Artículos
The genus Epanycles (Lepidoptera: Erebidae: Arctiinae: Ctenuchina): 145 years of monotypy
El género Epanycles (Lepidoptera: Erebidae: Arctiinae: Ctenuchina): 145 años monotípico
The genus Epanycles (Lepidoptera: Erebidae: Arctiinae: Ctenuchina): 145 years of monotypy
Revista de la Sociedad Entomológica Argentina, vol. 82, núm. 2, 2023
Sociedad Entomológica Argentina
Recepción: 06 Agosto 2022
Aprobación: 03 Marzo 2023
Abstract: The genus Epanycles Butler (Lepidoptera: Erebidae: Arctiinae: Ctenuchina) has been considered as monotypical for 145 years, including only Epanycles imperialis (Walker). Herein, we propose a new combination: E. satania new. comb. based on external morphology, genital morphology of males and females, morphological phylogeny, DNA barcoding, and geographical distribution. Epanycles satania was originally described for the genus Episcepsis Butler.
Keywords: Episcepsis, Neotropic, New combination, Taxonomy.
Resumen: El género Epanycles Butler (Lepidoptera: Erebidae: Arctiinae: Ctenuchina) ha sido considerado como monotípico desde hace 145 años, incluyendo solo a Epanycles imperialis (Walker). En base a datos de morfología externa, morfología de los genitales de machos y hembras, estudios de filogenia morfológica, información molecular de DNA barcoding y distribución geográfica, se propone una nueva combinación: Epanycles satania new comb. Esta última descrita y considerada dentro del género Episcepsis Butler.
Palabras clave: Episcepsis, Neotrópico, Nueva combinación, Taxonomía.
INTRODUCTION
The subtribe Ctenuchina (Lepidoptera: Erebidae: Arctiinae) includes more than 90 genera (Zerny, 1912) with a wide variety of wing coloration patterns, presence of different androconial organs, and diurnal habits. Within this subtribe, at least ten genera could be engaged in mimetic complexes with wasp species of the Suborder Apocrita (Hymenoptera). Several genera have been proposed and yet considered as monotypical (Hampson, 1898; Draudt, 1916-1919) and this would be related to the limited knowledge when descriptions were published. Moreover, data survey on species occurrence in many Neotropical countries is scarce. We have a fair knowledge of Mexico, Costa Rica, Ecuador, Peru, Brazil and French Guiana.
Recently, Grados et al. (2020) suggested that the genus Parascepsis Dognin, originally considered as monotypic, would actually include at least five species, based on material from Peru. Something similar occurs with Epanycles Butler, currently considered as monotypic (Hampson, 1898; Draudt, 1916-1919; Araujo et al., 2019). This genus was proposed by Butler (1876) who designated Euchromia imperialis Walker, 1854 as typespecies by monotypy. This species is easily recognized by its coloration pattern. A detailed study of Epanycles with the three species historically included in the genus can be found in Araujo et al. (2019).
This study proposes the non-monotypy of Epanycles based on external morphology, genital morphology of male and female, morphological phylogeny (Grados & Mantilla, unpublished data), DNA barcoding, and geographical distribution. According to our observations, this genus included two species: Epanycles imperialis (Walker) and Epanycles satania (Schaus) new. comb; this latter has been included historically in the genus Episcepsis Butler. The geographical distribution of E. imperialis in Peru is also presented and discussed, which will help future biogeographical studies on this species along the Neotropical region.
MATERIAL AND METHODS
For genital dissection, this study followed Robinson (1976) with modifications. The abdomen was immersed in a test tube filled with KOH (10%) in a water bath for 15 to 20 min. Scales were removed with fine brushes, chlorazol black was used for staining purposes (Cannon, 1937, 1941; Carayon, 1969) and then, stored in a microvial with glycerin. Adult specimens were photographed with a digital camera Nikon D80 and genital organs with a camera Canon EOS Rebel T6 and macro lens Canon MPE 65 mm. Terminology for wing venation follows Comstock & Needman (1898; 1899), Miller (1970), Wootton (1979) and Common (1990); for genital structures Klots (1970), and for coremata Kreusel (2000).
Sample sequencing of Epanycles imperialis was obtained from the Peruvian Amazon (Tambopata River, Madre de Dios) which is part of the Wired Amazon project, Discovery new species, that took place at Guelph University (Canada), corresponding code: RFEWA744-19 (GenkBank access number: BankIt2604995 gnl|uoguelph|RFEWA744-19.COI-5P OP028699). DNA sequences of Epanycles imperialis from French Guiana is: LNOUG055-18 (GenkBank access number: BankIt2604995 gnl|uoguelph|LNOUG055-18.COI-5P OP028700). DNA sequences identified as “Epanycles imperialis” from Central America were obtained from public data available in BOLD SYSTEM. Codes for each specimen are presented as follows: Costa Rica (Guanacaste Conservation Area): BLPCE695_08, BLPCN301_08 y BLPCM478_08; Panama: BCIPY865-21. Genetic distances were calculated with the aid of the software MEGA 11(Tamura et al., 2021).
The following collections were examined to the elaboration of the present work: Private Research Collection A. y R. Turrent, Mexico City, Mexico (CART); Museo de Historia Natural de la Ciudad de México, Mexico (MCM); Museo de Historia Natural, Universidad Nacional Mayor de San Marcos, Lima, Peru (MUSM); Natural History Museum, London, United Kingdom (NHMUK), and National Museum of Natural History, Smithsonian, Institution, Washington DC, USA (USNM).
RESULTS AND DISCUSSION
Taxonomy
EpanyclesButler, 1876.
EpanyclesButler, 1876: 425-426.
Type species: Euchromia imperialisWalker, 1854, by original designation.
Diagnosis: Head, thorax and abdomen brown with iridescent green scales. Antenna brown slightly iridescent green. A fold in the anal area on the ventral side of the hind wings, presenting androconial organs: (1A+2A) - 3A. Tegumen with two spiniform processes. Uncus lobe subrectangular. Presence of subscaphium. Ductus bursae sclerotized in part and bursa copulatrix rounded.
Epanycles imperialis (Walker, 1854)
Euchromia imperialisWalker, 1854: 241.
