Artículos
Description of a new species of Fannia (Diptera: Fanniidae) from the Argentinean Patagonia
Descripción de una nueva especie de Fannia (Diptera: Fanniidae) de la Patagonia Argentina
Description of a new species of Fannia (Diptera: Fanniidae) from the Argentinean Patagonia
Revista de la Sociedad Entomológica Argentina, vol. 82, núm. 2, 2023
Sociedad Entomológica Argentina
Recepción: 08 Marzo 2023
Aprobación: 08 Mayo 2023
Abstract: A new species of the genus Fannia (Diptera: Fanniidae) is described, Fannia adelaidae sp. nov. This new species was collected in the Patagonian region in the province of Neuquén, within the Lanín National Park using baited traps. Photographs of diagnostic characters are provided for both sexes. Its possible phylogenetic relationship with other species of the same region is discussed.
Keywords: Anthracina-group, Calyptratae, Southern South America, Taxonomy.
Resumen: Se describe una nueva especie del género Fannia (Diptera: Fanniidae), Fannia adelaidae sp. nov. Esta nueva especie fue colectada en la región Patagónica, en la provincia de Neuquén, dentro del Parque Nacional Lanín, usando trampas con cebo. Se proveen fotografías de los caracteres diagnósticos para ambos sexos. Se discute además brevemente su posible relación filogenética con otras especies de la misma región.
Palabras clave: América del Sur Austral, Calyptratae, Grupo anthracina, Taxonomía.
INTRODUCTION
Faniid flies (Diptera: Fanniidae), with approximately 300 known species occur in all geographic regions of the world. The species of Fanniidae are currently divided into five recognized genera: Fannia Robineau Desvoidy, Euryomma Stein, Piezura Rondani, Australofannia Pont, and Zealandofannia Domínguez & Pont (Domínguez & Pont, 2014).
Only Fannia and Euryomma are present in Argentina, and their species have been revised and phylogenetic and biogeographic hypothesis of their evolution have been recently proposed (Domínguez, 2007; Domínguez & Roig-Juñent, 2008, 2011, 2017). The Fanniidae are believed to be basically inhabitants of forests, and are relatively rare in open landscapes and wetlands (Rozkošný et al., 1997). The seven known species belonging to the Fannia anthracina species-group: Fannia albitarsis Stein, Fannia anthracina (Walker), Fannia bogoti (Stein), Fannia confusa Pont & Carvalho, Fannia pusilla (Bigot), Fannia schnusei Stein and Fannia setosa (Bigot) (Pont & Carvalho, 1994; Domínguez & Roig-Juñent, 2008) are endemic to the Argentinean and Chilean Patagonia and their distributions are related to the Notophagus Blume forests, with the exception of F. albitarsis which is also found in Peru, Bolivia, South Africa, New Zealand, and Australia. This very particular group shows a series of apomorphous characters such as the black to steely blue ground colour of the body; male fore leg with several tarsomeres white and often modified in shape, male mid leg with basal tarsomere with a ventral crest, followed by a short strong seta; male hind femur with a ventral tubercle or at least a ventral tuft of setae in apical half. However, although the family is believed to dwell in forests, as mentioned above, some species of the genus not belonging to the above-mentioned species group, such as Fannia fusconotata (Rondani) and Fannia heydenii (Wiedemann) have been found in open arid shrub lands and open woodlands of Prosopis Linneaus in the province of Mendoza (Domínguez, 2007).
Males of almost all species form swarms under tree branches and above forest paths, members of these swarms hover in the air like hover-flies (Syrphidae) (Rozkošný et al., 1997). Among the southern South American species, the males of F. fusconotata have been found swarming very low, near water streams and associated vegetation and F. hermani Domínguez was found swarming directly above a water stream (Domínguez, 2007).
The Lanín National Park, where this new species was collected is located in the Patagonian region of South America also known as southern South America. South America is considered a continent of hybrid origin and can be divided for biogeographic studies in two areas, namely the southern temperate and the northern tropical (Neotropical) area (Crisci et al., 1991). The biota of southern South America and northern South America never constitute a monophyletic group, given that many groups of plants and animals, including insects, endemic to southern South America are related to other southern temperate areas such as Australia, New Guinea, New Caledonia, Tasmania and New Zealand (Crisci et al., 1991). This is the case of the Fannia anthracina species-group (Pont & Carvalho, 1994), that includes species endemic to the Argentinean and Chilean Patagonia and which was found to fit this biogeographical pattern, i.e., it was more closely related to species of Fannia from New Zealand and Australia than to other species from the Neotropical region (Dominguez & Roig-Juñent, 2008, 2011).
