Articles
Revisiting the ambush bug genus Neoanthylla Kormilev, 1951 (Reduviidae, Phymatinae)
Revisitando el género de chinches emboscadoras Neoanthylla Kormilev, 1951 (Reduviidae, Phymatinae)
Revisiting the ambush bug genus Neoanthylla Kormilev, 1951 (Reduviidae, Phymatinae)
Revista de la Sociedad Entomológica Argentina, vol. 82, núm. 1, 2023
Sociedad Entomológica Argentina
Recepción: 28 Julio 2022
Aprobación: 01 Febrero 2023
Abstract: Neoanthylla (Kormilev, 1951) is a Neotropical genus of Phymatinae with three valid species: Neoanthylla bucki (Kormilev), Neoanthylla horvathi (Handlirsch), and Neoanthylla peruviana Kormilev. Morphologically, it is characterized by the elongated and oval profemur with the cuticular surface smooth and shiny; the procoxa long and subequal to the mesofemur; the prosternal carina long, reaching the anterior margin of the mesosternum; and the abdomen posteriorly entire. Neoanthylla species are scarcely documented in the literature, and the genital structures are unknown. In addition, the Brazilian species (N. bucki and N. horvathi) are known only by female specimens. Here, Neoanthylla is revisited, the diagnoses of the valid species are improved, images of the holotypes of N. horvathi and N. peruviana are provided, and the male of N. horvathi is described for the first time, including genital characters.
Keywords: Brazil, Heteroptera, Neotropical region, South America.
Resumen: Neoanthylla (Kormilev, 1951) es un género Neotropical de Phymatinae con tres especies válidas: Neoanthylla bucki (Kormilev), Neoanthylla horvathi (Handlirsch) y Neoanthylla peruviana Kormilev. Morfológicamente se caracteriza por el profémur alargado y ovalado con la superficie cuticular lisa y brillante; las procoxas largas y subiguales al mesofémur; la carina prosternal larga, alcanzando el margen anterior del mesosternum; y el abdomen entero posteriormente. Las especies de Neoanthylla están escasamente documentadas en la literatura y las estructuras genitales son desconocidas. Además, las especies brasileñas (N. bucki y N. horvathi) son conocidas sólo por hembras. Aquí, se revisa Neoanthylla, se actualizan las diagnosis de las especies válidas, se brindan imágenes de los holotipos de N. horvathi y N. peruviana, y se describe por primera vez el macho de N. horvathi, incluyendo los caracteres genitales.
Palabras clave: Brasil, Heteroptera, Región Neotropical, Sudamérica.
INTRODUCTION
Ambush bugs (Reduviidae, Phymatinae) are a charismatic subfamily composed of approximately 29 genera and more than 300 species, grouped into four tribes: Carcinocorini, Macrocephalini, Phymatini, and Themonocorini (Schuh & Weirauch, 2020). Morphologically, it is a very conspicuous group, characterized by the angular head with the bucculae surpassing the base of the labium, the proleg usually chelate to subchelate (Zhang et al., 2016), the reduced or absent protarsus, and the particular venation of the hemelytra (Weirauch et al., 2014; Schuh & Weirauch, 2020). Phymatini is a monophyletic group (Masonick et al., 2017) composed of five genera and approximately 115 species worldwide (Froeschner & Kormilev, 1989).
NeoanthyllaKormilev, 1951 is a rarely collected genus. It was described as a subgenus of Phymata, including the new species Phymata buckiKormilev, 1951 as the type species. Kormilev (1962a) elevated Neoanthylla to genus-level, considering that, despite being similar to Anthylla Stål, 1876, several characters were conspicuously different. He observed differences in the oval, elongated profemur nearly three times longer than wide, with the cuticular surface smooth and shiny and without granulations; the procoxa long and subequal to the mesofemur; the anterior branch of the prosternum longer, reaching the anterior margin of the mesosternum; and the abdomen entire posteriorly rather than bilobed. Currently, the genus comprises three species: N. bucki and Neoanthylla horvathi (Handlirsch, 1898) from Brazil, and Neoanthylla peruvianaKormilev, 1964 from Peru. The species are currently separated using the structure of the profemur and the shape of connexiva.
Neoanthylla peruviana is known from a male, and it is conspicuously different from the other species. Neoanthylla bucki and N. horvathi are known only from females and the principal criteria to separate them are the shape of the posterior lobe of the pronotum and the curvature of the lateral margin of the connexiva, with the latter character often barely perceptible. Kormilev (1964) described sexual dimorphism in the genus in the length and structure of the antennal segments, surprisingly he assumed this condition was constant in all three valid species, without a known description of male specimens of N. bucki or N. horvathi or the female of N. peruviana.
This study revisits the genus Neoanthylla, precipitated by the collection of one male specimen in Rio Grande do Sul (Brazil). Here, we describe for the first time the male of N. horvathi, including genitalic characters. In addition, we offer diagnoses for all valid species, a taxonomic key to species, images of the holotypes or additional examined material, and discuss the sexual dimorphism, and a new record registered by images of adult and immature specimens in iNaturalist.
