Calliphora vicina (Diptera: Calliphoridae): Growth rates, body length differences, and implications for the minimum post-mortem interval estimation
Calliphora vicina (Diptera: Calliphoridae): Tasas de crecimiento, diferencias de longitud corporal e implicaciones para la estimación del intervalo post-mortem mínimo
Calliphora vicina (Diptera: Calliphoridae): Growth rates, body length differences, and implications for the minimum post-mortem interval estimation
Revista de la Sociedad Entomológica Argentina, vol. 81, núm. 2, 2022
Sociedad Entomológica Argentina
Recepción: 27 Enero 2022
Aprobación: 26 Junio 2022
Abstract: Calliphora vicina Robineau-Desvoidy is a primary colonizing species of decomposing bodies, being predominant in the cold seasons in Buenos Aires province, Argentina. The minimum post-mortem interval (PMImin) was estimated using growth and larval length parameters as the accurate entomological evidence of the elapsed time since death. Specimens were reared at 12, 14, 16, and 18 °C, and the larval and pupal growth rates were determined. Differences in the developmental times were registered (43.2 days at 12 °C, 34.3 days at 14 °C, 29.1 days at 16 °C, and 24.3 days at 18 °C), with the lifespan shortened at higher temperatures, without changing the larval length range (~1.5–18.0 mm). Temperature also influenced the survival and size of emerged adults; the highest percentage of survival registered at 14 (29.73%) and 16 °C (38.65%), and more mortality in extreme temperatures (19.00% at 12 °C and 12.61% at 18 °C). Adults reared at 12 °C were the largest of all, females were longer than males, and body size showed an inverse relationship with survival. These results allowed the PMI estimation based on data from local populations instead of consulting papers based on other regions.
Keywords: Age estimation, Differential survival, Forensic entomology, Sexual size dimorphism.
Resumen: Calliphora vicina Robineau-Desvoidy es una especie colonizadora primaria en cuerpos en descomposición siendo predominante en las estaciones frías en Provincia de Buenos Aires, Argentina. Se estimó el intervalo post-mortem mínimo (PMImin) usando el crecimiento y longitud larvales como evidencia entomológica precisa del tiempo de muerte transcurrido. Se criaron especímenes a 12, 14, 16 y 18 °C para determinar las tasas de crecimiento larvales y pupales. Se registraron diferencias en los tiempos de desarrollo (43,2 días a 12 °C; 34,3 días a 14 °C; 29,1 días a 16 °C y 24,3 días a 18 °C), observándose tiempos de vida más cortos a mayor temperatura, sin alterarse los rangos de longitud larval (~1,5–18,0 mm). La temperatura también afectó la supervivencia y tamaño de los adultos emergidos; registrándose mayor porcentaje de supervivencia a 14 (29,73%) y 16 °C (38,65%) y mayor mortalidad a temperaturas extremas (19,00% a 12 °C y 12,61% a 18 °C). Los adultos criados a 12 °C fueron los más grandes, las hembras mayores que los machos con una relación inversa entre tamaño y supervivencia. Estos resultados permiten estimar el PMI basado en datos de poblaciones locales en lugar de consultar registros obtenidos en otras regiones.
Palabras clave: Dimorfismo sexual en el tamaño, Entomología forense, Estimación etaria, Supervivencia diferencial.
INTRODUCTION
Forensic entomology is based on the study of the insects and other arthropods, which sequentially colonize a decomposing corpse to feed on it, to prey on other organisms that develop inside this substrate or to use it as a refuge (Greenberg, 1991). Studies about succession have shown that the most common insects, associated with dead animals and humans, are the blowflies (Calliphoridae). Their developmental rates are directly related to environmental data (mainly temperature and humidity) (Greenberg, 1991; Centeno et al., 2002). As well, the species of flies associated with a body varies according to its decay process, body mass, kind of substrate, and cover in which they are found in each microhabitat, geographic region, and different seasons of the year (Greenberg, 1991; Marchenko, 2001; Oliva, 2001; Centeno et al., 2002; Matuszewski et al., 2014; Hadjer-Kounouz & Kamel, 2017). Therefore, forensic entomology research has to establish the date of death or minimum post-mortem interval (PMImin) in different environments and conditions that can modify and modulate the progress of this process. However, based on environmental conditions and the climate of a region, one species may be more abundant than the other ones (Centeno et al., 2002). There are several methods to establish the insect’s age by comparing the larval size (length or weight) with the development’s temperature and time, such as isomorphen/isomegalen diagrams and the thermal summation model (Grassberger & Reiter, 2001; Amendt et al., 2011). Furthermore, the differences in experimental methods or feeding substrate may also lead to discrepancies in the growth data.
At the species level, the individual size distribution, discriminated by sex or not, and growth rate of individuals depend on genetic and morphological differences, and they are also modeled by physiological factors in development, fertility, and the environment in which they develop (Parker & Johnston, 2006; Tarone et al., 2011). In addition, the breeding temperature (Woods et al., 2003; Stillwell & Fox, 2007; Tarone et al., 2011,Kotzé et al., 2015), quality or quantity of available food (Clark et al., 2006; Ireland & Turner, 2006; Rabêlo et al., 2011; Richards et al., 2013), and the number and sex of developing individuals (Johnson & Wallman, 2014; Macedo et al., 2018) can affect the range of stage size distributions.