Diagnosis: Brown head, thorax and abdomen with iridescent green scales; tornus margin of hind wings oval; presence of androconial organs in the form of long yellowish hairs tufts in a fold in the anal area on the ventral side of the hind wings; uncus longer than wide; coremata present (tube-shape, on the ventral side of the last abdominal segments); anal papillae higher than long; sclerotized part of antrum convex; sclerotized part of ductus bursae more than half of total length; bursa copulatrix about twice the width of anal papillae.
Description: A detailed redescription and photographs of male and female adults and genital character of both sexes can be found in Cerda (2017) and Araujo et al. (2019).
Examined material. AMAZONAS: 2 males, Cordillera del Condor, ca. Qda. Chinganasa (Qda. Ponce), 03°47S, 78°20’W, 14.xi.2003, 800 m, J. Grados & A. Asenjo; 3 males, idem except, 15.xi.2003, 850-1160 m. LORETO. 1 male, Soplín, Vargas, 00°22’30’’S, 74°40’30’’W,14-18.v.2010, 210 m, J.J. Ramírez leg.; 1 male, Río Curaray, Campamento Paiche, 01°29’25’’S, 75°23’53’’W, 02.iii.2008, 200 m, Williams Yawarcani; 1 male, Río Curaray, Campamento Paiche, 01°32’00’’S, 75°25’16’’W, 10.ii.2008, 215 m, Williams Paredes; 1 male, Río Arabela, Campamento 7B, 01°54’06’’S, 75°22’56’’W,18.xi.2007, 190 m, J. J. Ramírez leg.; 1 male, El Milagro, 03°57’S, 73°22’’W, 24.ix.2004, 120 m, J. J. Ramírez. UCAYALI. 1 male, 12 km SSE Lontananza, Río Yaquirana, 06°14’50’’S, 73°17’32’’W, 04-06.x.2008, 165 m, A. García; 2 males, 3.5 km SSO de Nevo Galicia (Río Abujao), 08°22’25’’S, 73°48’50’’W,22-25.ix.2011, 213 m, C. Espinoza; 1 male, 1 female, 29.5 km NO de Nuevo Mundo, Campamneto Kinteroni CX, río Sepa, 11°22’38’’S, 73°24’47’’W, 20-24.i.2010, 643 m, J. Grados. PASCO. 1 male, Comunidad 24 de junio, 10 km NO de Cajonari, 10°03’57’’S, 75°07’02’’W, 03-06.xi.2015, 359 m, L. Figueroa, GENITALIA # JGA - 1223, MUSM. JUNÍN. 1 male, 9.5 km NE de Soroja, Campamento Mashira JX, río Mayapo, 11°25’21’’S, 73°27’56’’W, 28.i.-02.ii.2010, 545 m, J. Grados, GENITALIA # JGA - 967, MUSM. CUSCO. 1 male, Río Urubamba, Conkariari, 11°40’S,72°59’W, 13.x.97, 350 m, J. Grados, LIGHT TRAP MV; 2 males, San Martín C, 11°47’S,72°41’W, 24.iii.97, 480 m, J. Grados; 3 males, idem except, 25.iii.97; 2 males, ídem except, 26.iii.97; 1 male, idem except, 28.iii.97; 1 male, idem except, 30.iii.97; 2 male, idem except, 31.iii.97; LIGHT TRAP MV; 1 male, Segakiato, río Camisea, 11°48’S, 72°52’W, 02.x.97, 330 m, J. Grados, LIGHT TRAP MV/UV; 1 male, idem except, 03.x.97; 1 male, Armihuari, 11°51’S, 72°46’W, 11.vi.97, 560 m, G. Valencia, LIGHT TRAP; 2 males, Las Malvinas, río Urubamba, 11°52’S, 72°56’W, 18.ix.97, 360 m, J. Grados, LIGHT TRAP MV; 1 male, Cashiriari, 11°52’S, 72°39’W, 04.xii.1997, 690 m, J. Grados, 7:00-8:00; 2 males, La Convención Echarate, CC., Timpia, 12°06’47.04’’S, 72°49’34.56’’W, 20-21.x.2009, 519 m, M. Alvarado y E. Rázuri, Light trap; 1 male, La Convención Echarate, CC. Pomareni, 12°15’28.38’’S, 72°50’8.8’’W, 08.xi.2009, 477 m, C. Carranza & C. Rossi, Light trap; 1 male, Campamento Mangoriari, 12°21’S, 73°02’W, 05.xi.2002, 1500 m, J. Grados; 1 male, idem except, 07.xii.2002; 1 male, idem except, 10.xii.2002; 1 male, 3.5 km ONO de Monte Carmelo, Echarate,12°26’’21.0’’S, 72°59’21.1’’W, 21.ii.2011, 1349 m, M. Alvarado & E. Rázuri; 4 males, La Convención, Echarate, CC. Ochigoteni, 12°39’31.36’’S, 73°08’ 57.71’’W, 19.x.2009, 1449 m, C. Carranza y C. Rossi, Light trap; 1 male, Campamento Segakiato,12°43’S, 73°18’’W, 01.xi.2002, 1850 m, J. Grados; 1 male, idem except, 02.xi.2002; 1 male, idem except, 08.xi.2002; 1 male, Campamento Comerciato, 12°47’S, 73°22’’W, 23.xi.2002, 1350 m, J. Grados; 1 male, Quebrada Santa Isabel, 13°02’S, 71°31’W, 13.v.2018, 1194 m, J. Grados; 1 male, San Pedro, 13°03’S, 71°32W, 27.ix.2004, 1400 m, J. Grados, HELIOTROPIUM BAIT- NIGHT. MADRE DE DIOS. 2 males, Río Los Amigos, CICRA, 12°33’S, 70°06’W, 22.vii.2006, 280 m, A. Asenjo; 1 female, Río Los Amigos, CICRA, 12°33’S, 70°06’W, 25.vii.2006, 280 m, A. Asenjo, HELIOTROPIUM BAIT- NIGHT; 1 female, idem except, 28.vii.2006; 1 male, idem except, 20.x.2006; 2 males, idem except, 28.x.2006; 3 