Artículos
Redescripción de Centruroides infamatus (Scorpiones: Buthidae) y descripción de una nueva especie de alacrán “rayado” del subgrupo “infamatus ” del centro- norte de México
Redescription of Centruroides infamatus (Scorpiones: Buthidae) and description of a new species of “striped” scorpion of the “infamatus” subgroup from north-central México
Redescripción de Centruroides infamatus (Scorpiones: Buthidae) y descripción de una nueva especie de alacrán “rayado” del subgrupo “infamatus ” del centro- norte de México
Revista de la Sociedad Entomológica Argentina, vol. 84, núm. 4, e0409, 2025
Sociedad Entomológica Argentina
Recepción: 11 Abril 2025
Aprobación: 30 Septiembre 2025
Resumen: Se redescribe Centruroides infamatus (C. L. Koch), con base en una población de la región de León, Guanajuato, en el centro de México ante la ausencia de una localidad tipo, ya que en la descripción original no se indica una localidad precisa para el ejemplar tipo. También se describe Centruroides rommeli sp. nov. con base en una población de Jalpa, en el municipio del mismo nombre al sur del estado de Zacatecas, México. Esta especie como parte de los alacranes “rayados” se ubica en el subgrupo “infamatus”. Se compara morfológicamente con C. suffusus (Pocock) y C. vittatus (Say), especies del mismo subgrupo con registro para el estado de Zacatecas. Se incluye también la redescripción de C. infamatus, debido a que la población de la nueva especie previamente se identificaba como C. infamatus. Centruroides rommeli sp. nov. es simpátrico con Mesomexovis spadix (Hoffmann) perteneciente a la familia Vaejovidae
Palabras clave: Complejo de especies, Gen mitocondrial RNAm 16S, Taxonomía, Zacatecas.
Abstract: Centruroides infamatus (C.L. Koch) is redescribed based on individuals obtained from a population from the León, Guanajuato region, in central Mexico. The species is redescribed because in the original description only the country is mentioned as the type locality with no further details. Also, Centruroides rommeli sp. nov. is described based on individuals obtained from a population of scorpions from Jalpa, municipality of Jalpa in the southern region of the state of Zacatecas, Mexico. This species is placed in the subgroup "infamatus" as part of the "striped" scorpions. Morphological comparisons are carried out with C. suffusus (Pocock) and C. vittatus (Say), both species that belong to the same subgroup talso recorded in Zacatecas. Additionally, is included the redescription of C. infamatus, because the population of the new species was previously identified with this specific name. Centruroides rommeli sp. nov. is sympatric with Mesomexovis spadix (Hoffmann), scorpions of the Vaejovidae family
Keywords: Mitochondrial gen RNAm 16S, Species complex, Taxonomy, Zacatecas.
INTRODUCCIÓN
Los alacranes son un grupo de artrópodos (Arachnida: Scorpiones) relativamente pequeño con aproximadamente 2,903 especies descritas a nivel mundial, incluidas en 24 familias (Rein, 2025). Por la importancia médica y biológica que tienen algunas de sus especies se le ha dedicado gran atención (Polis, 1990; Ponce-Saavedra & Francke 2004; Santibáñez-López et al., 2015; Ponce-Saavedra et al., 2016; González-Santillán & Possani, 2018; Ponce-Saavedra & Francke, 2019).
La familia Buthidae que con 94 géneros y 1296 especies a nivel mundial representa el 44,64 % del total conocido, en México, actualmente se encuentra representada por dos géneros. Por un lado, ChanekeFrancke, Teruel & Santibáñez-López, 2014, que forma parte de los denominados “microescorpiones” o “microbútidos” por su tamaño reducido y está formado por cuatro especies que son endémicas del sur de México (Kovařík et al., 2016). El otro género es Centruroides Marx 1890, que se encuentra distribuido desde el sur de los Estados Unidos de Norteamérica, hasta Centroamérica y el Caribe, así como la zona norte de Sudamérica (Fet & Lowe, 2000), y que en México está conformado por 60 especies (Francke & Ponce-Saavedra, s.f.).
Centruroides es el género responsable del problema de salud pública de alacranismo en México, ya que se registran un poco más de 319 mil accidentes por picadura de alacrán al año, de acuerdo con los datos del Sistema Único de Información para la Vigilancia Epidemiológica (SUIVE 2010-2024) que para 2024 reportó alrededor de 310 mil casos y en los primeros meses del 2025 se reportaron 212,590 casos según el boletín número 37 del SINAVE (2025); aunque según Francke (2019) podrían ser más de medio millón anual si se consideran los casos no registrados.
Las modificaciones ambientales provocadas por las actividades antrópicas benefician a los alacranes del género Centruroides cuyos hábitos de vida errantes, alta capacidad reproductiva y su corto ciclo de vida son favorables para tener éxito en los nuevos ambientes generados por las actividades antrópicas, las cuales les proporcionan diversos refugios que son utilizados como parte de su hábitat y facilita el encuentro de presas para su alimentación (Quijano-Ravell & Ponce-Saavedra, 2016a; Mullen & Sissom, 2019); sin embargo, se requiere de mayor esfuerzo de estudio en cuanto a diversidad de alacranes, distribución y factores asociados a la misma (Ponce-Saavedra & Francke, 2004; Quijano-Ravell et al., 2010; Ponce-Saavedra et al., 2015; Quijano-Ravell & Ponce-Saavedra, 2016a).
Centruroides infamatus (C.L. Koch, 1845) fue descrita con base en un ejemplar depositado en una colección particular en Nuremberg, Alemania y cuya procedencia exacta es desconocida, mencionando como localidad sólo a México. En 1932, Hoffmann decide asignar esta especie a poblaciones de diferentes estados de México, particularmente refiriendo a la región de Apatzingán, Michoacán; sin embargo, en la redescripción que hace de la especie toma poblaciones de otros estados y la morfometría de hembras y machos la presenta a partir de dos ejemplares de cada sexo procedentes de Jalapa, Veracruz, indicando una distribución muy amplia que explícitamente incluye al estado de Zacatecas y partes colindantes del estado de Durango. En estas poblaciones se incluía una amplia variación en tamaño, patrón de color, conteo de dientes pectíneos, forma y grado de desarrollo del tubérculo subaculear, forma y grado de desarrollo de las carenas, así como proporción entre longitud del carapacho y longitud del metasoma.
Hoffmann (1932) decidió que en esta amplia distribución geográfica se encontraban dos taxones subespecíficos: C. infamatus infamatus y C. infamatus ornatus, esta última descrita como especie válida por Pocock en 1902 para poblaciones de Jalisco y en consecuencia sinonimizada. La historia de esta especie continúa con Beutelspacher (2000) quien en el Catálogo de los alacranes de México acepta la sinonimia, pero considera insuficientes las diferencias indicadas por Hoffmann (1932) para asignarles el estatus taxonómico de subespecie y las propone como “formas”. En la mayoría de los trabajos posteriores se reconocen las poblaciones como subespecies (Fet & Lowe, 2000; Lourenço & Sissom, 2000; Ponce-Saavedra & Beutelspacher, 2001; De Armas & Martín-Frías, 2001; González-Santillán, 2001; De Armas et al., 2003;De Armas & Martín-Frías, 2008); aunque algunos de ellos ya apuntaban a la diferencia entre ellas (De Armas et al., 2003; Ponce-Saavedra & Moreno-Barajas, 2005), incluso se publica un trabajo que muestra diferencias a nivel molecular (Gantembein et al., 2001). Esta situación fue retomada y termina formalmente con el reconocimiento como especie válida y redescripción de C. ornatus Pocock, 1902 (Ponce-Saavedra et al., 2015) eliminando la categoría subespecífica para C. infamatus. En este trabajo fue necesario tomar la decisión sobre la o las poblaciones que pudieran representar a C. infamatus, lo que se hizo revisando la descripción original y adiciones posteriores, determinando que las poblaciones de la región de León, Guanajuato en el centro de México representarían adecuadamente a la especie y así se pudo tener un referente para comparaciones posteriores. La consecuencia fue que varias especies fueran incluidas en el “grupo infamatus” (Ponce-Saavedra & Francke, 2013a; Ponce-Saavedra et al., 2016). En el centro-occidente de México, tres especies de este grupo, con distribución en los estados de Jalisco, Guanajuato y Michoacán, son las más cercanas geográfica y morfológicamente a las poblaciones definidas como C. infamatus sensu stricto; por lo que se usaron para hacer la comparación y la redescripción de la especie. Estas especies son: C. ornatus Pocock, 1902, mencionada desde 1932 por Hoffmann como la más cercana y cuyas poblaciones se distribuyen al noroeste del estado de Michoacán, sur del estado de Guanajuato y sureste de Jalisco; Centruroides romeroiQuijano-Ravell, De Armas, Francke & Ponce-Saavedra, 2019, descrita de la sierra de Coalcomán en los límites de Michoacán y Jalisco y Centruroides ruanaQuijano-Ravell & Ponce-Saavedra, 2016b que se distribuye al noroeste del estado de Michoacán en la tierra caliente, en la localidad tipo de menor altitud sobre el nivel del mar (320 msnm) para estas especies.
Otras especies incluidas en el subgrupo “infamatus” se registran para el norte de México, las cuales tienen diferencias morfológicas importantes, las que, aunadas a la distribución geográfica (Fig. 1), características ecológicas y altitud sobre el nivel del mar, permiten separarles con facilidad (Ponce-Saavedra et al., 2016; González-Santillán et al., 2019; Ponce-Saavedra et al., 2022).
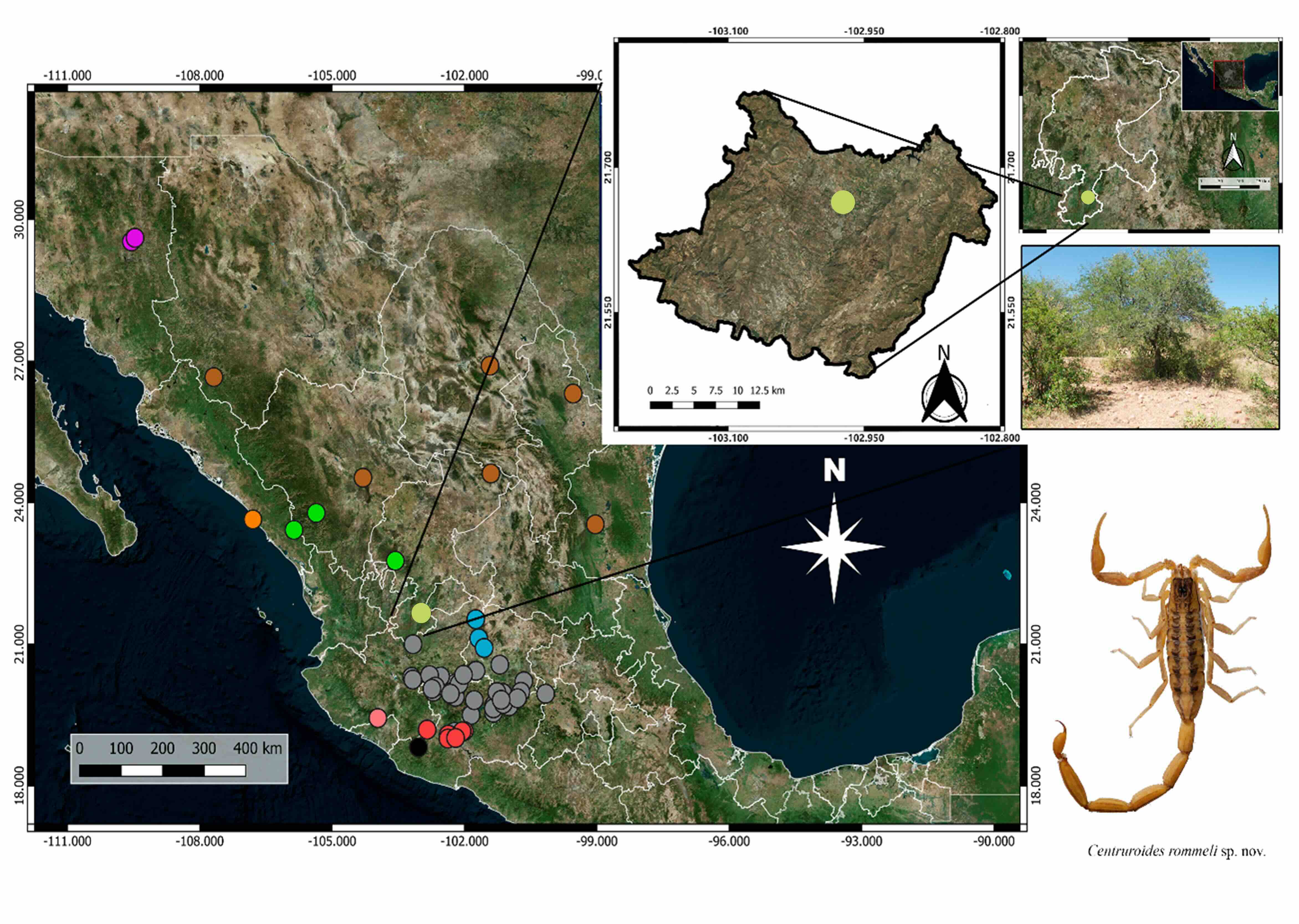
Figura 1
Mapa de la distribución de las especies del subgrupo infamatus
Referencias= : Centruroides infamatus, Centruroides ornatus, Centruroides vittatus, Centruroides suffusus, ***Centruroides ruana, Centruroides romeroi, Centruroides lauriadnae, Centruroides possani, Centruroides baldazoi, ***Centruroides rommeli sp. nov.
A pesar de que con la decisión tomada para hacer la comparación y validación de C. infamatus como especie, tanto en ese como en trabajos posteriores se consideran y/o mencionan algunos caracteres diagnósticos, sin embargo, en ninguno de ellos se incluyó una redescripción morfológica de la especie, lo que justifica la necesidad de disponer de ella con los criterios actuales y sin mezclar características de las otras poblaciones que Hoffmann en 1932 consideró, ni las que autores posteriores han definido como nuevas especies, anteriormente incluidas en el epíteto específico.
Esta redescripción ayudará a clarificar la situación taxonómica de otras poblaciones geográficamente dispersas en diferentes condiciones ambientales en varios estados del centro y norte de México, las cuales aún con la incorporación de las nuevas herramientas para la delimitación de especies, permanecen en duda sobre su identidad y que tentativamente están determinadas como C. infamatus y que son parte de este complejo de especies (Lourenço, 1980, 1981; De-Souza et al., 2009; Miranda-López et al., 2012; Ponce-Saavedra & Francke, 2004; Ponce-Saavedra & Moreno-Barajas, 2005; Lourenço & Da Silva, 2007; Ponce-Saavedra et al., 2015; Talal et al., 2015; Luna-Ramírez et al., 2017; Fet et al., 2018; Graham et al., 2019; Jochim et al., 2020; Ponce-Saavedra et al., 2022; Lourenço, 2023) y del que aún no se han separado todas las especies que le conforman (Ponce-Saavedra et al., 2022).
Con la descripción de Centruroides rommeli sp. nov. del estado de Zacatecas se incrementará a diez el número de especies del subgrupo, sumándose a las nueve especies ya descritas: Centruroides infamatus, C. vittatus, C. ornatus, C. suffusus, C. ruana, Centruroides. lauriadnaePonce-Saavedra & Francke, 2019; C. romeroi; Centruroides possaniiGonzález-Santillán, Galán-Sánchez & Valdez-Velázquez, 2019 y Centruroides baldazoiPonce-Saavedra, Linares-Guillén & Quijano-Ravell, 2022 (Fig. 2).
El estado de Zacatecas se ubica al centro-norte de México, limita al norte con Coahuila, al noreste con Nuevo León, al este con San Luis Potosí, al sur con Guanajuato, Jalisco y Aguascalientes, al suroeste con Nayarit y al oeste con Durango. En este estado se reconoce la presencia de tres especies del género Centruroides:
1) Centruroides suffusus (Hoffmann, 1932; Beutelspacher, 2000; De Armas & Martín-Frías, 2000; Fet & Lowe, 2000; Lourenço, 1997; De Armas et al., 2003; Teruel & Kovařík, 2010; Santibáñez-López et al., 2015; Ponce-Saavedra et al., 2016; Lucas & Meier, 2017; González-Santillán & Possani, 2018; Ponce-Saavedra & Francke, 2019; Martínez-Tejeda & Ponce-Saavedra, 2020)
2) Centruroides vittatus, (Hoffmann, 1932; Shelley & Sissom, 1995; Beutelspacher, 2000; De Armas & Martín-Frías, 2000; Fet & Lowe, 2000; De Armas et al., 2003; Teruel & Kovařík, 2010; Santibáñez-López et al., 2015; Ponce-Saavedra et al., 2016; Ponce-Saavedra & Francke, 2019)
3) Centruroides infamatus para el sur del estado (Hoffmann, 1932; Keegan 1980; Stahnke & Calos, 1977; Fet & Lowe, 2000; De Armas & Martín-Frías, 2008; Ponce-Saavedra & Francke, 2013a, 2013b; González-Santillán & Possani, 2018; Ponce-Saavedra et al., 2016; Lucas & Meier, 2017).
La población de Jalpa, ubicada en el sur del estado de Zacatecas, fue estudiada en el presente trabajo y representa la localidad tipo para C. rommeli sp. nov., determinándose así que para esta zona del país no hay registro válido para C. infamatus, dejando a esta especie hasta el momento solo en el estado de Guanajuato y otros registros en Aguascalientes, Michoacán y Nayarit, que se encuentran en proceso de descripción ya que corresponden a nuevas especies (Linares-Guillén, 2021).

Figura 2
Habitus de las especies que conforman el subgrupo infamatus
a. Centruroides infamatus. b. Centruroides ornatus. c. Centruroides vittatus. d. Centruroides suffusus. e. Centruroides ruana. f. Centruroides romeroi. g. Centruroides lauriadnae. h. Centruroides possani tomado de Gonzalez Santillán (2019). i. Centruroides baldazoi. j. Centruroides rommeli sp. nov. Escala= 5 mm.
Con estos aportes, la biodiversidad de escorpiones de México, que ya es el país de mayor riqueza específica en el mundo con 319 especies (Francke & Ponce-Saavedra, s.f.), se incrementará y además se genera información que facilitará delimitar otras especies y definir la situación taxonómica de todas aquellas poblaciones del subgrupo “infamatus” que aún permanecen en duda.