The main life traits for the laboratory-reared flies are related to size at each immature stage in the life cycle, developmental rates and survival, and number, size and sex of emerged adults. In particular, the immature stages and/or emerged adults of Calliphora vicina Robineau-Desvoidy, can show various size ranges, without the larval mass significantly affecting the growth rate (length and weight) (Pellitero & Bordas, 2007; Richards et al., 2013; Johnson & Wallman, 2014). However, starvation conditions and type of diet produce reduced sizes and an increase in development times (Pellitero & Bordas, 2007; Richards et al., 2013). Calliphora vicina is widely distributed throughout the world, commonly identified in forensic samples, being found in temperate and subtropical regions mainly during the cold months of the year or for long periods at temperatures up to 30 °C (Marchenko, 2001; Oliva, 2001; Centeno et al., 2002). Moreover, its life cycle has been described under constant (Reiter, 1984; Anderson, 2000; Donovan et al., 2006; Díaz Martin et al., 2014; Salimi et al., 2018) and fluctuating temperatures (Anderson, 2000; Oliva, 2001; Ames & Turner, 2003; Niederegger et al., 2010; Vinogradova & Reznik, 2015) between 4 and 30 °C, since development can be interrupted at higher temperatures. In Buenos Aires (Argentina), C. vicina is active from autumn to the spring, being the most common species in winter with fluctuating temperatures of 5-20 ºC, and ovipositing and developing (Oliva, 2001; Centeno et al., 2002). These studies showed that C. vicina growth was related to rearing temperature and geographical origin because of similar temperature regimes, and developed larvae and pupae from various regions showed different sizes and developmental times (Reiter, 1984; Anderson, 2000; Oliva, 2001; Donovan et al., 2006; Pellitero & Bordas, 2007; Díaz Martin et al., 2014; Hadjer-Kounouz & Kamel, 2017). Despite the studies on the development of this species, information is still lacking for populations from different geographical regions. To study of the fly population and their larval stages at a crime scene can help forensic scientists estimate the time of death, we evaluated the temperature effect on life traits such as size and development time of immature stages, survival and sizes of adults, and the sex ratios of C. vicina to improve the estimation of PMImin in the Argentinean population.
MATERIAL AND METHODS
Sampling and maintenance of flies. A laboratory colony of C. vicina was established by collecting adults in the fall using meat as bait and an entomological net in the gardens of the National University of Quilmes, Buenos Aires Province, Argentina (34°42’24’’S, 58°16’39’’W). In Buenos Aires (The Argentinean Pampas ecoregion), the climate is humid-warm, rains are distributed through the year with annual rainfalls of around 700-1,200 mm from Southwest to Northeast, with an annual mean temperature between 12-15 °C in winter and 20 °C in summer (Morello et al., 2012). Flies were identified using Mariluis & Schnack (2002) and Whitworth (2012) keys and kept in a cage [40 x 30 x 30 cm] at 22 ± 2 °C and 50 ± 5% relative humidity. Adults were fed 50:50 sugar: water solution, pure water, and milk powder ad libitum, including fresh beef to allow them to oviposit. The beef was checked until the flies laid the eggs. About 250 eggs from several flies were randomly collected and transferred to rearing cages (plastic boxes of 13.0 x 9.0 x 4.5 cm) containing 20 g of fresh beef in pine shavings 2 cm deep to homogenize the samples. Fresh meat was added every other day to reduce competition and starvation of larvae and larval size variability. Ten replicates for each constant temperature treatment (12, 14, 16, and 18 ± 1 °C) were performed using incubators (SEMEDIC I-290PF) and photoperiod 23:1 (darkness: light).
Growth under constant temperature regimes. The development period (mean ± standard error, SE) from oviposition until the emergence of adults for each rearing cage at each temperature regimen was determined. The average time (mean ± SE) to reach each developmental stage, the duration of each one (first, second, and third larval instars, pupal period), and the total development time from egg to adult were calculated. After hatching, three larvae were randomly collected per day, killed by immersion in water at 90 °C for 2 min, the total length measured, and the larvae preserved in 70% ethanol. Instars were determined by examining the morphology of the posterior spiracles with a stereomicroscope. All developed pupae length was also measured and observed daily until all adults emerged. Pupae survival was calculated by recording the number of all emerged adults at each temperature treatment. A hundred adult females and males for each range of temperatures were measured considering total body length (BL), head width (AW), and thorax high (TH) to obtain their relative sizes. The fly’s volume is roughly proportionate to the amount of ingested food in the immature stages, and the parallelepipedon structure is proportional to the body shape (Maldonado & Centeno, 2003). The sex ratios (female/male) were also measured. Measurements of the total length of larvae, the total length of pupae, and the size of adults were taken, with a micrometer using a Leica stereomicroscope, model type EZ4 (35×) (Bio-Optic S.R.L., Argentina).
Statistical analyses. The developmental time of immature stages was analyzed at each temperature using the Kruskal-Wallis ANOVAs on ranks for global comparisons (p< 0.05), followed by the Kruskal-Wallis all-pairwise comparison test for contrasts between treatments. To evaluate the relationship between the rearing temperature and sex of the adults, we ran two-factor analyses of covariance (ANCOVA) on thorax high (TH) and head width (AW), with body total length (BL) as the covariate (p< 0.05). Numerical differences are indicated in the text as means with 95% confidence intervals. To evaluate the relationship between body length and development time for each immature stage (4 levels: first, second, and third larval instars, pupal period) and rearing temperature (4 levels: 12, 14, 16, and 18 °C), a generalized linear mixed model (GLMM), fitted with a quasi-Poisson family with the logit link function, followed by a Fisher’s LSD test comparison between treatments was also performed. The model chosen was based on the Akaike information criterion (AIC). Adult sex ratios at each assessed temperature were determined using the normal distribution. The percentages of the emerged adults with the proportion test were analyzed. Data analyses were performed using InfoStat software applying an R interface (Di Rienzo et al., 2017).
