, 2022). Já no estuário do rio Piranhas-Açu a área ocupada é bastante expressiva, com 12.000 ha de tanques de salinas solares e 2.100 ha de tanques de carcinicultura, somando, tem-se um total de 14.100 ha (Saldanha, 2020). A área ocupada por estes dois tipos de empreendimentos é 43 vezes maior no Piranhas-Açu (área 3) do que no Timonha (área 1). Em Itarema, área 2, a aquicultura/carcinicultura ocupa uma área de 555 ha (Sema, 2022), enquanto a ocupação no Mamanguape (área 4) pela carcinicultura é de 82 ha (ICMBio, 2014).
Bryan-Brown et al (2020) apontam tendências globais de fragmentação dos manguezais como a conversão para aquicultura, agricultura e expansão urbana, que estão associadas com o desmatamento. Gilani et al (2021) e Grantham et al (2020) destacam também outros fatores como a construção de estradas, visualizada nas áreas 2, 3 e 4, respectivamente, em Porto dos Barcos (Itarema/CE), Piranhas-Açu (RN) e Mamanguape (PB). Consequentemente essas modificações causam desequilíbrios ambientais e impactos ecológicos que afetam as funções ecossistêmicas (Polidoro et al., 2010; ICMBio, 2018), como a diminuição de espécies de peixes estuarinos (Tran; Fischer, 2017), liberação de emissões de carbono, perda da biodiversidade e qualidade da água (Atwood et al., 2017; Hagger; et al., 2022).
Corte et al. (2021) fornecem evidências de que mesmo pequenos os fragmentos dos manguezais melhoram a biodiversidade e as funções ecossistêmicas, reconhecendo sua importância como integração do ambiente. Li et al. (2013), Estoque et al (2018) e Gilani et al (2021) destacam o estabelecimento de reservas naturais de manguezais como a forma mais eficaz de proteger e expandir os manguezais; a implementação de monitoramento eficaz; a instauração de programas e projetos educacionais; para que sejam perpetuados a estrutura e funcionamento dos ecossistemas para propiciar a oferta de serviços ecossistêmicos por estes ambientes. No Brasil, mesmo com um avançado arcabouço legislativo, a falta de fiscalização e monitoramento nos manguezais, sobretudo os que não estão localizados em unidades de conservação, se tornam um fator limitante à conservação deste ecossistema (Ferreira; Lacerda et al., 2016).
Segundo o Instituto Chico Mendes de Conservação da Biodiversidade (ICMBIO, 2018, p. 59) a presença de Unidades de Conservação (UCs) no Brasil circunscritas no ecossistema manguezal é de 120 unidades dentre as 2.071 UC existentes, correspondendo a uma área de 12.114 km², que representa 87% do ecossistema em todo o Brasil. Ressalta-se que nacionalmente os manguezais são resguardados pela lei nº 12.651/2012, que os delimitam em Área de Preservação Permanente (APP) (Brasil, 2012).
O estabelecimento de unidades de conservação exige uma série de medidas de planejamento e monitoramento para analisar e regulamentar a utilização dos recursos das áreas protegidas, visando o desenvolvimento sustentável nos ecossistemas englobados nas UCs de Uso Sustentável, e de preservação dos ambientes nas UCs de Proteção Integral (Brasil, 2000). Neste ponto, as três UCs envolvidas neste estudo fazem parte da categoria de Uso Sustentável. A área 1 (estuário do rio Timonha, no semiárido brando) está localizada na APA Delta do Parnaíba e a área 4 apresenta duas UCs, a ARIE Manguezais da Foz do Rio Mamanguape e a APA da Barra do Rio Mamanguape. O que contribui para um planejamento e controle dos usos dos recursos naturais das áreas (Paludo; Klonowsky, 1999; ICMBio, 2014).
Os dados apresentam uma complexa interação entre a distribuição dos manguezais e variáveis ambientais, como os padrões climáticos regionais. As análises indicam que a fragmentação dos manguezais é inversamente proporcional à precipitação média anual, sugerindo que o padrão climático com baixas precipitações e maior número de meses secos podem levar a uma maior divisão dos manguezais, além das extensas áreas de apicum, como observado no estuário do rio Piranhas-Açu (RN), onde a precipitação média anual é de aproximadamente 500 mm e esta área tem um período de 8 meses secos.
Além disso, a correlação estatística entre a fragmentação e a distribuição das espécies, embora significativa, não implica causalidade, o que ressalta a necessidade de uma compreensão mais profunda dos mecanismos subjacentes que regem essas relações ecológicas, pois a presença e distribuição das espécies de mangue são influenciadas por uma série de fatores, que vão dos limites fisiológicos das espécies a elementos antrópicos, como mudanças no uso do solo.
A expansão dos manguezais em áreas anteriormente não ocupadas e os impactos negativos causados pela fragmentação, como a perda de conectividade e os efeitos de borda, destacam a importância de medidas de conservação e manejo adequado desses ecossistemas. O estabelecimento, monitoramento e a manutenção de unidades de conservação contribuem diretamente para a gestão sustentável dos manguezais. Assim, é essencial um esforço contínuo para promover a conservação efetiva desses importantes ecossistemas costeiros.