Con Centruroides rommeli sp. nov. se alcanzan 61especies para el género en México y 320 especies totales incluyendo las siete familias hasta ahora registradas, cantidad que seguramente será incrementada en el corto plazo con más especies de este complejo y otros, también del género Centruroides en los que se está trabajando.
MATERIALES Y MÉTODOS
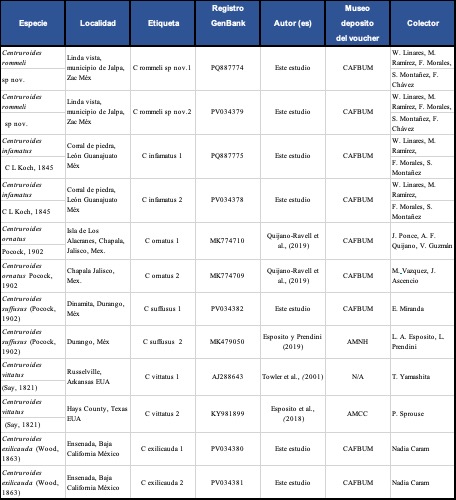
Para la redescripción de Centruroides infamatus se revisó la descripción original, las caracterizaciones morfológicas de Pocock (1902), Hoffmann (1932), Ponce-Saavedra et al., (1999); De Armas & Martín-Frías (2001); De Armas et al., (2003); De Armas & Martín-Frías (2008); Ponce-Saavedra & Francke (2013b); Ponce-Saavedra et al., (2016); además del trabajo de redescripción de Centruroides ornatus (Ponce-Saavedra et al., 2015). Posteriormente se hizo una caracterización morfológica y morfométrica con 31 ♂♂, 30 ♀♀ y 7 juveniles, además de comparar las secuencias de RNAm 16S obtenidas de cinco especies morfológicamente y/o geográficamente cercanas y pertenecientes al subgrupo “infamatus” sensu Ponce-Saavedra & Francke (2019) y de las cuales se cuenta con secuencias de ese fragmento de gen obtenidas de material recolectado por los autores y/o que se encuentran en GenBank (Tabla I).
En el caso de la nueva especie, para la caracterización morfológica, incluyendo el conteo de dientes pectíneos se revisaron 4 ♂♂, 14 ♀♀ adultas y 6 juveniles. El holotipo y la mayor parte de los paratipos (3 ♂♂, 4 ♀♀) quedarán depositados en la Colección Nacional de Arácnidos del Instituto de Biología de la Universidad Nacional Autónoma de México (CNAN-T01621, CNAN-T01622, CNAN-T01623, CNAN-T01624) mientras que un ♂ y el resto de los ejemplares en la colección de referencia del Laboratorio de Entomología “Biol. Sócrates Cisneros Paz” de la Facultad de Biología de la Universidad Michoacana de San Nicolás de Hidalgo (CAFBUM), con los datos CAFBUM-SB-620, CAFBUM-SB-621.
Históricamente se ha registrado en el estado de Zacatecas a Centruroides infamatus, por lo que la primera comparación de la población de la nueva especie en Jalpa se hizo con esta especie, para lo cual se usaron ejemplares de la CNAN (SC772) y de CAFBUM (SB-1727, 1728, 1737, 1738), provenientes de la región de León Guanajuato, tomados como referente de acuerdo con Ponce-Saavedra et al. (2015), ante la inexistencia de una localidad tipo.
Notas C inf_Leon GTO= Centruroides infamatus; C rom_JALPA, ZACATECAS= Centruroides rommeli sp. nov.; C inf_CORRALDEPIEDRA, LEONGTO= Centruroides infamatus; C suff_Vgro= Centruroides suffusus; C vitt= Centruroides vittatus; C lauriadnae= Centruroides lauriadnae; C sculpt= Centruroides sculpturatus; C exilicuada= Centruroides exilicauda
Para la comparación con otras especies del subgrupo “infamatus” y que tienen distribución en el norte del país, se consideró a Centruroides suffusus, especie de la que se usaron ejemplares de la Colección Nacional de Arácnidos CNAN (SC1226, SC1238) además de ejemplares de la Colección Aracnológica de la Facultad de Biología de la Universidad Michoacana de San Nicolás de Hidalgo CAFBUM (SB-1908, SB-1909); mientras que para comparar con Centruroides vittatus se usaron ejemplares de la CNAN (SC1331, SC1343) y de CAFBUM (SB-1910). En ambos casos también se revisaron las descripciones originales y adiciones posteriores (Say, 1821; Pocock, 1902; Hoffmann, 1932; Shelley & Sissom, 1995; Brown, 1996; Beutelspacher, 2000; De Armas & Martín Frías, 2000; De Armas et al., 2003; Teruel & Kovařík, 2010; Yamashita & Rhoads, 2013; Ponce-Saavedra et al., 2016).
Para la nomenclatura general se siguió a Stahnke (1970), dentición de los quelíceros según Vachon (1963), tricobotriotaxia de acuerdo con Vachon (1974), carenas del metasoma Francke (1977), carenas del pedipalpo Acosta et al. (2008), superficies del carapacho Prendini et al. (2003), hemiespermatóforo Monod et al. (2017); para los ojos laterales Loria & Prendini (2014), conteo y arreglo de setas de los esternitos de acuerdo con Ponce-Saavedra et al. (2022). Las medidas se hicieron acorde con Sissom et al. (1990).
Para las observaciones y la toma de medidas se utilizó un microscopio estereoscópico Carl Zeiss, Stemi DV4 y un vernier digital TTC con precisión de 0,01 mm. La toma de fotografías se hizo con una cámara réflex digital de 12,3 megapixeles marca Nikon modelo D90, con lente Micro-Nikkor 55 mm/3,5, un anillo de iluminación marca Amscope, modelo 144a. Posteriormente se almacenaron en Adobe Ligthroom Creative Cloud y finalmente, se exportaron a Adobe Photoshop Cs6® para la edición y correcciones de fondo. Para las estructuras más pequeñas se utilizó luz led y luz UV. Las fotografías se tomaron en el microscopio estereoscópico equipado con una cámara MU100 con lente FMA050 de AmScope de 10 megapixeles y posteriormente se exportaron a Adobe Photoshop Cs6® para mejorar las fotografías en exposición y contraste.
Para la elaboración de los mapas se usó QGIS versión 3,40,1 “Bratislava”, como insumos se usaron la carta de división política, estatal y municipal 1:250000 2023 de CONABIO y como capa base las imágenes satelitales del servidor Bing del Plugin QuickMapServices y una vista del hábitat.
Para la descripción de la nueva especie, el análisis morfométrico incluyó 32 ejemplares ♂♂ adultos, 3 de ellos correspondientes a la localidad de Jalpa, al sur del estado de Zacatecas; 8 de Centruroides infamatus (4 de la ciudad de León, Guanajuato y 4 de la localidad “Corral de piedra” en los suburbios de la misma ciudad); 5 de C. suffusus de dos localidades de Coahuila; 7 de C. vittatus de tres estados del país (Coahuila, Durango y Nuevo León); 3 de C. lauriadnae del estado de Sonora; 2 de C. sculpturatus de Guaymas, Sonora y 4 C. exilicauda de la península de Baja California. La matriz inicial de datos tuvo 69 variables (46 mediciones directas y 23 proporciones). Esta matriz fue depurada eliminando variables redundantes (valor de correlación múltiple de 0,8 o mayor), variables poco informativas (con eigenvalores muy bajos en los primeros tres componentes principales o aquellas con valores altos e interpretación equivalente en dos o tres de los primeros componentes en un Análisis de Componentes Principales), hasta llegar a una matriz poco ruidosa de 21 variables informativas (76.7% de variación explicada en los tres primeros componentes principales), formada por 10 mediciones directas y 11 proporciones (Tabla II).
Con esta matriz se corrió un Análisis Jerárquico de Agrupación para reflejar las tendencias de semejanza morfométrica entre las especies en estudio; un Análisis de Componentes Principales para evidenciar las variables que permiten separar a los ejemplares que representan a las especies y finalmente, un Análisis de Discriminantes Canónicos para evaluar la pertinencia del conjunto de variables utilizadas para conformar los grupos (especies) en análisis e identificar posibles variables diagnósticas para la nueva especie. Estos análisis se efectuaron utilizando el programa PAST v4,17 (Hammer et al., 2001).
Para el análisis molecular se hizo la extracción de ADN de los ejemplares a partir de tejido muscular de la patela del pedipalpo mediante el uso del protocolo de Fitz-Simmons (1997), amplificado mediante Reacción en Cadena de la Polimerasa (PCR por sus siglas en inglés) utilizando los “primers” previamente usados en estudios con alacranes (Gantenbein et al., 1999; Gantenbein et al., 2000; Gantenbein et al., 2001; Towler et al., 2001; Teruel et al., 2006; Ponce-Saavedra et al., 2009; Miranda-López, 2012; Quijano-Ravell et al., 2019). Para ello se utilizaron los primers con las siguientes secuencias: LRN 13398 5’-CGCCTGTTTATCAAAAACAT-3’ y LRJ 12887 5’-CCGGTTTGAACTCAGATCATGT-3’ bajo las condiciones sugeridas por Simon et al., (1994) para obtener secuencias del fragmento de gen mitocondrial RNAm 16S.
Una vez obtenidas las secuencias se alinearon mediante el método Clustal W usando el programa Mega XI (Tamura et al., 2021).
Se utilizaron 12 secuencias, las cuales tuvieron un tamaño final de 354 pares de bases y correspondieron a: dos secuencias de Centruroides suffusus de Durango, dos secuencias de Centruroides vittatus, una de Arkansas USA (Towler et al., 2001) y una más de Texas USA de Esposito et al., (2018); además de secuencias obtenidas por los autores que corresponden a dos secuencias de Centruroides ornatus, dos secuencias de ejemplares provenientes de localidades de la región de León, Guanajuato utilizados para la redescripción de Centruroides infamatus, además de dos secuencias de Centruroides rommeli sp. nov. de Jalpa, Zacatecas, todas ellas especies que son parte del subgrupo “infamatus”. También se incluyeron dos secuencias de Centruroides exilicauda de Baja California Sur, especie que se usó como grupo externo (Tabla I).
Las secuencias tuvieron un tamaño final de 354 pares de bases, de los cuales se observaron 197 sitios conservados, 157 sitios variables que contienen dos o más tipos de nucleótidos en esa posición, 142 sitios informativos para parsimonia y 19 sitios en el que la variación de un nucleótido es única para una secuencia (singleton).
Se obtuvo una tabla de distancias genéticas utilizando la p-distance como medida de divergencia, la cual indica la proporción respecto al número de diferencias, entre el total de nucleótidos comparados; la variación de la tasa entre los sitios se ajustó al modelo de distribución Gamma; obteniendo las distancias genéticas pareadas y la desviación estándar (Tabla III).
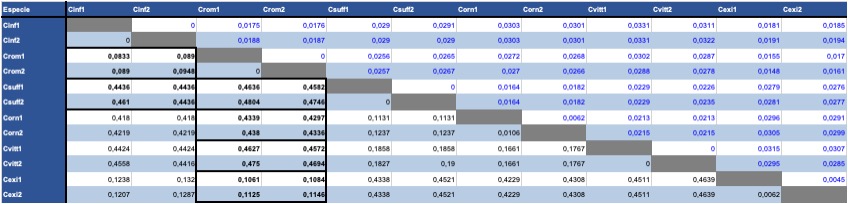
Notas Cinf= Centruroides infamatus; Csuff= Centruroides suffusus; Cvitt= Centruroides vittatus; Corn= Centruroides ornatus; Crom= Centruroides rommeli; Cexi= Centruroides exilicauda
Para recuperar las relaciones entre las especies, como una evidencia de su semejanza genética, se hizo una reconstrucción filogenética con tres métodos: Máxima verosimilitud, UPGMA y Máxima Parsimonia, evaluando con Bootstrapping la consistencia de los clados formados. Estos análisis se corrieron con el software MEGA XI (Tamura et al., 2021).
RESULTADOS
Análisis morfométrico
La matriz con menor número de variables informativas (21 variables), se utilizó en un análisis de agrupación con el método de Ward, para evaluar la semejanza entre las especies representadas, observando la cercanía morfométrica entre la población de Jalpa con las dos poblaciones de Centruroides infamatus del estado de Guanajuato, claramente separadas del grupo que contiene a todos los ejemplares representantes del resto de especies incluidas en el estudio (Fig. 3).
El Análisis de Componentes Principales mostró 76,7% de variación explicada, suficiente para considerarle una matriz informativa, confirmando la hipótesis de la cercanía morfométrica entre C. infamatus y C. rommeli sp. nov., así como la separación de las otras especies (Fig. 4). El Análisis Discriminante Canónico confirmó la utilidad del conjunto de variables utilizadas y mostró que la clasificación fue adecuada con una matriz de confusión con cero malas identificaciones y las pruebas estadísticas multivariantes significativas (p ≤ 0,05) (Tabla IV, Fig. 5). Es importante notar que en este análisis se probó la hipótesis emergente en los dos análisis anteriores, consistente en que las dos poblaciones de C. exilicauda pueden ser en realidad dos taxones distintos, lo que morfométricamente se corrobora y se plantea esta posibilidad para un trabajo posterior.
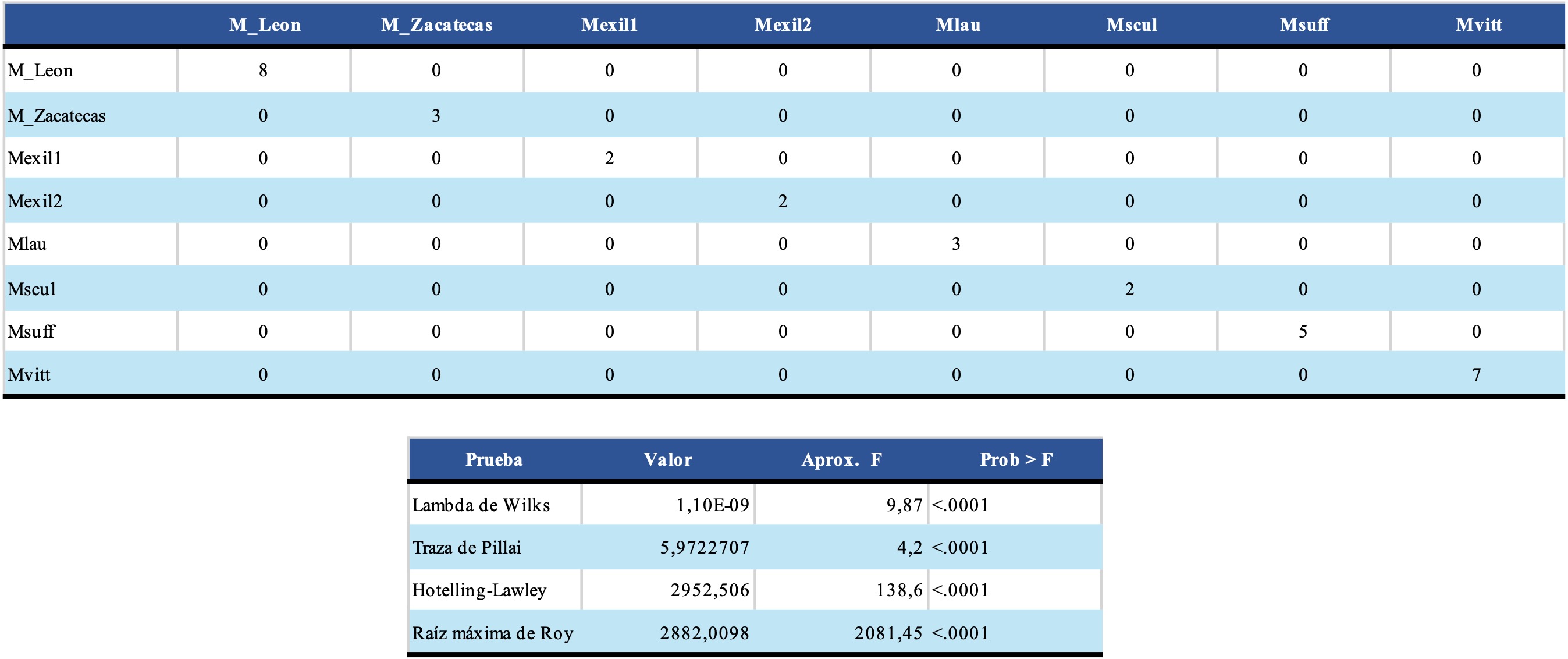
Notas M_Leon= Centruroides infamatus; M_Zacatecas= Centruroides rommeli sp. nov.; Mexil1= Centruroides exilicauda; Mvitt2= Centruroides aff. vittatus
Una vez reconocidas las variables que tienen mayor peso en la ordenación y para la discriminación de las especies, se probaron las variables individualmente mediante Análisis de Varianza de una vía (ANOVA) y se encontró que hay algunas variables que permiten separar (p<0,05) a C. infamatus y la nueva especie, con respecto de las otras especies utilizadas, tanto en el análisis de agrupamiento como en el ACP y el Análisis Discriminante. Estas variables son: la longitud de la vesícula (LVES); la longitud de la mano de la quela del pedipalpo (LMAN); las proporciones entre la longitud del segmento metasomal V con el segmento metasomal I (PLSMV/LSMI) y con la longitud de la vesícula (PLVSMV/LVES); la proporción entre la longitud de la mano y la longitud del segmento metasomal I (PLMAN/LSMI).