RESULTS
The developmental rate of C. vicina was significantly affected by temperature. At higher temperatures, development times of the eggs, first instar, second instar, third instar, and pupae decreased (Table I). It took around 43 days to become an adult at 12 °C; 34 days at 14 °C; 29 days at 16 °C, and 24 days at 18 °C; therefore, the total development time was shortened by 19 days between 12 and 18 °C (Fig. 1). Total development time was reduced to nearly 9 days at 14 °C, 14 days at 16 °C, and to 9 days at 18 °C, compared with the 43 days at 12 °C (Fig. 1). The pupal stage represented 60% of global development, regardless of the rearing temperature (Table I).
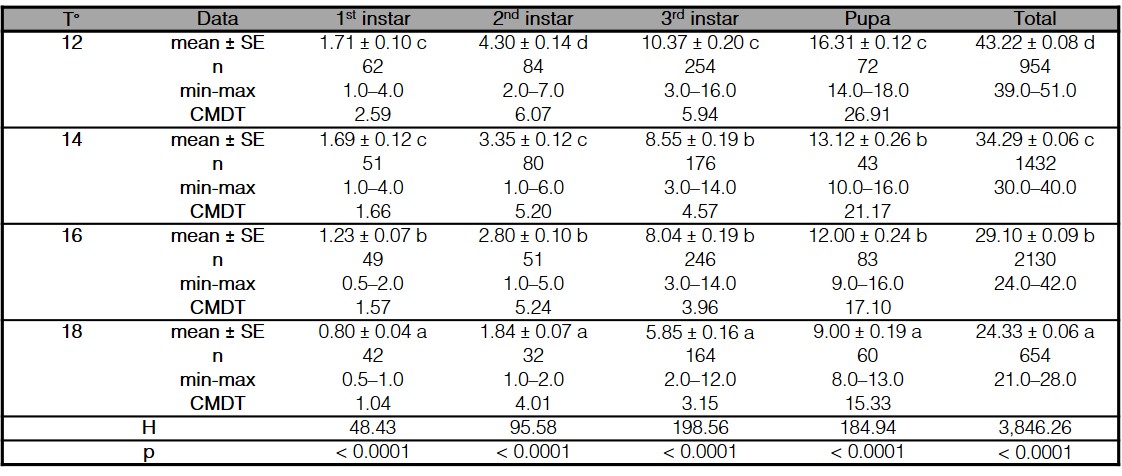
Table I.
Average development time (mean ± SE, days), time range to reach each stage (min-max, days), and cumulative mean development time (CMDT) of each immature stage of Calliphora vicina reared at different temperatures (°C).
n indicates the number of sampled individuals in each stage and treatment. Comparisons for each development stage different temperatures were analyzed with the Kruskall-Wallis ANOVA test on ranks for global comparisons (p< 0.05), followed by the Kruskall-Wallis all-pairwise comparison test for contrasts between treatments. Different letters within the same column indicate significant differences (p< 0.05).
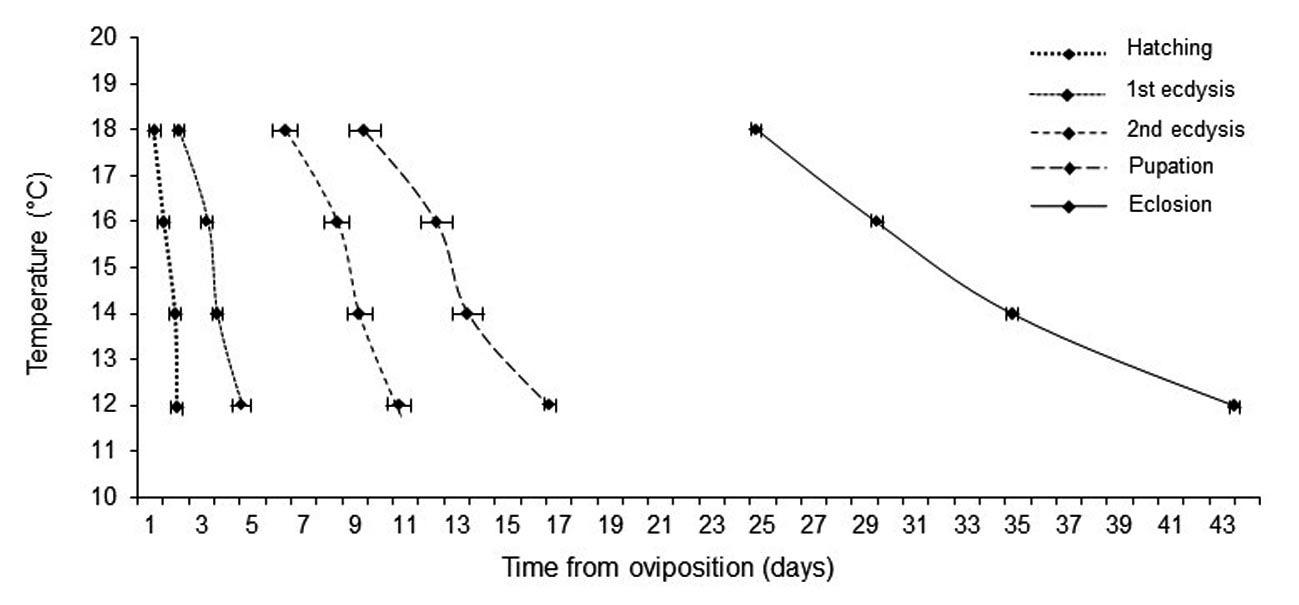
Fig. 1.
Isomorphen diagram of Calliphora vicina. Developmental time of immature stages to reach a particular developmental event (reference on the graph), at a range of constant temperatures (°C).