MATERIALS AND METHODS
Examined specimens. The description of the male of N. horvathi is based on a single specimen deposited in the entomological collection of the Museo de La Plata, Buenos Aires, Argentina (MLPA). Types and additional specimens were examined from high-resolution images of deposited material in the following institutions: Hungarian Natural History Museum, Budapest, Hungary (HNHM); Museu Anchieta, Porto Alegre, Rio Grande do Sul, Brazil (MGAP); National Museum of Natural History, Washington D.C., USA (NMNH), and Zoologische Staatssammlung, Munich, Germany (ZSM). Adult and immature specimens of Neoanthylla were photographed by Felipe Toro (Colombia) and registered in the iNaturalist website (https://www.inaturalist.org/observations/96723441).
Genital dissections and imaging. Dissections and imaging of structures follow Forero & Weirauch (2012). We used a dissecting scope Olympus SZ61 to dissect and manipulate the genitalia. For documentation we used a Canon EOS Rebel T7i digital camera, equipped with a Helicon Remote software v3.9.11W. Stacking was accomplished using Helicon Focus v8.1.0. For image editing, Adobe Photoshop CS6 v13.1.2 was used.
Terminology. For external structures, we follow Kormilev (1962b), Swanson (2019), and Schuh & Weirauch (2020). For male genitalia, we follow the terminology of Davis (1957, 1966) and Forero & Weirauch (2012).
Georeferencing and mapping. All examined specimens lacked geographical coordinates in their locality labels. These localities were georeferenced using the Global Gazetteer v.2.3 (http://www.fallingrain.com/world/index.html). A distribution map was created using QGIS ver. 3.24.3 (QGIS, 2022).
RESULTS
NeoanthyllaKormilev, 1951
Phymata (Neoanthylla) Kormilev, 1951, 2: 57, type species: Phymata bucki Kormilev [description as subgenus and sp. nov]; Kormilev, 1952, 23: 123, 128 [dorsal habitus image].
NeoanthyllaKormilev, 1962a, 89: 468 [elevated to genus, N. horvathi n. status, key to species]; Kormilev, 1964, 65: 138 [sp. nov.]. Froeschner & Kormilev, 1989, 6: 41–42 [key to species].
Diagnosis. Neoanthylla can be distinguished from other genera by the enlarged, ellipsoidal profemur with a flat and polished lateral surface and smooth dorsal margin (Fig. 1C), and the anterior branch of the mesosternal cross long, reaching the prosternum (Fig. 1E).
Remarks. According to Kormilev (1962a), its closely allied genera are Anthylla Stål, 1876 and KelainocorisKormilev, 1963, as they share the meso- and metatibiae convexly rounded on the dorsal surface, and laterally without rims. However, Neoanthylla can be distinguished by the rounded apex of the abdomen (bilobed in Anthylla, straight or bilobed in Kelainocoris), the shape and texture of the anterior femora (subtriangular, convex, and granulated in Anthylla; subtriangular but rough in Kelainocoris), and the longer mesosternal cross (not reaching prosternum in Anthylla; without information about this structure in Kelainocoris).

Fig. 1.
Neoanthylla horvathi, male. A. lateral view; B. dorsal view; C. proleg detail, lateral view; D-F. head and thorax, D. lateral view, E. ventral view, F. dorsal view; G. connexiva, lateral view. Arrows indicate key characters.
Key to species of Neoanthylla (based on Froeschner & Kormilev 1989)
l. Profemur with a longitudinal carina on external region, connexiva of abdominal segments II–IV with conspicuous protruding processes on posterolateral angles ....................................... N. peruviana
1´. Profemur smooth, without longitudinal carina, connexiva of abdominal segments II–IV with short processes on posterolateral angles ................................................................................................................. 2
2. Both processes of lateral expansion of posterior lobe acute, connexiva of abdominal segments III and IV with posterolateral angles produced as an angulated short protuberance...................... N. horvathi
2´. Anterior processes of lateral expansion of posterior lobe acute and posterior processes rectangular, connexiva of abdominal segments III and IV with posterolateral angles produced as a blunt tubercle N. bucki
Neoanthylla bucki (Kormilev)
Phymata (Neoanthylla) buckiKormilev, 1951, 2: 56 [Brazil]; Kormilev 1952, 23: 123, 128.
Neoanthylla bucki, Kormilev 1962a, 89: 468; Froeschner & Kormilev 1989, 6: 41–42 [key to species].
Diagnosis. Neoanthylla bucki can be diagnosed by the shape of the processes on the posterior lobe of the pronotum, the anterior acute and posterior rectangular (Fig. 3A); and the posterolateral angles of connexiva of abdominal segments II–IV with a small tuberculiform protuberance (Fig. 3A).
Distribution. Brazil: Rio Grande do Sul and Santa Catarina.
Examined material. Holotype: (from high quality images): 1 female, BRAZIL: RS, Vila Oliva, 28.i.1947 / Holotipo / Phymata buckiKormilev, 1951 / C J Drake Coll. 1956 / Neoanthylla bucki Kormilev, 1955 / Collection N. Kormilev.
Other specimens (high quality images): 1 female, BRAZIL: RS, Vila Oliva (Caxias do Sul), RS. 24.II.1952 (MGAP); 1 female, Vila Oliva (Caxias do Sul), RS. 13.I.1960. Pe. Pio Buck col. (MGAP); 2 females, Santa Catarina, 8.32-1.33, Coll. A. Jöhnk, Thaden Holst / Neoanthylla bucki, det. Kormilev, 1956 (ZSM).
