Articles
Drosophila suzukii Matsumura (Diptera: Drosophilidae): análisis de condiciones experimentales para su cría y estudio en el laboratorio
Drosophila suzuki Matsumura (Diptera: Drosophilidae): analysis of experimental conditions for its rearing and study in the laboratory.
Drosophila suzukii Matsumura (Diptera: Drosophilidae): análisis de condiciones experimentales para su cría y estudio en el laboratorio
Revista de la Sociedad Entomológica Argentina, vol. 82, núm. 1, 2023
Sociedad Entomológica Argentina
Recepción: 01 Diciembre 2022
Aprobación: 22 Marzo 2023
Resumen: Drosophila suzukii (Matsumura) (Diptera: Drosophilidae) es una especie de plaga invasora que utiliza una amplia variedad de frutas comerciales como sitio de cría y alimentación. Los daños al fruto generado por la oviposición de D. suzukii provoca pérdidas económicas considerables. Por lo tanto, el mantenimiento de D. suzukii en condiciones de laboratorio constituye una herramienta fundamental para la propuesta de investigación. Dos de los factores determinantes que afectan las condiciones de crianza en laboratorio son el medio de cultivo y la densidad. En este estudio, evaluamos el impacto de estos factores en tres componentes asociados con la adecuación darwiniana (fitness): tiempo de desarrollo, viabilidad larval-adulta y tamaño corporal, comparando los efectos de tres medios de cultivo: harina de maíz (polenta), puré de papa y ciruela cuando las moscas se criaron a tres densidades: 10, 30 y 50 larvas por 5ml de medio de cultivo. Nuestros resultados revelaron que las moscas criadas en el medio de cultivo de harina de maíz y a bajas densidades presentaron las mejores condiciones para el mantenimiento de D. suzukii en el laboratorio.
Palabras clave: Caracteres adaptativos, Efecto denso-dependiente, Medio de cultivo, Mosca de alas manchadas.
Abstract: Drosophila suzukii (Matsumura) (Diptera: Drosophilidae) is an invasive pest species that uses a wide variety of commercial fruit as a breeding and feeding resource. The damage to the fruit generated by D. suzukii´ oviposition causes considerable economic losses. Therefore, the maintenance of D. suzukii in the lab conditions constitutes a fundamental tool for research proposal. Two of the determining factors affecting the lab rearing conditions are the culture medium and density. In this study, we evaluated the impact of these factors on three components associated with fitness: development time, larval-adult viability and body size, comparing the effects of three culture media: cornmeal (polenta), mashed potato and plum when the flies were reared at three densities: 10, 30 and 50 larvae per 5ml of culture medium. Our results revealed that the flies reared the cornmeal culture medium and at lower densities provide the better conditions for the maintenance of D. suzukii under lab conditions.
Keywords: Adaptive traits, Density-dependent effect, Rearing medium, Spotted-wing flies.
Introducción
Drosophila suzukii Matsumura (Diptera: Drosophilidae), también conocida como la mosca de las alas manchadas, constituye una de las plagas emergentes más importantes a nivel mundial para la producción de fruta fina y de carozo (Cini et al., 2012). Las hembras presentan un ovipositor aserrado que les permite lacerar el epicarpio de las frutas sanas y maduras para oviponer, sin requerir que la fruta esté previamente dañada o sobremadura como ocurre con otras especies de Drosophila (Markow & O'grady, 2008). El daño ocasionado por la oviposición, la posterior alimentación de las larvas, y el ingreso de patógenos causa un deterioro prematuro del fruto (Walsh et al., 2011; Cini et al., 2012), provocando pérdidas económicas millonarias (Lee et al., 2011; Cini et al., 2012).
Ésta especie altamente polífaga presenta una gran capacidad plástica evidenciada por la diversidad de ambientes y de hospederos en los que se la ha encontrado (Kenis et al., 2016; Andreazza et al., 2017; Little et al., 2020). Se han descrito una gran variedad de hospederos de importancia comercial (arándanos, frutillas, cerezas, moras, duraznos, entre otros), así como frutos silvestres (una gran variedad plantas de los géneros Prunus, Lonicera y Vaccinium, entre otros). Indudablemente, durante su proceso de invasión, D. suzukii se expone a ambientes que presentan una amplia heterogeneidad, producto de las variaciones climáticas así como el hecho de encontrar recursos nutricionales nuevos. En estos casos, la plasticidad fenotípica podría ser un mecanismo beneficioso para lidiar con el desafío que representa la heterogeneidad ambiental (Shearer et al., 2016; Fanara & Werenkraut, 2017; Schlichting & Pigliucci, 1998; Lavagnino et al., 2020).