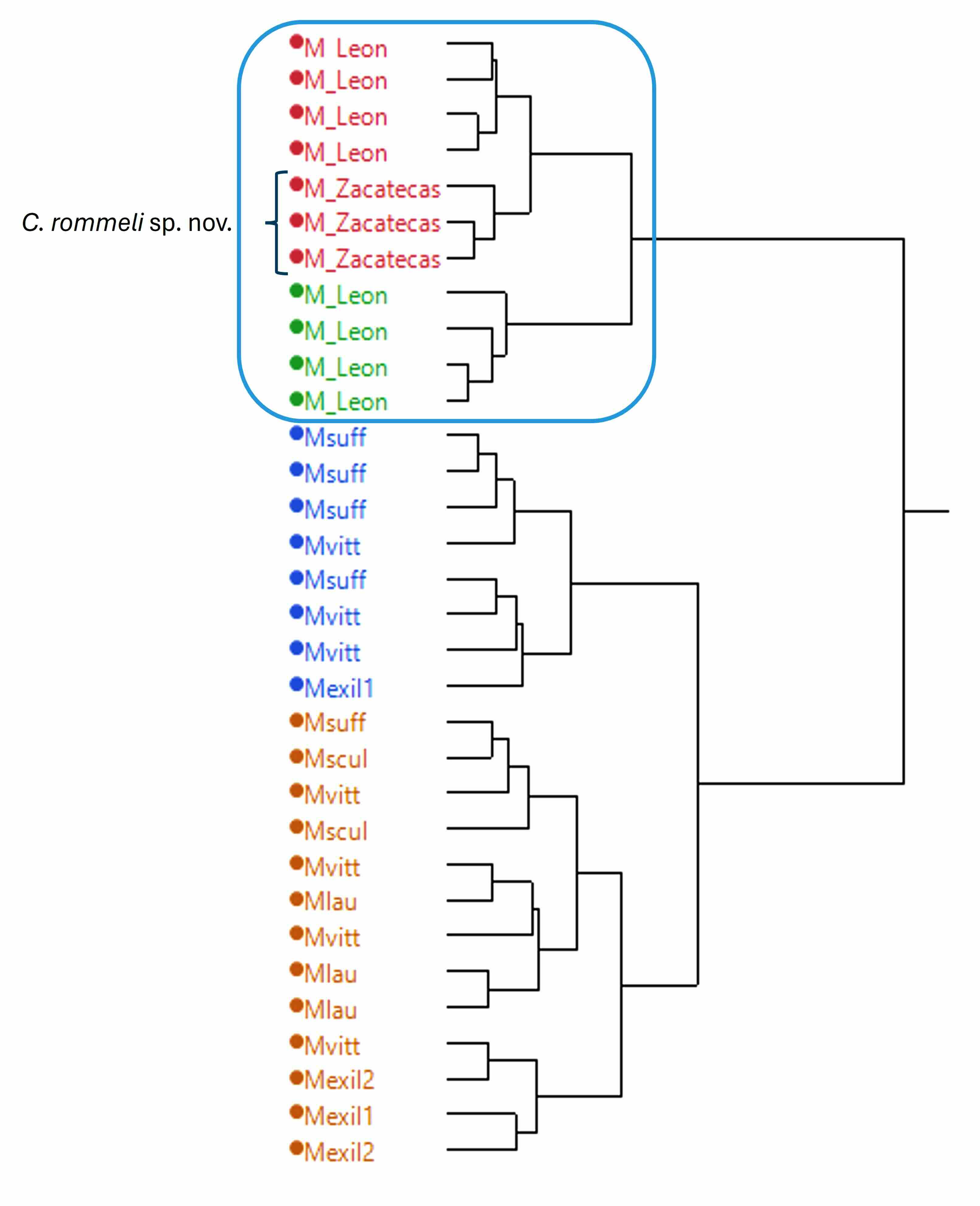
Figura 3
Dendrograma obtenido mediante el método de agrupación jerárquico de Ward con datos estandarizados
La longitud del cefalotórax con la longitud de la vesícula (LCEF/LVES) ofrecen separación en el componente principal 1(X), entre el grupo formado por C. rommeli sp.nov. y C. infamatus con respecto a las otras especies en análisis (variación explicada en dos componentes= 76,7%), lo que explica la separación en dos grandes grupos morfométricos observada en la figura 3.
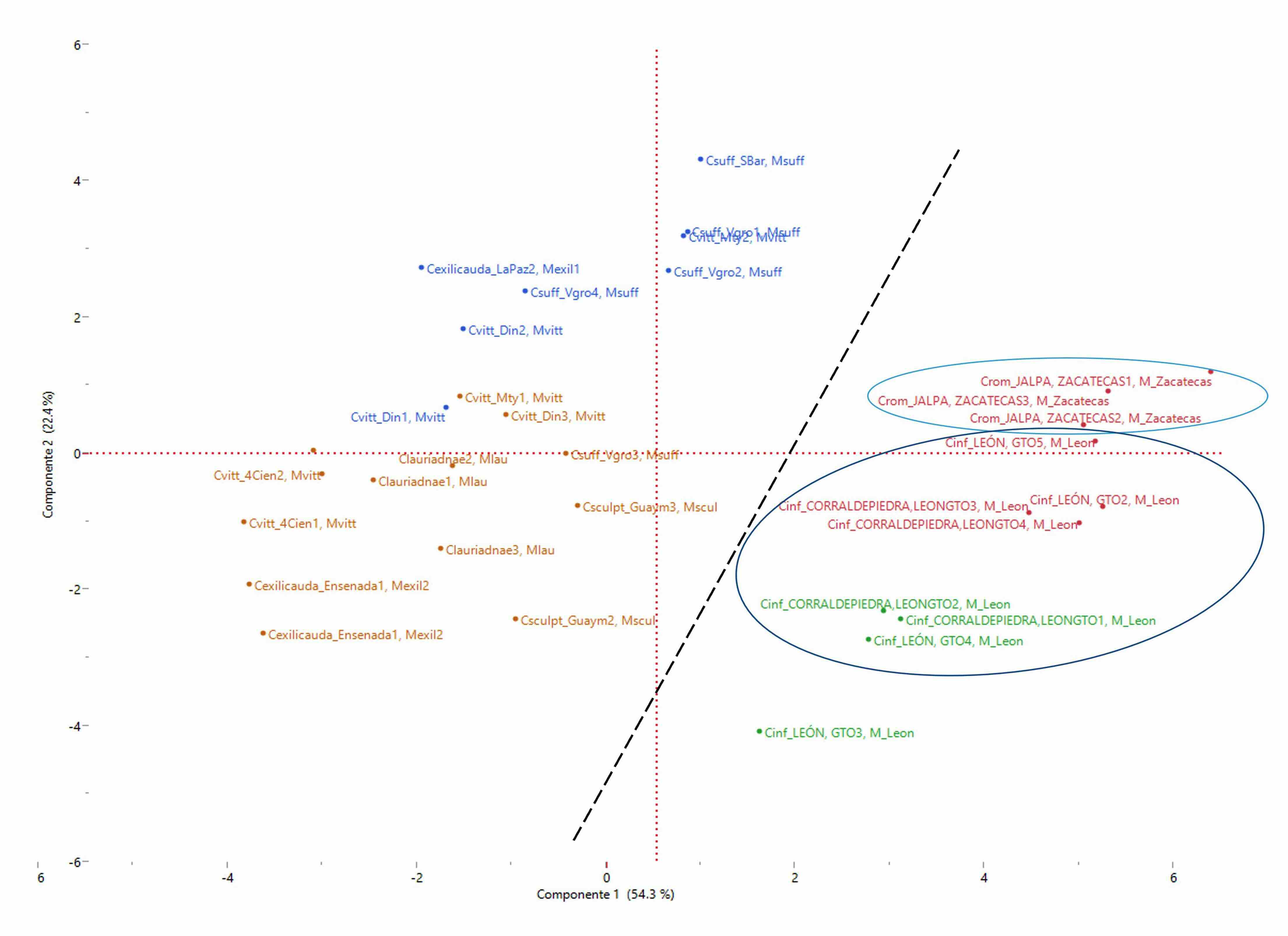
Figura 4
Gráfico obtenido con el Análisis de Componentes Principales
Nótese la separación en el componente principal 1(X), entre el grupo formado por Centruroides rommeli sp. nov. y Centruroides infamatus con respecto a las otras especies en análisis. Variación explicada en dos componentes= 76.7 %
La semejanza morfométrica entre la nueva especie y C. infamatus hace que en los gráficos resultantes de los tres análisis utilizados sus ejemplares aparezcan muy cercanos; sin embargo, se encontraron tres variables que individualmente tuvieron diferencia altamente significativa (p< 0,01) para poder separarles: La longitud de la vesícula (LVES) cuyo valor promedio ± Error estándar en mm para los ejemplares de C. infamatus de León fue de 5,42±0,20, por 6,22±0,12 en C. rommeli sp. nov.; en el mismo orden, la longitud de la mano (LMAN) 9,38±0,12 por 10,33±0,19 y la longitud del dedo fijo de la mano de la quela del pedipalpo (LDFIJO) 5,07±0,13 por 6,14±0,22.
Los resultados del Análisis Discriminante Canónico permitieron probar la hipótesis morfométrica sobre la diferencia multivariada entre la nueva especie y los ejemplares correspondientes a C. infamatus; así como la pertinencia de las variables utilizadas para separarles de las otras especies incluidas en el análisis (Tabla IV, Figura 5).
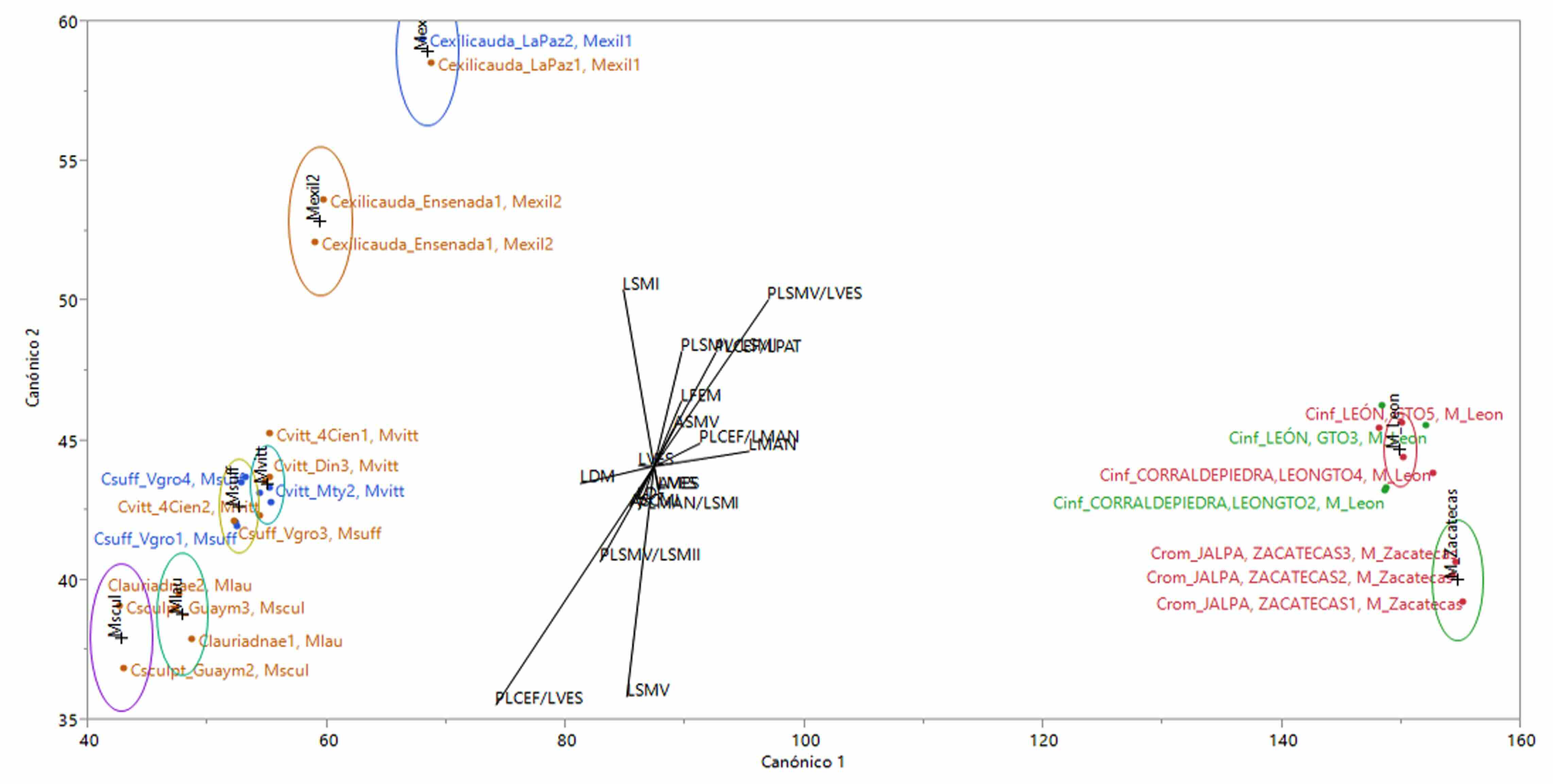
Figura 5
Gráfico obtenido con el Análisis Discriminante
Elipses de confiabilidad a 95%
Análisis molecular
Los resultados del análisis indican que las poblaciones de León y la localidad cercana a esta ciudad denominada “El corral de piedra” en el estado de Guanajuato, tienen una distancia genética que prácticamente les hace idénticos y representan la especie Centruroides infamatus C.L. Koch, 1845.
Las distancias genéticas de las secuencias de Centruroides rommeli sp. nov. con respecto a C. infamatus fueron entre 0,8 y 0,9 (≈0,01 de diferencia intragrupo). La comparación con otras especies del grupo mostró distancias genéticas de 0,44-0,46 con C. suffusus; 0,46-0,47 con C. vittatus y 0,42-0,43 con C. ornatus, mientras que con respecto al grupo externo C. exilicauda la distancia fue menor, alrededor de 0,11, lo que sugiere suficiente divergencia genética para la delimitación de la nueva especie.
La reconstrucción filogenética para representar las afinidades entre las especies mostró lo siguiente: Máxima verosimilitud (Fig. 6a) en el que C. infamatus muestra un valor Bootstrap de 100 % para la agrupación de las especies del subgrupo “infamatus” utilizadas en este estudio (C. suffusus, C. vittatus y C. ornatus); mientras que la nueva especie se recupera junto a las poblaciones de C. infamatus con 85% y finalmente las dos secuencias del grupo externo. Tanto la reconstrucción con el método de Máxima Parsimonia (Fig. 6b) como con UPGMA (Fig. 6c), se conserva el clado en el que se muestra la relación entre C. infamatus y C. rommeli sp. nov., aunque con valores ligeramente menores de soporte Bootstrapping; pero las secuencias por especie se recuperaron siempre juntas con 100 % de soporte.
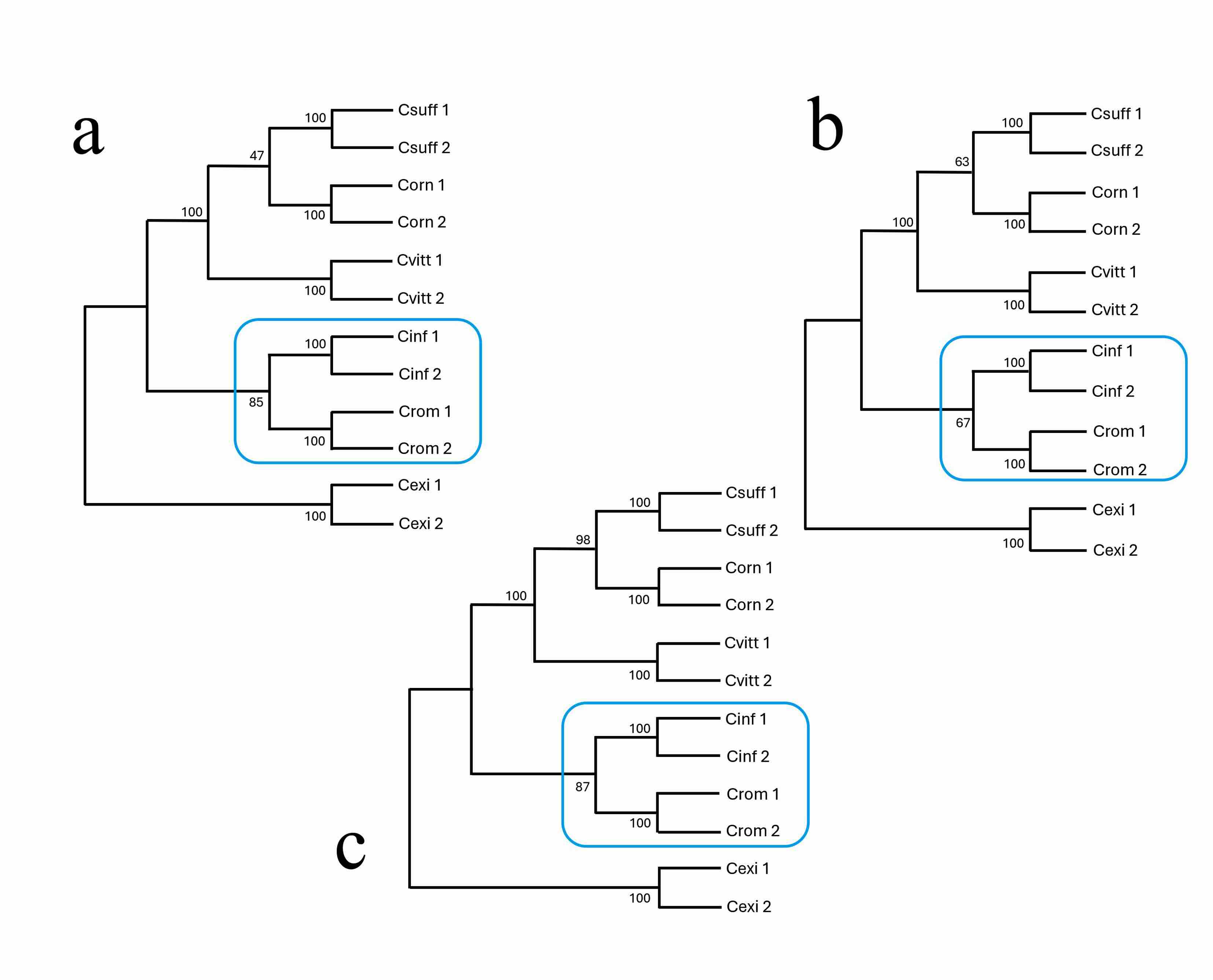
Figura 6.
Cladogramas obtenidos con diferentes métodos de reconstrucción filogenética como evidencia para delimitar Centruroides rommeli sp. nov. de Zacatecas, México. Métodos utilizados
a. Máxima verosimilitud. b. Máxima parsimonia. c. UPGMA
Diagnosis comparativa de Centruroides infamatus C.L. Koch
Antes de hacer la comparación morfológica de la nueva especie con Centruroides infamatus, se compara con especies que geográfica y morfológicamente pudieran confundirse con C. infamatus, una de ellas con distribución en Jalisco, Michoacán y Guanajuato (C. ornatus); otra que solo se conoce de la sierra de Coalcomán en los límites de Michoacán y Jalisco (C. romeroi) y una más en el valle de Apatzingán, Michoacán (C. ruana), por lo que la comparación se enfoca en estas especies. El resto de especies morfológicamente similares y que se ubican en el subgrupo “infamatus” sensu Ponce-Saavedra & Francke (2019), pueden separarse por su distribución geográfica más norteña o costera.
