Revisiones
Sintiencia en insectos: reflexiones éticas para una Entomología del siglo XXI
Sentience in insects: ethical reflections for a 21st-century Entomology
Sintiencia en insectos: reflexiones éticas para una Entomología del siglo XXI
Revista de la Sociedad Entomológica Argentina, vol. 84, núm. 4, e0411, 2025
Sociedad Entomológica Argentina
Recepción: 26 Julio 2025
Aprobación: 07 Octubre 2025
Resumen: En la Declaración de Conciencia Animal de Nueva York, la posibilidad de que los insectos posean experiencias conscientes ya fue reconocida. En este trabajo, se presenta una revisión basada en evidencias actuales que apoya dicha posibilidad y se ofrecen argumentos relevantes para el debate sobre su capacidad de sentir dolor y las implicancias éticas para la Entomología. Se exponen hallazgos relacionados a ocho criterios neurobiológicos y conductuales propuestos como indicadores de sintiencia. Se describen comportamientos complejos observados en insectos, como el juego, el aprendizaje social, el uso de herramientas, la transmisión cultural y el reconocimiento individual, los cuales desafían la visión tradicional de los insectos como autómatas. Se discuten las diferencias entre humanos e insectos, así como los sesgos y actitudes humanas que limitan nuestra empatía hacia estos animales. Como marco ético ante la incertidumbre, se propone el principio de precaución para guiar decisiones en investigación, educación, producción y control de plagas. Se promueve la adopción de métodos de colecta menos invasivos y se sugieren mejoras en las condiciones de bienestar en biofábricas. Finalmente, se identifican protocolos de sacrificio ético en dos etapas, diseñados para reducir al mínimo el sufrimiento individual.
Palabras clave: Bienestar animal, Capacidades cognitivas en insectos, Dolor en insectos, Ética en Entomología, Principio de precaución.
Abstract: In the New York Declaration on Animal Consciousness, the possibility that insects may have conscious experiences was acknowledged. In this work, a review based on current evidence is presented to support this possibility, and relevant arguments are offered for the debate on their capacity to feel pain and its ethical implications for Entomology. Findings related to eight neurobiological and behavioral criteria proposed as indicators of sentience are presented. Complex behaviors observed in insects—such as play, social learning, tool use, cultural transmission, and individual recognition—are described, challenging the traditional view of insects as automatons. Differences between humans and insects are discussed, along with human biases and attitudes that limit empathy toward these animals. As an ethical framework under uncertainty, the precautionary principle is proposed to guide decisions in research, education, production, and pest control. The adoption of less invasive collection methods is encouraged, and improvements in welfare conditions in insect biofactories are suggested. Finally, two-step ethical killing protocols designed to minimize individual suffering are identified.
Keywords: Animal welfare, Cognitive capacities in insects, Ethics in Entomology, Pain in insects, Precautionary principle.
INTRODUCCIÓN
Recientemente, más de 500 expertos en neurociencias entre ellos biólogos, médicos veterinarios, sociólogos y filósofos, firmaron en Nueva York la Declaración de Conciencia Animal (Andrews et al., 2024). En ella, reconocen la posibilidad real de que los insectos puedan tener experiencias conscientes, cuestión que se ha debatido desde hace tiempo (Dawkins, 2006; Barron & Klein, 2016; Klein & Barron, 2016; DeGrazia, 2020; Andrews, 2023; Andrews et al., 2025). La forma más básica de conciencia es tener experiencias subjetivas, es decir tener la impresión de que algo nos sucede como individuos (Barron & Klein, 2016), y se encuentra estrechamente ligada a la capacidad de los organismos para sentir dolor. El concepto inglés “sentience” y el español "sintiencia" (aún no oficialmente aceptado) hacen referencia a la capacidad de un organismo para experimentar el mundo de manera subjetiva, es decir, para valorar positiva o negativamente sus vivencias, como el placer, el hambre, la sed, la calidez o el dolor. Por su parte, el dolor se define como “una experiencia sensorial y emocional desagradable asociada con un daño tisular real o potencial, o descrita en términos de dicho daño” (Raja et al., 2020). Debido a que se trata de una vivencia inherentemente individual, resulta difícil afirmar con certeza si los insectos sienten dolor. Sin embargo, si la respuesta fuese afirmativa, entonces existiría una responsabilidad moral respecto al modo en que los tratamos.
La declaración firmada en Nueva York representa un paso clave de la comunidad científica internacional que debería llevarnos, como entomólogos, a reconsiderar nuestras prácticas a la hora de estudiar y manipular a estos organismos. En nuestro país, los Comités Institucionales para el Cuidado y Uso de Animales de Laboratorio (CICUAL) apenas mencionan a los invertebrados entre los sujetos cuyo bienestar merezca consideración. No obstante, en otros países ya se incluyen a cefalópodos y crustáceos en los protocolos de bienestar animal, y se han logrado avances significativos en el diseño de condiciones de cría y experimentación adecuadas para invertebrados (Freelance, 2019; van Huis, 2020; Fischer et al., 2023). Por tanto, sería deseable que todo entomólogo cuente con criterios bien fundamentados para tomar decisiones relacionadas con el bienestar de insectos en cautiverio, así como para asesorar a otros en dichas decisiones, ya sea en ámbitos científicos, educativos, productivos o comunitarios.
La sintiencia en insectos presenta múltiples dimensiones, todas valiosas e interesantes. Sin embargo, somos conscientes de que no podremos abordar todas en este artículo. Nuestro objetivo es ofrecer una visión general sustentada en evidencias actuales y presentar argumentos que sirvan de base para el debate con estudiantes, profesionales y personas interesadas en el tema. Empezaremos abordando el prolongado debate entre neurocientíficos sobre la posibilidad de que los insectos sientan dolor, para luego examinar los comportamientos complejos y estados emocionales que estos organismos pueden manifestar. A continuación, analizaremos las diferencias entre humanos e insectos, así como los sesgos y actitudes humanas que limitan nuestra empatía hacia estos animales. Dado que reflexionar sobre la sintiencia implica adoptar una perspectiva ética centrada en el individuo como sujeto capaz de experimentar sensaciones subjetivas, también abordaremos el dilema que plantea la aplicación de criterios éticos, tanto individuales como colectivos, al momento de definir qué tipo de consideración moral debería prevalecer (o ha prevalecido en la práctica) en nuestro trato hacia los insectos. Finalmente, propondremos algunas vías para repensar nuestras actitudes hacia los insectos en los distintos ámbitos en los que intervienen los entomólogos: la investigación científica, la enseñanza, la producción industrial y el control de plagas, a fin de que nuestras decisiones se fundamenten en criterios de responsabilidad y precaución acordes con la complejidad del tema.
EVIDENCIAS DE SINTIENCIA EN INSECTOS
Para abordar la cuestión de la sintiencia en cualquier organismo, es fundamental partir de un debate filosófico de larga data, conocido como el problema de otras mentes (Andrews, 2023); dicho problema señala que todo intento de comprender cómo podría ser la experiencia subjetiva de otro ser, especialmente si no es humano, está inevitablemente teñido de cierto grado de antropocentrismo y antropomorfismo (Nagel, 1974; de Waal, 1999; Harrison & Hall, 2010; de Souza Valente, 2025). Es decir, tendemos a atribuir a los animales emociones, pensamientos y vivencias humanas (Döring & Chittka, 2011; Scotto, 2024).
Desde nuestra perspectiva, los insectos no siempre muestran señales claras de dolor, y de hecho, exhiben comportamientos que, a primera vista, sugieren la ausencia total de sufrimiento. Un ejemplo frecuentemente citado es el del macho de mantis religiosa que continúa copulando mientras que la hembra le devora la cabeza, o el de ciertos grillos que llegan a ingerir sus propias vísceras tras sufrir heridas abdominales (Eiseman et al., 1984; Adamo, 2019). No obstante, es importante tomar con cautela este tipo de observaciones (Barrett & Fischer, 2024). A diferencia de los mamíferos, los insectos no siempre manifiestan una inmovilización análoga ante una lesión dolorosa, lo que podría deberse a que, dada su corta vida, la inmovilidad frente al dolor no ha resultado ser una estrategia adaptativa. En el caso del macho de mantis, por ejemplo, su aparente indiferencia al dolor durante el apareamiento contrasta con su respuesta defensiva frente a las conductas depredadoras de la hembra antes de que comience la cópula, apoyando la existencia de procesos de nocicepción (Elwood, 2023).
Esto refuerza la idea de que la perspectiva humana del dolor no siempre es adecuada para comprender el comportamiento de otros animales, especialmente los más alejados evolutivamente (Harrison & Hall, 2010; Rodríguez-Rodríguez, 2024). Así, por ejemplo, el sistema nervioso de los insectos, aunque pequeño y anatómicamente distinto al de los mamíferos, es capaz de realizar funciones sorprendentemente complejas con gran precisión (Simons & Tibbets, 2019; Makarova et al., 2021). En este sentido, es sabido que los insectos ven a pesar de no tener corteza visual (Sanes & Zipursky, 2010; Gibbons et al., 2022a). Este dato nos lleva a considerar que el dolor también podría procesarse a través de circuitos neuronales no homólogos a los mamíferos, pero funcionalmente equivalentes. Por lo tanto, la ausencia de evidencia no puede tomarse como evidencia de ausencia de dolor (Durrant, 2024).
1. Neurobiología y nocicepción
Considerando el conjunto de evidencia actualmente disponible, es más prudente hablar de la probabilidad de que un organismo experimente dolor que afirmar con certeza su presencia o ausencia (Barrett & Fischer, 2024). Con este enfoque, se han propuesto ocho criterios conductuales y fisiológicos aplicables a distintos grupos animales (Birch et al., 2021) y a insectos en general (Gibbons et al., 2022a, b), que se detallan en los siguientes apartados. Bajo esta perspectiva, cuantos más indicadores de dolor se observen, mayor será la probabilidad de que el organismo en cuestión posea dicha capacidad. Sin embargo, hasta el momento se han estudiado pocas especies de insectos en este campo, lo que dificulta las generalizaciones. Finalmente, es necesario subrayar que las diferencias entre los estados inmaduros y adultos en insectos podrían implicar distintos niveles de sintiencia, por lo que es esencial investigar ambos estados por separado.