Entre los efectos plásticos descritos que presenta D. suzukii pueden mencionarse modificaciones tanto en su tamaño corporal como en la melanización de su cutícula debido a cambios asociados a la temperatura de desarrollo (Shearer et al., 2016). Por un lado, se ha detectado un morfotipo “estival” de color amarillento y de menor tamaño y, por otro lado, un morfotipo "invernal" de mayor tamaño y de color más oscuro. Éste último, presentaría adaptaciones al clima frío producto de los cambios plásticos asociados a la temperatura durante el desarrollo (Shearer et al., 2016). Asimismo, se han descrito trabajos en los cuales se ha evaluado el efecto nutricional del medio durante el desarrollo larval. Diversos trabajos analizaron el efecto de la proporción Proteínas/Carbohidratos (P/C) tanto en D. suzukii (Hardin et al., 2015; Rendon et al., 2018; Young et al., 2018; Shu et al., 2022) como en otras especies de Drosophila (Grandison et al., 2009; Fanson et al., 2012; Tatar et al., 2014; Rodrigues et al., 2015), demostrando que un alto P/C disminuye el tiempo de desarrollo y aumenta la capacidad reproductiva en las hembras. Por otro lado, se reportaron también desventajas asociadas a las dietas ricas en proteínas, que están asociadas a una menor longevidad y una mayor susceptibilidad a la inanición en los adultos. En el caso del trabajo de Hardin et al. (2015) se demuestra que la densidad de larvas en el medio de cría también afecta la supervivencia y tiempo de desarrollo de las larvas, así como el tamaño de los adultos, pero este efecto se ve disminuido cuando el P/C es alto. Cabe también mencionar que la inoculación con levaduras vivas puede suplementar una dieta de bajo P/C, mejorando la supervivencia de las larvas y su tiempo de desarrollo (Bellutti et al., 2018).
La mayoría de los trabajos que analizan la nutrición y competencia intraespecífica de D. suzukii utilizan distintas frutas naturales o en puré para reflejar condiciones naturales (Shu et al., 2022; Silva-Soares et al., 2017) o, alternativamente, manipulan las concentraciones de nutrientes en medios artificiales con el objetivo de que estos sean comparables entre sí. Esto deja un vacío de conocimiento en cuanto a la comparación entre distintos medios de cría frecuentemente utilizados en el laboratorio y las densidades larvales óptimas en cada uno de ellos. Dada la importancia económica que subyace a la invasión de D. suzukii, estudiar las condiciones óptimas para su cría constituye información relevante para establecer protocolos de investigación que permitan analizar las causas, consecuencias y mitigación de esta invasión. El presente trabajo analiza el efecto que provocan en distintos caracteres adaptativos diferentes densidades larvarias de cría y dietas utilizadas regularmente en laboratorios tanto para esta especie como para otras especies de Drosophila. Ésta información permitirá conocer las condiciones óptimas de cría de D. suzukii en laboratorio, dando factibilidad al desarrollo de futuros proyectos de investigación teniendo en consideración el interés biológico y económico que genera la invasión de esta especie plaga.
Materiales y métodos
Generación del stock poblacional
Los individuos de D. suzukii utilizados en este análisis provinieron de una captura realizada en Luján (Universidad de Luján, Buenos Aires) en cultivos experimentales de ciruelas, duraznos y cerezas. A partir de 30 hembras colectadas en el campo, se generó un stock que se mantuvo en medio de laboratorio estándar a base de harina de maíz (Tabla I) en condiciones controladas: fotoperiodo (12:12 horas de luz:oscuridad) y temperatura (25°C). Los ensayos fueron realizados luego de tres generaciones en estas condiciones, con el objetivo de minimizar el posible efecto de adaptación al medio de cría.

Tabla I.
Composición de los medios de cría utilizados. Las cantidades por réplica fueron de 5 ml para la ciruela y harina de maíz.
Experimento de medio de cría
Se evaluó el efecto de la dieta, la densidad larvaria y la interacción entre estos factores sobre distintos caracteres asociados al fitness: tiempo de desarrollo (TD), viabilidad (V) y tamaño corporal en los adultos (TC). Para ello se realizó un experimento factorial completamente aleatorizado con 3 dietas y 3 densidades larvales. Las dietas evaluadas fueron: ciruela (homogenato de ciruelas, agar), papa (puré de papa instantáneo que consiste en almidón deshidratado) y harina de maíz (medio estándar para cultivo de D. melanogaster, cuyas fuentes de nutrientes son levadura (Saccharomyces cerevisiae Meyen), dextrosa y sacarosa) (Tabla I). Las densidades larvales analizadas fueron 10, 30 y 50 larvas por 5 ml de medio evaluado.
Para la obtención de larvas se tomaron del stock 80 parejas de adultos sexualmente maduros y se colocaron en un recipiente cilíndrico de acrílico (10 cm de alto por 5 cm de diámetro) cuyo fondo consiste en una placa de Petri con agar 3% (3 g agar sólido: 100 ml de agua destilada), unos granos de levadura comercial como fuente de alimento y homogenato de ciruela (Prunus domestica Linneo) para estimular la ovipuesta. En total se hicieron 10 réplicas de este dispositivo. La placa de Petri de cada réplica se renovó cada 24 horas y se incubó a 25°C hasta el momento de emergencia de las larvas del primer estadio. En todas las réplicas, se registró el momento en el que se colocó (Ti) y se retiró (Tf) la placa de Petri. Para cada combinación de dieta y densidad se sembraron larvas de primer estadio en viales de vidrio (10 cm de alto por 2.5 cm de diámetro) que contenían 5 ml del medio de cría correspondiente. Cada 12 horas se registró la emergencia de los adultos, los cuales se separaron por sexo y se congelaron para posteriores mediciones corporales. Por cada combinación de dieta y densidad se hicieron 6 réplicas que se mantuvieron en condiciones controladas de fotoperiodo (12:12 horas de luz:oscuridad) y temperatura (25 °C).