females, idem except, 29.x.2006; 1 female, idem except, 06.xi.2006; 1 male, idem except, 23.viii-15.ix.2008; 2 males, CICRA, Río Los Amigos, 12°33’36.3’’S, 70°06’17.3’’W, 9.v.2005, 380 m, P. Centeno; 2 males, idem except, 11.v.2005; 1 male, ídem except, 13.iv.2005; 1 male, Río Los Amigos, CICRA, 12° 34’7.35’’S, 70°05’56.8’’W, 06.xii.2005, 380 m, J. Grados leg. (Introducción al estudio de los artrópodos de la Amazonía tropical - CICRA 04-15.xii.2005); 1 female, idem except, 07.xii.2005; 2 males, idem except, 08.xii.2005; 2 males, idem except, 09.xii.2005; 1 male, Tambopata Reserve, Explorer's Inn, 12°50’S, 69°17’W, 11.xi.97, 250 m, J. Grados, HELIOTROPIUM BAIT- NIGHT; 1 male, Tambopata Reserve, 12°50’S,69°17’W, 4.xii.96, 250 m, AVZ Brower, J Grados, S Green, G Lamas, JS Miller & C Snyder, MV/ UV LIGHT TRAP; 1 male, Albergue Refugio Amazonas, 12°52’30’’S, 69°24’35’’W, 12.vi.2018, 231 m, J. D. Shoobridge et al., Voucher DNA barcoding Arct·00 744 JGA - MUSM, ARCT-001081 JGA COLLECTION; 1 male, Río Alto Madre de Dios, cerca de Atalaya, 12°53’S, 71°22’W, 2.v.2006, 587 m, P. Centeno, MV LIGHT TRAP 08:00 PM - 9:00 PM; 1 male, idem except, MV LIGHT TRAP 10:00- 11:00 PM; 1 male, idem except, MV LIGHT TRAP 00:00- 1:00 AM; 2 males, idem except, MV LIGHT TRAP 11:00 PM- 00:00 AM; 1 male, idem except, 21.viii.2006, MV LIGHT TRAP 8:00- 9:00 PM; 1 male, idem except, 22.viii.2006, MV LIGHT TRAP 08:00 PM- 9:00 PM; 1 male, idem except, MV LIGHT TRAP 11:00 PM- 00:00 AM; 1 male, idem except, 16.ii.2007, MV LIGHT TRAP 10:00 PM- 11:00 PM; 1 male, Tambopata, Research Center, 13°08’S, 69°36’W, 11.v.2001, 300 m, J. Grados, HELIOTROPIUM BAIT- NIGHT; 1 male, idem except, 13.v.2003; 1 male, idem except, 12.ix.2008; 1 male, idem except, 24.v.2002, Heliotropium bait 8-9 pm; 1 male, Tambopata, Research Center,13°08’S, 69°36’W, 12.v.2001, 300 m, J. Grados, LIGHT TRAP MV; 1 male, idem except, 15.v.2001; 1 male, P.N. Bahuaja-Sonene, 13°11’35’’S, 70°07’56’’W, 17.vi.2013, 353 m, J. Grados, E. Rázuri & J. Barrientos; 1 male, idem except, 18.vi.2013; 1 male, idem except, 05.vi.2013, GENITALIA # JGA - 1137, MUSM. PUNO. 1 male, Lanlacuni, 13°29’24.9’’S, 70°25’22.8’’W, 25.x.2010, 775 m, E. Huamaní.
Geographical distribution (Fig. 6): Occurs in the French Guiana, Venezuela, Brazil, Bolivia and Peru (Cerda, 2017, Araujo et al., 2019). It is very common throughout the Peruvian Amazon, also found at montane forests (1850 m).
Barcoding: The mitochondrial DNA sequence (Peruvian Amazon) of one specimen is as follows (Voucher MUSM-Arctiinae VB # 744). GenBank access number: BankIt2604995 gnl|uoguelph|RFEWA744-19.COI-5P
AACATTATATTTTATTTTTGGTATTTGAGCTGGAATAGTAGGAACATCTTTAAGATTATTAATTCGAGCAGAATTAGGTAATCCAGGATCATTAATTGGTGATGATCAAATTTATAATACAATTGTTACAGCACATGCTTTTATTATAATTTTTTTCATAGTTATACCAATTATAATTGGAGGATTTGGTAATTGATTAGTACCTCTTATATTAGGAGCTCCTGATATAGCTTTCCCCCGAATAAATAATATAAGTTTTTGACTTTTACCCCCTTCTCTTACTTTATTAATTTCAAGAAGAATCGTAGAAAATGGAGCAGGAACAGGATGAACAGTTTACCCCCCACTTTCATCTAATATTGCCCATGGCGGTAGTTCTGTTGATCTAGCTATTTTTTCCCTTCATTTAGCAGGTATTTCTTCAATTTTAGGTGCTATTAATTTTATTACTACAATTATTAATATACGATTAAATAATCTATCTTTTGATCAAATACCTTTATTTATTTGAGCTGTAGGAATTACTGCATTTTTATTATTATTATCTCTTCCTGTATTAGCAGGAGCTATTACTATACTTTTAACTGATCGTAATTTAAATACATCATTCTTTGATCCTGCTGGAGGAGGAGATCCTATTTTATACCAACATTTATTT

Fig. 1.
Epanycles imperialis. a-b. Male, 9.5 km NE from Soroja (Junín). a. Dorsal view. b. Ventral view. c-d. Female, 10 km NO from Cajonari (Pasco). c. Dorsal view. d. Ventral view. Scale bars = 5 mm.
Epanycles satania (Schaus, 1924) new. comb.

Fig. 2.
Epanycles satania nov. comb. Guanacaste, Costa Rica. a-b. Male. a. Dorsal view. b. Ventral view. c-d. Female. c. Dorsal view. d. Ventral view. Scale bars = 5 mm.
Episcepsis sataniaSchaus, 1924: 11.
Diagnosis: Tornus margin of hind wings angular; androconial organs in the fold of anal areal composed by short yellowish hairs and small hyaline area on hindwings; uncus as long as wide; spiniform structures of the uncus wide; coremata absent; anal papillae longer than high; sclerotized part of ductus bursae less than half of total length; distal side of the sclerotized area of antrum bilobed; bursa copulatrix almost three times the papillae width.