Un abordaje similar, basado en un gran número de indicadores fisiológicos, comportamentales y cognitivos, ha sido desarrollado detalladamente por la organización sin fines de lucro Rethink Priorities (Schukraft, 2019a,b, c, d).
● Criterio 1- Presencia de nociceptores: La nocicepción es el proceso mediante el cual un organismo detecta y responde de forma inconsciente a estímulos potencialmente dañinos, gracias a receptores especializados denominados nociceptores (Birch, 2021). En algunos insectos, se ha demostrado la existencia de neuronas multidendríticas ubicadas bajo la epidermis, cuyas membranas contienen canales iónicos sensibles que responden únicamente a estímulos nocivos (Gibbons et al., 2022a). En larvas de Manduca sexta (L) (Lepidoptera: Sphingidae), por ejemplo, estos canales han sido caracterizados como responsables de respuestas defensivas frente a daños en el tegumento (Caron et al., 2020). Además, estudios realizados en Drosophila melanogaster Meigen (Diptera: Drosophilidae) han utilizado mutantes con canales iónicos bloqueados, lo que confirma que la ausencia de estos canales impide las respuestas típicas ante estímulos dañinos (Tracey et al., 2003).
● Criterios 2 y 3- Regiones cerebrales integradoras e integración de la nocicepción: Un indicio relevante sobre la posibilidad de que un organismo experimente dolor es la existencia de regiones cerebrales capaces de integrar información sensorial proveniente de diversas fuentes, incluyendo la emitida por los nociceptores (Gibbons et al., 2022a). En los insectos, se han identificado varias estructuras cerebrales con funciones integradoras complejas, una de las más destacadas son los cuerpos fungiformes, considerados centros de aprendizaje, memoria e integración sensorial, y que han sido propuestos como asiento de una forma básica de “inteligencia” en insectos (Traniello & Avergués Weber, 2023). Estas estructuras están presentes en todos los insectos, excepto en los pececitos de plata (Zygentoma). Otra región clave es el complejo central, que participa en múltiples funciones, incluyendo la navegación espacial, el control de la locomoción, la memoria y, según evidencia reciente, también la nocicepción (Barron & Klein, 2016). El cuerno lateral es otra estructura relevante, asociada principalmente a la memoria olfativa y a la regulación de comportamientos relacionados con el olfato. Recibe impulsos de múltiples modalidades sensoriales, lo que refuerza su papel como centro de integración (Das Chakraborty & Sachse, 2021). Cabe destacar que estas estructuras no suelen estar presentes en los estados inmaduros de insectos con metamorfosis completa. En cambio, en especies con metamorfosis incompleta, se desarrollan de manera progresiva a lo largo de las etapas juveniles (Buehlmann et al., 2020; Thiagarajan & Sachse, 2022).
● Criterio 4- Analgesia: Un indicador importante de la posible experiencia de dolor en un organismo es la modulación de su respuesta conductual ante estímulos nocivos mediante compuestos que actúan sobre el sistema nervioso (Gibbons et al., 2022a; Pasquini et al., 2025). Esta modulación puede darse por dos vías: 1) la acción de neurotransmisores endógenos que el propio organismo produce para atenuar sus respuestas frente a estímulos reales o potencialmente dañinos, 2) la reacción a compuestos exógenos como anestésicos locales, analgésicos (incluidos opioides), ansiolíticos o antidepresivos. En ambos casos, estos compuestos modifican la respuesta del insecto ante estímulos nocivos ya que se atenúa la experiencia de dolor, angustia o daño (Gibbons et al., 2022a). Otros experimentos también han demostrado la acción de diversos compuestos opioides sobre insectos (Gibbons et al., 2022a; Santos-Silva et al., 2022). Por ejemplo, se ha demostrado que las cucarachas escapan más lentamente de una caja con temperatura elevada si se les inyecta morfina (Gritsai et al., 2004), y que la morfina disminuye la agresividad que despliegan las abejas al recibir una ronda de descargas eléctricas (Núñez, 1983).
● Criterio 5- Compromisos motivacionales: Este término hace referencia a la capacidad de tomar decisiones de forma flexible, lo que implica un procesamiento integrador de la información (Gibbons et al., 2022a, b). En este contexto, la flexibilidad conductual se interpreta como evidencia de que el insecto es capaz de evaluar el disvalor de un estímulo nocivo frente al valor de una posible recompensa. Para que se considere que existe un compromiso motivacional, debe demostrarse que el animal asigna valor a las distintas opciones disponibles y realiza comparaciones entre ellas utilizando una especie de “moneda común” interna (Gibbons et al., 2022a). Uno de los ejemplos más representativos de este tipo de comportamiento proviene de estudios realizados con abejorros (Bombus terrestris L.) (Hymenoptera: Apidae) (Gibbons et al., 2022b). En una primera etapa experimental, los insectos deben elegir entre dos ubicaciones: una de ellas está a una temperatura elevada, mientras que la otra se mantiene a temperatura ambiente y ambas contienen soluciones de sacarosa con igual concentración. Los comederos están etiquetados con diferentes colores y distribuidos espacialmente para que las abejas asocien su contenido a partir de estas señales visuales. Los abejorros muestran preferencia por alimentarse en el lugar con temperatura normal, tal como se esperaba. Además, continúan mostrando preferencia por la ubicación incluso cuando se quita la recompensa previamente asociada a la temperatura más baja, lo que demuestra memoria asociativa. En una segunda fase del experimento, se presenta una solución de sacarosa más concentrada en la ubicación con temperatura desagradable, mientras que en la otra la recompensa es muy inferior. Sorprendentemente, los abejorros eligen alimentarse en la ubicación incómoda, tolerando el estímulo aversivo del calor a cambio de una recompensa mayor. Incluso cuando se eliminan ambos estímulos (el calor y la sacarosa), los insectos siguen prefiriendo el lugar que les había reportado mayor beneficio. Este comportamiento demuestra que los abejorros no responden a estímulos de manera refleja, sino que son capaces de compensar motivaciones opuestas, como placer y malestar, basándose en la memoria de experiencias previas. Tal grado de flexibilidad sugiere un nivel de procesamiento cognitivo complejo, incompatible con una respuesta automática simple al dolor o a la recompensa.
● Criterio 6- Autoatención: La presencia de comportamientos de autoprotección, como el acicalamiento, el frotamiento o la atención dirigida hacia zonas específicas del cuerpo que han recibido un estímulo nocivo, es un indicador relevante de la posibilidad de experimentar dolor. Estas conductas sugieren que el organismo no solo detecta el daño, sino que además actúa de forma localizada para aliviar o mitigar el malestar, lo que implica una evaluación interna del estado corporal (Gibbons et al., 2022a; Crump et al., 2023). En abejorros, por ejemplo, se ha documentado un aumento significativo del acicalamiento de las antenas tras haber sido expuestas a calor, lo que indica una forma de autoatención específica al sitio afectado (Gibbons et al., 2024). Este tipo de comportamiento también se ha observado en orugas en estadios tardíos de desarrollo (Walters et al., 2001). Otro comportamiento relacionado es la autotomía, es decir, la eliminación voluntaria de apéndices dañados, conducta que se ha registrado en diversos insectos como chinches, grillos y bichos palo. Este mecanismo, si bien cumple una función adaptativa de escape o defensa, también ha sido interpretado como una posible manifestación de sensibilidad al dolor, ya que implica una respuesta localizada y específica ante el daño (Emberts et al., 2020; Elwood, 2023).
● Criterio 7- Aprendizaje asociativo: Un criterio importante para evaluar la sintiencia es la capacidad del animal para aprender por asociación en contextos que involucran estímulos nocivos. Esto ocurre cuando el organismo es capaz de vincular un estímulo neutro con una experiencia dolorosa o cuando aprende a evitar activamente estímulos dañinos (Pitman et al., 2009). Este tipo de aprendizaje asociativo supera el condicionamiento clásico o pavloviano, en el que un estímulo condicionado se presenta repetidamente junto a un estímulo incondicionado. Diversos estudios han documentado este tipo de comportamientos en insectos (Elwood, 2023). Por ejemplo, las abejas que han sobrevivido a un ataque de una araña cangrejo automatizada desarrollan un patrón de vigilancia intensificada: antes de posarse sobre una flor, inspeccionan visualmente el entorno con mayor detenimiento. Este comportamiento, que se puede considerar como una forma de ansiedad o incluso como un estado similar al trastorno de estrés postraumático en humanos, reduce significativamente la probabilidad de ser predadas (Ings & Chittka, 2008). Las larvas de mariposa también muestran capacidad de aprendizaje que persiste hasta la etapa adulta, lo que implica una transferencia de memoria a través de la metamorfosis (Blackiston et al., 2008). En langostas, se ha observado aprendizaje instrumental: aprenden a retirar una pata del agua para evitar una descarga eléctrica, o a mover una extremidad específica para impedir que se les administre un estímulo nocivo en la cabeza (Punzo, 1980). Incluso insectos decapitados pueden mostrar cierto grado de aprendizaje, siempre que conserven intactos sus ganglios protorácicos (Harris & Eisenstein, 1999). Un hallazgo particularmente llamativo se ha observado en adultos de D. melanogaster con patas amputadas, quienes tienden a evitar superficies calientes con mayor frecuencia que los individuos no lesionados. Este comportamiento ha sido interpretado como análogo a la alodinia en mamíferos, es decir, la percepción de dolor ante un estímulo que normalmente no lo provoca (Khuong et al., 2019).