Caracteres adaptativos analizados
El TD se definió como el tiempo transcurrido (en horas) desde que los huevos eclosionaron hasta la emergencia del adulto. La V se definió como la proporción de individuos que logró alcanzar la etapa adulta y se cuantificó a partir de la cantidad de moscas adultas sobre el total de larvas sembradas en cada tratamiento de densidad evaluado. Finalmente, el TC se estimó a partir de la medición del largo del ala izquierda. Para ello, el ala de cada individuo se removió y fotografió empleando una lupa Leica MZ6 que dispone de una cámara digital Canon Powershot S50 conectada a una computadora con el software Canon Utilities Remote Capture (Canon Inc., ©2000-2003). A partir de las fotografías obtenidas se realizaron mediciones lineales de la distancia entre la intersección de la vena longitudinal anterior con la segunda vena transversal hasta el extremo distal del ala utilizando el programa tpsDig (Rohlf, 2001). Las mediciones del TD y TC se hicieron para cada sexo por separado.
Análisis estadístico
Las variables asociadas al fitness fueron analizadas mediante modelos lineales generalizados mixtos (GLMMs), empleando como variables de efectos fijos la dieta, densidad y la interacción entre ambas. Adicionalmente para el caso de las variables TC y TD se incluyó el sexo como covariable, contemplando adicionalmente su interacción con las restantes variables y la variable aleatoria Réplica. Para el TD se empleó una distribución normal para los errores empleando una función de enlace logarítmica. Dado que se detectó el incumplimiento de homocedasticidad se emplearon diferentes estructuras de varianzas estimadas para todas las combinaciones de las variables de efectos fijos. La elección entre dichos modelos se realizó por el criterio de Akaike (AIC), siendo el modelo elegido aquel que presentaba la interacción dieta x densidad. Este modelo estimó una varianza para cada combinación de dieta y densidad. Para el TC se empleó una distribución normal para los errores utilizando una función de enlace identidad. Finalmente, para la viabilidad se utilizó una distribución binomial para los errores usando una función de enlace logit. Debido a que se detectó sobredispersión en el modelo implementado, se optó por adicionar una variable aleatoria al nivel de la observación (Harrison, 2014). Todos los GLMMs implementados fueron realizados utilizando la librería “glmmTMB” (Brooks et al., 2017) y el cumplimiento de los supuestos fue corroborado utilizando la librería “DHARMa” (Hartig & Hartig 2017). Posteriormente se aplicó la corrección por pruebas múltiples de Bonferroni.
Resultados
Se observó que todos los caracteres estudiados exhibieron diferencias significativas en función de la densidad larvaria y de la dieta suplementada (Tabla II yIII). En términos generales, el mejor desempeño para todos los caracteres estudiados se detectó en el medio estándar de harina de maíz y a densidades bajas de individuos. El efecto de la dieta reveló que el medio de harina de maíz presenta la mayor V respecto a los medios preparados con papa y con ciruela (odds 7,37 y 5,09 respectivamente) (Fig. 1A). Por su parte, también se detectó un efecto denso-dependiente detrimental para la V. En efecto, la V se vio incrementada significativamente en la densidad de 10 individuos respecto de las densidades de 30 y 50, con un odds de 4,17 y 8,13 respectivamente (Fig. 1A). A su vez, la densidad de 30 mostró un odds de 1,95 respecto a la densidad de 50 (Fig. 1A).
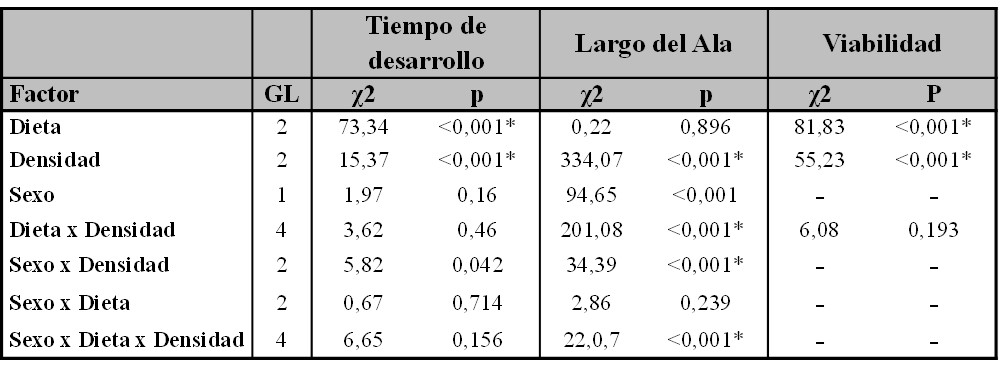
Tabla II.
Resultados de los GLMMs realizados para los caracteres de vida en Drosophila suzukii.
La significancia de las variables de efectos fijos fue testeada usando la prueba de Chi-cuadrado de Wald. * Representa significancia estadística después de la corrección por el método de Bonferroni. GL: grados de libertad.
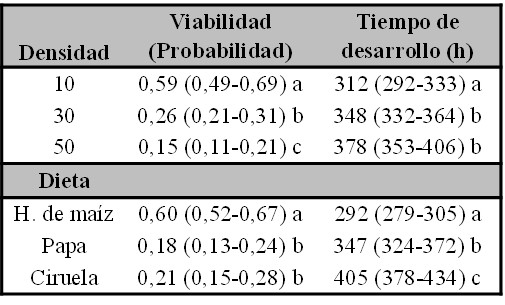
Tabla III.