Redescription.
Male (Fig. 2). Head.Labial palps brown with iridescent green, curved upwards reaching the vertex. Proboscis blackish brown. Frontoclypeus brown, wider than high and with the upper third iridescent green. Vertex and occiput iridescent green. Antennae bipectinate, blackish brown with bluish hue. Middle rami the same width as the flagellum axis. Thorax.Patagia, tegulae, meso and metascutum brown with iridescent green. Presence of a tymbal organ on the katepisternum. Legs brown with iridescent green. Epiphysis blackish brown. Forewing. Forewing span: 13 - 14 mm (n=3). Dorsal surface. Brown with slight greenish tint. Ventral surface. Brown with slightly blueish tint, except for the anal area which is only brown. Retinaculum brown. Hindwing. Dorsal surface. Brown with slight iridescent blueish tint. Hyaline area elongated and narrow from the base, below the disc cell and extending to before the beginning of Cu.. Ventral surface (Fig. 5b). Brown with slight iridescent blueish tint. Cell (1A+2A) - 3A modified in an elongated fold containing yellowish acicular scales. Abdomen. Brown with slight iridescent greenish tint. Two first tergites with hairy scales, dorsally; the third with few scales on dorsal part and anteriorly. Presence of coremata between the VII and VIII sternites. Genitalia (Fig. 3) (Genitalia # JGA - 1026, MUSM). Tegumen longer than wide, anterior side concave, trilobed; posterior side V-shaped with two divergent sclerotized hairy projections originating from the mesial margin. Uncus long at the base but sclerotized on its lateral parts; distal part subquadrate, sclerotized with a slight invagination on middle part and setae on the latero-distal part of its base. Juxta sclerotized and subcircular, with a constriction on its distal part. Subscaphium present, trapezoidal, with rounded edge at distal end. Valve elongated and almost straight, with the dorsal-distal part with a somewhat rounded evagination, notched distal end; ventral view, sinusoid at its base, wider at the juxta level, arched in its distal half, with setae on its mesal margin, somewhat denser at its apical end; dorsal view, mesal margin of dorsal part with numerous setae, distal area of ventral part with setae, distal end acute and sclerotized. Aedeagus wide and somewhat arched but sclerotized at its distal half. Vesica membranous, smaller than the aedeagus; with narrow, elongated, sclerotized areas on dorsal side.
Female (Figs. 2c-d). Similar to male, except for the following: frenulum light brown with two bristles, fasciculum above the CuP and androconial organ absent. Forewing. Forewing span: 14 mm (n = 1). Genitalia (Fig. 4d-f) (Genitalia # JGA - 1222, MUSM). Eighth tergite uniformly sclerotized. Eighth sternite with sclerotized areas along both sides. Anal papilla esclerotized and higher than wide, with setae throughout the surface. Posterior apophyses somewhat longer than the anterior. Antevaginal lamella membranous. Postvaginal lamella esclerotized. Ventral side of antrum sclerotized with a convex distal area. Ductus bursae enlarged with more than distal half sclerotized. Corpus bursae with signa on the entire surface.
Examined material. MEXICO: 1 male, Orizaba, Estado de Veracruz, junio (MCM); 1 female, Chajul [Boca de], Municipio Marqués de Comillas, Chiapas, de mayo de 1981 Leg. J. de la Maza, volando de día; 1 male, idem except, 8 de noviembre de 1992, Leg. A. Pescador; 1 female, Reserva de la Biosfera Montes Azules, estación Chajul, Municipio de Marqués de Comillas, Chiapas, 14 al 21 de octubre de 1991 Leg. J León; 1 male, idem except, 27 de enero al 1 de febrero de 1992. Leg. J León; 1 male, Ejido Loma Bonita, Municipio Marqués de Comillas, Chiapas, del 26 de diciembre de 1991 al 2 de enero de 1992. Leg. A Zacarías; 1 male, Ejido Loma Bonita [8km al este de Boca de Chajul], Municipio de Marqués de Comillas, Chiapas del 24 de mayo de 1992. Leg J. León. All in CART. GUATEMALA. 1 male, Panzos, Vera Paz, Champion / B.C.A. Lep. Het. Epanycles imperialis / Godman-Salvin, Coll. 97.–52. (NHMUK); 1 male, Alta Verapaz, Cobén, Santa Lucía Lachua, 270 m, 22-24.iv.2002, Col. Bailey y Monzón (Genitalia # JGA - 905, MUSM). COSTA RICA. 1 male, Guanacaste, nr. Santa Cecilia, El Bosque Nuevo, 11º03.38 N, 85º21.34 W, 27 Aug. 2012, at light, leg. M. Boppré (Genitalia # JGA - 1026, MUSM); 1 male, idem except, 23 Aug. 2012; 1 female, idem except, 05 Sep. 2012 (Genitalia # JGA - 1222, MUSM).
Geographical distribution: This species occurs in Mexico (Santa Rosa, type-locality; Type Cata. Nº 25889 U.S.N.M.) [Examined], Guatemala, Costa Rica and Panamá.
Comparative morphology
The genus Episcepsis was proposed by Butler (1877), including as type-species Episcepsis venata. This species was described on the basis of a single female collected at a tributary of the Amazonas River (Brazil) and a brief history on the genus was provided by Mantilla & Grados (2021). The type locality of Episcepsis satania is Santa Rosa (Mexico). At the end of its original description, Schaus (1924) mentioned that this species lacks a tuft of long hairs on the inner margin of hind wings, a condition that is similar to the typical androconial organs observed in species belonging to Episcepsis. This citation indicates some uncertainty of Schaus (1924) about the generic allocation of E. satania.
Epanycles imperialis (Euchromia imperialis) was described by Walker (1854) based on material collected by H.W. Bates at Santarem (Brazil). Apart from its unmistakable coloration, this species possesses a fold on the ventral side of hind wings, where androconial organs are present, and yellowish hairy scales (Fig. 5a). However, this fold is not very conspicuous and was unnoticed in the original species description (Walker 1854). Epanycles satania new. comb. presents a similar coloration pattern, in addition to the fold in the anal area of the ventral part of hind wings. The latter is very noticeable and has short needle-like scales (Fig. 5b). On the other hand, the hyaline area of hind wings is wide in Epanycles imperialis and very reduced in Epanycles satania new. comb.