● Criterio 8- Preferencia por la analgesia: Este criterio evalúa si un animal demuestra valorar los compuestos analgésicos o anestésicos en contextos de daño físico. Se considera que un animal muestra preferencia por la analgesia cuando, al estar lesionado, cumple con una o más de las siguientes condiciones: a) aprende a autoadministrarse analgésicos o anestésicos, b) prefiere ubicarse en entornos donde puede acceder a dichos compuestos, y c) prioriza la obtención de estos compuestos por encima de otras necesidades, como la alimentación (Gibbons et al., 2022a). Hasta el momento, solo se ha llevado a cabo un estudio orientado específicamente a evaluar este criterio en insectos, y los resultados no fueron concluyentes (Groening et al., 2017).
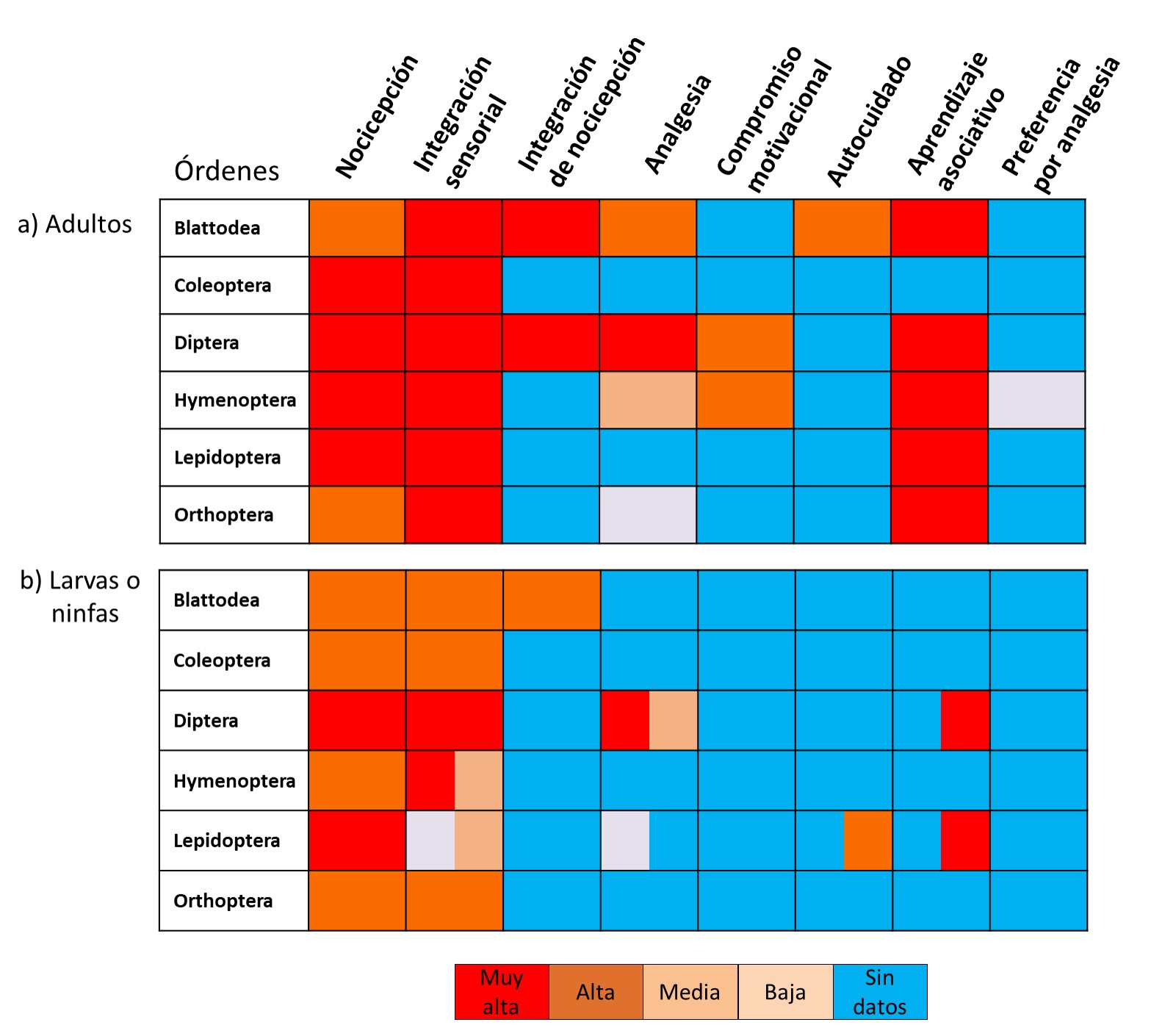
Figura 1.
Nivel de confianza para asegurar el cumplimiento de diversos criterios neurobiológicos que indican percepción de dolor de acuerdo a evidencias obtenidas de distintos órdenes de insectos.
a. En adultos. b. En larvas o ninfas. En el panel de inmaduros, celdas con dos colores indican distinto nivel de confianza para primeros estadios larvales (izquierda) o estadios larvales tardíos (derecha) (Modificada de Gibbons et al., 2022a)
Gibbons et al., 2022aLa Figura 1a muestra la probabilidad de cumplimiento de cada uno de los ocho criterios mencionados para evaluar la sintiencia en insectos adultos de distintos órdenes, según la evidencia actualmente disponible. La Figura 1b muestra esa misma probabilidad, pero en estados inmaduros. Entre los adultos, moscas y cucarachas cumplen seis de los ocho criterios (75 %). Por otro lado, las abejas, avispas y hormigas (Hymenoptera) cumplen cuatro criterios (50 %), las mariposas y polillas (Lepidoptera) y los grillos y saltamontes (Orthoptera) cumplen tres (40 %), mientras que los escarabajos (Coleoptera) cumplen apenas dos (25 %) (Fig. 1a). Debería quedar muy claro en este punto que los porcentajes en que se cumplen los criterios en cada orden no indican el porcentaje en que ese orden es sintiente. En cambio, la mención de porcentajes es simplemente para identificar en qué magnitud sería necesario aplicar medidas de precaución. Es importante señalar que la evidencia disponible proviene de un número limitado de especies dentro de cada orden, por lo que estas estimaciones deben interpretarse con cautela (Crump et al., 2023). En el caso de organismos inmaduros, la evidencia es escasa. En Diptera y Lepidopera se cumplen el 63 y 50 % de los ocho criterios; mientras que Blattodea (38 %), Coleoptera, Hymenoptera y Orthoptera (25 %) también cumplen algunos criterios de sintiencia (Fig. 1b). Es importante destacar que en el caso de Hymenoptera la mayoría de los estudios se han realizado en larvas de abejas. Dado que estas larvas son altamente dependientes y pasivas, sus respuestas podrían no representar adecuadamente el comportamiento de otros himenópteros inmaduros que, en condiciones naturales, requieren una mayor capacidad de toma de decisiones y respuestas activas ante estímulos.
2. Comportamientos complejos
El debate sobre si un organismo merece o no un trato ético suele centrarse en sus capacidades cognitivas. Algunos autores han afirmado que estas habilidades son requisitos previos para la sintiencia y la percepción del dolor (Dawkins, 2006; Elwood, 2011, 2023). Es razonable suponer que la evolución del dolor requiere ciertas funciones cognitivas, como la memoria y el aprendizaje, ya que el dolor cumple la función adaptativa de motivar al individuo a evitar daños futuros o mitigar los ya sufridos, aumentando así sus probabilidades de supervivencia (Villamor Iglesias, 2021).
Existen pruebas que demuestran sorprendentes capacidades cognitivas en insectos, especialmente en especies sociales (Chittka, 2022; Czaczkes, 2022; Chittka & Rossi, 2023), pero también en especies solitarias (Nieberding et al., 2018). Se han documentado formas complejas de comunicación —visuales, químicas, auditivas y mecánicas—, así como una variedad de conductas que desafían la visión tradicional de los insectos como seres puramente instintivos. A continuación, mencionaremos sólo algunos comportamientos poco conocidos o particularmente notables observados en insectos. Algunos trabajos ofrecen evidencia general o específica de ciertos grupos de insectos (Giurfa, 2013, 2015; Perry et al., 2017; Mikhalevich & Powell, 2020; Czaczkes, 2022; Gibbons, 2022a; Traniello & Avergués Weber, 2023; Wen et al., 2024a).
Aprendizaje social y transmisión cultural en insectos. Este tipo de aprendizaje ocurre cuando un individuo adquiere información al observar a un congénere (Leadbeater & Chittka, 2007). En insectos, hay evidencia de que el aprendizaje social está presente en contextos como la selección de sitios para alimentarse, oviponer o evitar depredadores (Danchin et al., 2018). La danza de las abejas, por ejemplo, constituye un lenguaje simbólico que permite a una obrera transmitir información sobre la ubicación de una fuente de alimento mediante movimientos precisos en la oscuridad de la colmena, combinados con señales químicas durante la trofalaxis. Aunque durante mucho tiempo se consideró una conducta innata, recientemente se ha comprobado que, al menos parte de esta habilidad, se adquiere por aprendizaje social (Chittka & Rossi, 2023; Dong et al., 2023). Vale mencionar que, para algunos autores, la mera capacidad de compartir información sobre la ubicación de las fuentes de alimento con sus conespecíficos y que éstos sean capaces de seleccionar a cuáles acudir en consecuencia, demuestra capacidades cognitivas equivalentes al planeamiento y cumplimiento de objetivos y deseos, consistentes con la sintiencia (Carruthers, 2007).