Valores medios predichos por el modelo con su intervalo de confianza del 95% estimados, entre paréntesis, para los factores Dieta y Densidad para la Viabilidad y el Tiempo de desarrollo. Las letras diferentes denotan diferencias significativas entre los valores medios.
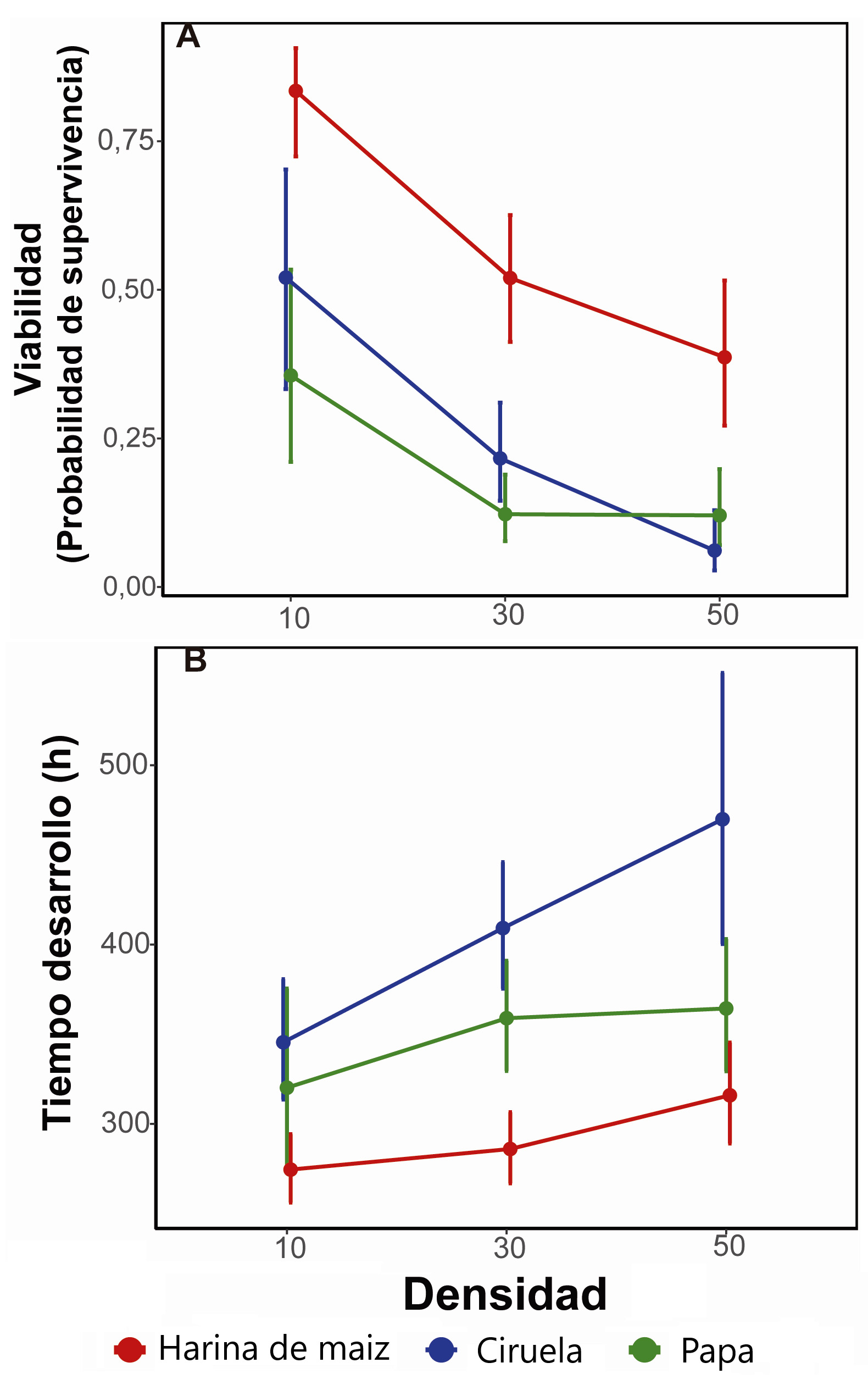
Fig. 1.
Valores medios con su intervalo de confianza del 95% para los valores esperados de los GLMMs para la viabilidad (A) y el tiempo de desarrollo (B) en función de la dieta y la densidad.
El TD de los individuos fue menor en el medio de harina de maíz respecto al medio de papa (16% menos) y el de ciruela (28% menos). Adicionalmente, en el medio con papa se detectó un menor TD que en el medio de ciruela (14,3% menos) (Fig. 1B). En cuanto a la densidad de larvas, también se detectó un efecto denso-dependiente ya que las moscas fueron más lentas en su desarrollo a medida que aumentó la densidad. Efectivamente, detectamos un incremento del 11% y del 22% del TD en las densidades de 30 y 50 respecto de la densidad de 10, respectivamente (Fig. 1B). Por su parte, la densidad de 50 individuos presentó un mayor TD (10% más respecto a la densidad 30). Cabe resaltar que ninguna de las interacciones para los caracteres V y TD fue significativa, sugiriendo que el tipo de alimentación y la densidad afectan estos caracteres de manera independiente.