Differences between both species regarding the morphology of components of genital capsules are: Epanycles imperiales presents an uncus longer than wide (vs. as long as wide in Epanycles satania new. comb.); anterior invagination of juxta wide (vs. narrow in E. satania new. comb.); mesal parts of valves separated in ventral view (vs. close to each other in E. satania new. comb.); spiniform projections from the posterior margin of the uncus wider in E. satanianew. comb. than in E. imperiales. Both species present the proximal support of sternite VIII and the sternite VIII sclerotized and T-shaped. Only E. imperialis presents a coremata. Females of E. satania new. comb. differ by having anal papillae longer than high (vs. higher than long in E. imperialis); sclerotized part of ductus bursae less than half of total length (vs. more than half of total length in E. imperialis); distal side of the sclerotized are of antrum bilobed (vs. convex in E. imperialis); bursa copulatrix almost three times the width of anal papillae (vs. two times in E. imperialis).
Geographical distribution
Two species were identified based on the samples analyzed: Epanycles imperialis and Episcepsis satania. Epanycles imperialis is known to occur in French Guiana, Venezuela, Brazil, Peru, Bolivia (Grados, 1999; Cerda, 2017; Araujo et al., 2019) and has also been reported in Mexico, Guatemala and Costa Rica (Hampson, 1898; Turren-Díaz & Pescador, 2013; Draudt, 1916; Araujo et al., 2019). E. satania new. comb. Occurs in Mexico, Guatemala, and Costa Rica (Schaus, 1924 and material examined in this study).
Both species are morphologically similar and very often misidentified (Beutelspacher, 1988; Hernández-Baz & Grados, 2004). Draudt (1916) presented the geographical distribution of E. imperialis, before the description of Episcepsis satania. According to our study, specimens from Central America belong to Episcepsis satania.
It is highly probable that the records of Araujo et al. (2019) regarding Epanyclesimperialis in Central America are, in fact, Episcepsis satania. Bold System sequences of “Epanycles imperialis” analyzed in this work correspond to the morphology of Episcepsis satania.
DNA Barcoding
From the Cytochrome oxidase 1 (CO1) sequencing, a matrix of genetic distances was obtained (Table I). The maximum value of genetic distances between specimens from Costa Rica (CR)and Panama (PA), identified as “Epanycles imperialis” was 0.19%; even three comparisons showed identical genotype. Comparing E. imperialsfrom Peru (PE) to specimens from Central America (“Epanycles imperialis”), genetic distances correspond to 2.6-2.8%. Genetic distances between a French Guiana specimen and others from Central America were 2.7-3.3%. Lastly, distances between Peru and French Guiana specimens were 2%.
Our results help us to discriminate between the two taxa, as proposed by Hebert et al. (2003) as well as for other Arctiinae species in the Neotropical region (Vincent, 2012; Vincent, & Laguerre, 2013; Vincent, 2018). However, it is important to mention that the differentiation threshold might vary (Wiemers & Fiedler, 2007), depending on the evolutionary history of the groups analyzed.
Based on genetic evidence, we hypothesized the occurrence of E. imperialis in South America and a second species mistakenly identified as such that occurs in Central America. At least, in relation to E. imperialis, the external morphological characters and the male genitalia of specimens from Brazil, French Guiana (Cerda, 2017; Araujo et al., 2019) and Peru have no significant differences. Although DNA barcoding is a powerful tool that could give us indications of the differentiation between two species, morphological and, biogeographical data should also be considered before nomenclatural changes are made.
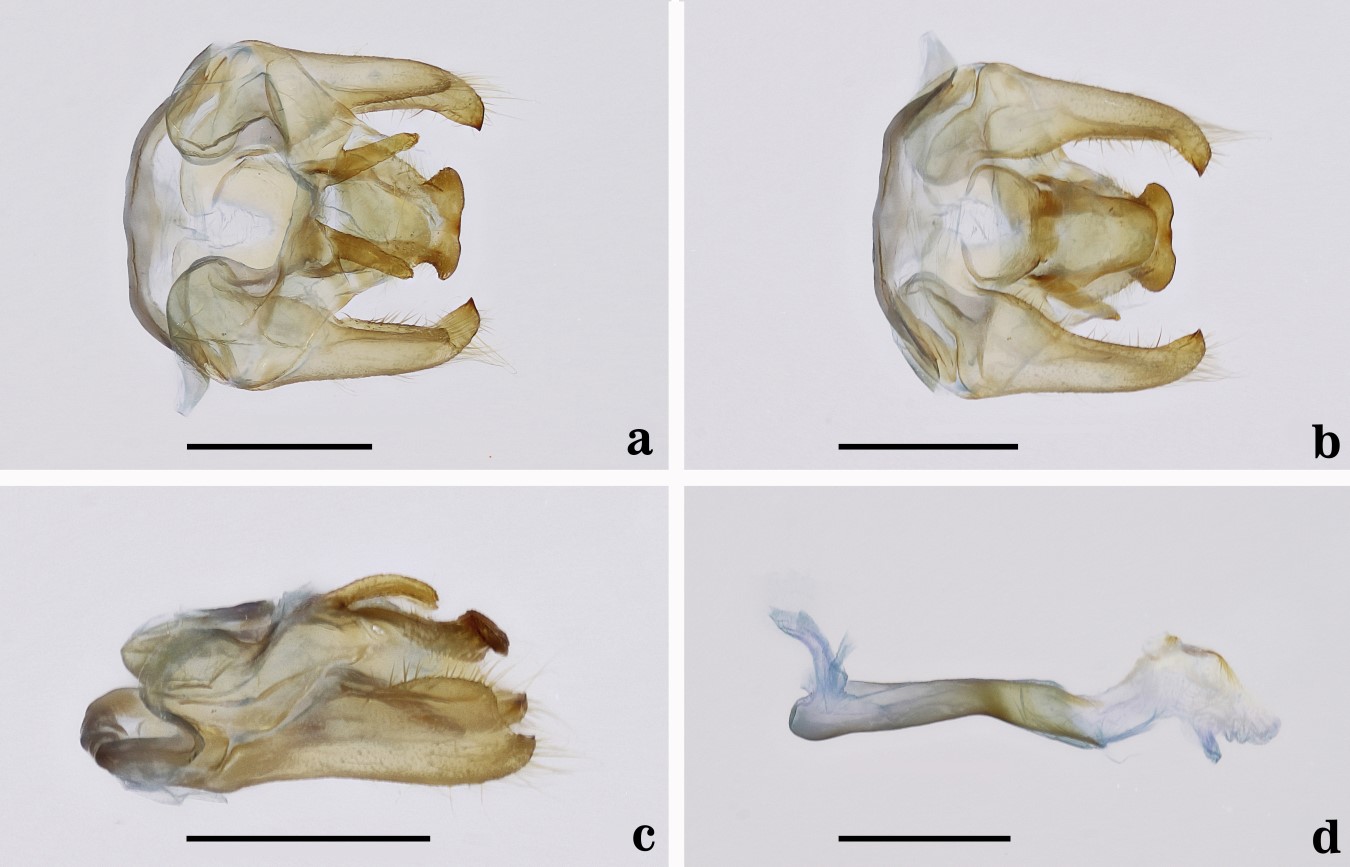
Fig. 3.