The size of the larvae of C. vicina was related to its age as a function of time by showing a rapid increase in all feeding phases; the first and second instars’ were similar in sizes independently of the rearing temperature (Table II). Larval length markedly increased from second to the third instar, the growth curve presented a change in the curvature, corresponding to the maximum value of the growth of this larval stage. A significant decrease in the post-feeding phase and pupal size was observed in all treatments, as the length gradually decreased until reaching a horizontal asymptote corresponding to the pupal stage (Fig. 2).
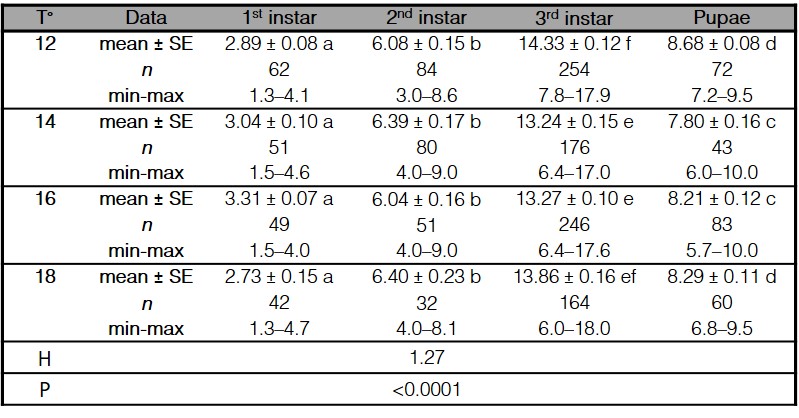
Table II.
Average length of immature stages (mean ± SE, mm) and total length range (min-max, mm) of Calliphora vicina reared at different temperatures (°C).
n indicates the number of sampled individuals in each stage and treatment. Comparisons for all development stages different temperatures were analyzed with the Kruskall-Wallis ANOVA test on ranks for global comparisons (p< 0.05), followed by the Kruskall-Wallis all-pairwise comparison test for contrasts between treatments. Different letters indicate significant differences (p< 0.05).
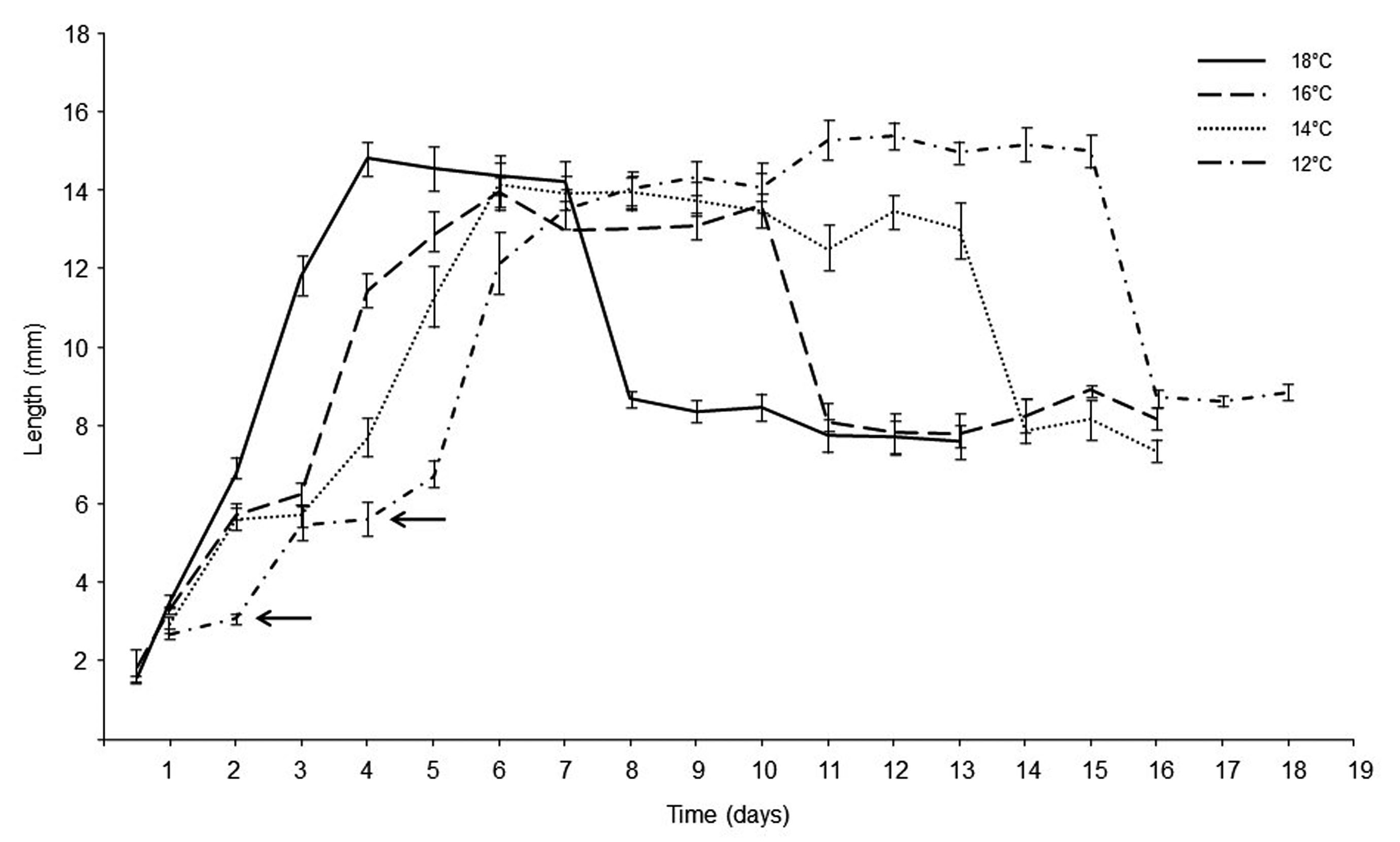
Fig. 2.