Finalmente, con respecto al TC, si se detectó un efecto interactivo significativo entre los factores sexo, medio y densidad (Fig. 2). Para facilitar su interpretación se presentan los efectos para cada sexo por separado. Considerando que las hembras exhibieron un mayor TC en cada combinación de condiciones de cría, excepto para la combinación dieta ciruela – densidad 50 larvas, se observó un efecto diferencial por sexo siendo las hembras las más afectadas por las densidades altas y la dieta de Ciruela (Tabla IV). En líneas generales y para ambos sexos, a menores densidades se detectaron los individuos con mayor tamaño. No se detectaron diferencias entre las dietas evaluadas en la densidad de 10 larvas por tubo (Tabla IV). El tipo de dieta afectó diferencialmente al TC en ambos sexos a medida que se incrementó la densidad, de modo tal que a densidad 50 los mayores TC corresponden a los individuos criados en harina de maíz (Tabla IV). En este sentido, la reducción del TC como consecuencia del aumento de la densidad varió según el tipo de alimento siendo los individuos con menor variación de tamaño los que se criaron alimentándose en el medio de harina de maíz.
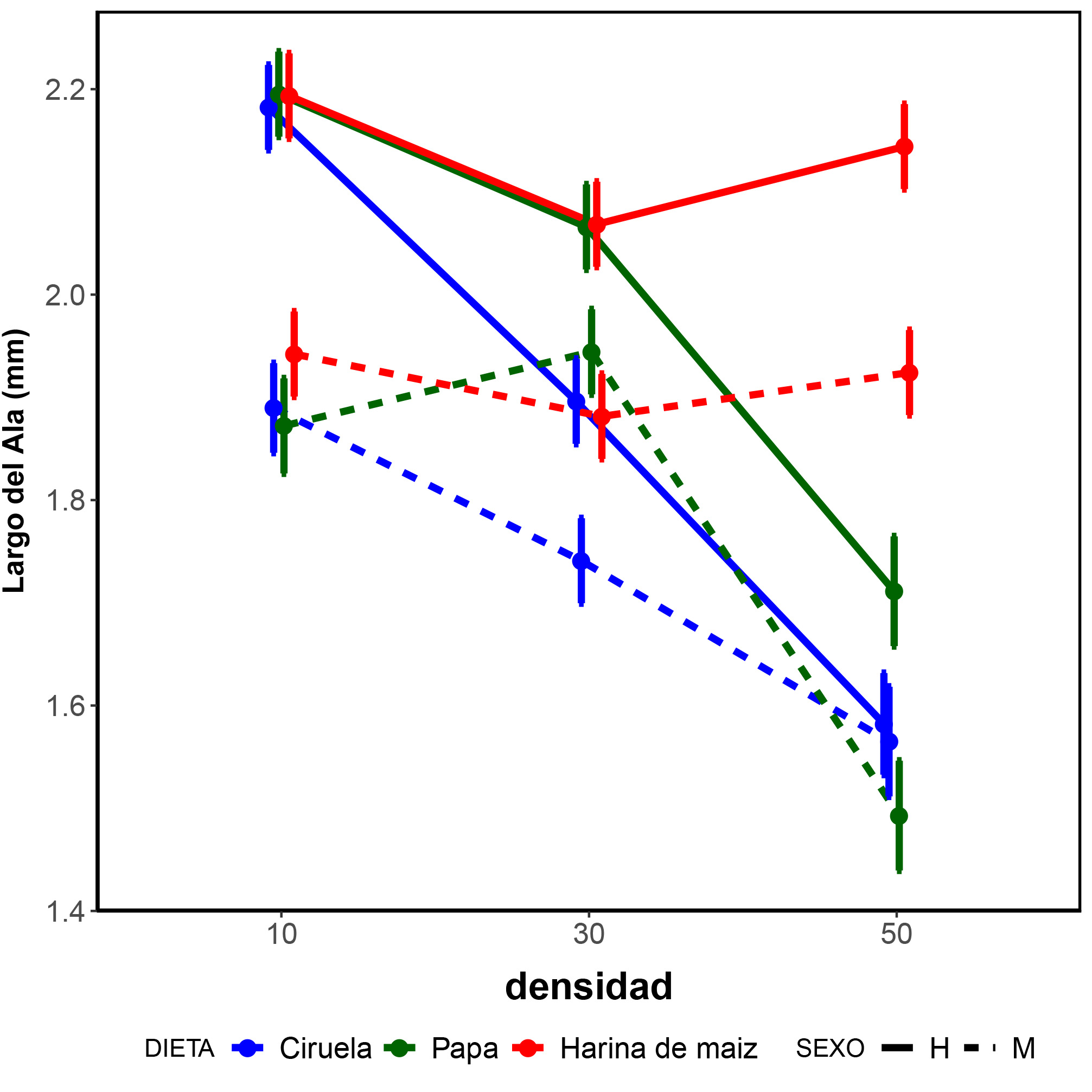
Fig. 2.
Valores medios con su intervalo de confianza del 95% para los valores esperados del largo del ala en función de la dieta, densidad y sexo.