The medical and hygienic importance of the wide-spread species of Fannia such as F. canicularis (Linnaeus), F. femoralis (Stein), F. incisurata (Zetterstedt), F. pusio (Wiedemann), F. scalaris (Fabricius) are well known. Fannia canicularis and F. scalarishave been reared from various decaying materials in gardens. Moreover, the larvae of F. scalaris are frequent in cesspools, latrines and dunghills, having also been reared, accompanied by F. canicularis from human faeces. Some of the most abundant species occur regularly in agricultural pens used for breeding pigs, cattle, horses or fowls, and in fur farms. The larvae apparently develop in animal droppings and dung (Rozkošný et al., 1997). Nevertheless, the medical and hygienic importance of most southern South American species is unknown. Fannia albitarsis has been found to be an important nuisance in poultry farms in the south of the province of Buenos Aires (Perotti, 1998). Fannia fusconotata, as well as F. canicularis and F. scalaris are believed to cause myiasis in man and in cattle (Mazza & Oribe, 1939; Oliva, 1997). However, numerous and relatively recent forensic studies have reported the presence of the genus Fannia in South America, mainly in Brazil, Colombia and Argentina (Souza & Linhares 1997; Carvalho et al., 2000; Wolff et al., 2001; Centeno et al., 2002; Iannacone, 2003; Aballay et al., 2008; Battán Horenstein et al., 2010). And in more specific studies about the family Fanniidae, Aballay et al. (2012) explored for the first time the importance of adult Fanniidae as PMI indicators, and Domínguez & Aballay (2008) and Quiroga & Domínguez (2010) described two new species of the genus associated to pig carcasses.
The main aim of this study is to describe a new species of de genus Fannia from southern South America, specifically from the Lanín National Park and to provide photographs of its diagnostic characters as well as a brief discussion of its relationship with previously described species of the Argentinean Patagonia.
MATERIAL AND METHODS
The study area, the Lanín National Park is located within the North Patagonian Andean Biosphere Reserve which encompasses the cold-temperate forests of the northern Patagonian mountainous stripe. These forests are geographically isolated from tropical and subtropical forests of South America, and shelter a large number of endemic plant and animal species. The vegetation of the Lanín National Park is composed of a large number of endemic species that vary along with the elevation of the region. It is dryer towards the East (Patagonian steppe); whereas toward the West, humid forests and intrusions of the Valdivian forests form the main landscape; and in higher altitudes of the Andean mountain ridge -beyond the forest line- high Andean environments can be found Morrone, 2006). The type series of Fannia adelaidae sp. nov. was collected next to the dry De Los Pinos river bed, beyond the Monkey Puzzles, i.e., Araucaria araucana forest.
Adults were collected using baited traps; the bait consisted of putrescine that is fermented bone meal. All the type series was collected next to the De Los Pinos river, near the Curruhué lake (39°52´36,26´´ S, 71°27´12,16´´W) and were conserved as dry pinned specimens. The material is deposited at the CEI collection, within the Laboratorio de Entomología, (IADIZA. CCT CONICET Mendoza, Argentina).
Measurements are expressed as follows: body length: anterior margin of head (frons), excluding antennae, to apex of abdomen; frons width: narrowest distance between eye margins; vitta width: measured at uppermost pair of frontal setae; frontal setae length: relative to length of flagellum; parafacial width: relative to width of flagellum at its base; shape of flagellum: length/width; palp shape: basal width relative to apical width; length of ventral pubescence of mid tibia: relative to tibial width.
The following abbreviations are used in the descriptions: Head: fr, frontal seta; pocl, postocular seta; orb, orbital seta. Thorax: acr, rows of acrostichal setulae; acr s, acrostichal seta; dc, dorsocentral seta; ial, intra-alar seta; npl, notopleura; prepm, proepimeral seta; pra, prealar seta; pprn, postpronotal seta; spal, supraalar seta. Legs: C, coxa; F, femur; T, tibia; ., anterior seta; ad, anterodorsal seta; av, anteroventral seta; ., dorsal seta; ., posterior seta; pv, posteroventral seta; pd, posterodorsal seta; ., ventral seta; ad, anterodorsal seta. Morphological terminology mainly follows McAlpine (1981) with the exception of the following terms for genitalia: pregonite and postgonite (paramere and gonopod of McAlpine).