Remarks. The pronotal processes on the lateral expansion of the posterior lobe have been used as a taxonomic character to separate N. bucki (anterior process acute and posterior rectangular) from N. horvathi (both processes acute) (Kormilev, 1962a; Froeschner & Kormilev, 1989). However, comparing the images and specimen, the differences appear to be minor. Moreover, we observed slight variations from side to side in the same specimen. For this reason, this character should be carefully considered or complemented with other characteristics.
Locating the type specimen of N. bucki was not easy. We searched for “Kormilev´s holotypes” without success, in Argentinean type-material catalogues (Museo Argentino de Ciencias Naturales "Bernardino Rivadavia" and Instituto Fundación Miguel Lillo); in Brazilian entomology collections (Museu de Ciencias Naturais da Fundação Zoo-Botânica do Rio Grande do Sul and MGAP); and the ZSM. Finally, we received the communication from one of the Phymatinae specialists saying than the holotype is currently deposited in the NMNH, information confirmed by Thomas Henry, Heteroptera curator of this institution.
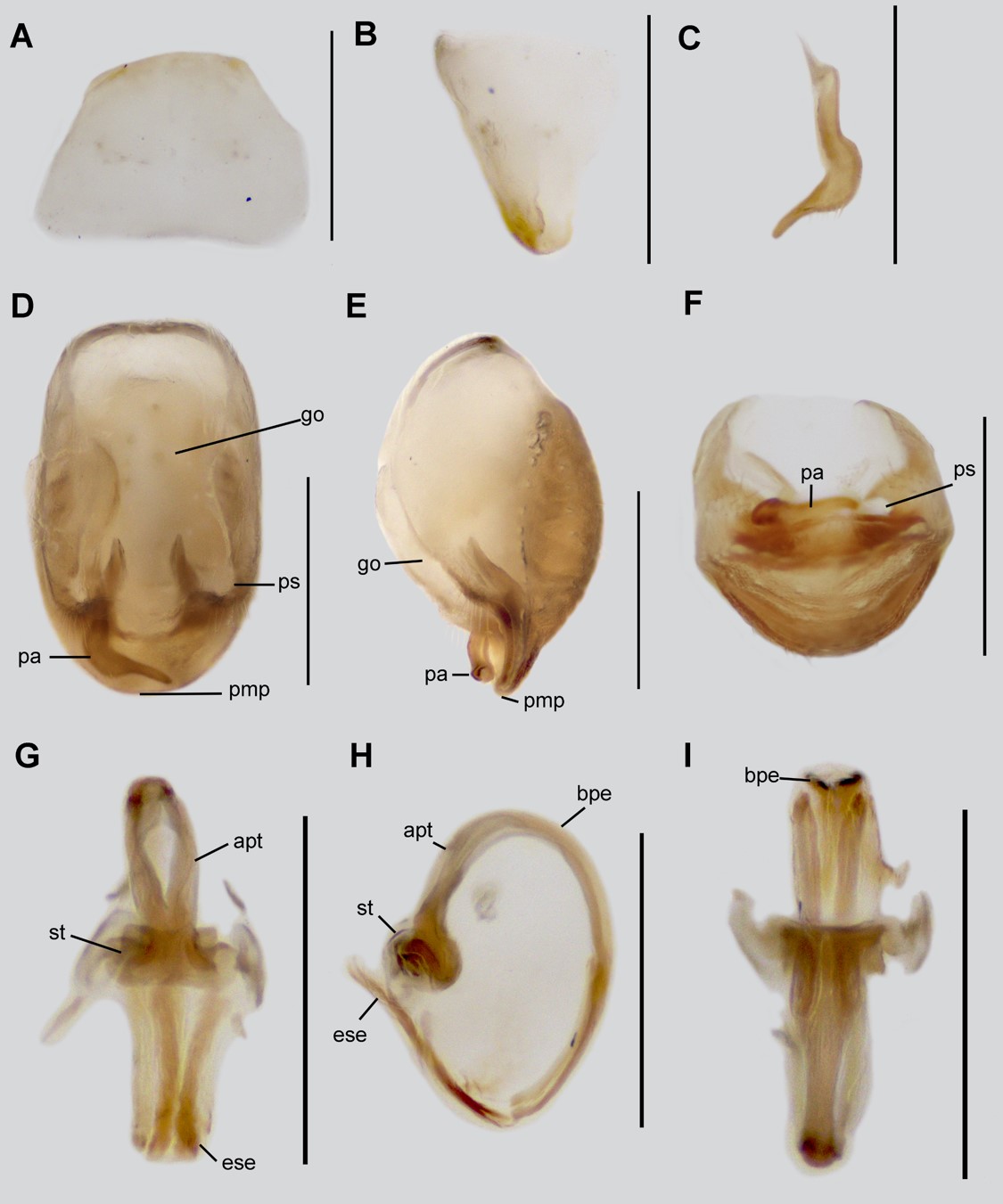
Fig. 2.