En estudios iniciados hace casi una década, investigadores lograron observar la transmisión cultural de habilidades individuales en colonias experimentales de Bombus terrestris (Alem et al., 2016). De toda la población, sólo dos de cien individuos aprendieron a resolver un problema concreto: tirar de hilos para acceder a una flor artificial exhibida bajo un plato transparente y conseguir una recompensa alimenticia. El resto de la colonia adquirió esta conducta al observarlos directamente o a través de una barrera de vidrio. Los aprendices se convirtieron luego en instructores, manteniendo así la conducta a través de generaciones (Alem et al., 2016; Danchin et al., 2024; Wen et al., 2024b).
Insectos solitarios como D. melanogaster también muestran comportamientos de transmisión cultural. En experimentos que generan dos fenotipos de machos diferentes, se pudo demostrar que las hembras son capaces de identificar y preferir para la cópula a machos que previamente habían observado copulando, estableciéndose así una tradición conductual que perdura en la población (Danchin et al., 2018).
Manipulación de objetos. Algunos insectos emplean herramientas de su entorno con fines distintos a la construcción de nidos (Pierce, 1986; Wen et al., 2024a). Por ejemplo, abejas que untan excremento de gallina alrededor de la colmena para repeler avispas (Mattila et al., 2020), hormigas que utilizan granos de arena para tapizar superficies tóxicas y así poder transitar sobre ellas (Wen et al., 2022), avispas solitarias que colocan cuerpos de hormigas muertas en sus nidos como defensa contra depredadores (Staab et al., 2014), o chinches predadoras que untan sus patas con látex pegajoso para aumentar el éxito al capturar presas (Chen et al., 2022).
Reconocimiento de conespecíficos. Algunas especies de avispas semisociales en el género Polistes Latreille (Hymenoptera: Vespidae) reconocen a sus congéneres y ajustan su nivel de agresividad en función de la "personalidad" del otro. En arenas experimentales se observó que, luego de observar contiendas entre individuos, las avispas son capaces de recordar a los que son particularmente agresivos y mostrarse sumisas frente a ellos (Simons & Tibbets, 2019; Tibbetts et al., 2021).
Aprendizaje inverso. El aprendizaje inverso es la capacidad de modificar el comportamiento para obtener una recompensa cuando cambian las reglas, es decir la capacidad de reaprendizaje. Esta habilidad es prueba de flexibilidad cognitiva y se ha observado en varios grupos de insectos, tales como larvas de D. melanogaster (Foley, 2017; Mancini et al., 2019), adultos de abejorros (Strang & Sherry, 2014), mariposas (Rodrigues et al., 2010), abejas (Pérez Claudio et al., 2018) y hormigas (Sanabria et al., 2024).
Atención médica. Se sabe que algunas hormigas cuidan de sus compañeras heridas: limpian cuidadosamente las lesiones y aplican secreciones glandulares con propiedades antibióticas. Además, si la herida es una pata, son capaces de discriminar el tratamiento y, si la lesión se encuentra en el fémur, realizan amputaciones que reducen la mortalidad por infección (Frank et al., 2023, 2024).
Comportamiento lúdico. Recientemente se ha comprobado que los abejorros juegan con bolitas de plástico en condiciones experimentales (Dona et al., 2022). Para que se considere comportamiento lúdico, la actividad debe realizarse en ausencia de estrés, no implicar recompensa directa ni mejora del fitness, y presentar variabilidad entre ejecuciones. Los abejorros cumplen con todos estos criterios, y, al igual que en vertebrados, los individuos jóvenes tienden a jugar más que los adultos (Wen et al., 2024b). Aunque es raro que se reporten comportamientos lúdicos en insectos, es probable que sean más frecuentes de lo que creemos (Zylinski, 2015).
3. Estados emocionales
En los últimos años ha crecido notablemente el interés por identificar estados emocionales en insectos (Bateson et al., 2011; Baracchi et al., 2017; Perry & Baciadonna, 2017). Estos estados se definen como procesos que integran componentes fisiológicos, neuronales, conductuales y subjetivos (Jirkof et al., 2019), y pueden inferirse experimentalmente a partir de respuestas ante estímulos ambiguos —aquellos que se sitúan entre dos estímulos previamente aprendidos como positivos o negativos— (Baracchi et al., 2017). La variación intraespecífica en estas respuestas sugiere la existencia de “personalidades” entre congéneres, con individuos más o menos optimistas, activos o retraídos, ansiosos o tranquilos (Mollá-Albaladejo & Sánchez-Alcañiz, 2021).
Estos estados afectivos se han asociado con la acción de neurotransmisores homólogos a los implicados en emociones en vertebrados, tales como la dopamina, la octopamina, la serotonina, el neuropéptido F (NPF) y el ácido gamma-aminobutírico (GABA) (Perry & Baciadonna, 2017). Estudios realizados en múltiples especies como abejas, moscas, grillos, hormigas, langostas y abejorros, han demostrado que estos compuestos modulan conductas complejas como agresión, memoria espacial, sesgo cognitivo negativo, expresión emocional ante recompensas inesperadas o división de tareas sociales (Tabla I). La diversidad de paradigmas empleados y la consistencia de los hallazgos refuerzan la idea de que los insectos poseen estados internos persistentes, compatibles con una forma básica de experiencia subjetiva.
Barreras para la empatía hacia los insectos
Las actitudes humanas hacia los animales son sumamente variables, tanto a nivel individual como colectivo. Factores como la cercanía filogenética con los humanos, las capacidades cognitivas y conductuales, la belleza percibida, la utilidad para las personas o una apariencia de vulnerabilidad tienden a generar una actitud negativa o positiva hacia ciertas especies (Serpell, 2004; Mather, 2019).
En el caso de los insectos, nuestras actitudes suelen estar guiadas por dos dimensiones principales: la emocional y la utilitaria (Kellert, 1993). Considerando pocas excepciones, tales como el caso de las mariposas por su belleza y las abejas por su utilidad (Mather, 2019), muchos grupos de insectos se ubican en el extremo negativo de ambas dimensiones: provocan rechazo, miedo o asco en el plano emocional, y en el plano utilitario suelen percibirse como perjudiciales o sin valor (Fig. 2). Las diferencias entre insectos y humanos son evidentes, y aunque puedan parecer obvias, conviene detenerse en algunas de ellas, ya que impactan directamente en nuestra empatía hacia estos animales. Su pequeño tamaño y el ínfimo peso individual, por ejemplo, favorecen una percepción de inferioridad, lo cual puede llevarnos a subestimar su complejidad neurológica. Cabe hacer aquí un paréntesis para señalar la enorme heterogeneidad de este grupo: en cuanto al tamaño corporal, por ejemplo, un bicho palo de 33 cm es unas 2500 veces más largo que un microhimenóptero que mide solo 0,13 mm (Flindt, 2006).
A menudo se ha argumentado que el reducido tamaño, el escaso número de neuronas y la aparente simplicidad del sistema nervioso de los insectos son barreras para el desarrollo de conciencia (Eiseman et al., 1984). Sin embargo, algunos insectos superan en peso a ciertos mamíferos pequeños, y sus cerebros contienen un número no mucho menor de neuronas que el de algunos reptiles (Makarova et al., 2021).
La corta duración de la vida de muchos insectos también tiende a disminuir nuestra valoración de su experiencia subjetiva. Existen casos extremos, como el de los adultos de ciertos Ephemeroptera, cuya vida se limita a unos cinco minutos (Sweeney & Vannote, 1982), o el de algunas polillas que viven apenas 24 horas. No obstante, en el otro extremo se encuentran hormigas reina que pueden superar los 30 años de vida (Porter & Jorgensen, 1988), así como isópteros con longevidades similares, que incluso mantienen parejas estables (Carey, 2001; Kartsev, 2014).
Otra razón que contribuye a la escasa empatía hacia los insectos es su enorme capacidad reproductiva, que tiende a diluir el valor percibido de cada individuo. Por ejemplo, la reina de Termes bellicosus (Smeathman) (Isoptera: Termitidae) puede poner unos 30.000 huevos al día; si consideramos una esperanza de vida de 10 años, eso representa cerca de 110 millones de huevos (Fenton, 1952). Incluso entre insectos solitarios, la fecundidad puede ser asombrosa: una pareja de moscas domésticas, en condiciones ideales y sin enemigos naturales, podría originar una descendencia estimada en 10¹⁴ individuos durante su vida (Weidhaas & Labrecque, 1970). Estos aspectos vuelven a los insectos vulnerables a sesgos humanos que nos instan a adjudicarles un valor inversamente proporcional a su cantidad. En otras palabras, tendemos a infravalorar lo que abunda en demasía y a sobrevalorar los bienes escasos. En el caso de los insectos, la ecuación entre su tamaño casi insignificante (que además nos induce a experimentar una condición de superioridad y de poder ilimitado sobre ellos) y su enorme tasa reproductiva, podrían operar como obstáculos que atentan incluso contra la voluntad de someter a discusión la cuestión sobre su status o relevancia moral.
La morfología tampoco ayuda a generar empatía. Si bien puede haber insectos que resultan atractivos, sobre todo por su coloración vistosa, la mayoría de ellos posee un exoesqueleto rígido e inexpresivo, que no transmite señales emocionales como lo haría un rostro mamífero. Además, el hecho de que el comportamiento de los insectos se rige principalmente por patrones instintivos, refuerza nuestra percepción de su carácter de autómatas o máquinas sin voluntad.
Por otro lado, la relación evolutiva entre humanos e insectos ha estado marcada por múltiples interacciones, muchas de ellas negativas. Nos pican, nos parasitan y transmiten enfermedades que, en algunos casos, han tenido efectos devastadores sobre poblaciones humanas. Esto ha generado una impronta cultural que nos lleva a asociarlos con miedo, asco o repulsión (Curtis, 2013; Lockwood, 2013). Algunas de estas emociones negativas podrían incluso tener un componente innato, heredado de nuestros ancestros, y manifestarse en nuestro subconsciente: no es casual que los insectos sean protagonistas recurrentes en nuestras pesadillas (Klein, 2011).