Male genitalia of Epanycles satania nov. comb. (Genitalia JGA # 1026-MUSM). a. Dorsal view. b. Ventral view. c. Lateral view. d. Aedeagus. Scale bars = 1 mm.
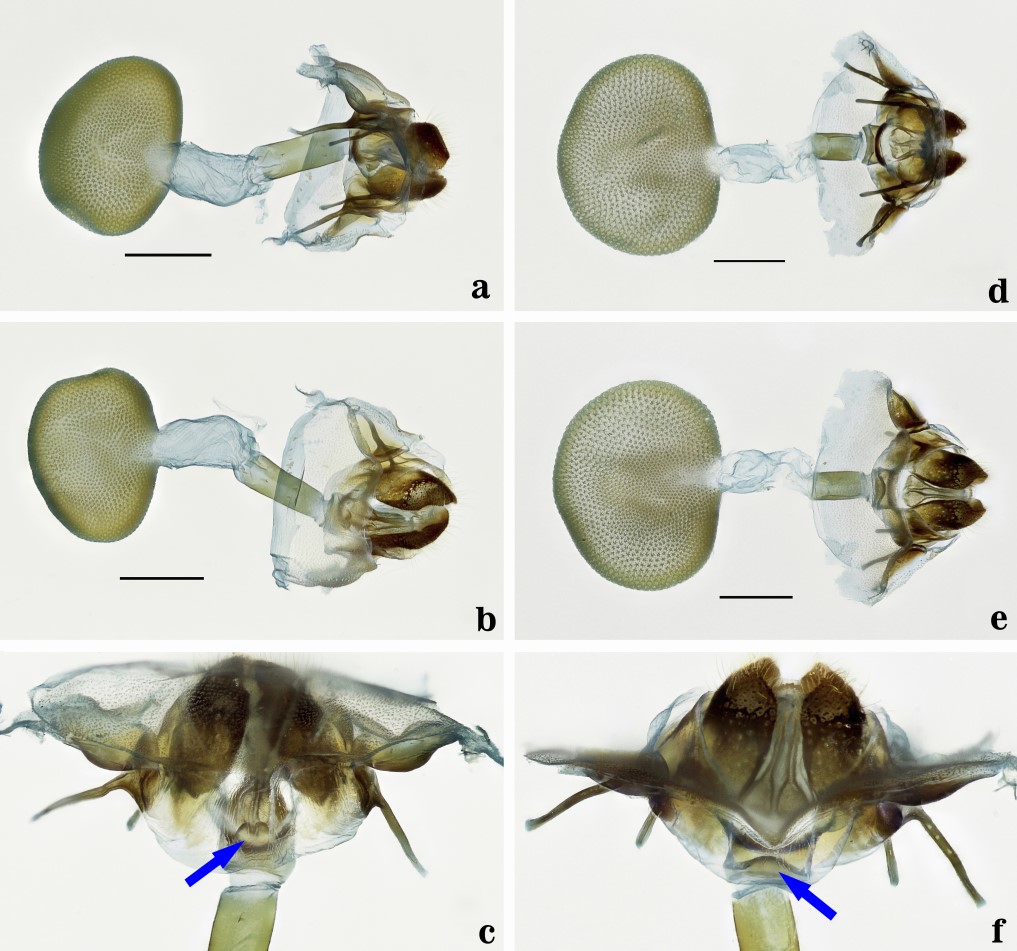
Fig. 4.
Female genitalia. a-c. Epanycles imperialis (Genitalia JGA # 1223-MUSM). a. Dorsal view. b. Ventral view. c. Antrum view. d-f. Epanycles satania (Schaus) nov. comb. (Genitalia JGA # 1222-MUSM). d. Dorsal view. e. Ventral view. f. Antrum view. Scale bars = 1 mm.

Fig. 5.
Ventral view of hindwings. a. Epanycles imperialis. b. Epanycles satania nov. comb.