Centruroides infamatus son animales más grandes (51-63 mm) (Fig. 1a) que C. ornatus (40-50 mm) (Fig. 1b). La pigmentación del carapacho de Centruroides infamatus presenta áreas inmaculadas a los lados de los ojos medios, además del surco medio anterior, posterior y transverso (Fig. 8c), mientras que en C. ornatus la pigmentación del carapacho es más difusa e intensamente pigmentada, cubriendo la mayor parte del carapacho (Fig. 7a).
En el mesosoma de Centruroides infamatus las bandas obscuras son casi del mismo grosor que la banda mesal clara (Fig. 2a), mientras que en C. ornatus las bandas obscuras son más gruesas que la banda mesal clara (Fig. 2b), aunque esto puede variar entre poblaciones.
En machos el conteo de dientes pectíneos es más alto en Centruroides infamatus (21-24, moda= 23) (Fig. 10a) que en C. ornatus (19-22, moda= 21) (Fig. 7b).
La placa basal de los peines de Centruroides infamatus es subrectangular, casi dos veces más ancha que larga (proporción ancho/largo=1,9), con el borde anterior casi recto y una muesca media tenue, mientras que en C. ornatus la placa basal es más cuadrangular (proporción ancho/largo ≈1,7) y con el margen anterior ligeramente cóncavo. Entre hembras el conteo de dientes pectíneos de Centruroides infamatus es de 18-22 (moda=20), mientras que en C. ornatus es menor, de 18-21 (moda=19). La placa basal de los peines en hembras de Centruroides infamatus es subcuadrangular, sin muesca en el borde anterior y proporcionalmente más ancha (proporción ancho/largo=2,05), con el margen posterior redondeado, mientras que en C. ornatus la placa es similar, pero con margen posterior más recto. El tubérculo subaculear es cónico y reducido en Centruroides ornatus (Fig. 7d) y está ligeramente más desarrollado en C. infamatus.
Otras poblaciones revisadas para este trabajo se ubican al norte del estado de Michoacán en los municipios de Álvaro Obregón, Charo, Cuitzeo, Cuto de la Esperanza, Ecuandureo, Huandacareo, Indaparapeo, Tarímbaro, Zinapécuaro; al occidente Comanja, Jacona, Jiquilpan, La Piedad, Puruándiro, Sahuayo, Venustiano Carranza, Yurécuaro, Zamora y al centro en Acuitzio del Canje, Morelia, Tiripetío y Zacapu las que al compararlas se determinaron como Centruroides ornatus.
La segunda especie, Centruroides romeroi (Fig. 2f) descrita de la sierra de Coalcomán en los límites de Michoacán y Jalisco, se distingue de C. infamatus por ser la especie más pequeña hasta ahora conocida en el subgrupo (33-45 mm) contrastando con los 51 a 63 mm de C. infamatus; C. romeroi tiene una pigmentación más uniforme en el carapacho, incluyendo las áreas laterales a los ojos medios, que están ligeramente pigmentadas (Fig. 7e).
En machos el conteo de dientes pectíneos de Centruroides infamatus es de 21-25 (moda=23), mayor que los 20-22 (moda=21) de C. romeroi (Fig. 7f). El segmento metasomal V de Centruroides infamatus es más alargado (proporción largo/alto= 3.2), largo 6,85-9,07 y alto 2,25-2,8; comparado con C. romeroi que es tres veces más largo que alto (proporción ≈3,0) (Fig. 7g). Entre hembras el conteo de dientes pectíneos de Centruroides infamatus es de 18-22 (moda=20), mientras que en C. romeroi es de 19-21 (moda=20). El segmento metasomal V en hembras de Centruroides infamatus es más alto (proporción largo/alto=2,4) largo 6,43-7,1 y alto 2,67-2,96 (Fig. 10e), mientras que en C. romeroi es más delgado (proporción largo/alto ≈2,7).
La tercera especie, Centruroides ruana (Fig. 2e) se distribuye al noroeste del estado de Michoacán en la tierra caliente, en la localidad tipo con menor altitud sobre el nivel del mar (320 msnm). Se diferencia en que Centruroides. infamatus tiene el carapacho con una granulación densa, mientras que en C. ruana es más dispersa (Fig. 7i); metasoma de Centruroides infamatus con carenas moderadamente desarrolladas, el segmento metasomal V aproximadamente 2,5 veces más largo que alto (Fig. 10d) y con la vesícula ovalada y ligeramente alargada (proporción largo/alto= 1,98 mm) (Fig. 10f); mientras que en C. ruana las carenass del metasoma son fuertes, bien desarrolladas; el segmento metasomal V es más largo, casi tres veces más largo que alto (proporción largo/alto= 2,8) (Fig. 7k) y la vesícula (Fig. 7l) es ligeramente más redondeada (proporción largo/alto=1,9).

Figura 7
Algunas características de tres especies del subgrupo “infamatus”
Centruroidesornatus: a. Carapacho. b. Placa basal de los peines. c. Segmento metasomal V. d. Vesícula. Centruroides romeroi: e. Carapacho. f. Placa basal de los peines. g. Segmento metasomal V. h. Vesícula. Centruroides ruana: i. Carapacho. j. Placa basal de los peines. k. Segmento metasomal V. i. Vesícula. Escala= 1 mm
En machos, la placa basal de los peines en Centruroides infamatus es subrectangular casi dos veces más ancha que larga (proporción ancho/largo= 1,9), con el borde anterior casi recto y una muesca media tenue, y con conteo de dientes pectíneos de 21-25 (moda= 23) (Fig. 10a), mientras que C. ruana presenta la placa basal de los peines de forma subcuadrangular (proporción ancho/largo= 1,5), es ligeramente cóncava y lobulada, y posee un conteo de dientes pectíneos ligeramente más alto con 24-26 (moda= 25) (Fig. 7j). El segmento metasomal V de Centruroides. infamatus es aproximadamente 2,5 veces más largo que alto (Fig. 10d); mientras que en C. ruana es más largo, casi tres veces más largo que alto (proporción largo/alto= 2,8) (Fig. 7k). Entre hembras, el conteo de dientes pectineos en Centruroides infamatus es de 18-22 (moda= 20), mientras que en C. ruana es de 22-24 (moda= 23). La placa basal de los peines en hembras de Centruroides infamatus es más ancha (proporción ancho/largo= 2.05) (Fig. 10b), mientras que en C. ruana es más cuadrangular (proporción ancho/largo ≈1,8).
La distribución de esta especie incluye varias localidades en la parte norte de la Depresión del Balsas incluyendo áreas de transición en los límites de esta región fisiográfica (Fig. 1).
Redescripción de Centruroides infamatus C.L. Koch
Taxonomía
Centruroides infamatus C. L. Koch (1845)
Tityus infamatus C. L. Koch, 1845. Tab: CCCLXXII, Fig: 873. Pag: 46-48.
Holotipo: Sexo desconocido. Iconotipo: Hembra. Localidad tipo: México. Zoologisches Museum, Humboldt, Universitat, Berlin Germany
Sinónimos
Tityus infamatus Koch, 1850:91.
Centrurus infamatus Kraepelin, 1891: 121, 125-127; Kraepelin, 1893: 263-283; Kraepelin, 1894: 95; Lönngber, 1897a: 191; Lönngber, 1897b:184-185; Pocock, 1898:387; Kraepelin, 1899: 88-90; Kraepelin, 1901:270; Kraepelin, 1905: 198; Kraepelin, 1908: 184; Lampe, 1910: 24, Berland, 1913: 254; Penther, 1913: 243; Strand, 1916: 59; Lampe, 1917: 196, Ochoterena 1920: 223; Kopstein, 1921:143.
Centruroides infamatus infamatus Hoffmann, 1932: 328, 331-334, figuras 88-89; Hoffmann, 1937: 203-204; Hoffmann, 1938: 319; Díaz Nájera, 1964: 19- 27; Díaz Nájera, 1966: 111-116; Díaz Nájera, 1975: 3-33; Stahnke & Calos,1977:118; Keegan, 1980: 19, 71, 72; Francke, 1984: 98; Dehesa-Dávila, et al., 1989: 281-286; Dehesa-Dávila, et al., 1994: 1487-1492; Dehesa-Dávila, et al., 1996: 331-338; Fet & Lowe, 2000: 109,110; Lourenço & Sissom, 2000: 133; De Armas & Martín-Frías, 2001: 313, 315; González-Santillán, 2001:26; Ponce-Saavedra & Beutelspacher, 2001: 103; Towler et al., 2001: 158-163; Ponce-Saavedra & Moreno-Barajas, 2005; Chávez-Haro, 2011: 16; González-Sponga, 2011: 108; Possani et al., 2011: 149.
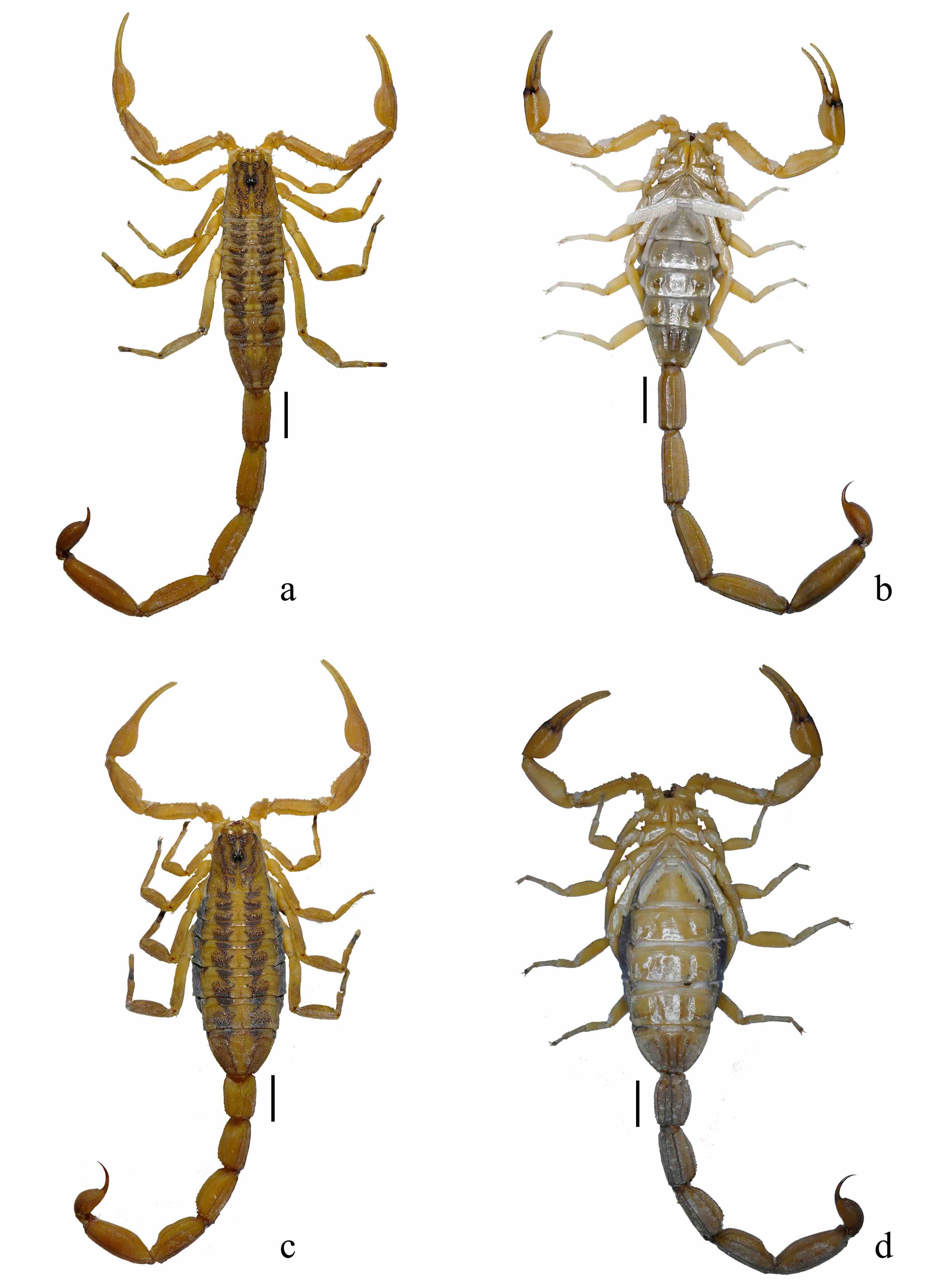
Figura 8
Habitus de Centruroides infamatus
a. macho vista dorsal. b. macho vista ventral. c. hembra vista dorsal. d. hembra vista ventral Escala= 5 mm
Distribución: México. Guanajuato. La región del municipio de León, incluyendo los ambientes urbanos de la ciudad; Municipio de Romita, Guanajuato.
Material Revisado: CAFBUM: 5♂, 7♀, León, Guanajuato, 21.1264N, -101.6688W, 1815 msnm, 8.VII.2001, Cols. n/a. 14♂, 2 juveniles, 13♀, 4 juveniles; Corral de piedra, Municipio de León, 21.0945N, -101.7433W, 1801 msnm 09.X.2020, Cols. W. Linares, S. Montañez, F. Morales, M. Ramírez. 2♂, 4♀; El Pochote, Municipio de Romita, 20.9222N, -101.5550W, 1750 msnm.VI.2016. Cols. E. Pavel Miranda. CNAN: 1♂; Colonia Jardines del Moral, Municipio de León, Guanajuato, 21.1454N, -101.6903W, 1827 msnm, sin fecha, Cols. n/a. CNAN:SC732. 9♂, 1 juvenil, 6♀; León, Guanajuato, 21.1264N, -101.6688W, 1815 msnm, 1.V.2004 Cols. P. Berea, CNAN:SC772. GUANAJUATO.
Alacranes de tamaño mediano para el género (43-60 mm); los machos (Fig. 8a, b) sexualmente maduros presentan dos tamaños: “pequeños” (47,13±1,5 mm, n= 9) y “grandes” entre 53,10±4,10 mm, n= 25).
Coloración: Color base amarillo a ocre, dorsalmente más obscuro. Carapacho con pigmentación difusa en la parte anterior, dos áreas inmaculadas en ambos lados de los ojos medios, igual que las áreas laterales, aunque éstas presentan unas pequeñas manchas irregulares (Fig. 9a), patrón que representa a las especies agrupadas en los alacranes “rayados” subgrupo “infamatus” sensu Ponce-Saavedra y Francke (2019). Pedipalpos ligeramente pigmentados sobre las carenas, obscuro en la articulación del dedo móvil y dentículos de ambos dedos. Patas ligeramente manchadas por el dorso, ventralmente inmaculadas.
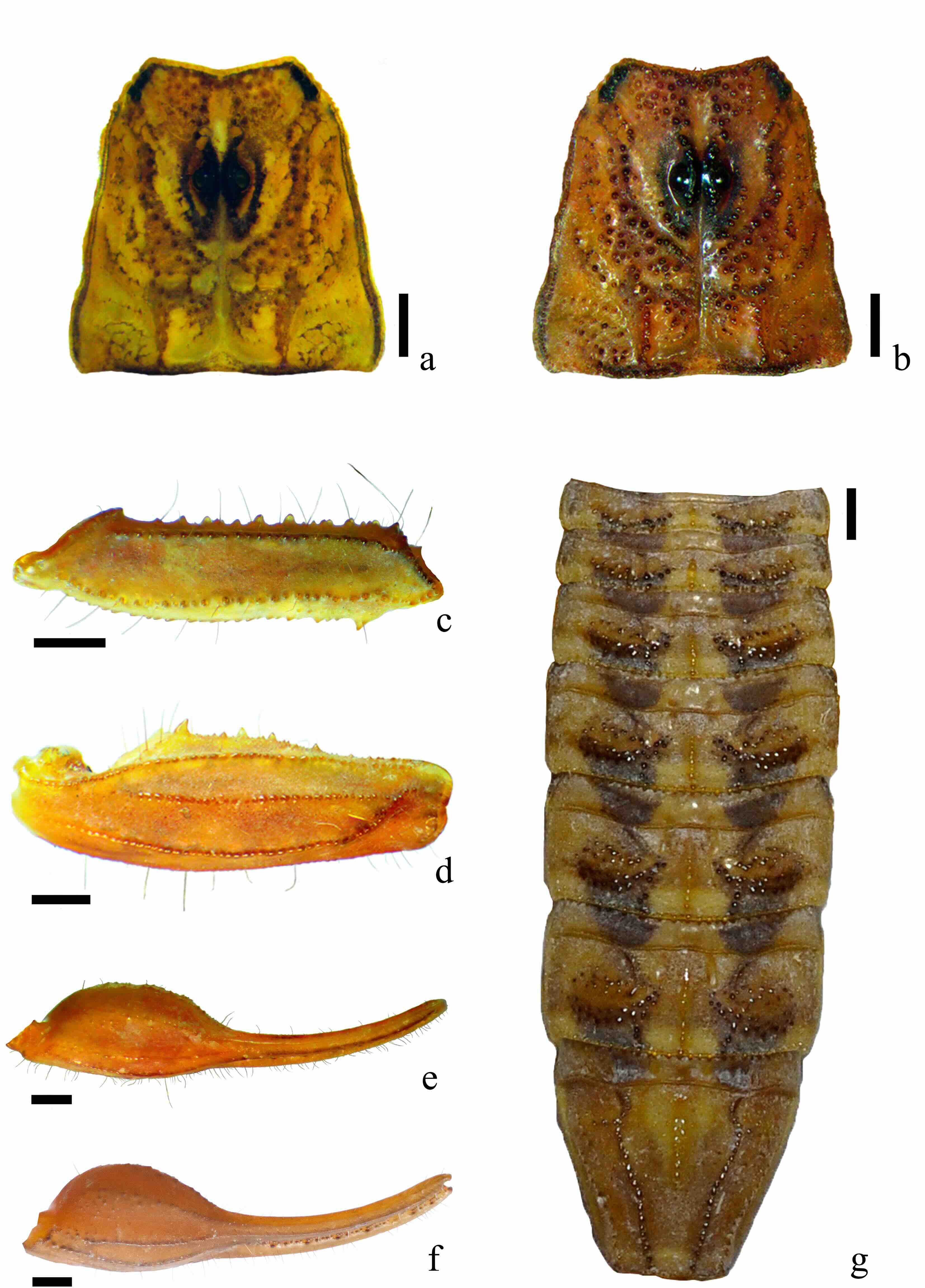
Figura 9
Características diagnósticas de Centruroides infamatus
a. Patrón de color de carapacho. b. Ornamentación de carapacho. c. Fémur vista dorsal. d. Patela vista dorsal. e. Mano de macho vista dorsal. f. Mano de hembra vista dorsal. g. Mesosoma, terguitos I-VII. Escala= 1 mm
Mesosoma (Fig. 9g) dorsalmente con manchas de pigmento negro sobre los terguitos, las cuales forman dos bandas longitudinales a los lados de una banda mesal inmaculada; bandas negras ligeramente más anchas (1,44 mm±0,1 n= 20) que la mesal (1,25 mm±0,14 n= 20); pigmento de las manchas intenso en preterguito y posterguito de las placas I-III unidas por pigmentación reticulada; IV-VI con una línea oblicua que une estas manchas; VII en su mayoría inmaculado, el pigmento se reduce a las carenas medias laterales. Ventralmente terguitos inmaculados y lustrosos; VII ligeramente más obscuro que el resto. Metasoma dorsalmente inmaculado; ventralmente hay pigmento obscuro a negro sobre las carenas mientras que los espacios intercarinales son esencialmente del color base. Vesícula ligeramente más obscura que el resto del telson, ventralmente con pigmentación difusa ligera a poco aparente; aguijón rojizo obscuro en su mitad distal, particularmente hacia la punta.