Growth parameters of Calliphora vicina. Growth curves in the length of C. vicina over time from hatching to pupation at different temperature (°C) regimes. Arrows indicate 1st and 2nd moulting.
The third instars and pupae developed at 12 and 18 °C were longer than the individuals reared at 14 and 16 °C (Fig. 2, Table II). Body length was significantly affected by the immature stages (GLMM: df = 3; p< 0.0001), but not by the rearing temperature (df = 3; p= 0.1794), being the interaction between both variables also significant (df = 9; p< 0.0001). Then, each immature stage could be estimated based on its body length from the first instar until the peak of the third feeding stage, independently of the rearing temperature (Table III).
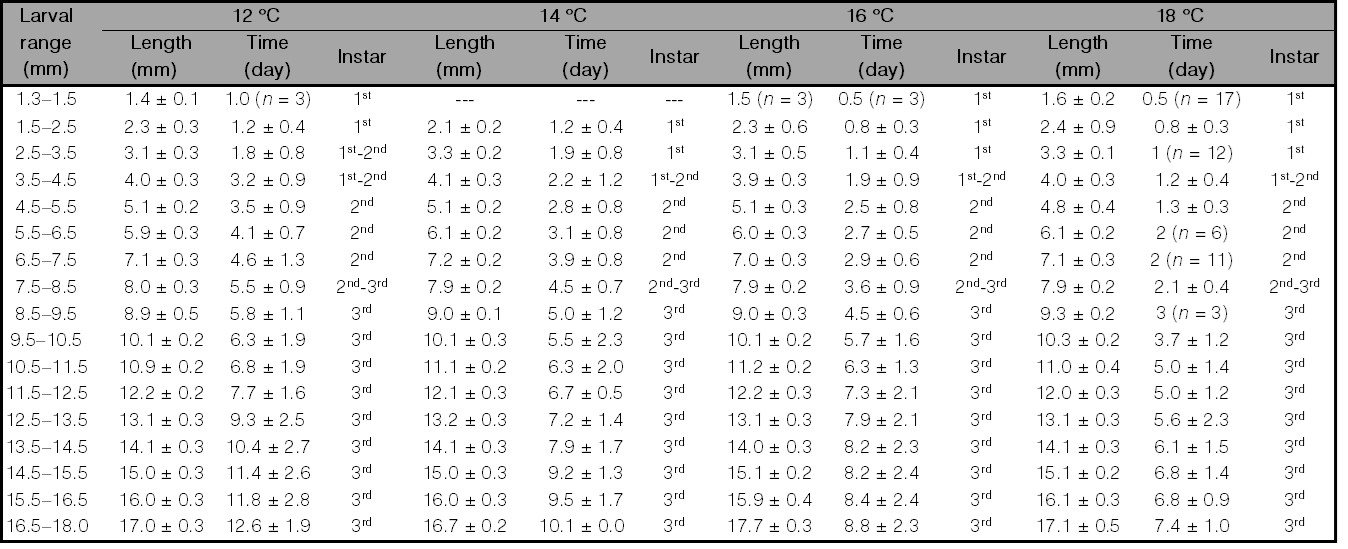
Table III.
Length larval range (mm), average length (mean ± SE, mm), and average developmental time (mean ± SE, days) of larvae of Calliphora vicina reared at different assessed temperature (°C).
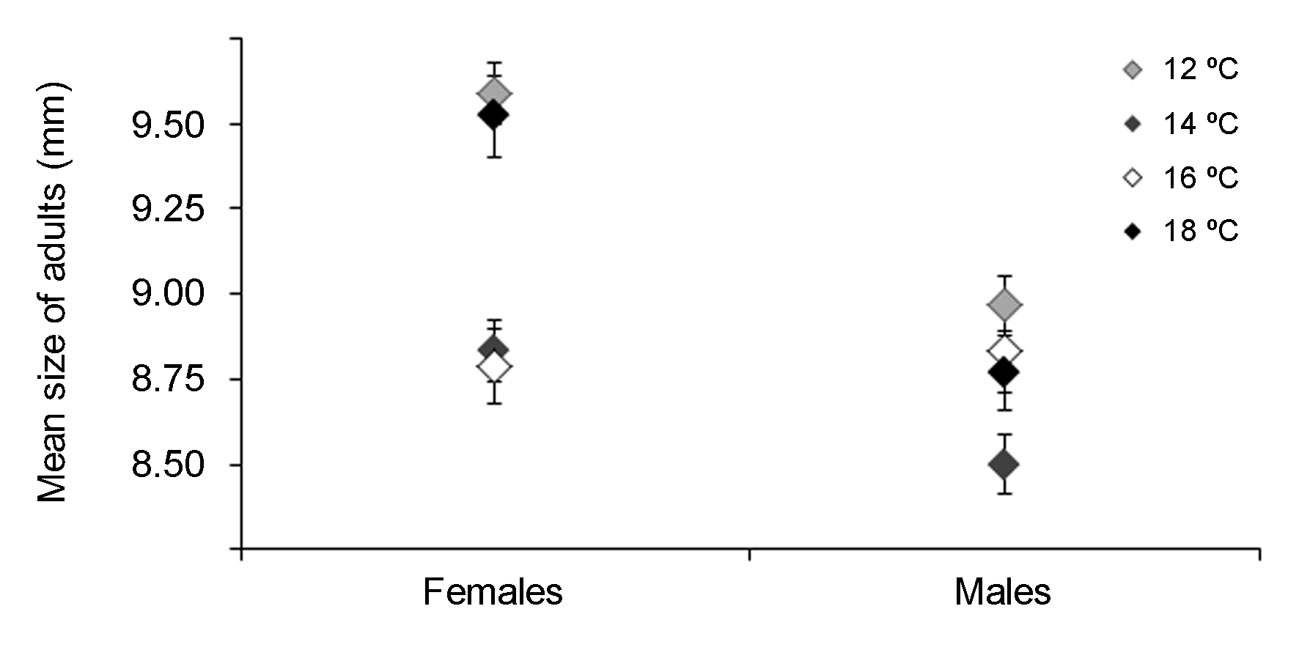
Fig. 3.