For genitalia examination, the abdomen was removed from a dry specimen and heated in 10% KOH for 10–15 minutes. The abdomen was then transferred to acetic acid, and then to glycerine. The postabdominal structures were separated from the rest of the abdomen. Examination of genitalia was done using a compound microscope. After examination, the genitalia and the rest of the abdomen were placed in glycerine in a plastic microvial and pinned directly under the specimen.
Photographs were obtained using a Leica S6D Stereomicroscope with an attached digital colour Leica EC3 camera.
RESULTS
Fannia adelaidae Domínguez, new species
LSID: urn:lsid:zoobank.org:act:A640E63D-706F-4834-A0FC-2543FF51D2AC
Diagnosis: Medium-sized species, completely black, except for abdomen which is shining dark blue. Male thorax with 4+5 very thin and irregular acr; fore tarsi black, not expanded nor flattened; mid tarsomere 1 with a basal ventral crest, followed by a short and stout seta; C3 bare at apex of posterior margin; F3 curved, with a very prominent preapical protuberance on ventral surface. Epandrium strongly broadened in anterior margin, bell-shaped; surstyli simple, marginal, shorter than epandrium and strongly oar-shaped. Female parafacialia light silvery grey, with a very striking division line between the dark black frons and parafacilia at the level of the antennal scape.
Description
Male: body length 5–7 mm.
Head: Frontal vitta black, at narrowest point 1-1.5x width of anterior ocellus. Frons covered with silvery pruinescence, at uppermost pair of fr 0.5x width of anterior ocellus. Nine to 10 frontal setae with weaker setae in between, 1.4x length of flagellum. Eyes bare, antero-internal facets much larger than remaining; pocl hair-like, as long as flagellum. Face and parafacial silvery grey; parafacial at base of flagellum 0.4-0.5x width of same and bare. Facial carina black. Gena dark grey to black. Scape and pedicel black; flagellum 2.3x as long as wide and heavily covered with dull-grey pruinescence. Arista black, very short pubescent. Palpi dark brown to black, the apex 1.5 to 2x width of base.
Thorax: Shiny black; 4+5 very thin and irregular acr, 1 prescutelar acrs; 2+3 dc. pprn covered with long, hair-like setae; 1 pra dorsad spal, 0.33x length of same, and another the same size next to transverse suture; prealar area with very thin hair-like ground setulae; 2 prepm densely surrounded by long hair-like setulae.
Legs: Completely black. F1 with 1 row of pd as long as femoral width; 4 rows of hair-like p, longer than femoral width; 1 row of pv, short at base and as long as femoral width at apex. T1 with a long subapical pd; 1 apical v and 1 apical pv. Foretarsi black, not expanded nor flattened (Fig. 1a). F2 with 1 row of ad, hair-like and shorter than femoral width; 1 row of av, basally as long as femoral width and in femoral width; 1 row of av, basally as long as femoral width and in apical ¼ forming a very short and closely placed row; 1 row of long pv hair-like in basal 3/4, shorter and stouter towards apex that form a small clump at apex; 1 row of p, long and hair-like throughout. T2 thin, with a strong subbasal protuberance; ventral pubescence long, as long as tibial width; 1 submedial and 4 apical a; 1 subapical ad; 1 strong apical av-v; 1 submedial and 1 apical pd; 1 apical p; 1 very long apical pv (Fig. 1b). Mid tarsomere 1 with a basal ventral crest, followed by a short and stout seta (Fig. 1b). C3 bare at apex of posterior margin (Fig. 1c). F3 curved, with a very prominent preapical protuberance on ventral surface; 1 row of ad longer and dorsally directed towards apex; 1 row of av as long as femoral width at base, longer at apex; with a preapical tuft of long pv 2.5x femoral width, coinciding with the preapical ventral protuberance; pd and p with short and thin setae in single rows (Fig. 1c). T3 with 2 short d of almost equal length; 1 submedial and 1 apical ad, lacking medial row; 2 medial and 1 apical av (Fig. 1c).