Superficie del carapacho con gránulos irregulares y dispersos, más abundantes en la parte media; carenas medias posteriores formadas por gránulos irregulares pigmentados (Fig. 9b). Pedipalpos en machos con carenas granulares moderadamente desarrolladas en fémur y patela (Fig. 9c, d), manos ovaladas, dos veces más largas que anchas (proporción largo/ancho= 1,94±0,21) (Tabla II,Fig. 9e); en machos carenas en su mayoría lisas; hembras con todas las carenas bien desarrolladas a fuertes, granulares y la mano de forma más globosa (proporción largo/ancho= 1,76±0.11) (Tabla II, Fig. 9f).
Placa basal de los peines de los machos rectangular, casi dos veces más ancha que larga (proporción ancho/largo (1,89±0,1) (Tabla II, Fig. 10a), con una muesca media en el borde anterior de la placa. Hembras con placa pectinal subcuadrangular, sin muesca en el borde anterior y proporcionalmente más ancha (proporción ancho/largo 2,05) y con el margen posterior redondeado (Fig. 10b); el conteo de dientes pectíneos en machos es 21-23 (moda= 22; n= 74), hembras 18-22 (moda= 20; n= 104).
Metasoma con quilla submediana ventral ligeramente crenulada en I-II y bien desarrolladas en III-IV; II y III con la lateral supramediana y lateral inframediana granulares a crenuladas poco desarrolladas, las dorsolaterales ligeramente dentadas (Fig. 10c); segmento metasomal V en machos con carenas muy débiles, aparentes solo por pigmento (Fig. 10d), en las hembras están presentes las carenas ventrolaterales, medias y las dorsolaterales apreciables al menos en la base (Fig. 10e), además de que es más alto (proporción largo/alto= 2,4; machos= 3,2).
Vesícula ovalada en machos, dos veces más larga que alta (proporción largo/alto= 2.03) (Fig. 10f), en las hembras más redondeada (proporción largo/alto= 1,6) (Fig. 10g). Tubérculo subaculear reducido a un gránulo o un pequeño diente cónico, tanto en machos como en hembras.
Hemiespermatóforo ensanchado (Fig. 11a) con la quilla basal bien esclerotizada; quilla interna fuerte desde la apófisis interna hasta la flexión del pie; reborde externo del pedicelo desarrollado; lámina interna del pedicelo lobulada, diente basal desarrollado en forma de gancho (Fig. 11b).
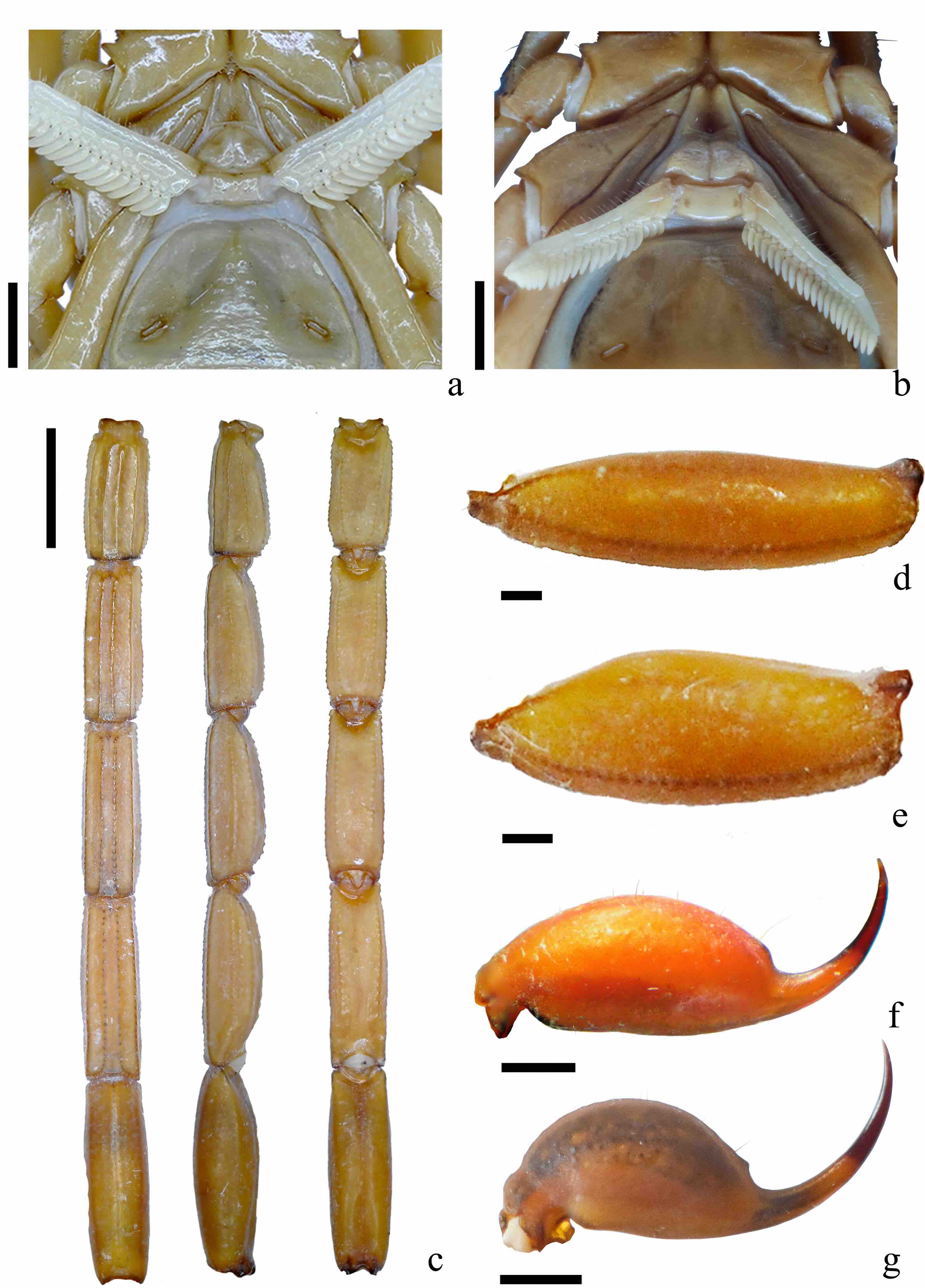
Figura 10
Características diagnósticas de Centruroides infamatus
a. Placa basal de los peines de macho. b. Placa basal de los peines de hembra. c. Metasoma. d. Segmento metasomal V de macho. e. Segmento metasomal V de hembra. f. Vesícula de macho. g. Vesícula de hembra. Escala= 1 mm
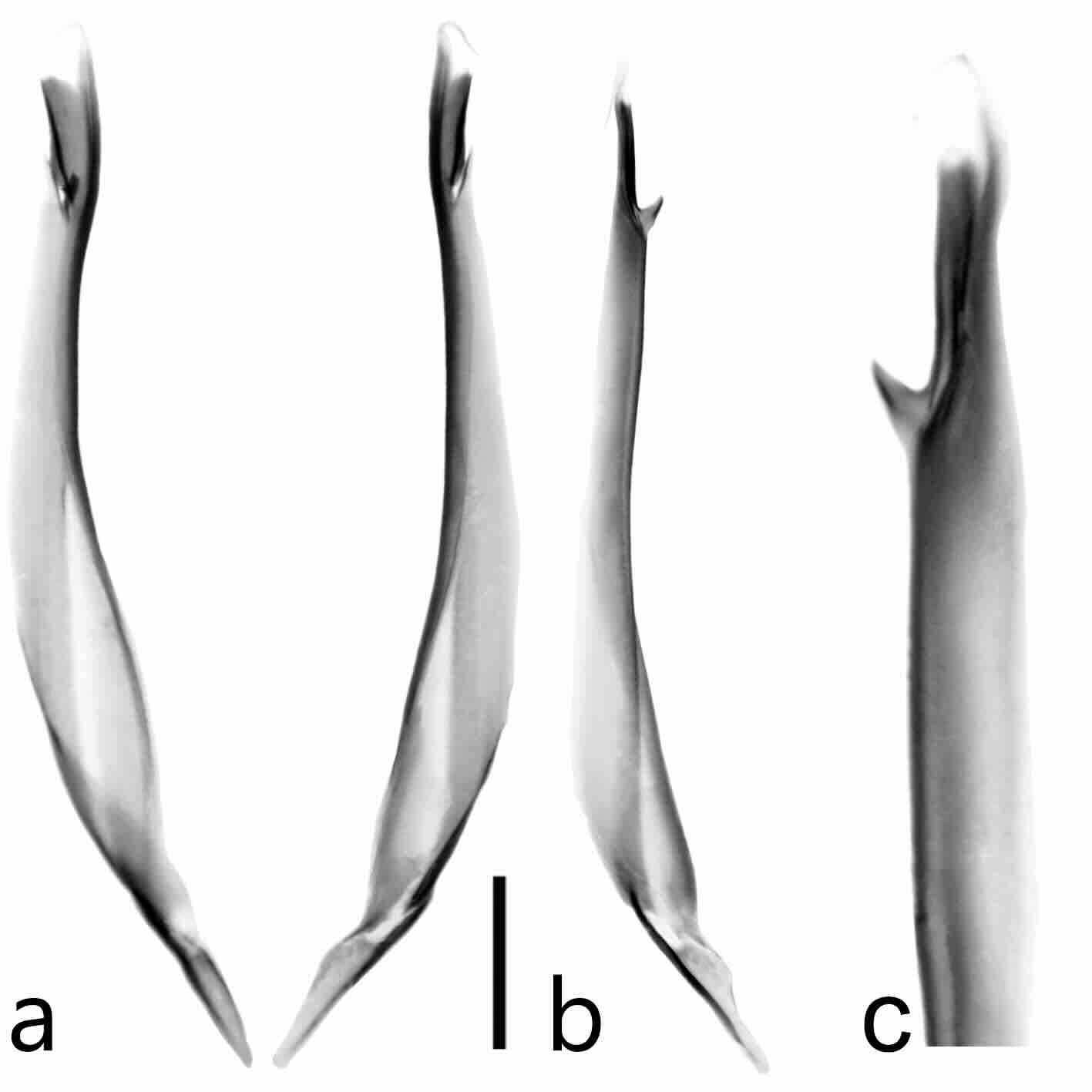
Figura 11
Hemiespermatóforo de Centruroides infamatus
a. Vista frontal. b. Vista lateral. c. Detalle de la región de los lóbulos. Escala= 1 mm
Centruroides rommeli sp. nov.Centruroides rommeli sp. nov.
Clasificación
Orden Scorpiones
Familia Buthidae C.L. Koch, 1837
Subfamilia Centruroidinae Kraus, 1955
Género Centruroides Marx, 1890
Material tipo: México: Zacatecas: Municipio de Jalpa: Holotipo ♂ (CNAN-T01621), 5 km del municipio de Jalpa 21.6639N, -102.9928W 1444msnm, 8.IV.2005, Colecta directa, Col: E. Miranda; 5 paratipos ♀♀ (CNAN-T01622), 5 km del municipio de Jalpa 21.6639N, -102.9928W 1444msnm, 8.IV.2005, Colecta directa, Col: E. Miranda 1 paratipo ♂ y 1 paratipo ♀ (CAFBUM-SB ); 3 paratipos ♂♂, Colonia Linda vista, Municipio de Jalpa, Zacatecas, 21.6601N, -102.9710W, 1427msnm, 4.X.2020, Colecta directa diurna Cols: W. Linares, M. Ramírez, F. Morales, S. Montañez, F. Chávez. (CNAN-T01623); 13 paratipos ♀♀ , Colonia Linda vista, Ubicada en el Municipio de Jalpa, Zacatecas, 21.6601N, -102.9710W, 1427msnm, 4.X.2020, Colecta directa diurna Cols: W. Linares, M. Ramírez, F. Morales, S. Montañez, F. Chávez (CNAN-T01624) y 1 paratipo♀ (CNAN SC773), Jalpa, Zacatecas, 1982, Cols: A. Durán.
Etimología: El nombre específico está dedicado al Dr. Carlos Rommel Beutelspacher Baigts, naturalista chiapaneco que incursionó en el estudio de las mariposas y los alacranes, dejando un importante legado científico en estas áreas de la entomología. Las orquídeas y la vegetación de Chiapas son ahora su pasión, además de la poesía y la pintura, actividades que ha cultivado con gran sensibilidad.
Diagnosis comparativa de Centruroides rommeli sp. nov.
La nueva especie difiere de las otras del género y estados vecinos del norte de México de la siguiente forma: con Centruroides infamatus (Fig. 2a) difieren en el tamaño, ya que son más pequeños (48-60 mm en machos, 48-53 mm en hembras); mientras que los ejemplares de la nueva especie miden entre 53 y 63 mm en machos y 40 a 53 mm en hembras.
La granulación del carapacho gruesa y dispersa en Centruroides infamatus (Fig. 9b), mientras que en la nueva especie es irregular y más abundante (Fig. 16c); la placa basal de los peines de la nueva especie es más ancha (proporción ancho/largo= 1,7) y el conteo de dientes pectíneos es más alto (23-25) (Fig. 16d), mientras que en C. infamatus la placa basal es 1,9 veces más ancha que larga y el conteo de dientes pectíneos es menor (21-23) (Fig. 10a); los esternitos de C. infamatus presentan menor cantidad de setas (III: 30, IV: 18, V: 30, VI: 14, VII: 8) (Fig. 12a), mientras que en la nueva especie hay mayor número de setas en todos los esternitos (III: 32, IV: 22, V: 38, VI: 12, VII: 10), (Fig. 12b).