Neoanthylla horvathi, male genitalia. A-B. Sternite 8. A. ventral view; B. lateral view; C. Paramere; D-F. Pygophore; D. dorsal view; E. lateral view; D. caudal view; G-I. Aedeagus; G. dorsal view; H. lateral view; I. ventral view. Abbreviations: bpe, basal plate extension; go, genital opening of pygophore; pa, paramere; pc, processus capitati; pmp, posterior margin of pygophore; ps, paramere socket; st, struts. Scale bar: 0.5 mm.
Neoanthylla horvathi (Handlirsch)
Phymata horvathiHandlirsch, 1898, 48: 382 [Brazil].
Neoanthylla horvathi, Kormilev, 1962b, 89: 471; Froeschner & Kormilev, 1989, 6: 41–42 [key to species].
Diagnosis. Very similar to N. bucki, but it can be distinguished by the acute processes of the lateral expansion of the posterior lobe of pronotum (Figs. 1B, 3B), and shortly produced posterolateral angles of connexiva of abdominal segments III–IV (Figs. 1G, 3B).
Redescription. MALE: Total length: 8.70 mm (clypeus-apex abdomen), maximum width 3.40 mm (apex of connexivum). COLORATION: Head (Figs. 1D, 1F): Dark brown with several whitish punctures, mandibular plates, and postocular region in dorsal view pale brown, ocelli orange. Antenna: Scape dark brown, pedicel and distiflagellomere brown, basiflagellomere pale brown. Labium: dark brown. Thorax (Figs. 1B, 1D, 1F): Pronotum pale brown with several brown and dark brown spots, whitish punctures on whole structure, slightly more conspicuous on longitudinal medial region, posterior margin of posterior lobe whitish; scutellum orange, supracoxal lobes pale brown with several whitish punctures. Legs (Figs. 1A, 1C): Procoxa brown with a medial longitudinal dark brown band; protrochanter pale brown, profemur and tibia orange polished, protarsus yellowish; meso- and metalegs yellowish, meso- and metafemur with a pair of dark brown transversal bands on the medial and apical region respectively, metatibia with a slightly transversal brown band. Hemelytron: Corium brown, lateral margins pale brown, with several whitish punctures; membrane dark brown. Abdomen: Pale brown, medial longitudinal carina dark brown, pygophore yellowish. VESTITURE: Body with rather sparse, fine, suberect setae; surface covered by whitish protruding punctures. Legs: Apex of meso- and metatibia with dense, long setae ventrally. Abdomen: Connexiva of the abdominal segments II to IV with conspicuous tubercles on the posterolateral angles. STRUCTURE: Head (Figs. 1D, 1F): Pentagonal in dorsal view, subquadrate in lateral view; anteocular region with a bifurcate tubercle behind antennophores, clypeus slightly elevated over mandibular plates; postocular region with a pair of elevated longitudinal carinas, ocelli positioned laterally and with a pair of short tubercles on them. Eyes: Quarter hemispherical in dorsal view, ovoid in lateral view. Antenna: scape 0.3 mm; pedicel 0.4 mm; basiflagellomere 0.3 mm; distiflagellomere 2.20 mm. Labium: First visible labial segment as long as the second visible labial segment. Thorax (Figs. 1D-F): Collar with acute angles, anterior pronotal lobe trapezoidal; lateral notch absent; posterior pronotal lobe hexagonal, slightly depressed medially, disc with paired longitudinal medial carinas, humeral angles with an acute protruding process and another small acute process lateral behind, posterior margin smooth, the posterior angles with a short tubercle. Scutellum triangular, anterior margin with a medial tubercle. Metaepimeron with a short tubercle. Mesosternum with medial transversal carina. Legs: Procoxa cylindrical and thick, with a tubercle on the proximal dorsal region; protrochanter nearly oval; profemur nearly oval-elongate, with an apical acute short process, ventrally with a row of short processes; protibia curved, falcate, ventrally with a row of short acute processes. Meso- and metacoxa oval and globose; meso- and metatrochanter pentagonal; meso- and metafemur cylindrical, mesofemur slightly wider and shorter than metafemur; meso- and metatibia cylindrical, nearly as long as meso- and metafemur. Tarsi two-segmented, the first tarsal segment conspicuously shorter than the second, pretarsal claws simple, semiparallel, meso- and metatarsal claws orthogonal and simple. Hemelytron: Macropterous, wings surpassing the abdominal apex. Abdomen (Fig. 1G): Nearly rectangular, elongated, and basally slightly narrow; abdominal segments V and VI with lobate ultraconnexivum, in lateral view the anterior margin of the ultraconnexivum on the abdominal segment V is acute and on the abdominal segment VI is sinuate; abdominal sternites with a medial longitudinal carina. Sternite VIII only sclerotized ventrally, trapezoidal, anterior and posterior margin smooth. MALE GENITALIA: Pygophore elongate ovoid (Fig. 2D); ventral surface rugose (Fig. 2E); proctiger membranous, with the anterior portion thinner than the posterior one; posteromedial margin flat (Fig. 2E); genital opening (pgo) not separated in anterior and posterior openings; area surrounding paramere socket (ps) with short erect setae (Fig. 2D). Paramere elongated, slightly wider on the medial region, apical half compressed apically, curving apically, and long delicate setae on dorsal and ventral surfaces of apical half before curvature (Fig. 2C). Phallus with “processus capitati” (=promotor apodeme, pc) fused on medial line (Fig. 2G), basal plate extension (=pedicel, bpe) strongly curved in lateral view (Fig. 2H), phallosoma without dorsal phallothecal sclerite (Fig. 2H), struts elongated (Figs. 2G-I, str).