Body total length (mean ± SE mm) of females and males Calliphora vicina emerged at different assessed temperatures (°C).
Although developmental times of both sexes were similar in the assessed temperatures, females were larger than males (F = 12.61; df = 1, 788; p= 0.0004), adults’ sizes varied with the temperature (F = 8.91; df = 6, 1,576; p< 0.0001), and interaction between temperature and sex was also significant (F = 3.03; df = 6, 1,576; p= 0.0059). Adults reared at 12 °C were the largest of all, flies reared at 18 °C were mid-sized, and the smallest adults, and with similar sizes, developed at 14 and 16 °C. In the different rearing temperatures, females were bigger than males except females and males reared at 16 °C, which presented similar sizes (Fig. 3). No significant differences were observed in the sex ratios when flies were reared at 12, 16, and 18 °C, whereas it was biased toward females when flies developed at 14 °C (Table IV). Temperature also influenced the proportion of adults’ emergence. The highest survival percentage in the middle-rank of temperatures was observed (38.65% and 29.73% at 16 and 14 °C, respectively), and the highest mortality occurred on the extreme of the temperature ranges (19.00% and 12.61% in 12 and 18 °C, respectively) (Table IV). A higher adult’s size was indirectly related to the reduction in competition over the substrate as the larger flies presented a lower survival rate at 12 and 18 °C (Fig. 3, Table IV).
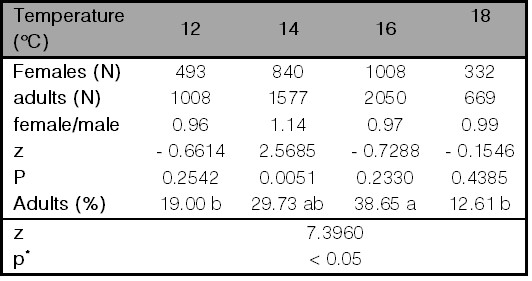
Table IV.
Productivity, sex ratio, and percentage of adults emerged of Calliphora vicina reared at different temperatures (°C).
Comparisons between sexes at each assessed temperature were analyzed using the normal distribution test (p< 0.05). The proportion of adults observed in all temperatures was compared with the proportion test. Different letters indicate significant differences (p< 0.05).
DISCUSSION
The most important result of this study was to estimate age based on size of immature stages of C. vicina from Argentina, relevant data that can be used to calculate minimum PMI, and predict the population parameters of C. vicina in South America. The temperature affected the development of C. vicina, as an increase in temperature from 12 to 18 °C caused an increase in metabolic activity by reducing the growth rates. Besides, the third instars, pupae, and adults developing at 12 and 18 °C were larger than adults at 14 and 16 °C. Although the developmental times for both sexes were similar, sexual size dimorphism was observed, with females being larger than males, with their sizes were also associated with the rearing temperature. Further, body size showed an inverse relationship with survival as flies reared at 12 and 18 °C presented a lower quantity of specimens reaching adulthood, compared to those obtained at 14 or 16 °C.
Forensic entomology relates the changes of larvae age to their lengths based on the rearing temperatures to estimate the PMI. However, despite the importance of accurate and precise data sets, several studies report different developmental rates for the same species. Several studies that published data of C. vicina under similar ranges of temperature have revealed some intraspecific variability in the developmental times and/or immature sizes concerning our data and between them (Reiter, 1984; Anderson, 2000; Donovan et al., 2006; Díaz Martín et al., 2014). This may lead to incorrect estimations in PMI based on observed differences in the time required to reach the pupal stage of C. vicina in several sites and rearing conditions. Developing larvae from Austria reached the pupal stage 4–6 days sooner than those from Argentina at similar temperatures (18–19 °C) (Reiter 1984). Conversely, the larvae and pupae of C. vicina from Argentina reared at 16 °C develop one day earlier than those from Canada and Italy (Anderson, 2000; Defilippo et al., 2013). It is interesting to note that constant temperatures under 16 °C, after the peak in growth, inhibit the viability of pupae by causing a stationary state arrest in samples from Austria and The United Kingdom (Reiter 1984; Donovan et al., 2006). But our results are consistent with data from Spain and Italy, which states C. vicina can develop below 16 °C (Defilippo et al., 2013; Díaz Martín et al., 2014). When comparing our results with that obtained from Spain at similar temperatures, total development duration of C. vicina from Argentina was shortened by 25 days when eggs were incubated at 12 °C and by 28–33 days at 14–16 °C versus when developed at 10 or 15–20 °C (Díaz Martín et al., 2014). Besides, larval development times shortened by 3–4 days in C. vicina from Argentina reared at 14 and 18 °C regarding developing larvae at 14 and 19 °C in London (Donovan et al., 2006). If we consider that Donovan et al. (2006) did not include the pupal or total developmental times, this discrepancy could be broader. In our study, the pupal stage over the other ones was predominant, taking place 60% of the total developmental time. This stage also involved 59–61% of the developmental time when larvae grew on swine meat at different temperatures (Anderson, 2000; Ames & Turner, 2003; Díaz Martín et al., 2014). However, when larvae fed on camel meat, goat meat, or fish, the pupal period was 45–48% of the total time (Hadjer-Kounouz & Kamel, 2017), but only ~39% when the development occurred at higher temperatures (Kamal, 1958). In temperate climates, such as that of the Pampas region in Argentina, there is marked seasonality in the Calliphoridae (Oliva, 2001; Centeno et al. 2002, 2004). Especially, C. vicina is active in the coldest months from last Autumn until the early Spring (Oliva 2001, Centeno et al. 2002, 2004). When the environmental temperatures go high, the fitness of C. vicina could have been affected by their competitors’ species adapted to the warm seasons as Lucilia sericata (Meigen), Sarconesia chlorogaster (Wiedemann) or Cochliomyia macellaria (Fabricius) (Oliva, 2001; Centeno et al., 2002, 2004). Therefore, lower relative abundance is registered in C. vicina compared with these species.