Centruroides rommeli sp. nov. tiene apariencia de “cola delgada”, ya que los segmentos metasomales son cuatro veces más largos que altos y en el segmento metasomal V las carenas con gránulos son muy débiles y dispersos (Fig. 16a), mientras que en C. infamatus son apenas tres veces más largos que altos y en el segmento metasomal V las carenas son obsoletas, solo evidenciadas por pigmento (Fig. 10c)
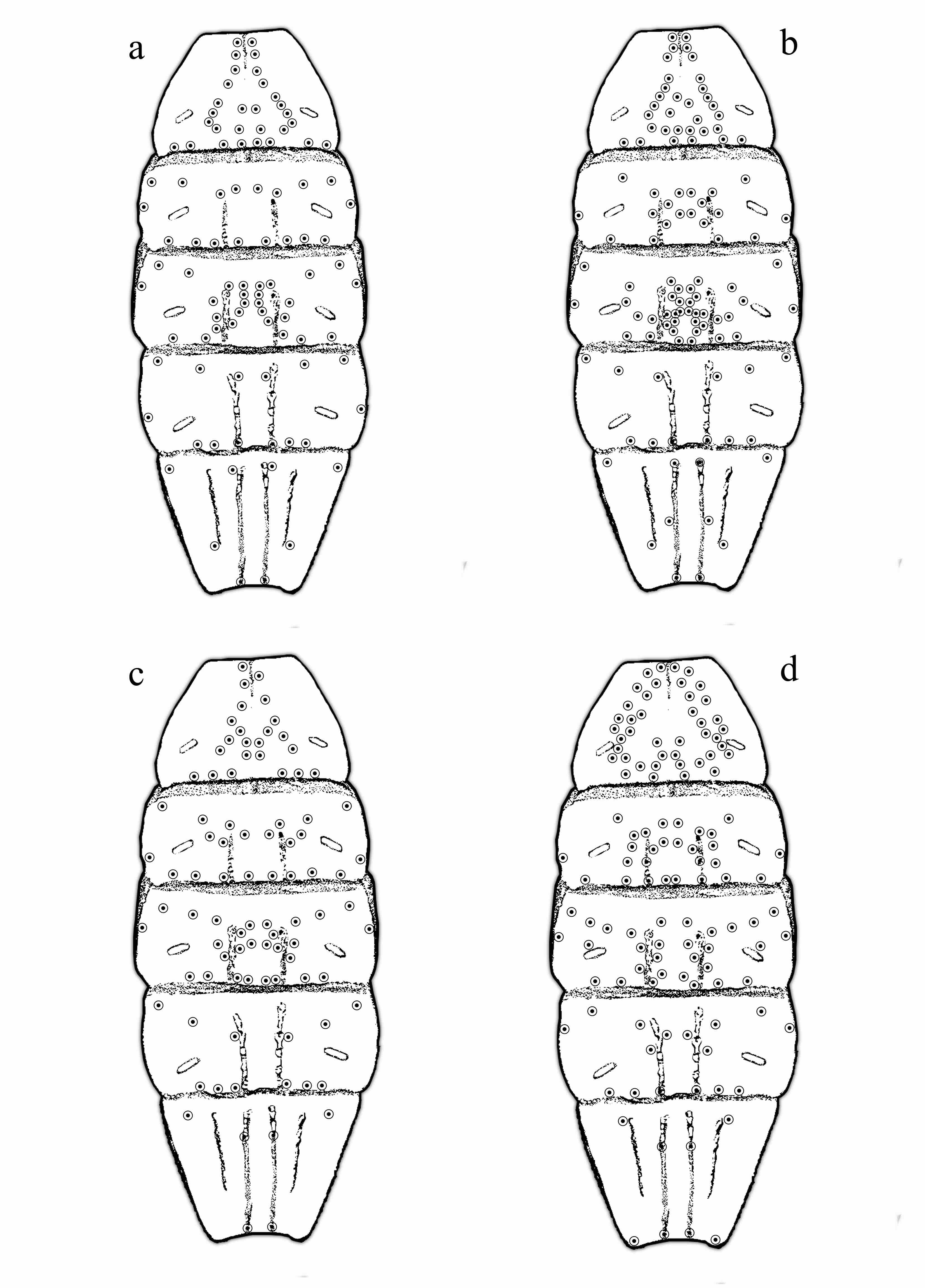
Figura 12
Patrón de setas en esternitos
a. Centruroides infamatus. b. Centruroides rommeli sp. nov. c. Centruroides vittatus. d. Centruroides suffusus
Con Centruroides vittatus (Fig. 2c) que se distribuye al norte del estado y se extiende hacia Coahuila, Chihuahua y Nuevo León, la nueva especie difiere por presentar el surco medio anterior del carapacho inmaculado y con pigmentación difusa (Fig. 13a); mientras que C. vittatus presenta una pigmentación intensa en la parte anterior del carapacho la cual se extiende hacia los ojos laterales y posteriormente en diagonal hacia los lados del surco medio posterior formando un triángulo (Fig. 13e)
En Centruroides rommeli sp. nov, la placa basal de los peines es subrectangular (proporción ancho/largo= 1.7), borde anterior convexo y el posterior recto, el conteo pectinal es menor (22-25) mientras que en Centruroides vittatus la placa basal de los peines es subcuadrangular, (proporción ancho/largo= 1.5), borde anterior recto y el posterior es curvado; el conteo de dientes pectíneos es claramente más alto (27-30). La nueva especie presenta más setas en los esternitos III: 32, V: 38 y VII: 10 (Fig. 12b) con respecto a C. vittatus que tiene III: 23, V: 28 y VII: 6 (Fig. 12c)
Centruroides rommeli sp. nov. presenta la vesícula es ovalada 2,1 veces más larga que alta y el tubérculo subaculear es cónico y reducido (Fig. 13b), mientras que en C. vitattus es 2,5 veces más larga que alta y el tubérculo subacuelar es cónico y más desarrollado, con orientación hacia la punta del aguijón (Fig. 13f)
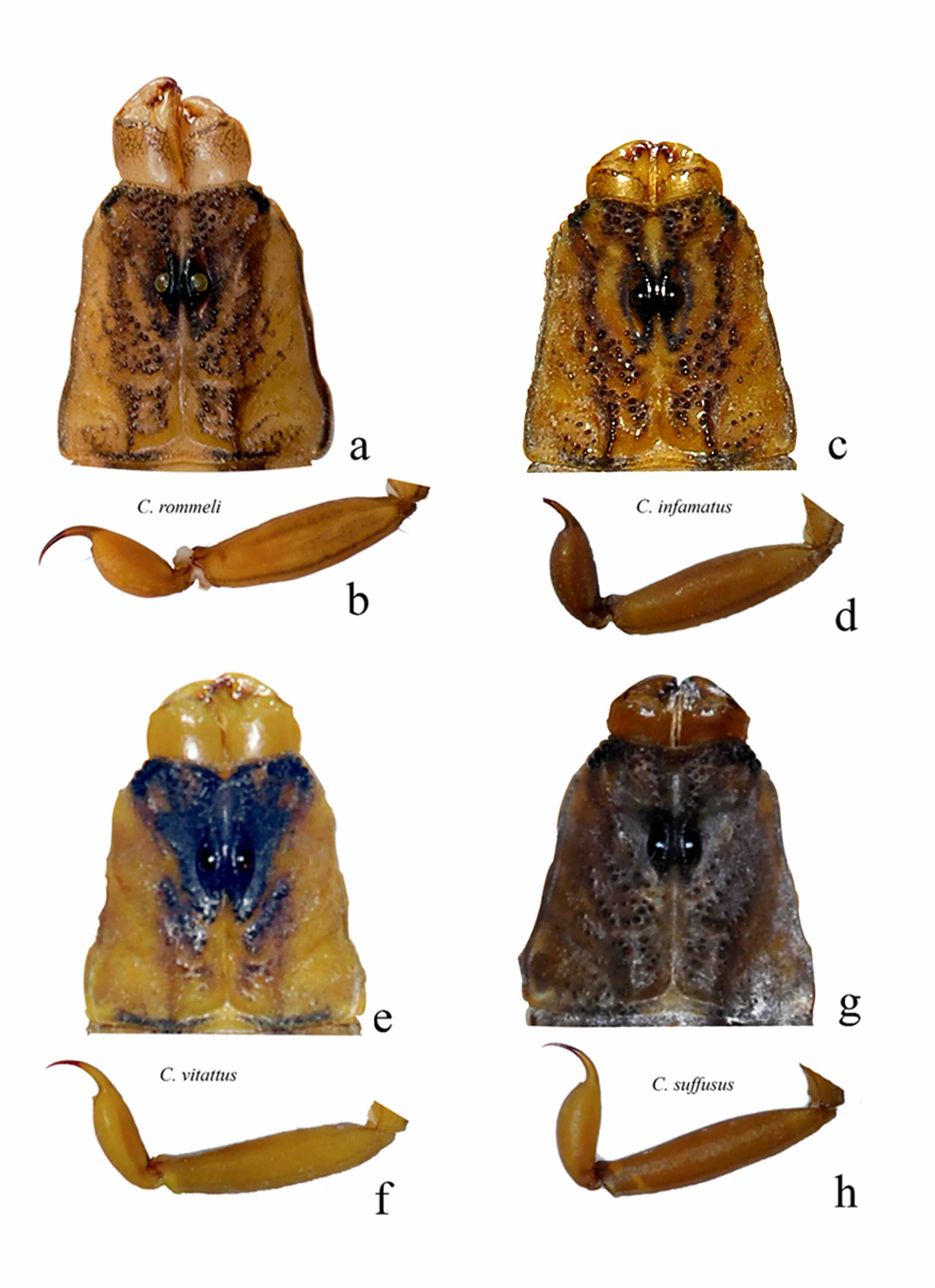
Figura 13
Algunas características comparativas de Centruroides rommeli con tres especies del subgrupo “infamatus”, dos que habitan en el norte de México
a-b. Carapacho y Segmento metasomal V + vesícula de Centruroides rommeli. c-d. Carapacho y Segmento metasomal V + vesícula de Centruroides infamatus. e-f. Carapacho y Segmento metasomal V + vesícula de Centruroides vittatus. g-h. Carapacho y Segmento metasomal V + vesícula de Centruroides suffusus
Finalmente, Centruroides suffusus (Fig. 2d), se diferencia de la nueva especie por ser de apariencia esbelta y alargada (proporción largo metasoma/largo mesosoma ≈2,7) y de mucho mayor tamaño alcanzando hasta 90 mm en machos según Hoffmann (1932) contrastando con los 50-60 mm y apariencia robusta (proporción largo de mesosoma/largo de metasoma 2,08) de la nueva especie, la cual también puede diferenciarse por la placa basal de los peines que presenta el borde anterior convexo y el posterior recto, a diferencia de la placa en C. suffusus cuyo borde anterior es recto y el posterior ligeramente curvado; también se aprecian diferencias importantes en el arreglo y número de setas de los esternitos, ya que la nueva especie presenta menor número de setas en los esternitos: III: 32, IV: 22, VI: 12 y mayor en V: 38 y VII: 10 (Fig. 12b), mientras que C. suffusus es más setoso en III: 42, IV: 24, V: 30 y menos en VI: 16, VII: 8 (Fig. 12d), sin considerar las microsetas dispersas que no presentan un patrón

Figura 14
Habitus de Centruroides rommelisp. nov
a. Macho vista dorsal. b. Macho vista ventral. c. Hembra vista dorsal. d. Hembra vista ventral. Escala= 5 mm
Además en Centruroides rommeli sp. nov. el segmento metasomal V es más corto y más alto (proporción largo/alto= 3,8) (Fig. 13b), mientras que en C. suffusus el segmento es delgado y alargado, ≈4,5 veces más largo que alto; la nueva especie presenta la vesícula redondeada (proporción largo/alto= 2,1) y el tubérculo subaculear es cónico y reducido, mientras que en C. suffusus la vesícula es ligeramente más ovalada, (proporción largo/alto= 2,3) y el tubérculo subaculear más desarrollado y más cercano a la base del aguijón (Fig. 13h)
Descripción del holotipo macho
Coloración: Dorsalmente amarillo marrón, patas y superficie ventral ligeramente más claras, segmento metasomal V y vesícula ligeramente más obscuros que el resto del cuerpo (Fig. 14a, b).
Carapacho (Fig. 15a), con pigmentación difusa anteriormente, ojo lateral posterior con pigmentación negra que se extiende ligeramente hacia atrás; pigmentación de la superficie difusa en el área posterior a los ojos laterales tornándose más intensa conforme se extiende y angostándose hacia los lados de los ojos medios hasta que se interrumpe por los surcos medios transversos que son inmaculados; posterior a este, la pigmentación continúa y se extiende hasta el borde posterior del carapacho al menos en el área prolateral, mientras que en el área media se interrumpe por el surco posterior transverso que es inmaculado, en esta zona se aprecian dos áreas redondas sin pigmento; parte posterior de los ojos laterales con dos áreas deprimidas con pequeñas manchas irregulares; dos áreas inmaculadas a los lados de los ojos medios, un par de manchas oblicuas se extienden desde el ojo posterior lateral hacia el surco medio posterior formando una línea, sin llegar a unirse en el surco medio posterior; las áreas laterales del carapacho se observan casi inmaculadas, con pequeñas machas de pigmento irregular.
Ojos laterales negros (Fig. 15b) rodeados por pigmentación obscura que se extiende hacia el ocelo menor anterodorsal (ADMi), presentan arreglo tipo 4A (Loria & Prendini, 2014); tubérculo ocular negro, surco medio posterior inmaculado solo en la parte anterior. Surco medio anterior con pigmentación difusa, similar al surco medio posterior en las dos terceras partes distales, áreas posterolaterales con pigmentación reticulada fina, similar en posterguito I. Márgenes laterales con pigmentación fina y obscura que coalece hacia el borde posterior del carapacho.
Quelíceros con pigmentación reticulada anteroposteriormente, área central y dedos inmaculada, excepto la punta de los dientes que son rojizos (Fig. 15c). Trocánter, fémur y patela de Pedipalpos, dorsalmente con manchas tenues irregulares, articulación del dedo móvil e hileras de los dientecillos con pigmentación marrón rojiza (Fig. 15 d-f).
Preterguitos I-II del Mesosoma (Fig. 15g) con una mancha de pigmento obscuro subrectangular, en terguitos III-VI esta mancha es trapezoidal y pigmentación reticulada la que llega hasta el borde posterior del posterguito. Terguitos III-VI con una línea de pigmento oblicua, que inicia desde la mancha del preterguito y se une con la pigmentación del posterguito. Terguito VII no comparte este patrón, ya que se observa una mancha de pigmento reticulado en el preterguito; además del pigmento en carenas dorsales hay pigmentación irregular entre éstas y la lateral supramediana. Esternitos III-VI sin un patrón de coloración, VII con tenue pigmentación entre carenas submedianas y laterales.

Figura 15
Características diagnósticas de Centruroides rommeli
a. Patrón de color de carapacho. b. Ojos laterales. c. Quelícero. d. Fémur vista dorsal. e. Patela vista dorsal. f. Mano de macho vista dorsal. g. Mesosoma, terguitos. Escala= 1 mm
Patas ventralmente de coloración más clara que el resto del cuerpo; dorsal y dorsolateralmente ligeramente pigmentadas, pata I con pigmentación más tenue. Metasoma (Fig. 16a) dorsalmente con pigmentación muy tenue en segmentos I-V, pigmentación infuscada entre las carenas ventrales y más tenue entre las carenas ventrolaterales, todas las carenas con pigmento; segmento V con pigmento sobre carenas ventrolaterales y ventromedianas. Vesícula ligeramente más obscura que el resto del metasoma, aculeus rojizo, más obscuro hacia la punta (Fig. 16b).
Carapacho con borde anterior ligeramente convexo, granulación irregular y dispersa, gránulos gruesos en la parte anterior que se extienden lateralmente hacia los ojos laterales y posteriormente hacia las carenas superciliares, las cuales son gruesas y crenuladas, granulares en la parte posterior donde se bifurca en dos áreas de gránulos finos que se van engrosando hacia la parte distal de los ojos medios (Fig. 16c). Gránulos gruesos abundantes en la zona media junto a los ojos medios y hacia las carenas medias posteriores. Carenas medias con granulación gruesa a fina.
Ojos medios redondos y separados (0,47 mm); tres ojos laterales redondos subiguales, el anterior ligeramente más pequeño, ocelos accesorios en arreglo tipo 4A (Loria & Prendini, 2014). Los bordes laterales del carapacho finamente granulados, borde posterior con gránulos grandes y pequeños. Las áreas inmaculadas presentan una granulación muy fina.
Quelíceros (Fig. 15c) con la superficie finamente granulada, carena transversal granular, con tres macrosetas distales y tres basales, área retrolateral y ventral cubiertas con setas largas. Dedo fijo con el diente distal grande y agudo, subdistal mediano y romo, medio y basal, más grandes, con apariencia bicúspide. Dedo móvil con el diente distal mediano y romo, subdistal más grande y agudo, medio y basal pequeños en apariencia bicúspide, con una fila de tres setas menores en la parte distal: ms3 se localiza en la parte distal ventroexterna, de forma oblicua a esta se encuentra la seta ms2 y de forma oblicua a esta se encuentra la ms1, dorsal-proximal a los dientes distal y subdistal.
Pedipalpos con granulación fina en la superficie, carena dorsal en trocánter con gránulos gruesos, tres setas basales y dos distales; carena ventral desarrollada y con gránulos gruesos, seis setas basales y 10 a 12 distales; fémur con granulación irregular en la superficie dorsal prolateral y dorso-retrolateral, carenas dorsales y externa completas, granuladas, carena externa dentada, diente basal bien desarrollado, interna con gránulos irregulares e incompleta en la parte distal, carena ventral con granulación dispersa hacia la parte distal (Fig. 15d).
Patela (Fig. 15e) con la superficie finamente granulada, carena dorsomediana granulada, ligeramente sinuosa en la parte media, incompleta en la parte proximal, dorsal prolateral granulada y completa, dorsal retrolateral crenulada en la parte proximal y granular en la parte distal, ventral prolateral crenulada y desarrollada, ventral-retrolateral con dientes medianos y pequeños separados; carina prolateral con siete dientes, cuatro grandes y tres más pequeños, retrolateral presenta granulación débil, crenulada en la parte proximal y granulada en la parte distal.
Mano (Fig. 15f) 1,9 veces más larga que ancha y 2 veces más larga que alta, 0,7 veces la longitud del fémur y 2,1 veces más alta, 0,6 la longitud de la patela y 1,4 veces más alta; carenas digital y digital secundaria con gránulos en la parte basal a lisas distalmente, dorsal marginal y ventroexterna lisas, interna, ventral accesoria y externa secundaria finamente granuladas.
Dedo móvil con 8 hileras de dientecillos medios, hilera terminal formada por tres dientecillos, 9 gránulos accesorios externos y 10 gránulos accesorios internos separados por 1 a 5 gránulos más pequeños, más setoso en la parte proximal, ventralmente con granulación fina y dispersa, dedo fijo con 7 hileras de dientecillos medios y arreglo de gránulos accesorios internos y externos similar al dedo móvil.
Patas setosas, espolones tarsales en patas I-II simétricas, III-IV son asimétricas con el epolón externo bífido y el interno simple. Terguitos (Fig. 15g) con preterguitos I-VI con granulación fina, densa y uniforme en toda la superficie, borde posterior del preterguito finamente granulado, borde posterior del posterguito con granulación irregular. Carena media granular, terguito I con dos gránulos grandes anteriores y tres más pequeños posteriores, II con cuatro gránulos grandes y dos más pequeños, a partir del III la carena llega a la mitad del terguito y está formada por 6 gránulos grandes y 4 gránulos más pequeños; granulación en terguito IV con 12 gránulos irregulares, el posterior más grande; terguito V con 13 gránulos en la carena, cinco de ellos en la parte anterior; VI con la carena formada por 16 gránulos, una doble fila de 5 pares de gránulos y posteriormente 6 gránulos grandes y 4 más pequeños; VII con la granulación de la carena media crenulada proximalmente, granulada distalmente, carenas dorsales granuladas y bien desarrolladas, superficie intercarinal y zonas laterales con granulación fina y dispersa, gránulos del borde posterior menos desarrollados.
Placa basal de los peines subrectangular, muesca media aparente, margen posterior recto, proporción longitud/ancho= 1,5; conteo de dientes pectíneos 23-23; placas genitales subtriangulares, ligeramente lobuladas en la zona media posterior, dos setas anteriores y un par medio basal; esternón subtriangular y profundo con un reborde poco desarrollado, seis setas, un par anterior, uno medio y uno posterior (Fig. 16d).
Esternitos con granulación muy fina a lisa, carenas débiles y lisas en los terguitos III-VI, VII con carenas medias lisas en la parte proximal a crenuladas distalmente, carenas laterales crenuladas; esternito III con 32 setas, en la parte anterior con un arreglo de 3 setas pareadas en la primera hilera, en la segunda 10 pares y la tercera hilera de seis setas sobre el borde posterior, todas forman un arreglo triangular, distribuidas de la parte media anterior en diagonal hacia el espiráculo; esternito IV con 22 setas, tres pares anteriores ligeramente recurvadas, 4 pares mediales entre los espiráculos, en esta misma línea un par medio basal en los bordes laterales y seis setas más en el borde posterior del esternito; 38 setas en esternito V, cuatro pares anteriores, al centro 26 setas agrupadas, un par medio en los bordes laterales y seis setas en el borde posterior del esternito; en VI 12 setas, tres pares anteriores formando un semicírculo y en el borde posterior 6 setas; en VII 10 setas, un par de setas anteriores, un par sobre la parte dorsal de las carenas laterales, 6 en carena media, un par en la parte proximal, un par en la zona media y un par en el borde posterior, también se aprecian algunas microsetas sin un patrón definido; carenas de crenuladas a lisas, bien desarrolladas (Fig. 12b).