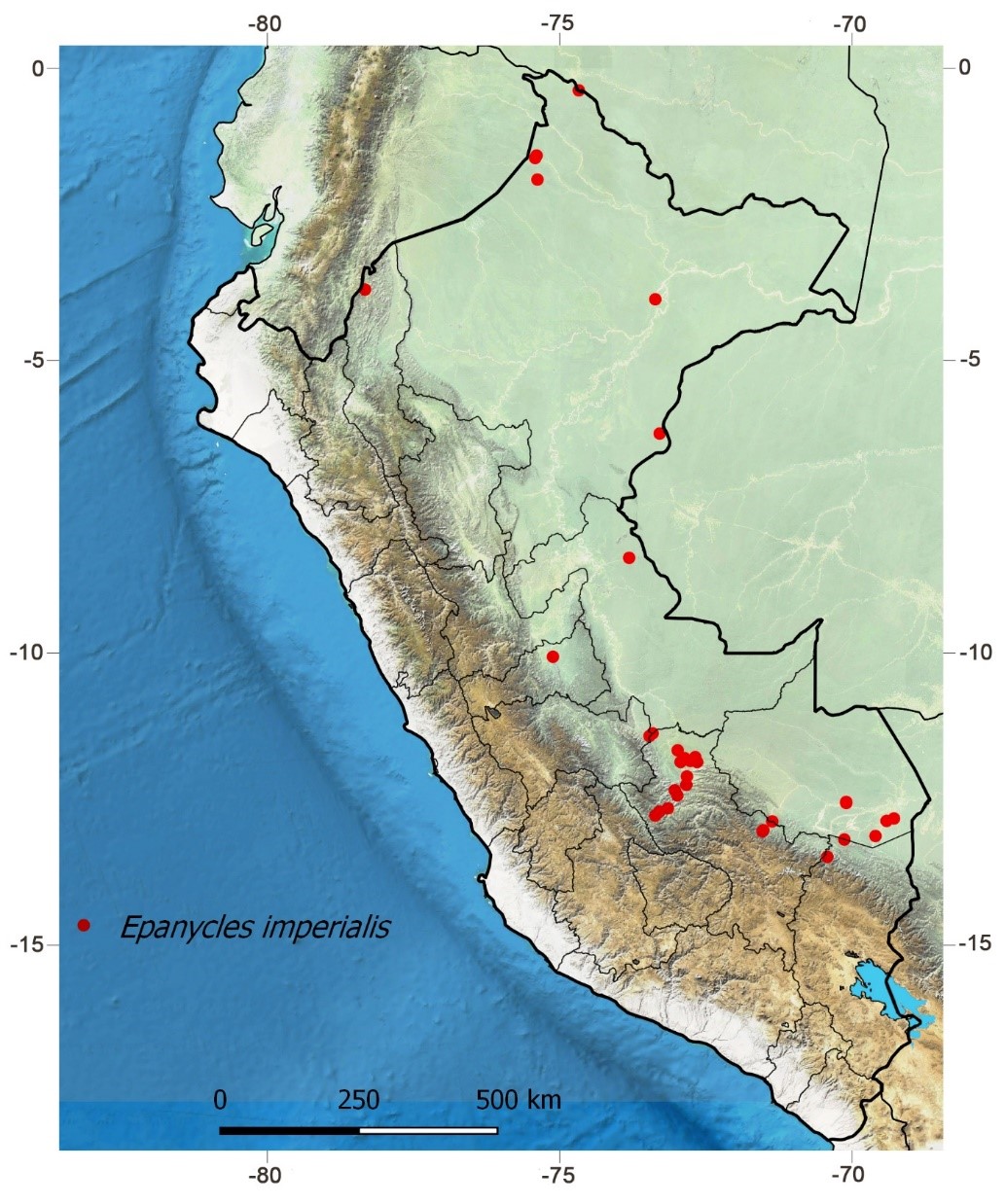
Fig. 6.
Geographical distribution of Epanycles imperialis in Peru.
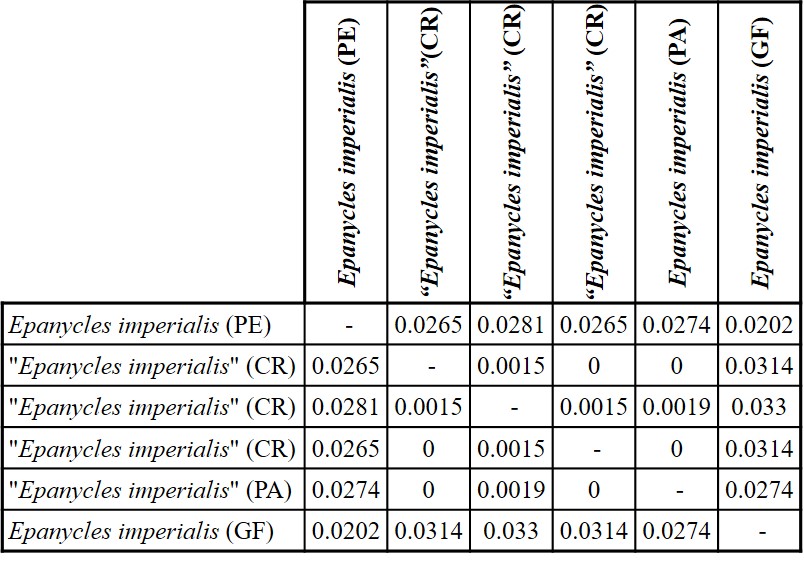
TABLE I.
Genetic distance calculated from DNA barcode sequence (Kimura-2-parameter) between Epanycles imperialis (Peru) and “Epanycles imperialis” from Costa Rica and Panama (Epanycles satania).
CONCLUSION
The genus Epanycles has been considered monotypic for more than 145 years. In this study, we propose that Epanycles imperialis and Episcepsis satania share a common evolutionary history and should be reunited in the same genus, based on data from external morphology, genital morphology of males and females, geographic distribution, DNA barcoding and phylogenetic studies of some genera of Ctenuchina (Grados & Mantilla, unpublished data). Epanycles satania new. comb. is proposed as a new generic combination.
Acknowledgments
The authors would like to thank Alberto Zilli and Geoff Martin for permission to examine type specimens deposited at NHMUK. To Rainforest Expeditions for the invitation to participate in Discovery new species project. To M. Boppré for the loan of material from Costa Rica and J. Cerda for sharing information on French Guiana specimens. To Christian Wieser, Michel Laguerre and Carlos Vaamonde for collecting, identifying and sharing the sequence of French Guiana specimen, respectively. Suggestions of two anonymous reviewers significantly improved this manuscript. This work is part of the master’s thesis of Juan Grados.
REFERENCES
Araujo, E.C., Pinheiro, L.R., & Duarte, M. (2019) Comparative morphology and taxonomy of the tiger moth genus Epanycles Butler (Lepidoptera, Erebidae, Arctiinae, Arctiini, Ctenuchina), with notes on related genera. Zootaxa, 4613(1), 187-199.