Although differences in the developmental times are registered, there have not been registered changes in the larvae length range (~2–18 mm) between several studies (this study; Reiter, 1984; Kaneshrajah & Turner, 2004; Donovan et al., 2006; Richards et al., 2013; Díaz Martín et al., 2014; Hadjer-Kounouz & Kamel, 2017). However, we registered the first instars smallest than previously observed by others, and significant variability in length between third instars and pupae reared on different temperatures, being third larvae and pupae larger when developed at 14 and 16 °C. Detrimental effects on larval survival and size have previously registered in crowded cultures of C. vicina and C. vomitoria (Linnaeus) (Kaneshrajah & Turner, 2004; Ireland & Turner, 2006; Pellitero & Bordas, 2007). The sizes of third instars and pupae of C. vicina were also affected by the quantity and quality of the feeding substrate. When larvae fed on the decomposed liver, they were significantly smaller those who were fresh and frozen liver (Richards et al., 2013); and when larvae fed brain, heart, or under conditions presenting starvation, they lost weight on the post-feeding stage, by developing the smallest puparia (Kaneshrajah & Turner, 2004; Ireland & Turner, 2006; Pellitero & Bordas, 2007). These results are also supported in L. sericata (Meigen), in which growth rates also vary on different tissues, by growing slower, and were significantly smaller when reared on liver compared to other organs (Clark et al., 2006).
We also observed a discrepancy in size between emerging adults under different temperatures, being smaller in intermediate temperatures. Maybe this fact is an effect of the intraspecific competition on pupal development as bigger sizes were related to a lower percentage of emerged flies. Density, but not diet, also affected the size of adults of C. vomitoria and L. sericata (Ireland & Turner, 2006; Macedo et al., 2018). However, adults of L. sericata were also affected by type of tissue (lung, liver, heart) and species (pig, cow) in which they developed (Clark et al., 2006). These discrepancies in developmental times and sizes could result from either the different experimental designs (temperature regimes, diet) or naturally among populations’ variations. Based on the differences observed in the development of C. vicina from Argentina versus Austria, Canada, Italy, Spain, and the United Kingdom, more studies should be carried out to better understand the physiological characteristics of different populations as this could impact PMI estimation. Finally, we recommend future experiments on the maximum and minimum thresholds of development in C. vicina and other species of forensic importance to ensure comprehensive knowledge of insect life history for PMI estimation.
Acknowledgments
This work was supported by the Agencia Nacional de Promoción Científica y Tecnológica (ANPCyT) [grant number PICT2016-2025] and Universidad Nacional de Quilmes (UNQ) [grant number PUNQ-53/1026] grants from Argentina.
REFERENCES
Amendt, J., Richards, C.S., Campobasso, C.P., Zehner, R., & Hall, M.J.R. (2011) Forensic entomology: applications and limitations. Forensic Science, Medicine, and Pathology, ., 379-392.
Ames, C., & Turner, B. (2003) Low temperature episodes in development of blowflies: implications for postmortem interval estimation. Medical and Veterinary Entomology, 17, 178-186.
Anderson, G.S. (2000) Minimum and maximum development rates of some forensically important Calliphoridae (Diptera). Journal of Forensic Sciences, 45, 824-832.
Centeno, N.D., Almorza, D., & Arnillas, C. (2004) Diversity of Calliphoridae (Insecta: Diptera) in Hudson, Argentina. Neotropical Entomology, 33, 387-390.
Centeno, N.D., Maldonado, M., & Oliva, A. (2002) Seasonal patterns of arthropods occurring on sheltered and unsheltered pig carcasses in Buenos Aires Province (Argentina). Forensic Science International, 126, 63-70.
Clark, K., Evans, L., & Wall, R. (2006) Growth rates of the blowfly, Lucilia sericata, on different body tissues. Forensic Science International, 156, 145-149.
Defilippo, F., Bonilauri, P., Dottori, M., & Bonilauri, P. (2013) Effect of temperature on six different developmental landmarks within the pupal stage of the forensically important blowfly Calliphora vicina (Robineau-Desvoidy) (Diptera: Calliphoridae). Journal of Forensic Sciences, 58, 1554-1557.
Di Rienzo, J.A., Casanoves, F., Balzarini, M.G., Gonzalez, L., Tablada, M., & Robledo, C.W. (2017) InfoStat, Grupo InfoStat (accessed December 23, 2019).
Díaz Martin, B., Rodríguez, A.L., & Bordas, M.I.S. (2014) Primeros resultados sobre desarrollo de Calliphora vicina (Diptera: Calliphoridae) bajo condiciones controladas de temperatura. Ciencia Forense, 11, 241-260.
Donovan, S.E., Hall, M.J.R., Turner, B.D., & Moncrieff, C.B. (2006) Larval growth rates of the blowfly, Calliphora vicina, over a range of temperatures. Medical and Veterinary Entomology, 20, 106-114.