Metasoma (Fig. 16a) con carenas bien desarrolladas en segmentos I-III, carenas dorsolaterales y carenas laterales supramedianas crenuladas, iniciando con un diente romo desarrollado y en la parte distal con dientes fuertes y desarrollados; carenas lateroventrales crenuladas en la parte proximal a granuladas en la parte distal, carenas ventrales submedianas bien desarrolladas, crenuladas a granuladas, carena lateral inframediana granulada, poco desarrollada en I, en II-V se aprecian algunos gránulos dispersos; carenas dorsolaterales poco desarrolladas, granuladas en IV iniciando con un diente romo desarrollado; carena lateral supramediana granulada en la parte proximal a crenulada y lisa en la parte distal; carenas dorsolaterales en V lisas, poco desarrolladas proximalmente e incompletas distalmente, carenas supramedianas, lateroventrales y ventralsubmedianas con gránulos débiles y dispersos.

Figura 16
Características diagnósticas de Centruroides rommeli
a. Metasoma. b. Vesícula de macho. c. Ornamentación de carapacho. d. Placa basal de los peines de macho. e. Placa basal de los peines de hembra. f. Vesícula de hembra. g. Patrón de color de carapacho de hembra. h. Hemiespermatóforo y detalle de la región de los lóbulos. Escala= 1 mm
Superficie intercarinal finamente granulada, segmentos I-IV con un patrón de seis setas distribuidas en tres pares en la parte externa de las carenas ventrales submedianas, el primer par sobre el borde anterior, el segundo en la parte media ubicados de manera asimétrica y uno más sobre el borde posterior, segmento metasomal V también con seis setas ubicadas entre el espacio de la carena ventral mediana y ventrolaterales, con el mismo patrón que en los segmentos anteriores; parte externa de las carenas ventrolaterales con un patrón de dos pares de setas en I-IV, el primero en posición anterior y otro en el cuarto posterior; segmento V con tres pares, el primero en la parte externa de las carenas ventrolaterales, el segundo en el cuarto posterior y el tercero sobre las carenas ventrolaterales y el borde posterior.
Telson (Fig. 16b) con la vesícula ovalada, tres veces más larga que alta (proporción longitud/alto= 2,1), superficie finamente granulada; superficie dorsal acanalada y con una carina media poco desarrollada que llega hasta la base del aguijón; tubérculo subaculear cónico muy débil, con cuatro macrosetas ventrales, el primer par en posición media lateral, el segundo ubicado en la base del tubérculo; aguijón grande y curvo.
Hemiespermatóforo flageliforme, diente o gancho basal espiniforme poco desarrollado, carena basal delgada pero desarrollada, distal más desarrollada y esclerotizada; tronco laminar, recto y solo ligeramente ensanchado en la parte media, carenas interna y externa recorren hasta más de la mitad del tronco, axial engrosada hacia el flexor del pie el cual está poco desarrollado y débilmente esclerotizado (Fig. 16h).
Variación:
Dimorfismo sexual típico del género con el metasoma del macho aproximadamente dos veces más largo que el mesosoma, mientras que en las hembras es de 1.56 veces; placa basal en machos es rectangular con una muesca evidente en el borde anterior, posterior recto (Fig. 16d), mientras que en las hembras es más cuadrangular y el borde posterior recurvado (Fig. 16e), conteo de dientes pectíneos 23-25 (moda 24, n= 8) en machos, 21-24 (modas 21, 23, n= 42) en hembras; carenas de las manos de los pedipalpos y del metasoma en hembras con mayor desarrollo; segmento metasomal V en machos es 3,7 veces más largo que alto, en hembras es más alto en la parte media dando una apariencia ahusada (proporción largo/alto= 2,7). Vesícula más alargada en machos, proporción longitud/alto= 2,1 (Fig. 16b); en hembras 1,7 (Fig. 16f, Tabla II).
Hay variación en el tamaño de los individuos adultos, entre 53-63 mm en machos (Fig. 14 a, b), entre 40 y 48 mm en hembras (Fig. 14 c, d). La parte anterior del carapacho presenta variación en la coloración de intensa a difusa, así como el pigmento en el surco medio anterior que puede encontrase inmaculado a ligeramente pigmentado. La granulación del carapacho también es variable, aunque en hembras en general es menos densa (Fig. 16g)
Aspectos ecológicos: La población a partir de la que se describe Centruroides rommeli sp. nov., se ubica en el municipio de Jalpa en la parte sur del estado de Zacatecas, en la Sierra Madre Occidental (Fig. 1). La localidad tipo se ubica en las coordenadas 21.6601N, -102.9710W en la colonia “Linda vista” a 1,420 msnm, la vegetación en los alrededores es de bosque tropical caducifolio, con áreas perturbadas en diferente grado, debido al uso de suelo para pastizales usados para pastoreo de ganado, así como extensas áreas de cultivo.
Los alacranes fueron capturados en colecta directa diurna rodando piedras y troncos que se encuentran entre la vegetación; en las áreas cultivadas adyacentes no se encontraron ejemplares. La nueva especie comparte el hábitat con otro escorpión de la familia Vaejovidae, Mesomexovis spadix (Hoffmann, 1931).
Conclusiones: De la comparación morfológica y morfométrica se obtuvieron diferencias suficientes para caracterizar a Centruroides infamatus sensu stricto y validarla como una especie que puede diferenciarse de otras morfológicamente similares que se encuentran en el centro-norte de México, incluido C. rommeli sp. nov.
Las diferencias morfológicas se soportan también con las distancias y afinidades genéticas que se pudieron obtener con el análisis molecular de los fragmentos de gen RNAm16S utilizados para este fin.
Con los tres tipos de evidencia utilizadas, se puede soportar la existencia de Centruroides rommeli sp. nov. como una especie válida que habita en la zona de Jalpa, Zacatecas, población anteriormente identificada como C. infamatus, especie aquí redescrita.
Se modifica la distribución de C. infamatus, anteriormente muy amplia y que ahora se restringe al estado de Guanajuato.
En otros estados en los que sus poblaciones se tienen identificadas como Centruroides infamatus, actualmente están en estudio. Al momento, con evidencias similares a las que utilizamos para este trabajo, se puede decir que corresponden a tres especies nuevas más (Ponce-Saavedra et al., en prep.).
Agradecimientos
Se agradece al Consejo Nacional de Humanidades, Ciencia y Tecnología de México (Conahcyt) por la beca de Posgrado otorgada al segundo autor durante su estancia en el Programa Institucional de Maestría en Ciencias Biológicas de la Universidad Michoacana de San Nicolás de Hidalgo y la beca posdoctoral en la misma Universidad otorgada a la tercera autora. También se agradece a la Coordinación de la Investigación Científica de la Universidad Michoacana de San Nicolás de Hidalgo por el apoyo económico al primer autor para el desarrollo del proyecto. Agradecemos especialmente al Dr. Andrés Ojanguren Affilastro (MACN-Conicet) por su exhaustiva y meticulosa revisión de nuestro manuscrito. Sus acertados comentarios dedicación y detalle como revisor son apreciados por los autores. Finalmente, los especímenes fueron recolectados bajo el documento SPARN/DGVS/074492/24, Permiso de Colector Científico de la Secretaría de Medio Ambiente y Recursos Naturales (SEMARNAT), México, otorgado a Margarita Vargas Sandoval, Directora y Curadora Principal del CAFBUM, de la Facultad de Biología del Laboratorio de Entomología, Universidad Michoacana de San Nicolás de Hidalgo).
REFERENCIAS
Acosta, L. E., Candido, D. M., Buckup, E. H., & Brescovit, A. D. (2008). Description of Zabius gaucho (Scorpiones, Buthidae), a new species from southern Brazil, with an update about the generic diagnosis. The Journal of Arachnology, 36(3), 491-501. https://doi.org/10.1636/H07-28.1
Berland, L. (1913). Note sur un Scorpion muni de deux queues [ARACHN.]. Bulletin Société Entomologlque De France. 251
Beutelspacher, B. C. R. (2000). Catálogo de los alacranes de México. Universidad Michoacana de San Nicolás de Hidalgo.
Brown, C. A. (1996). Interpopulation and Intersexual Variation in Pectine Tooth Counts in Centruroides vittatus (Scorpionida, Buthidae). The Journal of Arachnology, 24(3), 262-264. http://www.jstor.org/stable/3705937
Chávez-Haro, A.L. (2011). Alacranismo en León, Guanajuato, México. En G. D’Suze, G. A. Corzo-Burguete, J. F. Paniagua-Solís & P. Álvarez León (Eds) Emergencias por animales ponzoñosos en las Américas (pp. 1-16). Dicresa SA.
De Armas, L. F., & Martín-Frías E. (2000). Presencia del alacrán Centruroides vittatus (Say) (Scorpiones: Buthidae) en el estado de Durango, México. Revista Nicaragüense de Entomología, 47, 11-13.
De Armas, L. F., & Martín-Frías E. (2001). Estudio Taxonómico y Biogeográfico de los Alacranes Mexicanos del Género Centruroides (Scorpiones: Buthidae), Principales Problemas. En T. Payán Porras, M. Guerra Álvarez, A. Flores Díaz & M. A. Aguayo Saviñón (Eds.), Reflexiones sobre el megaproyecto del Istmo de Tehuantepec. Recursos, problemática y futuro (pp. 311-315). Universidad Autónoma Metropolitana.
De Armas, L. F., Martín-Frías, E., & Estévez-Ramírez, J. (2003). Lista anotada de las especies mexicanas del género Centruroides Marx 1890 (Scorpiones: Buthidae). Revista Ibérica de Aracnología, 8, 93-98.
De Armas, L. F., & Martín-Frías E. (2008). El Género Centruroides Marx 1890 (Scorpiones: Buthidae), en el Estado de Veracruz, México. Boletín de la Sociedad Entomológica Aragonesa, 43, 7-22.
Dehesa-Dávila, M. (1989). Epidemiological characteristics of scorpion sting in Leon, Guanajuato, México. Toxicon, 27(3), 281-286. https://doi.org/10.1016/0041-0101(89)90175-X
Dehesa-Dávila, M., Martin, B. M., Nobile, M., Prestipino, G., & Possani, L. D. (1994). Isolation of a toxin from Centruroides infamatus infamatus Koch scorpion venom that modifies Na+ permeability on chick dorsal root ganglion cells. Toxicon, 32(12), 1487-1493. https://doi.org/10.1016/0041-0101(94)90307-7
Dehesa-Dávila, M., Ramírez, A. N., Zamudio, F. Z., Gurrola-Briones, G., Liévano, A., Darszon, A., & Possani, L. D. (1996). Structural and functional comparison of toxins from the venom of the scorpions Centruroides infamatus infamatus, Centruroides limpidus limpidus and Centruroides noxius. Comparative Biochemistry and Physiology Part B: Biochemistry and Molecular Biology, 113(2), 331-339. https://doi.org/10.1016/0305-0491(95)02031-4
De-Souza, C.A.R., Candido, D.M., Lucas S.M., & Brescovit, A.D. (2009). On the Tityus stigmurus complex (Scorpiones, Buthidae). Zootaxa, 1987(1), 1-38. https://doi.org/10.11646/zootaxa.1987.1.1
Díaz Nájera, A. (1964). Alacranes de la República Mexicana. Identificación de ejemplares capturados en 235 localidades. Revista del Instituto de Salubridad y Enfermedades Tropicales, 24, 19-27.
Díaz Nájera, A. (1966). Alacranes de la República Mexicana. Clave para identificar especies de Centrurus. Revista de Investigación de Salud Pública, 26, 111-116.
Díaz Nájera, A. (1975). Listas y datos de distribución geográfica de los alacranes de México (Scorpionida). Revista del Instituto de Salubridad y Enfermedades Tropicales, 35, 3-33.
Esposito, L. A., Yamaguti, H. Y., Pinto-da-Rocha, R., & Prendini, L. (2018). Plucking with the plectrum: phylogeny of the New World buthid scorpion subfamily Centruroidinae Kraus, 1955 (Scorpiones: Buthidae) reveals evolution of three pecten-sternite stridulation organs. Arthropod Systematics & Phylogeny, 76(1), 87-122. https://doi.org/10.3897/asp.76.e31942
Fet, V., Kovařík, F., Gantenbein, B., Kaiser, R. C., Stewart, A. K., & Graham, M. R. (2018) Revision of the Mesobuthus caucasicus complex from Central Asia, with descriptions of six new species (Scorpiones: Buthidae). Euscorpius, 255, 1-77. https://dx.doi.org/10.18590/euscorpius.2018.vol2018.iss255.1
Fet, V. & Lowe, G. (2000). Family Buthidae C.L. Koch 1837. En V. Fet, W. D. Sissom, G. Lowe & M. E. Braunwalder (Eds.), Catalog of the Scorpions of the World (1758-1998) (pp. 54-286). New York Entomological Society. https://mds.marshall.edu/bio_sciences
Fitz-Simmons, N. N. (1997). Male Marine Turtles. Gene Flow. Philopatry and Mating Systems of the Green Turtle Chelonia mydas. [Tesis Doctoral. Universidad de Queensland, Australia]. https://doi.org/10.14264/365841
Francke, O. F. (1977). Scorpions of the genus Diplocentrus Peters from Oaxaca, México. The Journal of Arachnology, 4, 145-200.
Francke, O. F. (1984). Centrurus limpidus Karsch, 1879 and Centruroides ornatus Pocock, 1902 (Arachnida, Scorpiones): proposed conservation. Z. N. (S.) 2446. Bulletin of the Zoological Nomenclature, 41(2), 98.
Chaneke Francke, O. F., Teruel, R., & Santibáñez-López, C. E. (2014) A new genus and a new species of scorpion (Scorpiones: Buthidae) from southeastern Mexico. The Journal of Arachnology, 42(3), 220-232. http://www.jstor.org/stable/24365296
Francke, O. F. (2019). Scorpions (Arachnida: Scorpiones) from the Cuatro Ciénegas Basin. En F. Álvarez & M. Ojeda (Eds.), Animal Diversity and Biogeography of the Cuatro Ciénegas Basin, Cuatro Ciénegas Basin: An Endangered Hyperdiverse Oasis (pp. 53-59). Springer. https://doi.org/10.1007/978-3-030-11262-2_4
Gantenbein, B., Fet, V., Largiadèr, C. R., & Scholl, A. (1999). First DNA phylogeny of Euscorpius Thorell, 1876 (Scorpiones: Euscorpiidae) and its bearing on taxonomy and biogeography of this genus. Biogeographica, 75, 49-65.
Gantenbein, B., Fet V., Barker M. D., & Scholl, A. (2000). Nuclear and mitochondrial markers reveal the existence of two parapatric scorpion species in the Alps: Euscorpius germanus (C. L. Koch, 1837) and E. alpha Caporiacco, 1950, stat. nov. (Scorpiones, Euscorpiidae). Revue Suisse Zoologie, 107(4), 843-869.
Gantenbein, B., Fet, V., & Barker, M.D. (2001). Mitochondrial DNA markers reveal a deep, divergent phylogeny in Centruroides exilicauda (Wood) (Scorpiones: Buthidae). En V. Fet & P. A. Selden (Eds), Scorpions 2001. In Memoriam Gary A. Polis (pp. 235-244). British Arachnological Society.
González-Santillán, E. (2001). Catálogo de escorpiones de la Colección Nacional de Arácnidos (CNAN). [Tesis, Facultad de Ciencias, Universidad Nacional Autónoma de México, México, D. F]. https://hdl.handle.net/20.500.14330/TES01000297144
González-Santillán, E., & Possani, L. D. (2018). North American scorpion species of public health importance with a reappraisal of historical epidemiology. Acta tropica, 187, 264-274. https://doi.org/10.1016/j.actatropica.2018.08.002
González-Santillán, E., Galán-Sánchez, M. A., & Valdez-Velázquez, L. L. (2019). A new species of Centruroides (Scorpiones, Buthidae) from Colima, Mexico. Comptes Rendus Biologies, 342(9-10), 331-344. https://doi.org/10.1016/j.crvi.2019.10.002
González-Sponga, M.A. (2011). Escorpiones, características, distribución geográfica y comentarios generales. En G. D’Suze, G. A. Corzo-Burguete, J. F. Paniagua-Solís & P. Álvarez León (Eds), Emergencias por animales ponzoñosos en las Américas (pp. 108). Instituto Bioclon.
Graham, M. R., Myers, E. A., Kaiser, R. C., & Fet, V. (2019). Cryptic species and codiversification in sand scorpions from the Karakum and Kyzylkum deserts of Central Asia. Zoologica Scripta, 48(6), 801–812. https://doi.org/10.1111/zsc.12381
Hammer, Ø., Harper, D. A. T., & Ryan, P. D. (2001). PAST: Paleontological statistics software package for education and data analysis. Palaeontologia Electronica, 4(1), 1-9. https://www.nhm.uio.no/english/research/resources/past/
Hoffmann, C.C. (1931). Monografías para la entomología médica de México. Monografía Num. 2, Los escorpiones de México. Primera parte: Diplocentridae, Chactidae, Vaejovidae. Anales del Instituto de Biología Universidad Nacional Autónoma de México, 2(4), 291-408.
Hoffmann, C. C. (1932). Monografías para la Entomología Médica de México. Monografía Num. 2, Los escorpiones de México. Segunda parte: Buthidae. Anales del Instituto de Biología Universidad Autónoma de México, 3-4, 283-361.
Hoffmann, C. C. (1937). Nota acerca de los alacranes del Valle de Mezquital, Hidalgo. Anales del Instituto de Biología Universidad Nacional Autónoma de México, 8(1-2), 203-204.
Hoffmann, C. C. (1938). Nuevas consideraciones acerca de los alacranes de México. Anales del Instituto de Biología Universidad Nacional Autónoma de México, 9, 317-337
Jochim, E. E., Broussard, L. M., & Hendrixson, B. E. (2020). Integrative species delimitation and taxonomic status of the scorpion genus Vaejovis Koch, 1836 (Vaejovidae) in the Santa Catalina Mountains, Arizona. Euscorpius, 316, 1-11. https://mds.marshall.edu/euscorpius/vol2020/iss316/1/
Keegan, H. L. (1980). Scorpions of medical importance. Jackson: Univ. Press of Mississippi, 19, 17-72.
Koch, C. L. (1845). Die Arachniden: Getreu nach der Natur abgebildet und beschrieben. Der C.H. Zeh'schen Buchhandlung.