Por último, cabe señalar que, por lo general, tendemos a alinear cuestiones ligadas a la consideración moral per se hacia individuos y especies; con las ventajas o desventajas que dicha consideración podría reportarnos, incluso llegando a confundir motivaciones genuinamente morales (el valor moral adjudicado a un organismo vivo como fin en sí mismo, por el sólo hecho de serlo) con la utilidad práctica que pueda llegar a reportarnos, ya sea por su valor en el circuito productivo, por el rol que juega en el ecosistema, etc. En otras palabras, asumimos que nuestra consideración moral estará en el fondo supeditada a una evaluación implícita de las ventajas o desventajas que los insectos pueden reportarnos en términos de nuestras propias metas, de modo tal que nuestro nivel de consideración empática hacia los insectos se topará, en última instancia, con nuestros intereses egoístas, los cuales impedirán que el altruismo sobrepase ciertos límites y atente contra nuestras propias metas adaptativas. Por otra parte, la psicología moral experimental ha estudiado algunos sesgos que afectan nuestras evaluaciones morales y que podríamos extrapolar a nuestras actitudes diferenciales hacia el mundo animal en general y a los insectos en particular. Uno de ellos es el llamado “efecto Knobe”, consistente en adjudicar malas intenciones a agentes que provocaron malos resultados (daños) como efecto colateral de sus acciones (Knobe, 2013). Este sesgo podría llevarnos a juzgar inconscientemente a los insectos como “malos” o “buenos” en función de nuestra evaluación subjetiva de los beneficios o daños que nos causan, como si fueran de algún modo “culpables” de provocar esos eventos negativos, incluso cuando la razón nos indica que, en términos generales, no cabe adjudicar categorías morales a los animales por carecer de libre albedrío -su comportamiento está mucho más sujeto a patrones genéticos pre-determinados cuya finalidad es la autopreservación y no “causarnos daño”. De allí que, por ejemplo, si pudiéramos erradicar mosquitos transmisores de enfermedades endémicas, o insectos venenosos causantes de numerosas muertes humanas, probablemente no dudaríamos en provocar su extinción; mientras que tendemos a adjudicar estatus y dignidad moral per se a aquellos insectos (como las abejas) que nos reportan grandes beneficios directos o indirectos.
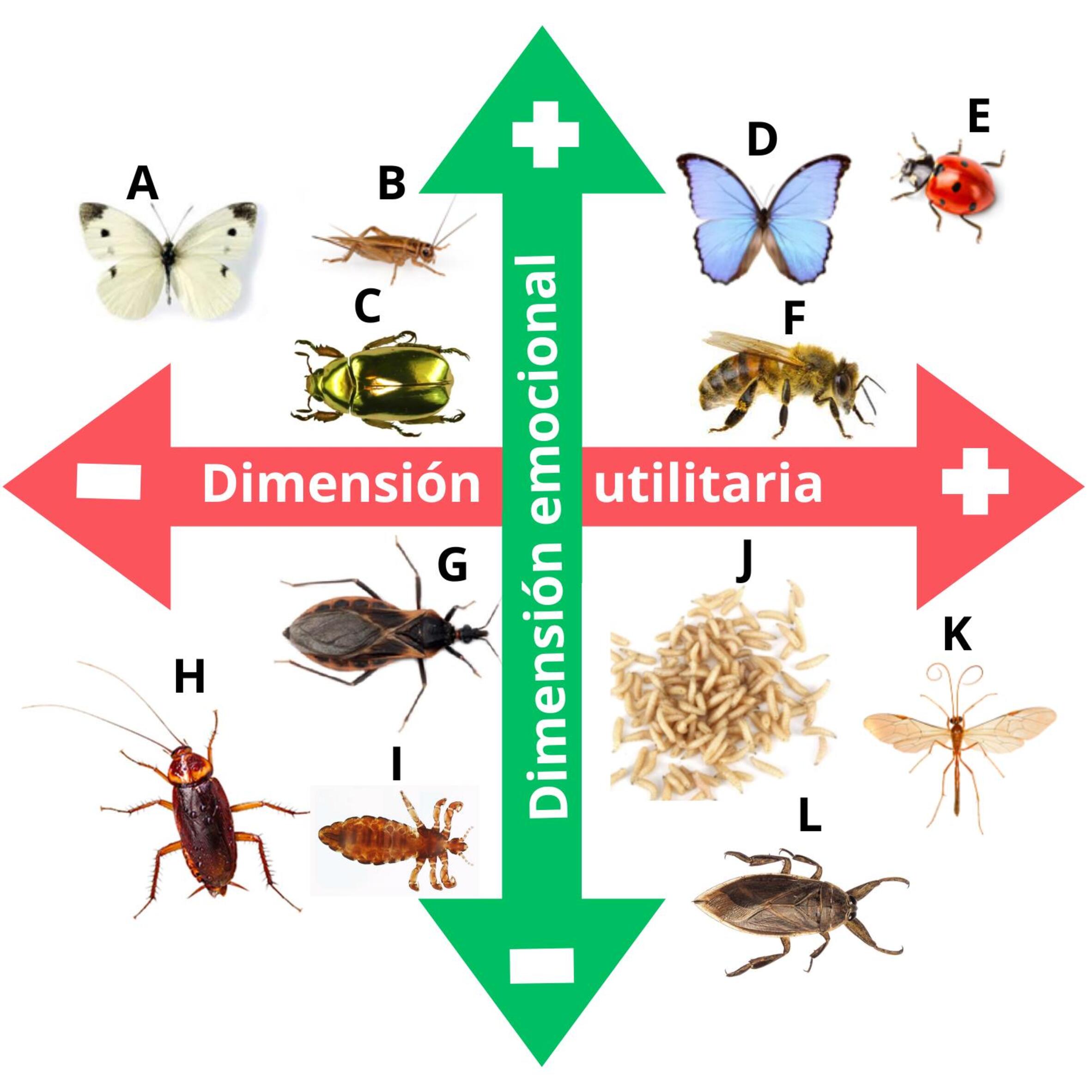
Figura 2
Las dos dimensiones o ejes principales que suelen guiar nuestras actitudes hacia los insectos: eje afectivo-emocional (vertical) y eje utilitario (horizontal), con extremos positivos y negativos
Se presentan ejemplos de insectos que podrían ubicarse en cada cuadrante, seleccionados desde el punto de vista de los autores. Afectivo-emocional positivo / Utilitario negativo (arriba izquierda): A- ciertas especies de mariposas, B- grillos, C- escarabajos, apreciados por su estética o simbolismo, pero percibidos como dañinos para los cultivos. Afectivo-emocional positivo / Utilitario positivo (arriba derecha): D- mariposa monarca, E- vaquita de San Antonio, F- abeja. Despiertan simpatía por su estética, tienen carácter icónico en distintas culturas y ofrecen servicios ecosistémicos (ej. especies bandera en conservación, polinización, control biológico). Afectivo-emocional negativo / Utilitario negativo (abajo izquierda): G- vinchuca, H- cucaracha, piojo. Asociados a enfermedad, suciedad y molestias. Afectivo-emocional negativo / Utilitario positivo (abajo derecha): J- larvas de dípteros K- avispa parasitoide, L- chinche acuática. Baja simpatía pese a su utilidad (ej. larvas saneadoras de heridas, avispa parasitoide usada en control biológico, chinche acuática bioindicadora de calidad de agua (modificada de Serpell, 2004). La dimensión afectivo-emocional se ha abreviado como “emocional” en la figura
Implicaciones éticas para entomólogos
Llegados a este punto, resulta evidente que no podemos afirmar con certeza absoluta que los insectos sean capaces de experimentar dolor, y es posible que nunca alcancemos esa certeza. En este trabajo hemos partido de un examen de las evidencias disponibles sobre la posibilidad de sintiencia y facultades cognitivas complejas en insectos, asumiendo que las mismas proporcionarían una base firme para justificar cierto grado de consideración moral hacia ellos.
Dado que, tal como hemos intentado poner de manifiesto, dichas evidencias son limitadas e interpretables, en esta sección partiremos de la asunción del principio de precaución (PP) como guía general para el desarrollo de una Entomología responsable, comenzando por la exposición y análisis de sus argumentos fundamentales, para luego proponer una serie de aplicaciones prácticas concretas tendientes a la reducción de daños en diversos ámbitos que involucran el uso de insectos. Este marco defendido por Fischer et al. (2025) respalda el enfoque de Birch et al. (2021), que establece que tener alta o muy alta confianza en al menos cinco de los ocho criterios constituye evidencia suficiente para justificar medidas precautorias. Además, los autores ofrecen un argumento histórico condicional: si en 1970 se extendieron protecciones legales a las aves (AWA (Animal Welfare Act), 1970) con evidencia que sólo satisfacía tres de los ocho criterios (y con menor calidad científica que la actual), entonces es coherente adoptar precauciones equivalentes para los insectos hoy, ya que la evidencia actual es más robusta y abundante. Este umbral representa una base suficiente para actuar sin necesidad de certeza absoluta, especialmente cuando los riesgos éticos de omitir acciones pueden ser significativos.
1. El Principio de Precaución
El PP emerge como una herramienta crítica para guiar decisiones éticas cuando nos enfrentamos a la incertidumbre sobre posibles daños derivados de nuestras acciones (Raffensperger & Tickner, 1999; Kriebel et al., 2001). Originalmente aplicado en políticas ambientales —donde se insta a actuar ante amenazas serias "aun con ausencia de certeza científica" (ONU (Organización de las Naciones Unidas), 1992)—, este principio adquiere relevancia paradigmática al trasladarse al debate sobre el bienestar de los insectos. Steel (2015) lo describe como un concepto "influyente pero controvertido", diseñado para promover respuestas proporcionales ante riesgos inciertos sin caer en parálisis por análisis. Propone que una versión coherente del PP debe ser compatible con el uso de evidencia científica, servir como guía bajo incertidumbre y evitar decisiones arbitrarias. Sin embargo, su aplicación en la ciencia enfrenta una paradoja: mientras en la vida cotidiana operamos bajo máximas precautorias ("mejor prevenir que lamentar"), en la experimentación animal persiste un enfoque reactivo que exige evidencia concluyente de dolor antes de implementar salvaguardas éticas (Sunstein, 2002).