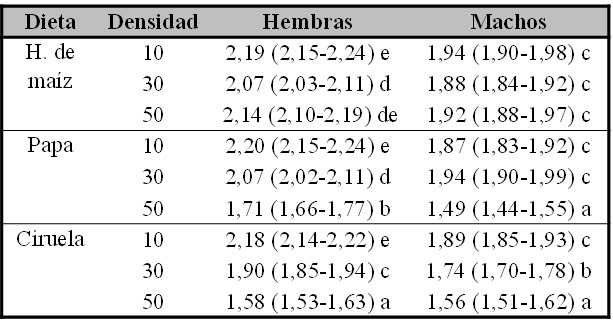
Tabla IV.
Valores medios predichos por el modelo con su intervalo de confianza del 95% estimado, entre paréntesis, para el factor Tamaño corporal, estimado a partir del largo del ala (mm). Las letras diferentes denotan diferencias significativas entre los valores medios.
Discusión
Desde la detección de D. suzukii en el país, se desarrollaron numerosos estudios de campo que reflejan la gran amplitud de recursos que puede utilizar D. suzukii evidenciado en la diversidad de frutales naturales en donde se la ha recolectado (Díaz et al., 2015; Santadino et al., 2015; Lavagnino et al., 2018; De la Vega et al., 2020; Dettler et al., 2021). Este carácter generalista que presenta la especie determina que el estudio de las técnicas de colecta y, principalmente de cultivo en el laboratorio, sea un paso fundamental para su manejo y mantenimiento. Dos de los factores determinantes para el mantenimiento de líneas de moscas en el laboratorio son la dieta y la cantidad de larvas por vial. En este sentido, el presente trabajo evaluó el impacto de estos factores en tres componentes asociados al fitness. En relación al efecto de la dieta, los menores tiempos de desarrollo y mayores niveles de tamaño corporal y viabilidad detectados en el medio de harina de maíz indican que el medio convencional de mantenimiento empleado para D. melanogaster Meigen, constituye un medio idóneo para la cría de D. suzukii.
Con respecto a la densidad de larvas, se detectó un efecto negativo significativo respecto a los componentes de fitness evaluados, ya que el incremento de la misma ocasionó un aumento en los TD y una disminución de la V y del TC. Trabajos previos empleando diferentes medios de cría han detectado un efecto negativo tanto para el TD (Hamby et al., 2016; Plantamp et al., 2017) como para la viabilidad larva-pupa (Silva-Soares et al., 2017), a medida que la densidad es incrementada. No obstante, algunos trabajos detectaron un efecto diferencial de la dieta dependiendo de la densidad de individuos/huevos utilizada por vial (Hardin et al., 2015). Burrack et al. (2013) encontraron 4,2 ± 1,3 larvas por fruto en arándanos, y Yu et al. (2013) un promedio de 2,64 ± 0.84 larvas por mora. Por otro lado, la cantidad de larvas de D. suzukii por fruto registradas en la bibliografía es variable, y depende principalmente del tamaño de los frutos infestados. Considerando el tamaño de los frutos y la baja movilidad de las larvas, es esperable que exista competencia entre éstas, y que se encuentren efectos de la densidad sobre el desempeño de las larvas fuera del ambiente del laboratorio.
En cuanto al efecto de los medios evaluados, los mismos difieren en sus características nutricionales, siendo el medio a base de harina de maíz el que presenta la mayor relación de proteínas a carbohidratos (P/C), seguido por el medio a base de ciruela. Sin embargo, el medio de ciruela presentó la menor concentración de nutrientes totales (P+C) dado su alto contenido de agua. Además, el medio de papa fue suplementado con levaduras vivas, las cuales constituyen una fuente de proteínas, mientras que el homogenato de ciruelas no fue adicionado con levadura. La dieta de harina de maíz contuvo levaduras muertas dado que fue cocida en olla a presión, por lo que presentó una cantidad fija de proteínas, mientras que las levaduras vivas convierten los carbohidratos en proteínas (además de otros compuestos como etanol que pueden ser nocivos).
Los resultados obtenidos son coincidentes con lo observado por estudios previos, en el cual el medio de mayor P/C provee las mejores condiciones para el desarrollo de D. suzukii teniendo en cuenta las tres medidas asociadas al fitnessque se evaluaron (Hardin et al., 2015; Young et al., 2018; Shu et al., 2022). Además, el efecto negativo de la densidad de larvas sobre el tamaño corporal se vio disminuido significativamente en el medio de harina de maíz (el de >P/C) y el de papa (con el agregado de levaduras vivas). Esto coincide también con los trabajos mencionados anteriormente donde se controló tanto la densidad de larvas (Hardin et al., 2015; Shu et al., 2022) como la presencia de levaduras (Young et al., 2018).
En el caso del agregado de levaduras, se hipotetizó que éstas reemplazan la falta de proteínas en los recursos de menor relación P/C, por los cuales D. suzukii presenta preferencia tanto para la oviposición como para la alimentación (Young et al., 2018). En el trabajo realizado por Young et al. (2018) el medio de bajo P/C con presencia de levaduras (comparable al medio de puré) presentó igual viabilidad al de alto P/C y ausencia de levaduras vivas (comparable al medio de harina de maíz). Nuestros resultados difieren de estos probablemente debido a que el medio de papa fue preparado con una solución de metil-parabeno (“nipagin”) la cual limita el desarrollo de las levaduras y posiblemente reduzca su efecto positivo. A pesar de esto no se recomienda el uso del medio a base de papa para la cría de D. suzukii ya que la mortalidad de adultos es muy elevada (Gandini, observación personal). El alto contenido de proteínas de la dieta de harina de maíz podría entonces reemplazar las levaduras halladas normalmente en las frutas de las que se alimenta D. suzukii. En condiciones naturales las levaduras son introducidas por medio de la lesión provocada por el ovipositor de la hembra. Sin embargo se ha comprobado que las lesiones en la fruta provocadas por otros medios en lugar de mejorar las condiciones, aumentan los efectos negativos de la densidad larval sobre el TD y el TC (Plantamp et al., 2017).