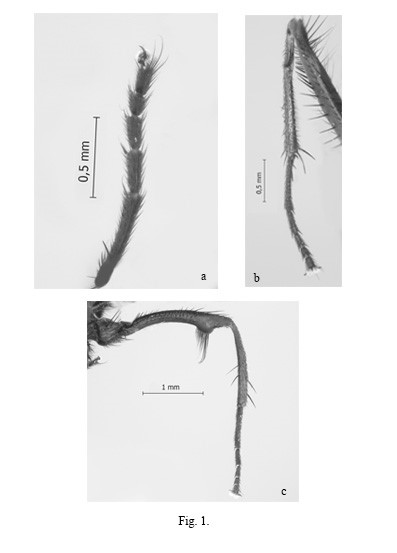
Fig. 1.
Fannia adelaidae Domínguez. Male: a) Fore tarsi, anterior surface; b) Mid leg, posterior surface (vc: ventral crest, s: stout setae, avr: anteroventral row); c) Hind leg, anterior surface.
Wings: translucent to very slightly brown tinted. Lower calypter broad and oval, upper rounded, both white with dark brown outer margins. Halteres dark brown to black.
Abdomen: Weakly elongated, tergite 2 slightly broader than tergite 4. Shining dark blue. Sternite 1 covered with few medium hair-like setae.
Postabdomen: Posterior margin of sternite 5 rounded (Fig. 2a). Hypandrium thin, hypandrial arms with lateral triangular extension; postgonites inconspicuous; hypandrial base with a circular perforation (Fig. 2b); aedeagus membranous. Epandrium strongly broadened in anterior margin, bell-shaped; cercal plate fused tapering at apex (anterior) and very slightly bifid in this region; bacilliform process cork-screw shaped; surstyli simple, marginal, shorter than epandrium and strongly oar-shaped; weakly connected to internal margin of epandrium (Fig. 2b, c).
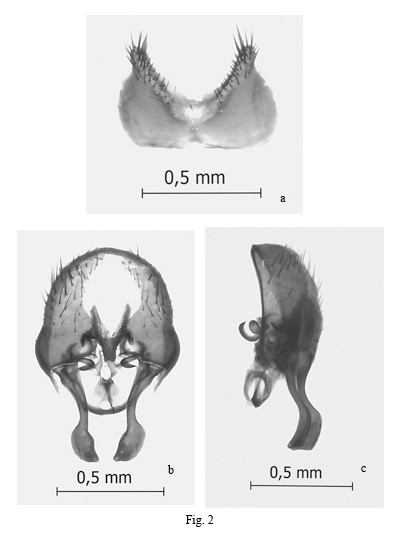
Fig. 2.
Fannia adelaidae Domínguez. Male genitalia: a) Sternite V, ventral view; b) epandrium and related structures, ventral view; c) Epandrium and related structures, lateral view. (ep: epandrium, cp: cercal plate, bp: bacilliform process, hyp: hypandrium, sur: sursstylus)
Female, differs from male as follows:
Head (Fig. 3a): Ocellar triangle long, reaching lower orb. Frons shining black and vitta dusty black, very slightly dark reddish, the distance between eye margins wider than 0.33x width of the head. Viewed from above, parafacialia strikingly light silvery grey, with a very defined division line between frons and parafacilia at the level of the antennal scape. Ten strong fr, slightly longer than flagellum and bearing 6 to 7 setulae half as long, in between. Upper and lower orb present, and 1 row of fronto-orbital setulae.
Legs: F1 with 1 row of short .. Fore tarsi normal in shape, dark brown to black. F2 with 1 row of short and setulose av; 1 row of pv setae. T2 cone-shaped, the base narrower than the apex, pubescence on ventral surface absent. Mid tarsomere lacking basal ventral crest and seta. F3 lacking preapical ventral protuberance, with 1 row of long ad that are longer and stouter towards the apex; 1 row of short av, and 2 subapical long av; posteroventral surface bare.
Abdomen: Heart-shaped, tergite 2 more than 2x width of tergite 4, this shaped more pronounced than in male. Shining dark blue.
Ovipositor and spermathecae (Fig. 3b): Cerci normal, short; anal plate longer than broad and uniformly covered with setulae; sternite 8 reduced to an anterior pair of circular plates bearing two setae and 4 to 5 setulae; postabdominal spiracles 7 and 8 on tergite 6. Three smooth-surfaced pear-shaped spermathecae.

Fig. 3.
Fannia adelaidae Domínguez. Female: a) Head; b) Ovipositor and spermathecae.