En el contexto de los insectos, esta contradicción se agudiza. El paradigma hegemónico —basado en una ética epistemocéntrica— posterga la consideración moral hasta demostrar sintiencia de manera irrefutable, ignorando que la naturaleza subjetiva del dolor hace imposible tal certidumbre (Birch, 2017a, b). Esta resistencia se arraiga en dos temores: (1) que el PP paralice la investigación al imponer restricciones prematuras, y (2) que equiparar invertebrados con vertebrados en derechos éticos implique costos logísticos y económicos insostenibles. No obstante, como señala Birch (2017a, b), esta postura omite que el PP no es binario: admite gradientes interpretativos, desde versiones débiles (que ponderan costos-beneficios) hasta fuertes (que prohíben acciones ante cualquier riesgo potencial). Stewart (2002) sintetiza cuatro aplicaciones prácticas: desde la no exclusión de actividades riesgosas hasta la prohibición absoluta, pasando por el uso de márgenes de seguridad y tecnologías menos invasivas.
La propuesta del Principio de Precaución para la Sintiencia Animal (PPSA) de Birch (2017a, b) adapta este marco a la ética entomológica: cuando exista evidencia plausible —aunque no concluyente— de que un insecto pueda experimentar daño (p. ej., presencia de nociceptores, aprendizaje asociativo o analgesia), se justifica implementar medidas preventivas proporcionales. Este enfoque ha sido profundizado por Browning & Birch (2022), quienes discuten su aplicación tanto en política pública como en ciencia, y subrayan la importancia de combinar evidencia parcial con razonamiento precautorio para fundamentar decisiones éticas. En el ámbito de la investigación científica, este marco adquiere un valor particular: permite fundamentar protocolos éticos incluso ante incertidumbre, apelando a la inferencia de la mejor explicación cuando los animales muestran indicadores como aprendizaje aversivo, evitación dirigida o decisiones costo-beneficio frente al dolor (Birch et al., 2021; Browning & Birch, 2022; Andrews et al., 2024). Esto es especialmente urgente en escenarios de escala masiva, como biofábricas donde billones de individuos son sacrificados anualmente, o en investigaciones que emplean métodos letales para conservar especies (Lövei et al., 2023). Como advierte D'Silva et al. (2024), el principio de precaución no solo debe guiar las decisiones científicas, sino también las prácticas productivas emergentes, garantizando condiciones que reduzcan el sufrimiento potencial. Frente a los extremos —la inacción por incertidumbre o la prohibición categórica—, el PPSA ofrece una vía pragmática: priorizar alternativas no letales, optimizar protocolos de sacrificio y reconocer que, en ausencia de consenso, la carga de la prueba debe recaer en quienes afirman que los insectos no sufren (Crump et al., 2023).
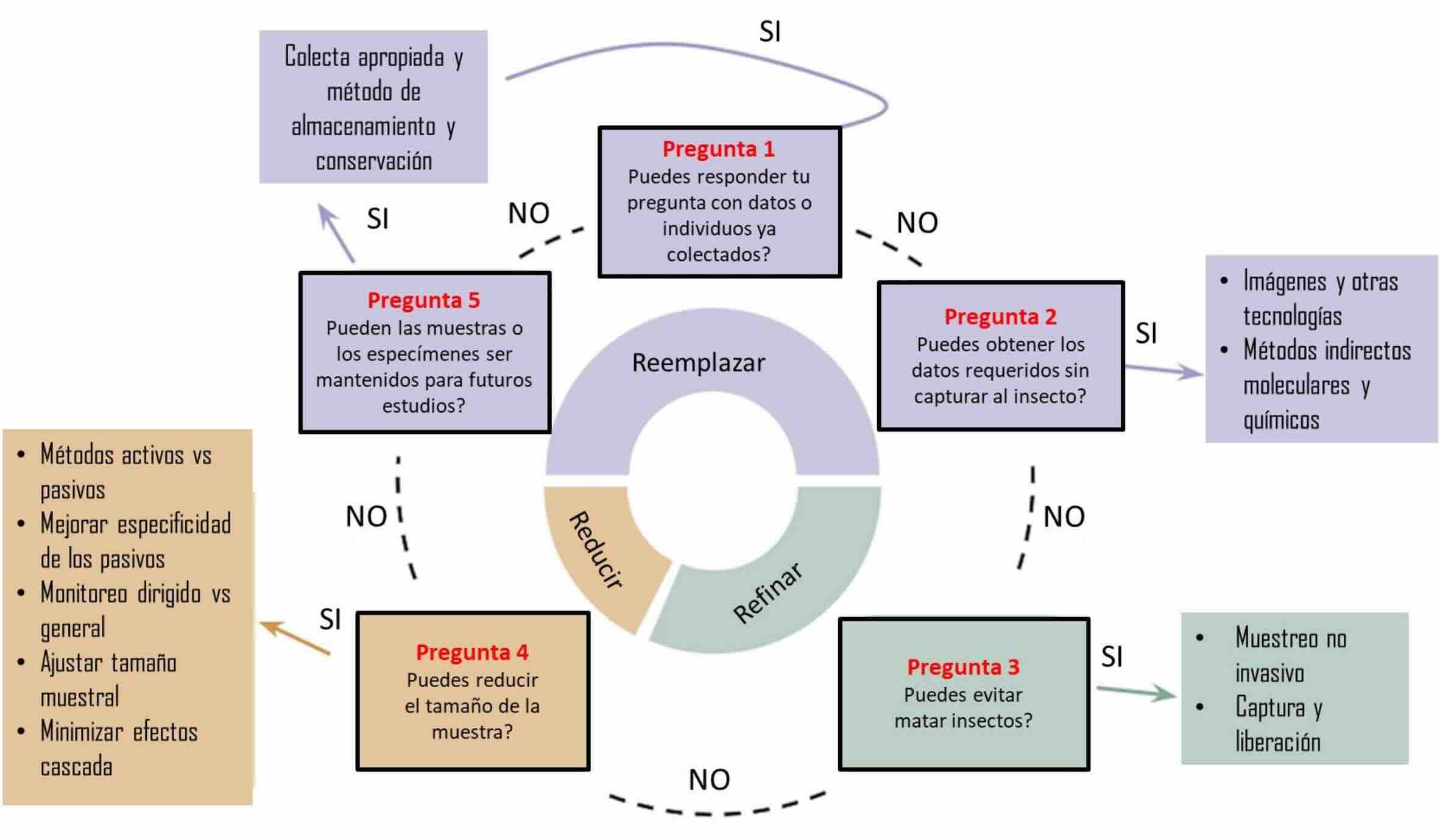
Figura 3
Propuesta de flujo de toma de decisiones para reemplazar, refinar y reducir las capturas de insectos en la investigación biológica, indicando métodos alternativos que pueden ayudarnos a implementar estas mejores prácticas (modificada de Montero-Castaño et al., 2022)
En definitiva, el PP no busca frenar el progreso científico, sino reorientarlo hacia una entomología responsable. Hasta el momento no se ha recabado ninguna evidencia que nos haga presuponer que los insectos no sienten dolor. La precaución no es sinónimo de prohibición, sino de innovación: desarrollar tecnologías que minimicen el daño, repensar la docencia sin colecciones letales y regular industrias desde un enfoque de Cinco Libertades adaptadas (Rowe et al., 2024) (ver punto 4, Producción industrial). La pregunta ya no es si los insectos sienten, sino cómo actuar ante la posibilidad de que lo hagan.
Comenzar a considerar el bienestar de los insectos nos enfrenta no solo a su enorme diversidad biológica, sino también a la pluralidad de contextos en los que interactuamos con ellos. Cada uno de estos contextos, implican dilemas éticos específicos que no pueden abordarse con una única lógica. Cuatro de estos contextos: investigación, educación, producción industrial y control de plagas son abordados a continuación.
2. Investigación
En el ámbito académico, varios autores han señalado una paradoja persistente en estudios de ecología centrados en la conservación de insectos: estos trabajos, cuyo objetivo es preservar especies, emplean métodos de muestreo letales que implican el sacrificio de millones de individuos (Fischer & Larson, 2019;Lövei et al., 2023; Lövei & Ferrante, 2024). Esta aparente contradicción, sacrificar individuos en masa para conservar especies, plantea una pregunta ética crucial: ¿pueden justificarse estos estudios frente al daño que infligen? El Código del Colector de insectos (Trietsch & Deans, 2018) es un excelente artículo que puede ayudar a un debate ético a quienes se inicien en la investigación con insectos. Sería siempre recomendable que docentes y directores responsables de proyectos puedan dialogar con los involucrados sobre los métodos a utilizar para la captura de insectos, integrando aspectos científicos y éticos (Stack Whitney & Whitney, 2024).
En este sentido, el uso de métodos no letales representa una estrategia efectiva para reducir el daño que causamos al estudiar insectos (Barrett & Fischer, 2024). Aunque no siempre es posible implementarlos, cada investigador debería evaluar caso por caso, y tomar decisiones acordes a las preguntas y medios disponibles (Fig. 3). Una de las estrategias más relevantes es aprovechar insectos muertos por otras causas, tanto científicas como no científicas. En estudios de ecología de insectos esto sería particularmente difícil porque es necesaria la estandarización de los diseños de muestreo a fin de que sean replicables. Sin embargo, en otras áreas de la Entomología, no debiera descartarse tal posibilidad. En el ámbito científico, los especímenes colectados pero descartados por no ajustarse a los objetivos originales de un proyecto (conocidos como “bycatch” en inglés) pueden reutilizarse en investigaciones posteriores. Trampas de caída y trampas Malaise suelen generar grandes volúmenes de bycatch (Fischer & Larson, 2019;González et al., 2020). Las trampas activas, que ofrecen algún tipo de atractivo al grupo de insectos de interés, siempre deberían ser preferidas a las pasivas (Montero-Castaño et al., 2022).