FEMALE (Fig. 3B): Similar to male, except as follows: basiflagellomere conspicuously longer than the pedicel and hemelytron not surpassing the abdominal apex. The genitalic structures were not examined.
Distribution. Brazil: Rio Grande do Sul and Minas Gerais.
Examined material. Holotype (from high quality images): 1 female, BRAZIL, Minas Gerais, 1897, ex-coll. of Fruhstorfer / Coll. Mus. Nat. Hung. / Phymata horvathi Hand. / TYPUS / Horvathi det. Handlirsch / Neoanthylla horvathi det. Handlirsch, N. Kormilev 55 (HNHM).
Other specimens: 1 male, BRAZIL: RS, São Francisco de Paula, 09-11 vii 2021, R. Brugnera. leg. (MLPA).
Remarks. This is the first formal description of the male of N. horvathi and of the internal genitalic structures within of the genus Neoanthylla. In addition, we confirmed the sexual dimorphism of the antennae in N. horvathi with the basiflagellomere conspicuously shorter than the pedicel in the male and longer than the pedicel in the female.
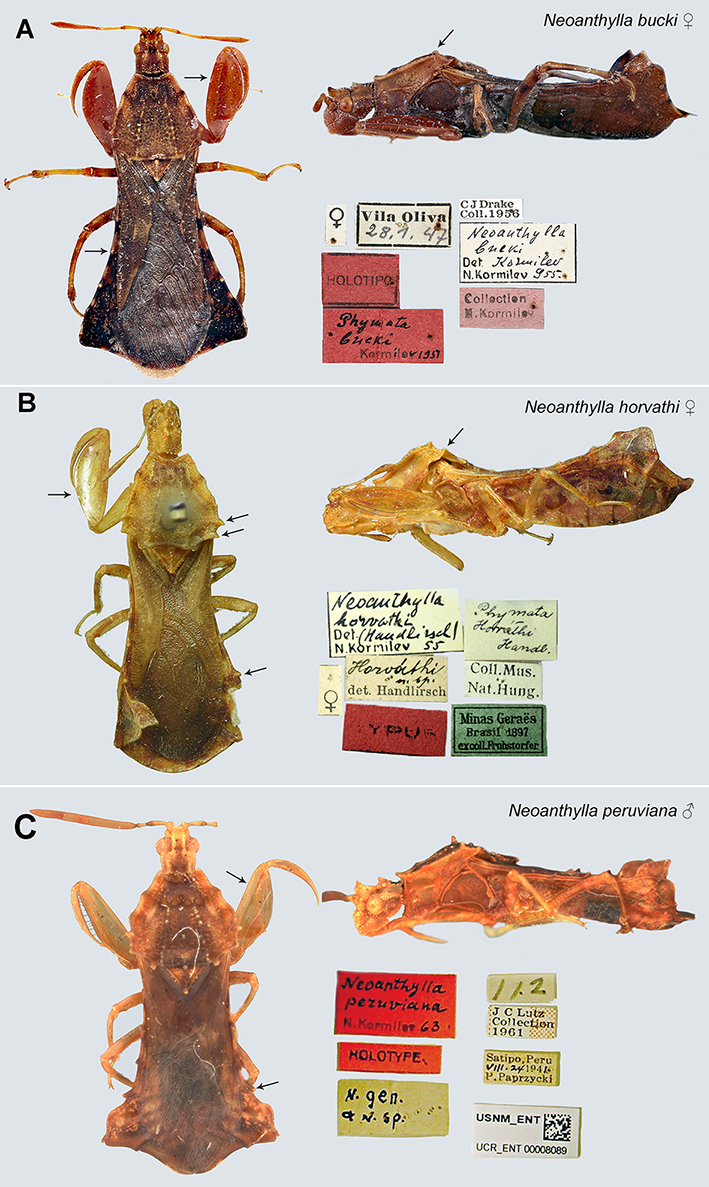
Fig. 3.
Neoanthylla type material. A. Neoanthylla bucki. Female holotype, lateral and dorsal views, labels, images by Alyssa Seemann (NMNH); B. Neoanthylla horvathi. Female holotype, lateral and dorsal views, labels, images by A. Somogyi (HNHM); C. Neoanthylla peruviana. Male holotype, lateral and dorsal views, labels. Arrows indicate key characters.
Neoanthylla peruviana Kormilev
Neoanthylla peruvianaKormilev, 1964, 65: 138 [Peru]; Froeschner & Kormilev, 1989, 6: 41–42 [key to species].
Diagnosis. Easily separated from the other two species by the longer profemur (Fig. 3C), the concave outer region of the profemur, the longitudinal carina on the profemur, and the finger-like produced posterolateral angles of the connexiva of abdominal segments II–IV (Fig. 3C).
Distribution. Peru: Satipo.
Examined material (from high quality images). Holotype male, PERU: Peru, Satipo, P. Paprzycki col. VIII. 24, 1941 / J.C. Lutz collection / Neoanthylla peruviana Kormilev 63 / n. gen n. sp / USNM_ENT UCR_ENT00008089 (USNM).