Material examined. Type series only.
Type material: Holotype ♂, Argentina: Neuquén: Parque Nacional Lanín, Lago Curruhué, Arroyo los Pinos, 1090 msnm; 39°52´36,26´´S, 71°27´12,16´´W 14/XII/2013 Patitucci leg (baited trap). Paratypes, 2♂♂, same data as holotype; 1♂ same data as holotype except for: Olea leg; 15 ♀♀ same data as holotype; 5 ♀♀ same data as holotype except for: Olea leg; 6♀♀ same data as holotype except for: Mulieri leg.
Distribution: Argentina: Province of Neuquén, De Los Pinos river, near the Curruhué lake.
Etymology. The species name is a matronym in honour of Adelaida Esther Canizzo, to whom it is dedicated in loving memory.
DISCUSSION
Fannia adelaidae sp. nov. could belong to the antracina species-group (Pont & Carvalho, 1994). It shares many diagnostic morphological characters as well as a concordant geographical distribution with the species of this group. The most relevant of these morphological characters are: the black and dark steely blue colouration; the presence of a basal ventral crest, followed by a short and stout seta on mid tarsomere 1; the ventral protuberance of the hind femur, as well as many features of the male genitalic structure. But, in this new species, the fore-tarosomeres are black and with a normal shape, not white, nor modified in shape and the presence of these features are strong apomorphies of the anthracina species-group. Nevertheless, a phylogenetic analysis with the inclusion of F. adelaidae sp. nov. could clarify its position within Fanniidae and its phylogenetic relatedness to species of Fannia from Australia and New Zealand.
As mentioned above, the type series of F. adelaidae sp. nov. was collected next to the dry De Los Pinos river bed, out of the Monkey Puzzles, i.e., Araucaria araucana forest, which is a novelty for a species of this genus because fanniids are believed to be basically inhabitants of forests, and are relatively rare in open landscapes and wetlands, although some exceptions have been found in species collected in Argentina (Domínguez, 2007). It would however be interesting to investigate if it really is related to this type of environment or if it is associated with the surrounding forests and was found outside the forest because of the use of baited traps.
Although F. adelaidae sp. nov. was not collected in association with a pig carcass, it was with the use of a baited trap, using putrescine, which could indicate some importance in forensic studies that should be explored as well as its medical relevance. This is the third species of the genus Fannia that is described as potentially important as PMI indicators, along with Fannia sanihue Dominguez & Aballay (2008) from the province of Mendoza and Fannia yunguensisQuiroga & Dominguez (2010) from the province of Jujuy, specifically in the yungas forests. These species, as well as F. adelaidae sp. nov. are interesting because they are from relatively unexplored, as far as the family Fanniidae is concerned, and remote regions of Argentina and belong phylogenetically to very different species groups within the family which is why they could be very valuable to aid in local forensic studies in the regions in which they were collected.
Acknowledgments
I am very grateful to Drs Pablo Mulieri, Luciano Patitucci, and Sofía Olea for collecting these specimens. I am especially thankful to Lic. Diego Miras for his help with the photographs. This study was funded by PICT-2019-0496 (MinCyT) and PIP 11220200100979CO (CONICET).
REFERENCES
Aballay, F.H., Murúa, A.F., Acosta, J.C. & Centeno, N.D. (2008) Primer registro de artropodofauna cadavérica en sustratos humanos y animales en San Juan, Argentina. Revista de la Sociedad Entomológica Argentina, 67, 157-163.
Aballay, F.H., Domínguez, M.C. & Fernández Campón, F. (2012) Adult Fanniidae associated to pig carcasses during the winter season in a semiarid environment: initial examination of their potential as complementary PMI indicators. Forensic Science International219(1-3), 284.e1–284.e4
Battán Horenstein, M., Linhares, A.X., Rosso, B., & García, M.D. (2010) Decomposition and dipteran succession in pig carrion in central Argentina: ecological aspects and their importance in forensic science. Medical and Veterinary Entomology, 24, 16-25.
Carvalho, L.M.L., Thyssen, P.J., Linhares, A.X., & Palhares, F.A.B. (2000) A checklist of arthropods associated with pig carrion and human corpses in southeastern Brazil. Memórias do Instituto Oswaldo Cruz, 95, 135-138.
Centeno, N., Maldonado, M., & Oliva, A. (2002) Seasonal patterns of arthropods occurring on sheltered and unsheltered pig carcasses in Buenos Aires Province (Argentina). Forensic Science International, 126, 63-70.