Promover la coordinación entre investigadores interesados en reutilizar el bycatch contribuiría significativamente a reducir el número total de insectos colectados (Spears & Ramírez, 2015). La utilización de refugios artificiales que hospeden a los insectos en ciertos hábitats y puedan mostrar su ocurrencia sin tener que matarlos, también es interesante (Salman, 2020). Asimismo, insectos que mueren de forma no intencional, por ejemplo, muertos por el tráfico vehicular en rutas, pueden ser aprovechados para la investigación (Russo, 2025).
Las colecciones entomológicas también constituyen una valiosa fuente de información. Su creciente digitalización facilita el acceso a datos y especímenes sin necesidad de nuevas capturas (Short et al., 2018; Svenningsen & Schingel, 2024). A pesar de que muchas publicaciones científicas exigen conservar especímenes de referencia en colecciones institucionales, esta práctica no siempre se cumple, y numerosos insectos colectados terminan deteriorándose o están inaccesibles para la comunidad científica.
Un gran número de tecnologías emergentes reemplazan a técnicas de muestreo letales para el estudio de insectos (van Klink et al., 2022; Fig. 4). El limitado conocimiento taxonómico de algunos grupos y la falta de fondos para la investigación científica son limitantes para su empleo en nuestro país y en otros países del Neotrópico. Las cámaras trampa permiten la detección e identificación de insectos sin necesidad de capturarlos, sobre todo cuando se integran con sistemas de inteligencia artificial (Costello et al., 2016; Bjerge et al., 2023; Zeuss et al., 2024; Prudic, 2024; Roy et al., 2024) y los sistemas de muestreo automatizado por sonido han demostrado su eficacia para identificar especies de insectos con gran precisión (Kohlberg et al., 2024). Los radares por su parte, pueden detectar grandes enjambres de insectos, y proporcionar información detallada sobre insectos voladores, incluyendo su tamaño, forma, velocidad y trayectoria (Chapman et al., 2011; Rhodes et al., 2022).
En la última década, se han desarrollado métodos genéticos no letales que permiten obtener ADN sin sacrificar al ejemplar (Chua et al., 2023). Así, por ejemplo, se analiza material genético de insectos colectados en capullos pupales (Ožana et al., 2020), de partes seccionadas del cuerpo, como alas o patas (Keyghobadi et al., 2021), o de muestras tomadas del ambiente, ya sea suelo, agua o aire (Roger et al., 2022; Bierman & Lloyd, 2024; Weber et al., 2024). En contextos agrícolas, es posible estimar mediciones de procesos ecológicos como la depredación, observando sus indicios en presas centinela, tales como orugas de plastilina (Ferrante et al., 2024), sin que sea necesaria la determinación de abundancia de predadores mediante colectas de insectos.
Si tenemos en cuenta las inquietudes expresadas por diversos técnicos e investigadores, así como lo que señala la bibliografía, podríamos situar a las técnicas no letales en el lugar más respetuoso desde el punto de vista ético. Les seguirían métodos como el uso de redes de arrastre, donde los insectos colectados pueden ser sacrificados por enfriamiento y posterior congelación. Las trampas de caída causan una muerte bastante rápida, siempre que el conservante se coloque en concentraciones adecuadas y no se diluya con la lluvia. En el extremo menos ético quedarían las trampas adhesivas, que provocan una muerte lenta y traumática por inanición o aplastamiento, y que deberían utilizarse como último recurso (Stack Whitney & Whitney; 2024).
Finalmente cabe mencionar que la ciencia abierta, concebida como el conocimiento accesible, compartido y desarrollado a través de redes colaborativas (Wilkinson et al., 2016; Vicente-Sáez & Martínez-Fuentes, 2018), ofrece la oportunidad de disminuir el daño a los insectos en investigación. Facilitar el acceso a datos previos y promover la reutilización de información, puede reducir la necesidad de nuevas colectas (Cuff et al., 2024). Gracias a las técnicas de metaanálisis, que son posibles cuando los datos están disponibles para quien los requiera (Wittman & Aukema, 2020), es posible responder diversas preguntas en el campo de la Entomología.
Los entomólogos también podemos intervenir en decisiones relacionadas con la experimentación con insectos. A medida que se acumula evidencia sobre la complejidad conductual, neurofisiológica y afectiva de los insectos, se vuelve cada vez más difícil justificar su exclusión automática de los marcos éticos que regulan el uso de animales en ciencia. Freelance (2019) sostiene que, frente a la incertidumbre sobre su sintiencia, pero con indicios crecientes de su posibilidad, es necesario adoptar un enfoque ético precautorio. En lugar de esperar certeza absoluta, propone medidas mínimas de respeto y protección que incluyan diseños experimentales responsables, aplicación flexible de los principios de reemplazo, reducción y refinamiento (3Rs), y la minimización del daño durante la experimentación.
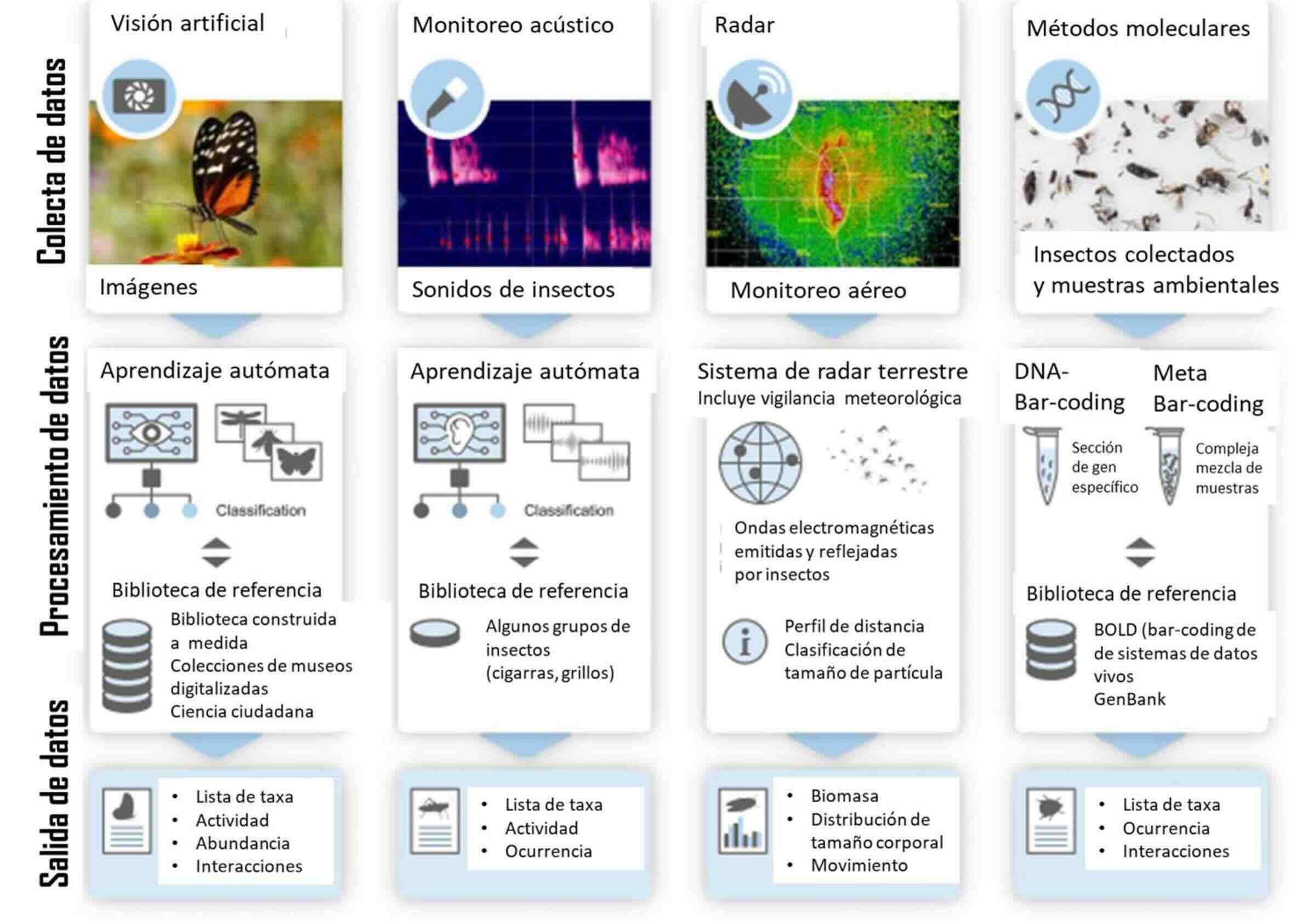
Figura 4.
Colecta, procesamiento y variables obtenidas en cuatro tecnologías emergentes que pueden ser utilizadas para evitar el uso de métodos no letales en Entomología: visión artificial, monitoreo acústico, radar y métodos moleculares (modificada de van Klink et al., 2022)
Este enfoque se complementa con el marco ampliado de los 12 Rs propuesto por Brink & Lewis (2023), que integra aspectos de bienestar animal, valores sociales e integridad científica. Esta propuesta expande los 3Rs clásicos para incluir principios como respeto, responsabilidad, regulación, reproducibilidad, relevancia y transferibilidad; y enfatiza la necesidad de una ciencia éticamente robusta, culturalmente sensible y científicamente rigurosa. Ambos enfoques convergen en un punto esencial: la necesidad de actuar éticamente incluso bajo condiciones de incertidumbre, reconociendo la posibilidad de sufrimiento en insectos y la responsabilidad de minimizarlo. Aplicar estos marcos no implica obstaculizar la investigación científica, sino más bien fortalecer su legitimidad, su calidad metodológica y su coherencia con los valores de una ciencia responsable, justa y cuidadosa. Volveremos sobre este tema al hablar de insectos que se crían a escala industrial.