Grassberger, M., & Reiter C. (2001) Effect of temperature on Lucilia sericata (Diptera: Calliphoridae) development with special reference to the isomegalen- and isomorphen-diagram. Forensic Science International, 120, 32-36.
Greenberg, B. (1991) Flies as forensic indicators. Journal of Medical Entomology, 28, 565-577.
Hadjer-Kounouz, S., & Kamel, L. (2017) Development of Calliphora vicina (Robineau-Desvoid) (Diptera: Calliphoridae) under different biotic and abiotic conditions. Journal of Entomology and Zoology Studies, ., 683-691.
Ireland, S., & Turner, B. (2006) The effects of larval crowding and food type on the size and development of the blowfly, Calliphora vomitoria. Forensic Science International, 159, 175-181.
Johnson, A.P., & Wallman, J.S. (2014) Effect of massing on larval growth rate. Forensic Science International, 241, 141-149.
Kamal, A.S. (1958) Comparative study of thirteen species of sarcosaprophagous Calliphoridae and Sarcophagidae (Diptera) I, Bionomics. Annals of the Entomological Society of America, 51, 261-270.
Kaneshrajah, G, & Turner, B. (2004) Calliphora vicina larvae grow at different rates on different body tissues. International Journal of Legal Medicine, 118, 242-244.
Kotzé, Z., Villet, M.H., & Weldon, C.W. (2015) Effect of temperature on development of the blowfly, Lucilia cuprina (Wiedemann) (Diptera: Calliphoridae). International Journal of Legal Medicine, 129, 1155-1162.
Macedo, M.P., Arantesa, L.C., & Tidon, R. (2018) Sexual size dimorphism in three species of forensically important blowflies (Diptera: Calliphoridae) and its implications for postmortem interval estimation. Forensic Science International, 293, 86-90.
Maldonado, A.M., & Centeno, N. (2003) Quantifying the potential pathogens transmission of the blowflies (Diptera: Calliphoridae). Memórias do Instituto Oswaldo Cruz, 98, 213-216.
Marchenko, M.I. (2001) Medico-legal relevance of cadaver entomofauna for the determination of the time since death. Forensic Science International, 120, 89-109.
Mariluis, J.C., & Schnack, J. (2002) Calliphoridae de la Argentina, Sistemática, ecología e importancia sanitaria (Diptera, Insecta). Actualizaciones en Artropodología Sanitaria Argentina (ed. Salomon O.S.), pp 23-37. Fundación Mundo Sano, Buenos Aires, Argentina.
Matuszewski, S., Konwerski, S., Frątczak, K., & Szafałowicz M. (2014) Effect of body mass and clothing on decomposition of pig carcasses. International Journal of Legal Medicine, 128, 1039-1048.
Niederegger, S., Pastuschek, J., & Mall, G. (2010) Preliminary studies of the influence of fluctuating temperatures on the development of various forensically relevant flies. Forensic Science International, 199, 72-78.
Oliva, A. (2001) Insects of forensic significance in Argentina. Forensic Science International, 120, 145-154.
Parker, J., & Johnston, L. (2006) The proximate determinants of insect size. Journal of Biology, ., 15.
Pellitero, J.M., & Bordas, M.I.S. (2007) Development of Calliphora vicina Robineau-Desvoidy (Diptera, Calliphoridae) under starvation conditions. Boletín Sociedad Entomológica Aragonesa, 41, 413-417.
Rabêlo, K.C.N., Thyssen, P.J., Salgado, R.L., Araújo, M.S.C., & Vasconcelos, S.D. (2011) Bionomics of two forensically important blowfly species Chrysomya megacephala and Chrysomya putoria (Diptera: Calliphoridae) reared on four types of diet. Forensic Science International, 210, 257-262.
Reiter C. (1984) Zum Wachstumsverhalten der Maden der blauen Schmeiss fliege Calliphora vicina. The growth behaviour of the maggots of the blue blowfly Calliphora vicina. Zeitschrift fur Rechtsmedizin, 91, 295-308.
Richards, C.S., Rowlinson, C.C., Cuttiford, L., Grimsley, R., & Hall, M.J.R. (2013) Decomposed liver has a significantly adverse effect on the development rate of the blowfly Calliphora vicina. International Journal of Legal Medicine, 127, 259-262.
Salimi, M., Rassi, Y., Oshaghi, M., Chatrabgoun, O., Limoee, M., & Rafizadeh, S. (2018) Temperature requirements for the growth of immature stages of blowflies species, Chrysomya albiceps and Calliphora vicina, (Diptera: Calliphoridae) under laboratory conditions. Egyptian Journal of Forensic Sciences, ., 1-6.
Stillwell, R.C., & Fox, C.W. (2007) Environmental effects on sexual size dimorphism of a seed-feeding beetle. Oecologia, 153, 273-280.
Tarone, A.M., Picard, C.J., Spiegelman, C., & Foran, D.R. (2011) Population and temperature effects on Lucilia sericata (Diptera: Calliphoridae) body size and minimum development time. Journal of Medical Entomology, 48, 1062-1068.
Vinogradova, E.B., & Reznik, S.Y. (2015) Influence of constant and changing temperatures on the larval development of Calliphora vicina (Diptera: Calliphoridae). Acta Societatis Zoologicae Bohemicae, 79, 149-154.
Whitworth, T. (2012) Identification of Neotropical blow flies of the genus CalliphoraRobineau-Desvoidy (Diptera: Calliphoridae) with the description of a new species. Zootaxa, 3209, 1-27.
Woods, H.A., Makino, W., Cotner, J.B., Hobbie, S.E., Harrison, J.F., Acharya, K., & Elser, J.J. (2003) Temperature and the chemical composition of poikilothermic organisms. Functional Ecology, 17, 237-245.