Crisci, J.V., Cigliano, M.M., Morrone, J.J., & Roig-Ruñent, S.A. (1991) Historical biogeography of southern South America. Systematic Zoology, 40, 152-171.
Domínguez, M.C. (2007) A taxonomic revision of the Southern South American species of the genus Fannia Robineau-Desvoidy (Diptera: Fanniidae). Papéis Avulsos de Zoología47, 289-347.
Domínguez, M.C., & Aballay F.H. (2008) A new species of the genus Fannia Robineau-Desvoidy (Diptera: Fanniidae) collected on pig carrion in Mendoza, Argentina. Annales Zoologici, 58, 819-824.
Domínguez, M. C., & Pont, AC. (2014) Fanniidae (Insecta: Diptera). Fauna of New Zealand, 71, 1-91.
Domínguez, M.C., & Roig-Juñent, S.A. (2008) A phylogeny of the family Fanniidae Schnabl (Insecta: Diptera: Calyptratae) based on adult morphological characters, with special reference to the austral species of the genus Fannia. Invertebrate Systematics, 22, 563-587.
Domínguez, M.C., & Roig-Juñent, S.A. (2011) Historical biogeographic analysis of the family Fanniidae Schnabl, with special reference to the austral species of the genus Fannia (Diptera: Fanniidae). Revista Chilena de Historia Natural, 84, 65-82.
Domínguez, M.C., & Roig-Juñent, S.A. (2017) Phylogeny and taxonomic revision of the genus Euryomma Stein (Diptera: Calyptratae: Fanniidae). Arthropod Systematics and Phylogeny, 75, 303-326.
Iannacone, J. (2003) Artropofauna de importancia forense en un cadáver de cerdo en el Callao, Perú. Revista Brasileira de Zoologia, 20, 85-90.
Mazza, S., & Oribe, H.R. (1939) Miasis urinaria por Fannia fusconotata (Rondani), en Formosa. Investigaciones sobre Dipteros Argentinos. I Miasis. Publicaciones Misión de Estudios de Patología Regional Argentina, Jujuy, 41, 66‑69.
McAlpine, J. F. (1981) Morfology and terminology Adults. Manual of Nearctic Diptera Agricultura Canada Monograph, n. 27, v.1 (ed. McAlpine, J.F., Peterson, B.V., Shewell, G.E.; Teskey., H. J., Vockeroth, J.R & Wood, D.M.), pp. 9‑63. Research Branch, Ottawa.
Morrone, J. J. (2015). Biogeographical regionalisation of the Andean region. Zootaxa, 3936, 207-236.
Oliva, A. (1997) Insectos de interés forense de Buenos Aires (Argentina). Primera lista ilustrada y datos bionòmicos. Revista del Museo de Ciencias Naturales, Bernardino Rivadavia, 7, 13‑59.
Perotti, A. (1998) Mosca sinantrópicas (Diptera: Muscidae y Fanniidae) asociadas a producciones avícolas del centro-sudeste bonaerense. Natura Neotropicalis, 29, 145‑154.
Pont, A.C., & Carvalho, C.J.B. (1994) Neotropical Fanniidae (Diptera): A key to the Fannia anthracina-group. Entomologist’s monthly Magazine, 130, 229‑238.
Quiroga, N.I., & Domínguez, M.C. (2010) A new species of the genus Fannia Robineau- Desvoidy (Diptera: Fanniidae) belonging to the canicularis species-group, collected on pig carrion in the Yungas of the province of Jujuy, Argentina. Studies on Neotropical Fauna and Environment, 45, 95-100.
Rozkošný, R., Frantisek, G., & Pont, A.C. (1997) The European Fanniidae (Diptera). Acta Scientiarum Naturalium Academiae Scientiarum Bohemicae, Brno, 31, 1‑80.
Souza, A.M., & Linhares, A.X. (1997) Diptera and Coleoptera of potential forensic importance in southeastern Brazil: relative abundance and seasonality. Medical and Veterinary Entomology, 11, 8-12.
Wolff, M., Uribe, A., Ortiz, A., & Duque, P. (2001) A preliminary study of forensic entomology in Medellín, Colombia. Forensic Science International, 120, 53-59.
Notas de autor
mcdomin@mendoza-conicet.gov.ar.