Remarks. Neoanthylla peruviana is morphologically the most distinct species within the genus, with a longitudinal carina on the profemur and cuticular processes more protruding and conspicuous than in the other species. Kormilev (1964) described a male holotype of N. peruviana, but the female was not described or included in the examined material (see below).
DISCUSSION
Male and female confusing descriptions. The information and documentation of Neoanthylla species are particularly scarce. Kormilev (1951) described Phymata .Neoanthylla. bucki from a female and offered diagnostic characters, a short description, and a key to separate it from the allied Anthylla. A year later, Kormilev (1952) included a dorsal image of Phymata (Neoanthylla) bucki holotype, without additional comments about the genus or species. Subsequently, he elevated Neoanthylla to genus, and diagnosed it, transferred Phymata horvathi to the genus, redescribed N. bucki and N. horvathi (emphasizing on the differences between them), and offered a key to genera and species and illustrations of the pronotum, propleuron, connexivum and left profemur and tibia (Kormilev, 1962a). He also confirmed that males of both species were unknown.
In 1964, Kormilev described N. peruviana. However, in the introduction, he stated “All three species of Neoanthyllahave sexual dimorphism...”, but N. bucki and N. horvathi were known only from females. Similarly, N. peruviana, was based solely on the male holotype (females were not even included in examined material). The illustrations accompanying the description of N. peruviana are similarly confusing: Figure 1 illustrated the connexiva of the ostensibly unknown male of N. bucki (in accordance with the figure legend), but he did not include a formal description or mention the examined specimens in his study. In addition, Figures 1–3 are illustrations of the connexiva: N. bucki and N. horvathi in a ventral view, and N. peruviana in a dorsal view; and this inconsistent treatment tarnishes any comparisons made using these figures. The description of the sexual dimorphism in the antennal segments and the illustrations offered by Kormilev lead us to think that he actually examined additional material of Neoanthylla, but for some inexplicable reason, he did not include the males of the species known only from females. It also seems possible that Kormilev simply assumed the sexual differences between the species would remain constant intraspecifically. However, this assumption seems premature, given that antennal ratios are not reliably consistent in other phymatine species (Kormilev, 1962a; Swanson, 2013; Masonick & Weirauch, 2020).
Finally, Neoanthylla was included in Froeschner & Kormilev’s (1989) catalog with a key to genera and species but without additional descriptions or comments. Moreover, the key to species used different characters than the first one created by Kormilev (1964) and included contradictions with previous data for the species (e.g., texture of profemur).
Morphological complexity and problematic taxonomic characters. Phymatinae is a group with a complex morphology, evidenced by the conspicuous cuticular processes and ornamentations. In this regard, Kormilev´s descriptions can be difficult to follow because he applied a particular terminology, including a unique system of acronyms, to thoracic and abdominal expansions. In addition to this, there is a high level of plasticity among the cuticular processes and sexual dimorphism, observed in other related genera to Neoanthylla (e.g., Phymata, Masonick & Weirauch, 2020).
Considering these aspects, this study has two fundamental limitations: on the one hand, the extreme scarcity of collected specimens makes impossible to compare the intraspecific variability. To evaluate the taxonomic value of some important characters within Neoanthylla is very difficult without series of both male and female specimens. On the other hand, some characters are difficult to observe and interpret, owing to external factors like pinning position or state of preservation. In the case of the structure of the sternal carinae (=anterior branch of the prosternum), one of the most used characteristics at the generic and specific levels, we had difficulty observing the variation in Neoanthylla specimens examined because this structure is usually covered by the prolegs.
The genitalia in Phymatinae apparently exhibits little variability within the subfamily, at least within the most studied genera (Kormilev, 1962a;Swanson, 2019). Given this trend, the genital character complex is usually not considered informative and is rarely documented, with a few exceptions: Carcinocoris (Rabitsch et al. 2006), Phymata (Kormilev, 1962a), Lophoscutus (Kormilev & van Doesburg, 1992), and Macrocephalus (Kormilev, 1962b). However, we think that recent taxonomic studies should document as many morphological structures as possible because it is not always predictable which character systems might become important in the future (Ang et al., 2013). In the case of Neoanthylla, given this possibility, we document the male genitalia here for the first time.
Northern South American immature specimen record. Recently, a record of one adult and one nymph of Neoanthylla from western Andean cordillera in Colombia (Envigado, Antioquia, Colombia) (Fig. 4 A–C) appeared on the iNaturalist website. This seems to be the northernmost record of the genus, which until now, was only known from Peru and southern Brazil (Fig. 5). By the position of the bug in the image, it is impossible to identify the adult specimen to species level with reasonable certainty, and it is worse for the nymph, which life stages were unknown until now. However, the adult and nymph exhibit the diagnostic characters of the genus, particularly on the profemur. In the nymph, the processes on the head, anterodorsal lobe of the thorax, and abdomen are more conspicuous than in the adult (including dorsal midlongitudinal processes, which are only present in immatures), but the posterodorsal lobe processes (relevant for species identification) are not visible in the specimen of the image. Considering these aspects, we treat this record with caution. Future studies with additional material could help to confirm the identity and distributional record.

Fig. 4.
Neoanthylla sp. recorded in Colombia. A. nymph specimen, lateral view; B. adult and immature specimens; C. nymph, frontal view.