La relevancia de estos resultados presentados en este trabajo recae en su utilidad para la cría de esta especie en condiciones de laboratorio. Esto es una condición importante para la generación de conocimiento científico sobre características fisiológicas y ecológicas de esta especie que pueda ser utilizado en el futuro para la planificación de planes de manejo que puedan mitigar los daños producidos sobre cultivos y frutas silvestres. Adicionalmente, el conocimiento acerca de los puntos donde esta especie fue encontrada es relevante dado que evidencia un continuado avance en la distribución de esta especie y zonas con potencial de ser invadidas que aún no han sido colonizadas.
Agradecimientos
Este trabajo fue financiado por el subsidio PICT-2017-2525 de la Agencia Nacional de Promoción de la Investigación, el Desarrollo Tecnológico y la Innovación (ANPCyT). Se agradece la colaboración para la realización de muestreos de personal del Instituto Nacional de Tecnología Agropecuaria (INTA) a la Ing. Silvia Norma Tapia (E.E.A. Yuto, Jujuy), Ing. Daniel Lamberti (Agencia de Extensión Rural Perico, Jujuy), Ing. Claudia F. Funes (E.E.A Famaillá, Tucumán), María Carla Chiandussi, (E.E.A La consulta, Mendoza), Lic. Ines Gonzales de Prada (Centro Universitario Tilcara, FFyL-UBA) y Dra. María Florencia Barbarich (Centro Universitario Tilcara, FFyL-UBA).
Referencias
Andreazza, F., Bernardi, D., Dos Santos, R.S.S., Garcia, F.R. M., Oliveira, E.E., Botton, M., & Nava, D.E. (2017). Drosophila suzukii in southern neotropical region: current status and future perspectives. Neotropical Entomology, 46, 591-605.
Bellutti, N., Gallmetzer, A., Innerebner, G., Schmidt, S., Zelger, R., & Koschier, E.H. (2018). Dietary yeast affects preference and performance in Drosophila suzukii. Journal of pest science, 91, 651-660.
Brooks, M.E., Kristensen, K., Van Benthem, K.J., Magnusson, A., Berg, C.W., Nielsen, A., Skaug, H.J., Machler, M., & Bolker, B.M. (2017). glmmTMB balances speed and flexibility among packages for zero-inflated generalized linear mixed modeling. The R journal, 9, 378-400
Burrack, H.J., Fernandez, G.E., Spivey, T., & Kraus, D.A. (2013). Variation in selection and utilization of host crops in the field and laboratory by Drosophila suzukii Matsumara (Diptera: Drosophilidae), an invasive frugivore. Pest Management Science, 69, 1173-1180.
Cini, A., Ioriatti, C., & Anfora, G. (2012). A review of the invasion of Drosophila suzukii in Europe and a draft research agenda for integrated pest management. Bulletin of Insectology, 65, 149-160.
De la Vega, G.J., Corley, J.C., & Soliani, C. (2020). Genetic assessment of the invasion history of Drosophila suzukii in Argentina. Journal of Pest Science, 93, 63-75.
Dettler, M.A., Barrientos, G.N., Martinez, E., Ansa, M.A., Santadino, M.V., Coviella, C.E., & Virgala, M.B.R. (2021). Nivel de infestación a campo de Zaprionus indianus Gupta y Drosophila suzukii (Matsumura) (Diptera: Drosophilidae) en Ficus carica L. (Rosales: Moraceae) y Rubusidaeus L. (Rosales: Rosaceae) en el noreste de la provincia de Buenos Aires. Revista de la Sociedad Entomológica Argentina, 80, 43-47.
Díaz, B.M., Lavagnino, N., Garrán, S., Hochmaier, V., & Fanara, J.J. (2015). Detección de la mosca plaga Drosophila suzukii (Matsumura) en la región de Concordia (Entre Ríos). Libro de resúmenes de las XV Jornadas Fitosanitarias Argentinas, 287.
Fanara, J.J, & Werenkraut, V. (2017). Phenotypic plasticity in Drosophila cactophilic species: the effect of competition, density, and breeding sites. Insect Science, 24, 675-683.
Fanson, B.G., Fanson, K.V., & Taylor, P.W. (2012). Cost of reproduction in the Queensland fruit fly: Y-model versus lethal protein hypothesis. Proceedings of the Royal Society B: Biological Sciences, 279, 4893–4900.
Grandison, R.C., Piper, M.D.W., & Partridge, L. (2009). Amino-acid imbalance explains extension of lifespan by dietary restriction in Drosophila. Nature,462, 1061–1064.
Hamby, K.A., Bellamy, E., Chiu, D., Lee, J.C., Walton, V.M., Wiman, N.G., & Biondi, A. (2016). Biotic and abiotic factors impacting development, behavior, phenology, and reproductive biology of Drosophila suzukii. Journal of pest science, 89, 605-619.