Beutelspacher, C.R. (1988) Catálogo de la colección Roberto Müller (Lepidoptera) del Museo de Historia Natural de la Ciudad de México VI. Familias Castniidae, Zygaenidae, Ctenuchiidae y Nolidae. Anales del Instituto de Biología UNAM, 58(1), 455-473.
Butler, A.G. (1876) On the subfamilies Antichlorinae and Charideinae of the lepidopterous families Zygaenidae and Arctiidae. Journal of the Linnean Society of London, 12(63), 408-433.
Butler, A.G. (1877) Illustrations of Typical Specimens of Lepidoptera Heterocera in the collection of the British Museum Vol I. British Museum, London, England.
Cannon, G. (1937) A new biological stain for general purposes. Nature, 139, 549.
Cannon, G. (1941) On Chlorazol black E and some other new stains. Journal of the Royal Microscopical Society, 61, 88-95.
Carayon, J. (1969) Emploi du noir chlorazol en anatomie microscopique des insects. Annales de la Société entomologique de France (N.S.), 5(1), 179-193.
Cerda, J.A. (2017) Ctenuchina de Guyane Française Lepidoptera, Erebidae, Arctiinae, Arctiini (partie 1). Mémoires de la Société linnéenne de Lyon, 7, 1-183 + 20 pls.
Common, I.F.B. (1990) Moths of Australia. University Press, Melbourne, Australia.
Comstock, J.H., & Needman, J.G. (1898) The wings of insects. American Naturalist, 32(373, 374, 376, 377, 378, 380, 382, 384), 43-48, 81-89, 231-257, 335-340, 413-424, 561-565, 769-777, 903-911.
Comstock, J.H., & Needman, J.G. (1899) The wings of insects. American Naturalist, 33(386, 391 & 395), 117-126, 573-582, 843-860.
Draudt M. ([1916-1919]) Familie: Syntomidae. Die Gross-Schmetterlinge der Erde, Vol. 6 (ed., Seitz, A.), pp. 37-230. A. Kernen, Stuttgart.
Grados, J. (1999) Lista Preliminar de los Ctenuchinae (Lepidoptera: Arctiidae) de la Zona Reservada Tambopata - Candamo, Madre de Dios, Perú. Revista peruana de Entomología, 41, 9-14
Grados, J., Mantilla, K., & Ramírez, J.J. (2020) The genus Parascepsis Dognin, 1923 (Lepidoptera, Erebidae, Arctiinae, Ctenuchina) in Peru, with the description of three new species, a new combination and their geographical distributions. Zootaxa, 4868(2), 221-242.
Hampson, G. (1898) Catalogue of the Lepidoptera Phalaenae in the British Museum. Vol. I. British Museum, London, England.
Hebert, P. D., Ratnasingham, S., & De Waard, J. R. (2003) Barcoding animal life: cytochrome c oxidase subunit 1 divergences among closely related species. Proceedings of the Royal Society of London, Series B, Biological Sciences, 270(Suppl. 1), S96-S99.
Hernández-Baz, F., & Grados, J. (2004) Lista de los Ctenuchinae (Insecta: Lepidoptera: Arctiidae) del estado de Veracruz, México y algunas notas sobre su riqueza en el continente americano. Folia Entomológica Mexicana, 43(2), 203-213.
Klots, A. (1970) Lepidoptera. Taxonomist's Glossary of Genitalia in Insects (ed. Tuxen, S.L.), pp. 97-111. Munksgaard, Denmark.
Kreusel, B. (2000) Phylogenetische Analyse der “Ctenuchinae” (Lepidoptera: Arctiidae). Dissertation zur Erlangung des Doktorgrades (Dr. re. nat.) der Mathematish-Naturwissenschaftlichen Fakultät der Rheinischen Friedrich-Wilhelms-Universität Bonn.
Mantilla, K., & Grados, J. (2021) Four new Neotropical species of Episcepsis Butler, 1877 (Lepidoptera: Erebidae: Arctiinae). Zootaxa, 5020(2), 352-366.
Miller, L.D. (1970) Nomenclature of wing veins and cells. Journal of Research on the Lepidoptera, 8(2), 37-48.
Robinson, G.S. (1976) The preparation of slides of Lepidoptera genitalia with special reference to the Microlepidoptera. Entomologist's Gazette, 27(2), 127-132.
Schaus, W. (1924) New species of moth in the United States National Museum. Proceedings of the United States National Museum, 65(7), 1-74.
Tamura, K., Stecher, G., & Kumar, S. (2021) MEGA 11: Molecular Evolutionary Genetics Analysis Version 11. Molecular Biological and Evolution, 38(7), 3022-3027.
Turrent-Díaz, R., & Pescador, A. (2013) La fauna de las tribus Euchromiini y Ctenuchini (Lepidoptera, familia Erebidae) de la región de Chaujil Chiapas (Marques de Comillas). Revista de la Sociedad Mexicana de Lepidopterología, Nueva Serie, 1(1), 1-15.
Vincent, B. (2012) A review of the genus Gorgonidia Dyar, 1898, with the description of one new species (Lepidoptera: Erebidae, Arctiinae, Arctiini). The European Entomologist, 4(1), 33-54.
Vincent, B. (2018) Two new species of the Neotropical Lophocampa hyalinipuncta (Rothschild) group (Lepidoptera, Erebidae, Arctiinae). ZooKeys, 788, 57-67.
Vincent, B., & Laguerre, M. (2013) Four new Neotropical Lophocampa species with a redescription of Lophocampa atriceps (Hampson) (Lepidoptera, Erebidae, Arctiinae). ZooKeys, 264, 47-69.
Walker, F. (1854) List of the specimens of Lepidopterous insects in the collection of the British Museum: Part 1- Lepidoptera Heterocera. Trustees of British Museum, London, England.
Wiemers, M., & Fiedler, K. (2007) Does the DNA barcoding gap exist? – a case study in blue butterflies (Lepidoptera: Lycaenidae). Frontiers in zoology, 4(1), 1-16.
Wootton, R.J. (1979) Function, homology and terminology in insect wings. Systematic Entomology, 4, 81-93.
Zerny, H. (1912) Syntomidae. Lepidopterorum Catalogus, Pars 7 (ed. Wagner, H.), pp. 179. W. Junk, Berlin.
Notas de autor
gradosjuan@hotmail.com