©Felipe Toro (https://www.inaturalist.org/photos/160721025).
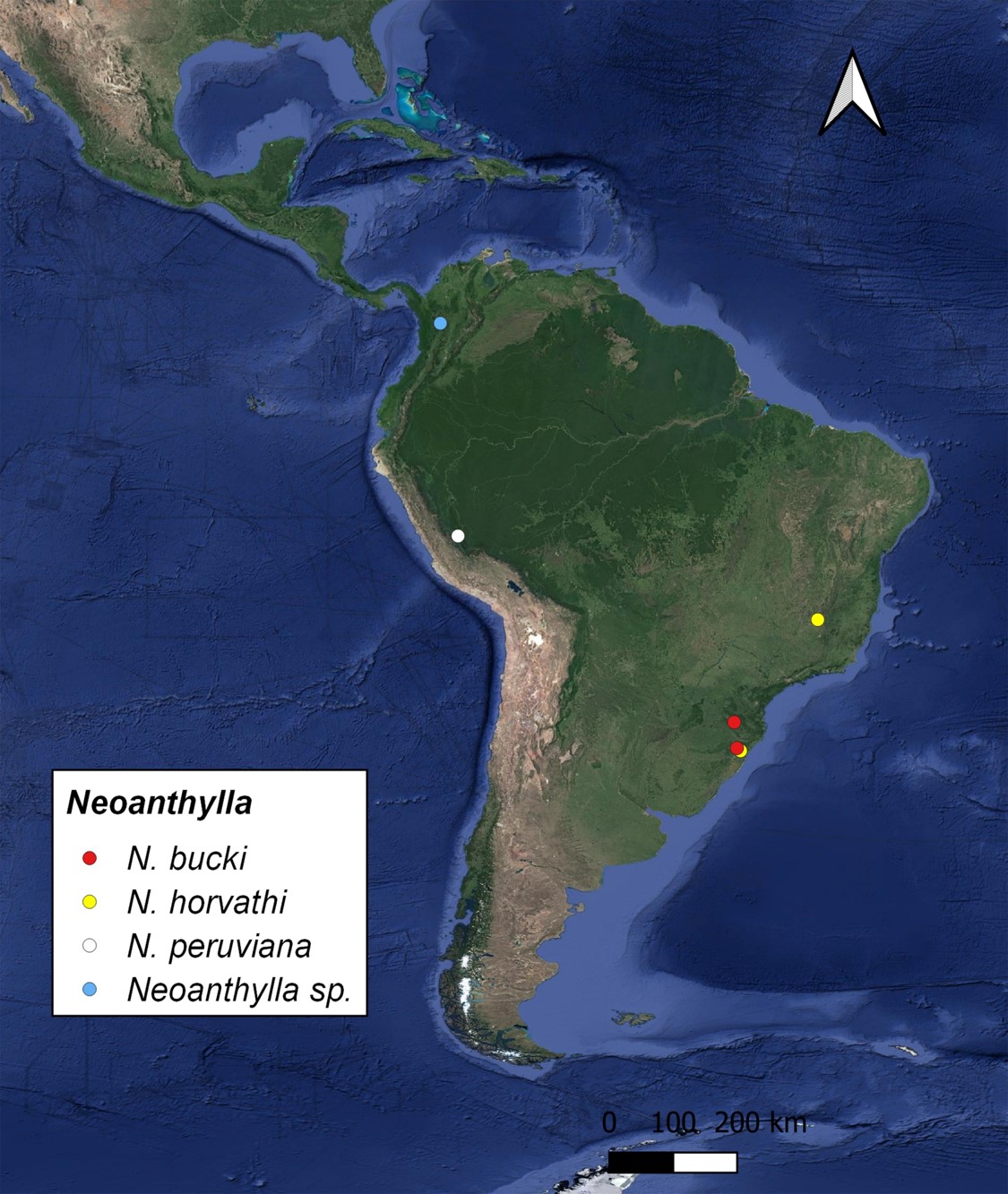
Fig. 5.
Distribution map of the species of Neoanthylla.
Acknowledgments
We are grateful to Ricardo Brugnera (UFRG, Brazil), who donated the Neoanthylla specimen. We also thank the curators of the institutions from which images of type and additional specimens were provided: Anna Somogyi (HNHM), Alana Cioato (MGAP), Thomas Henry and Alyssa Seemann (NMNH), Bärbel Stock-Dietl and Michael Raupach (ZSM). Felipe Toro (www.instagram.com/felipetorocardona) provided images of adult and immature specimens. Three anonymous reviewers helped improve the manuscript with their comments. CONICET postdoctoral and doctoral fellows provided financial support for VCH and PSV, respectively.
REFERENCES
Ang, Y., Wong, L.J. & Meier, R. (2013) Using seemingly unnecessary illustrations to improve the diagnostic usefulness of descriptions in taxonomy–a case study on Perochaeta orientalis (Diptera, Sepsidae). ZooKeys, 355, 9–27. https://doi.org/10.3897/zookeys.355.6013
Davis, N.T. (1957) Contributions to the morphology and phylogeny of the Reduvioidea (Hemiptera: Heteroptera). Part I. The morphology of the abdomen and genitalia of Phymatidae. Annals of the Entomological Society of America, 50, 432–443.