Hardin, J.A., Kraus, D.A., & Burrack, H.J. (2015). Diet quality mitigates intraspecific larval competition in Drosophila suzukii. Entomologia Experimentalis et Applicata, 156, 59-65.
Harrison, X.A. (2014). Using observation-level random effects to model overdispersion in count data in ecology and evolution. PeerJ, ., e616
Hartig, F., & Hartig, M.F. (2017). Package ‘DHARMa’. R Development Core Team.Vienna, Austria.
Kenis, M., Tonina, L., Eschen, R., van der Sluis, B., Sancassani, M., Mori, N., Haye, T., & Helsen, H. (2016). Non-crop plants used as hosts by Drosophila suzukii in Europe. Journal of Pest Science, 89, 735-748.
Lavagnino, N.J., Diaz, B.M., Cichón, L.I., de la Vega, G.J., Garrido, S.A., Lago, J.D., & Fanara, J.J. (2018) More records of the invasive pest Drosophila suzukii in the South American continent. Revista de la Sociedad Entomológica Argentina, 77, 27-31.
Lavagnino, N.J., Imberti, M., Flaibani, N., Ortiz, V., & Fanara, J.J. (2020). Contribution of population-level phenotypic plasticity to the invasiveness of Zaprionus indianus (Diptera: Drosophilidae). European Journal of Entomology, 117, 118-128.
Lee, J.C., Bruck, D.J., Dreves, A.J., Ioriatti, C., Vogt, H., & Baufeld, P. (2011). In Focus: Spotted wing Drosophila, Drosophila suzukii, across perspectives. Pest management science; 67, 1349-1351.
Little, C.M., Chapman, T.W., & Hillier, N.K. (2020). Plasticity is key to success of Drosophila suzukii (Diptera: Drosophilidae) invasion. Journal of Insect Science, 20, 5.
Markow, T. A., & O'grady, P. (2008). Reproductive ecology of Drosophila. Functional Ecology, 22, 747-759.
Plantamp, C., Estragnat, V., Fellous, S., Desouhant, E., & Gibert, P. (2017). Where and what to feed? Differential effects on fecundity and longevity in the invasive Drosophila suzukii. Basic and Applied Ecology, 19, 56-66.
Rendon, D., Buser, J., Tait, G., Lee, J.C., & Walton, V.M. (2018). Survival and fecundity parameters of two Drosophila suzukii (Diptera: Drosophilidae) morphs on variable diet under suboptimal temperatures. Journal of Insect Science, 18, 8.
Rodrigues, M.A., Martins, N.E., Balancé, L.F., Broom, L.N., Dias, A.J., Fernandes, A.S.D., Rodrigues, F., Sucena, E., & Mirth, C.K. (2015). Drosophila melanogaster larvae make nutritional choices that minimize developmental time. Journal of insect physiology, 81, 69-80.
Rohlf, F.J. (2001). TPSDigª. Department of Ecology & Evolution. New York Univ, Stony Brook, NY.
Santadino, M.V., Riquelme Virgala, M.B., Ansa, M.A., Bruno, M., Di Silvestro, G., & Lunazzi, E.G. (2015) Primer registro de Drosophila suzukii(Diptera: Drosophilidae) asociado al cultivo de arándanos (Vaccinium spp.) de Argentina. Revista de la Sociedad Entomologica Argentina, 74, 183-185
Schlichting, C.D., & Pigliucci, M. (1998). Phenotypic evolution: a reaction norm perspective. Sinauer associates incorporated, MA (USA).
Shearer, P.W., West, J.D., Walton, V.M., Brown, P.H., Svetec, N., & Chiu, J.C. (2016). Seasonal cues induce phenotypic plasticity of Drosophila suzukii to enhance winter survival. BMC ecology, 16, 1-18.
Shu, R., Uy, L., & Wong, A.C.N. (2022). Nutritional phenotype underlines the performance trade-offs of Drosophila suzukii on different fruit diets. Current Research in Insect Science, ., 100026.
Silva-Soares, N.F., Nogueira-Alves, A., Beldade, P., & Mirth, C.K. (2017). Adaptation to new nutritional environments: larval performance, foraging decisions, and adult oviposition choices in Drosophila suzukii. BMC ecology, 17, 1-13.
Tatar, M., Post, S., & Yu, K. (2014). Nutrient control of Drosophila longevity. Trends in Endocrinology & Metabolism, 25, 509-517.
Walsh, D.B., Bolda, M.P., Goodhue, R.E., Dreves, A.J., Lee, J., Bruck, D.J., Walton, V.M., O’Neal, S.D., & Zalom, F.G. (2011). Drosophila suzukii (Diptera: Drosophilidae): invasive pest of ripening soft fruit expanding its geographic range and damage potential. Journal of Integrated Pest Management, ., G1-G7.
Young, Y., Buckiewicz, N., & Long, T. A. (2018). Nutritional geometry and fitness consequences in Drosophila suzukii, the Spotted Wing Drosophila. Ecology and evolution, ., 2842-2851.
Yu, D., Zalom, F.G., & Hamby, K.A. (2013). Host status and fruit odor response of Drosophila suzukii (Diptera: Drosophilidae) to figs and mulberries. Journal of Economic Entomology, 106, 1932-1937.
Notas de autor
jjfanara@ege.fcen.uba.ar