Kopstein, F. (1921). Die Skorpione des Indo-Australischen Archipels. Mit Grundlage der in Hollländischen Samrnlungen, Vornähmlich des Rijks-Museums in Leiden, Vor-Handenen Arten. Zoologische Mededeelingen, 13, 115-144.
Kovařík, F., Teruel, R., & Lowe, G. (2016). Two new scorpions of the genus Chaneke Francke, Teruel et Santibáñez-López, 2014 (Scorpiones: Buthidae) from southern Mexico. Euscorpius, 218, 1-20. https://dx.doi.org/10.18590/euscorpius.2016.vol2016.iss218.1
Kraepelin, K. (1891). Revision der Skorpione. I. Die Familie der Androctonidae. Beiheft zum Jahrbuch der Hamburgischen Wissenschaftlichen Anstalten, 8, 121-127.
Kraepelin, K. (1893). Revision der Skorpione. II. Scorpionidae und Bothriuridae. Beiheft zum Jahrbuch der Hamburgischen Wissenschaftlichen Anstalten, 9, 263-286
Kraepelin, K. (1894) Revision der Skorpione. Mitteilungen aus dem Naturhistorischen Museum de Hamburgo. Beiheft zum Jahrbuch der Hamburgischen Wissenschaftlichen Anstalten, 12, 95.
Kraepelin, K. (1899). Scorpiones und Pedipalpi. In F. Dahl (Ed.), Das Tierreich (Vol. 8, pp. 88-90). R. Friedlander und Sohn Verlag.
Kraepelin, K. (1901). Catalogue des Scorpions des collections du Museum d'histoire naturelle de Paris. Bulletin du Muséum d'Histoire Naturelle, 49, 265-274.
Kraepelin, K. (1905). Die geographische Verbreitung der Scorpione. Zoologische Jahrbücher, Abteilung für Systematik, Geographie und Biologie der Thiere, 22, 321-364.
Kraepelin, K. (1908). Die sekundären Geschlechtscharaktere der Skorpione, Pedipalpen und Solifugen. Mitteilungen aus dem Naturhistorischen Museum, 25(2), 181-225.
Lampe, E. (1917). Katalog der Skorpione, Pedipalpen und Solifugen des Naturhistorischen Museums der Residenzstadt Wiesbaden. Jahrbücher des Nassauischen Verein für Naturkunde, 70(1), 185-203.
Linares-Guillén, J.W. (2021). Análisis sistemático del complejo “Infamatus” del género Centruroides (Scorpiones: Buthidae) en México. [Tesis de Maestría. Universidad Michoacana de San Nicolás de Hidalgo. Morelia, Michoacán]. http://bibliotecavirtual.dgb.umich.mx:8083/xmlui/handle/DGB_UMICH/6445
Lönnberg, E. (1897a). Skorpioner och pedipalper i Upsala Universitets Zoologiska Museum. Entomologisk Tidskrift, 18,175-192.
Lönnberg, E. (1897b). Om Skorpionemas och Pedipalpernas geografiska Utbredning. Entomologisk Tidskrift, 18, 184-185.
Loria, S. F., & Prendini, L. (2014). Homology of the Lateral Eyes of Scorpiones: A Six-Ocellus Model. PLOS ONE, 9(12), 1-30.
Lourenço, W. R. (1980). Contribution à la connaissance systématique des Scorpions appartenant au "complexe" Tityus trivittatus Kraepelin, 1898 (Buthidae). Bulletin du Muséum National d'Histoire Naturelle (4e série), 2, 793-843.
Lourenço, W. R. (1981). Sur la systématique des scorpions appartenant au complexe Tityus stigmurus (Thorell, 1877) (Buthidae). Revista Brasileira Biologia, 41(2), 351-62.
Lourenço, W. R. (1997). Additions à la faune de scorpions néotropicaux (Arachnida). Revue suisse de Zoologie, 104(3), 587-604.
Lourenço, W. R., & Sissom, W. D. (2000). Scorpiones. En J. Llorente Bousquets, E. González Soriano & N. Papavero (Eds.), Biodiversidad, taxonomía y biogeografía de artrópodos de México: Hacia una síntesis de su conocimiento (Vol. 2, pp. 115-135). Facultad de Ciencias, UNAM, CONABIO y BAYER.
Lourenço, W. R., & Da Silva, E.A. (2007). New evidence for disrupted distribution pattern of the ‘Tityus confluens’ complex, with the description of a new species from the State of Pará, Brazil (Scorpiones, Buthidae). Amazoniana, 19(3/4), 77-86.
Lourenço, W. R. (2023). A new species of Bothriurus Peters (Scorpiones: Bothriuridae) from ‘Parque Estadual da Serra dos Martírios/Andorinhas’ in the State of Pará, Brazil. Faunitaxys, 11(4), 1-7. https://doi.org/10.57800/faunitaxys-11(4)
Lucas, S. M., & Meier, J. (2017). Biology and distribution of scorpions of medical importance. In J. Meier & J. White (Eds.), Handbook of clinical toxicology of animal venoms and poisons (pp. 205-219). CRC Press.
Luna-Ramirez, K, Miller, A.D., & Rašić, G. (2017). Genetic and morphological analyses indicate that the Australian endemic scorpion Urodacus yaschenkoi (Scorpiones: Urodacidae) is a species complex. PeerJ, 5, 2759. https://doi.org/10.7717/peerj.2759
Martínez-Tejeda, F. Y., & Ponce-Saavedra, J. (2020). Nuevos registros de alacranes (Arachnida: Scorpiones) en áreas naturales protegidas del norte de México. Boletín de la Sociedad Mexicana de Entomología, 6(3), 90-94.
Miranda-López, E. P. (2012). Análisis sistemático del complejo “Pusillus” (Scorpiones: Vaejovidae) en México [Tesis de maestría, Universidad Michoacana de San Nicolás de Hidalgo]. http://bibliotecavirtual.dgb.umich.mx:8083/xmlui/handle/DGB_UMICH/1684
Monod, L. Cauwet, L., González-Santillán, E. & Huber, S. (2017) The male sexual apparatus in the order Scorpiones (Arachnida): a comparative study of functional morphology as a tool to define hypotheses of homology. Frontiers in Zoology, 14(1), 1-48. https://doi.org/10.1186/s12983-017-0231-z
Mullen, G. R., & Sissom, W. D. (2019). Scorpions (Scorpiones). In G. R. Mullen, L. A. Durden, & J. G. King (Eds.), Medical and veterinary entomology (pp. 489-504). Academic Press. https://doi.org/10.1016/B978-0-12-814043-7.00023-6
Ochoterena, I. (1920). El alacrán de Durango (Centrurus exilicauda Wood). Memorias y Revista de la Sociedad Científica "Antonio Alzate", 37, 215-226.
Penther, A. (1913). Beitrag zur Kenntnis Amerikanischer Skorpione. Annalen des Kaiserlich-Koniglichen Naturhistorischen Hofmuseums in Wien, 27, 239-254.
Pocock, R. I. (1898). Descriptions of some new Scorpions from Central and South America. The Annals and Magazine of Natural History: Zoology, Botany, and Geology, 1(7), 421-431.
Pocock, R. I. (1902). Arachnida, Scorpiones, Pedipalpi and Solifugae. Biologia Centrali-Americana.
Polis, G. A. (1990). Ecology. En G. A. Polis (Ed.), The biology of scorpions (pp. 247-293). Stanford University Press.
Ponce-Saavedra, J., Moreno-Barajas, R., & Beutelspacher, C. R. (1999). Características morfológicas de Centruroides infamatus Koch y C. limpidus Karsch (Scorpiones: Buthidae), especies comunes en el Estado de Michoacán, México. En Memorias del XXXIV Congreso Nacional de Entomología. Sociedad Mexicana de Entomología.
Ponce-Saavedra, J., & Beutelspacher, C. R. (2001). Alacranes de Michoacán. Ediciones Michoacanas.
Ponce-Saavedra, J., & Francke, O. F. (2004). Una nueva especie de alacrán del género Centruroides Marx (1890) (Scorpionbes, Buthidae) de la depresión del Balsas, México. Acta zoológica mexicana, 20(2), 221-232. https://doi.org/10.21829/azm.2004.2022340
Ponce-Saavedra, J., & Moreno-Barajas, R. (2005). El Género Centruroides Marx 1890 (Scorpiones: Buthidae) en México. Biológicas, 7, 42-51.
Ponce-Saavedra, J., Francke, O. F., Cano-Camacho, H., & Hernández-Calderón, E. (2009). Evidencias morfológicas y moleculares que validan como especie a Centruroides tecomanus (Scorpiones, Buthidae). Revista Mexicana de Biodiversidad, 80(1), 71-84. http://doi.org/10.22201/ib.20078706e.2009.001.585
Ponce-Saavedra, J., & Francke, O. F. (2013a). Actualización taxonómica sobre alacranes del Centro Occidente de México. Dugesiana, 20(2), 73-79. https://doi.org/10.32870/dugesiana.v20i2.4088
Ponce-Saavedra, J., & Francke, O. F. (2013b). Clave para la identificación de especies de alacranes del género Centruroides Marx 1890 (Scorpiones: Buthidae) en el Centro Occidente de México. Biológicas, 15(1), 52-62.
Ponce-Saavedra, J., Quijano-Ravell, A. F., Teruel, R., & Francke, O. F. (2015). Redescription of Centruroides ornatus Pocock, 1902 (Scorpiones Buthidae), a montane scorpion from central Mexico. Revista Ibérica de Aracnología, 27, 81-89.
Ponce-Saavedra, J., Francke, O. F., Quijano-Ravell, A. F., & Cortés Santillán, R. (2016) Alacranes (Arachnida: Scorpiones) de importancia para la salud pública en México. Folia Entomológica Mexicana (nueva serie), 2(3), 45-70. http://www.acaentmex.org/folia/revista/Num%202016_3/45-70.pdf
Ponce-Saavedra, J., & Francke, O. F. (2019). Una especie nueva de alacrán del género Centruroides (Scorpiones: Buthidae) del noroeste de México. Revista Mexicana de Biodiversidad, 90, e902660. https://doi.org/10.22201/ib.20078706e.2019.90.2660
Ponce-Saavedra, J., Linares-Guillén, J. W., & Quijano-Ravell, A. F. (2022). Una nueva especie de alacrán del género Centruroides Marx (Scorpiones: Buthidae) de la costa Noroeste de México. Acta Zoológica Mexicana, 38(1),1-24. https://doi.org/10.21829/azm.2022.3812517
Possani, L. D., Gurrola, G.B., Schwartz, E. F., & Restano-Cassulini, R. (2011). Bioquímica y biología molecular de los venenos de escorpiones de importancia médica en el continente americano. En G. D’Suze, G. A. Corzo-Burguete, J. F. Paniagua-Solís & P. Álvarez León (Eds) Emergencias por animales ponzoñosos en las Américas (pp. 147-180). Instituto Bioclon.
Prendini, L., Crowe, T. M., & Wheeler, W. C. (2003). Systematics and biogeography of the family Scorpionidae (Chelicerata: Scorpiones), with a discussion on phylogenetic methods. Invertebrate Systematics, 17, 185-259. https://doi.org/10.1071/IS02016
Quijano-Ravell, A. F., De Armas, L. F., Francke, O. F., & Ponce-Saavedra, J. (2019). A new species of the genus Centruroides Marx (Scorpiones, Buthidae) from western Michoacán State, México using molecular and morphological evidence. Zookeys, 859, 31-48. https://doi.org/10.3897/zookeys.859.33069
Quijano-Ravell, A. F., & Ponce-Saavedra, J. (2016a). Uso del hábitat por Centruroides ornatus Pocock 1902 (Scorpiones: Buthidae) en el cerro “EL Águila” Morelia, Michoacán, México. Entomología Mexicana, 3, 85-90.
Quijano-Ravell, A. F., & Ponce-Saavedra, J. (2016b). A new species of scorpion of the genus Centruroides (Scorpiones: Buthidae) from the state of Michoacán, Mexico. Revista mexicana de biodiversidad, 87(1), 49-61. https://doi.org/10.1016/j.rmb.2016.01.022
Quijano-Ravell, A. F., Ponce-Saavedra, J., Francke, O. F., & Villaseñor-Ramos, M. A. (2010). Nuevos registros y distribución actualizada de Centruroides tecomanus Hoffman, 1932 (Scorpiones: Buthidae). Ciencia Nicolaita, 52, 179-189.
Rein, J. O. (2025). The Scorpion Files. Trondheim: Norwegian University of Science and Technology. https://www.ntnu.no/ub/scorpion-files/
Santibáñez-López, C. E., Francke, O. F., Ureta, C., & Possani, L. D. (2015). Scorpions from Mexico: From species diversity to venom complexity. Toxins, 8(1), 1-18. https://doi.org/10.3390/toxins8010002
Say, T. (1821). Order II POLYMEROSTOMATA. Journal of the Academy of Natural Sciences of Philadelphia, II, 61-62.
Shelley, R. M., & Sissom, W. D. (1995). Distributions of the scorpions Centruroides vittatus (Say) and Centruroides hentzi (Banks) in the United States and Mexico. Journal of Arachnology, 23, 100-110.
Simon, C., Frati, F., Beckembach, A., Crespi, B., Liu, H., & Flook, P. (1994). Evolution weighting and phylogenetic utility of mitochondrial gene sequences and compilation of conserved polymerase chain reaction primers. Annals of the entomological Society of America, 87, 651-701. https://doi.org/10.1093/aesa/87.6.651
Sissom, W. D., Polis, G.A., & Watt, D. D. (1990). Field and laboratory methods. En Polis, G.A. (Ed.), The Biology of Scorpions (pp. 445-461). Stanford University Press.
Sistema Nacional de Vigilancia Epidemiológica/Sistema Único de Información. (2025). Boletín Epidemiológico. Dirección General de Epidemiología. Secretaría de Salud, 8(42), 1-88. https://www.gob.mx/salud/documentos/boletinepidemiologico-sistema-nacional-de-vigilancia-epidemiologica-sistema-unico-de-informacion-387843
Stahnke, H. L. (1970). Scorpion nomenclature and mensuration. Entomological news, 81(12), 297-316.
Stahnke, H.L., & Calos, M. (1977). A key to the species of the genus Centruroides Marx (Buthidae, Scorpionida). Entomological News, 88(5-6), 111-120
Strand, E. (1916). Ueber einige Arachniden aus Buea in Kamerun. Schaftliche Ergebnisse der Deutschen Zentral Afrika Expedition 1907-1908, unter Fuhrung Adolg Friedrichs, Herzogs zu Mecklenberg. Archiv Für Naturgeschichte, 81(A11), 139-149.
SUIVE. (2024). Dirección General de Epidemiología. Secretaría de Salud. https://www.gob.mx/salud/acciones-y-programas/historico-boletin-epidemiologico
Talal, S., Tesler, I., Sivan, J., Ben-Shlomo, R., Tahir, H. M., Prendini, L., Snir, S., & Gefen, E. (2015). Scorpion speciation in the Holy Land: multilocus phylogeography corroborates diagnostic differences in morphology and burrowing behavior among Scorpio subspecies and justifies recognition as phylogenetic, ecological and biological species. Molecular phylogenetics and evolution, 91, 226-237. http://doi.org/10.1016/j.ympev.2015.04.028
Tamura, K., Stecher, G. & Kumar, S. (2021). MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Molecular Biology and Evolution, 38(7), 3022-3027. https://doi.org/10.1093/molbev/msab120
Teruel, R., Fet, V., & Graham, M. R. (2006). The first mitochondrial DNA phylogeny of Cuban Buthidae (SCORPIONES: BUTHOIDEA). Boletín Sociedad Entomológica Aragonesa,1(39), 219-226.
Teruel, R., & Kovařík, F. (2010) The true identity of the enigmatic scorpion Centruroides subgranosus (Kraepelin, 1898), with some taxonomic comments on Centruroides vittatus (Say, 1821) and Centruroides suffusus Pocock, 1902 (Scorpiones: Buthidae). Euscorpius, 97, 1-9. https://dx.doi.org/10.18590/euscorpius.2010.vol2010.iss97.1
Towler, W. I., Ponce-Saavedra, J., Gantenbein, B., & Fet, V. (2001). Mitochondrial DNA reveals a divergent phylogeny in tropical Centruroides (Scorpiones: Buthidae) from Mexico. Biogeographica, 77(4), 157-172.
Vachon, M. (1963). De l'utilité, en systématique, d'une nomenclature des dents des chélicères chez les Scorpions. Bulletin du Muséum National d'Histoire Naturelle, 35(2), 161-166.
Vachon, M. (1974). Études des caractères utilisés pour classer les familles et les genres des scorpions (Arachnides). 1. La trichobothriotaxie en arachnologie. Sigles trichobothriaux et types de trichobothriotaxie chez les Scorpions. Bulletin du Muséum National d'Histoire Naturelle (3e série), 104(140), 857-958.
Yamashita, T., & Rhoads, D. D. (2013). Species delimitation and morphological divergence in the scorpion Centruroides vittatus (Say, 1821): insights from phylogeography. PLoS One, 8(7), e68282. https://doi.org/10.1371/journal.pone.0068282
Koch, C.L. 1850. Scorpionen. En: C. L. Koch (Ed.), Übersicht des Arachnidensystems (volume 5, pp. 86-92). C.H. Zeh.
Lampe, E. (1910). Verzeichnis der Neuerwerbungen des Naturhistorischen Museums. Jahrbücher des Nassauischen Vereins für Naturkunde, 63, 18-25.
Notas
Centruroides rommeli sp. nov. Holotipo y paratipos se depositaron en la Colección Nacional de Arácnidos del Instituto de Biología de la Universidad Nacional Autónoma de México (CNAN-T01621, CNAN-T01622, CNAN-T01623, CNAN-T01624) y dos ejemplares en la colección de referencia del Laboratorio de Entomología “Biol. Sócrates Cisneros Paz” de la Facultad de Biología de la Universidad Michoacana de San Nicolás de Hidalgo (CAFBUM), con los datos (CAFBUM-SB-620, CAFBUM-SB-621)
Centruroides infamatus. Topotipos depositados en la Colección Nacional de Arácnidos del Instituto de Biología de la Universidad Nacional Autónoma de México CNAN (SC772) y en la colección de referencia del Laboratorio de Entomología “Biol. Sócrates Cisneros Paz” de la Facultad de Biología de la Universidad Michoacana de San Nicolás de Hidalgo (CAFBUM), con los datos de CAFBUM (SB-1727, 1728, 1737, 1738)
Notas de autor
wilfrido.linares@umich.mx
Información adicional
redalyc-journal-id: 3220