Davis, N.T. (1966) Contributions to the Morphology and Phylogeny of the Reduvioidea (Hemiptera: Heteroptera). Part III. The Male and Female Genitalia. Annals of The Entomological Society of America, 59, 911–924.
Forero, D. & Weirauch, C. (2012) Comparative genitalic morphology in the New World resin bugs Apiomerini (Hemiptera, Heteroptera, Reduviidae, Harpactorinae). Mitteilungen aus dem Museum fur Naturkunde in Berlin - Deutsche Entomologische Zeitschrift, 59, 5–41.
Froeschner, R.C. & Kormilev, N.A. (1989) Phymatidae or ambush bugs of the world: a synonymic list with keys to species, except Lophoscutus and Phymata (Hemiptera). Entomography, 6, 1–76.
Handlirsch, A. (1898) Zwei neue Phymatiden. Verhandlungen der Kaiserlich-koniglichen zoologísch-botanischen Gesellschaft in Wien, 48, 382–383.
Kormilev, N.A. (1951) Phymatidae Argentinas (Hemiptera) con observaciones sobre Phymatidae en general. Revista del Instituto Nacional de Investigación de las Ciencias Naturales, "Bernardino Rivadavia," Ciencias Zoológicas, 2 (2), 45–110.
Kormilev, N.A. (1952) Notas sobre Phymatidae Neotropicales IV (Hemipt. Reduvioidea). Misión de Estudios de Patología Regional Argentina,23, 123–133.
Kormilev, N.A. (1962a) Revision of the Phymatinae (Hemiptera, Phymatidae). The Philippine Journal of Science, 89 (3-4), 287-486, plates 1-19 (imprint date: 1960).
Kormilev, N.A. (1962b) Notes on American Phymatidae II (Hemiptera, Reduviodea). Journal of the New York Entomological Society, 70, 47–58.
Kormilev, N.A. (1963) Notes on American Phymatidae III (Hemiptera- Heteroptera). Proceedings of the Entomological Society of Washington, 65, 215–226.
Kormilev, N.A. (1964) A third species of Neoanthylla Kormilev, 1951, from Peru (Hemiptera: Phymatidae). Proceedings of the Entomological Society of Washington, 66, 137–140.
Kormilev, N.A. & van Doesburg, P.H. (1992) Notes on Macrocephalinae (Heteroptera: Phymatidae), Zoologische Mededelingen, 66, 417–421.
Masonick, P., Michael, A., Frankenberg, S., Rabitsch, W. & Weirauch, C. (2017) Molecular phylogenetics and biogeography of the ambush bugs (Hemiptera: Reduviidae: Phymatinae). Molecular Phylogenetics and Evolution, 114, 225–233. https://doi.org/10.1016/j.ympev.2017.06.010
Masonick, P. & Weirauch, C. (2020) Taxonomic revision of the Nearctic erosa species group of Phymata Latreille, 1802 (Heteroptera: Reduviidae: Phymatinae). Canadian Journal of Arthropod Identification, 41: 1–90.
QGIS (2022). QGIS Geographic Information System. QGIS Association. http://www.qgis.org
Rabitsch, W., Cui, J. & Cai, W. (2006) Two new ambush bugs from Yunnan, southern China (Insecta, Heteroptera, Reduviidae, Phymatinae), Denisia, 19, 581–588.
Schuh R.T. & Weirauch C. (2020) True bugs of the world (Hemiptera: Heteroptera). Classification and natural history. 2nd ed. Siri Scientific Press, Manchester, UK. 768 pp.
Stål, C. (1876) Enumeratio Phymatidarum. In: Enumeratio Hemipterorum vol. 5. Konglinga Svenska Akademiens- Handlingar, 14, 131–136.
Swanson, D.R. (2013) A review of the ambush bugs (Heteroptera: Reduviidae: Phymatinae) of Michigan: identification and additional considerations for two common eastern species. The Great Lakes Entomologist, 46: 154–164.
Swanson, D.R. (2019) A new species of ambush bug (Heteroptera: Reduviidae: Phymatinae) from Ecuador and Peru. Proceedings of the Entomological Society of Washington, 121 (2), 279–289. https://doi.org/10.4289/0013-8797.121.2.279
Weirauch, C., Bérenger, J.M., Berniker, L., Forero, D., Forthman, M., Frankenberg, S., Freedman, A., Gordon, E., Hoey-Chamberlain, R., Hwang, W. S., Marshall, S. A., Michael, A., Paiero, S. M., Udah, O., Watson, C., Yeo, M., Zhang, G. & Zhang, J. (2014) An Illustrated Identification Key to Assassin Bug Subfamilies and Tribes (Hemiptera: Reduviidae). Canadian Journal of Arthropod Identification, 26.
Zhang, J., Gordon, E.R.L., Forthman, M., Hwang, W.S., Walden, K., Swanson, D.R., Johnson, K.P., Meier, R. & Weirauch, C. (2016) Evolution of the assassin’s arms: insights from a phylogeny of combined transcriptomic and ribosomal DNA data (Heteroptera: Reduvioidea). Scientific Reports, 6, 1–8. https://doi.org/10.1038/srep22177
Notas de autor
valeoptera@gmail.com