3. Educación
En el ámbito de la docencia superior, muchos cursos de Entomología incluyen la colecta de insectos y la presentación de una colección entomológica como requisito para aprobar. Dichas colectas a menudo son necesarias, ya que permiten a los estudiantes involucrarse en profundidad en los conocimientos, técnicas y prácticas de la Entomología. Sin embargo, a la luz de lo discutido hasta el momento, es previsible que aumente la resistencia por parte de docentes, y especialmente de estudiantes, frente a la realización de estas colectas (Byrne, 2023; Sandhi et al., 2023). Al igual que en otros ámbitos que implican la utilización o sacrificio de seres vivos con fines educativos, sería recomendable incorporar un espacio de reflexión ética en los programas de estudio. Este espacio permitiría analizar críticamente las razones que justifican el uso de material biológico y brindar a los estudiantes la posibilidad de fundamentar sus decisiones en torno al tratamiento de los insectos (Trout et al., 2010; Fore & Barrett, 2024; Barrett et al., 2025).
En este sentido, una alternativa válida consistiría en reemplazar la colecta de ejemplares por imágenes fotográficas obtenidas por los propios estudiantes, ya que los docentes suelen contar con material conservado que permite la observación de detalles morfológicos y la validación de las identificaciones (Fischer & Larson, 2019). Asimismo, actualmente existen numerosos sitios web que ofrecen imágenes de alta calidad y gran nivel de detalle de una amplia variedad de especies, lo que facilita la apreciación de la diversidad morfológica dentro de cada taxón. Entre estas plataformas destaca iNaturalist (Barrett et al., 2023a).
4. Producción industrial
Se calcula que entre 1 y 1.2 billones de insectos se crían anualmente en Europa, con proyecciones de crecimiento sostenido a nivel global, ya que la industria se posiciona como una alternativa más sostenible frente a la ganadería convencional (Rowe, 2020a, b; Lähteenmäki-Uutela et al., 2021;Barrett & Adcock, 2023). En comparación con la ganadería tradicional, la cría de insectos ofrece ventajas significativas: menor costo ambiental, alta eficiencia en la conversión de alimento en biomasa, menores emisiones de gases de efecto invernadero, bajo consumo de agua, posibilidad de cría en espacios reducidos y escaso riesgo de transmisión de patógenos al ser humano (De Goede et al., 2013). Además, el hecho de que algunas especies puedan criarse exitosamente sobre excrementos de pollos, ganado o incluso de humanos ha favorecido el desarrollo de modelos de economía circular (Moruzzo et al., 2021; Cattaneo et al., 2024; Safavi et al., 2024). Entre las especies más importantes cultivadas a escala industrial se encuentran la mosca soldado negra (Hermetia illucens L., Diptera: Stratiomyidae), el gusano de la harina (Tenebrio molitor L., Coleoptera: Tenebrionidae) y diversas especies de grillos, principalmente Acheta domesticus (L.) (Orthoptera: Gryllidae) (Barrett, 2024a).
En la actualidad, ya se sacrifican más insectos que vertebrados terrestres de consumo, principalmente mamíferos y aves, y las proyecciones indican que, de reemplazar entre el 25 % y el 100 % del alimento balanceado utilizado en acuicultura, se requeriría una producción aún mayor (Rowe, 2020a,Fig. 5). Esta tremenda expansión de la cría de insectos con fines industriales conducirá a la utilización de millones, e incluso miles de millones de individuos para alcanzar volúmenes de producción proteica comparables a los sistemas tradicionales. En consecuencia, también serán muy grandes los desafíos que enfrentaremos en términos de bienestar animal (Pali-Schöll et al., 2019; Tiwasing & Pate, 2019; van Huis, 2019; Lambert et al., 2022; Bermúdez-Serrano et al., 2023; Barrett, 2024b).
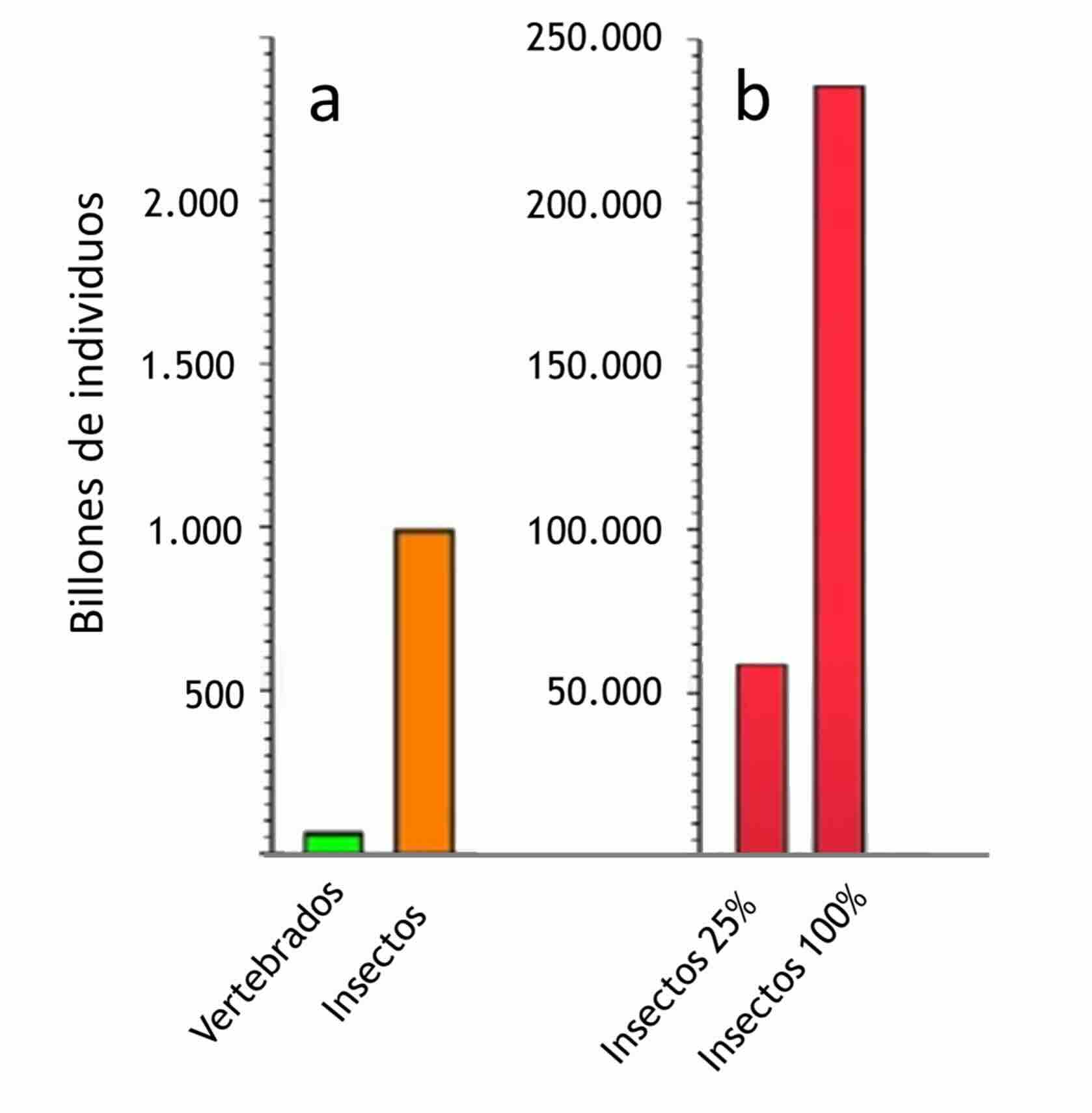
Figura 5
Número de individuos utilizados para alimento humano y/o para alimento de otros animales destinados al consumo humano
a. Datos de 2020. b. Estimado del número de individuos que serían necesarios si se reemplazara un 25 % o un 100 % del total de proteínas actualmente empleadas para alimentar peces, por proteínas obtenidas de insectos (datos de Rowe, 2020a)
El uso masivo de insectos como solución ecológica no debe ignorar su bienestar. Estos organismos aún carecen de protección legal significativa (Magalhães-Sant’Ana, 2009), y podrían reproducirse los mismos errores de la ganadería intensiva, como el hacinamiento, el sufrimiento silencioso o la automatización sin consideración ética (Barrett & Fischer, 2023).
Frente al avance industrial, los entomólogos que participan en el campo de la cría en masa de insectos deben aplicar la responsabilidad ética tanto a nivel individual como colectivo. Como plantean Crespi-Abril & Rubilar (2021), es necesario cuestionar el modo en que se ha “entrenado a los investigadores para desensibilizarse y reconfigurar su vínculo ético con los animales, naturalizando su reificación”. El bienestar de los insectos no puede abordarse con una única regla. Por lo tanto, se propone adaptar el marco de las “Cinco Libertades” (Webster, 1994) al contexto de los insectos, ajustándolo según la especie y la etapa de desarrollo. Este enfoque considera las libertades de: acceso a alimento y agua; comodidad física; ausencia de dolor, lesiones y enfermedades; expresión del comportamiento natural; y ausencia de miedo y angustia. Tales consideraciones resultan especialmente relevantes en insectos holometábolos, debido a las marcadas diferencias entre larvas y adultos (van Huis, 2021; Delvendahl et al., 2022; Barrett & Fischer, 2023; Klobučar & Fischer, 2023).
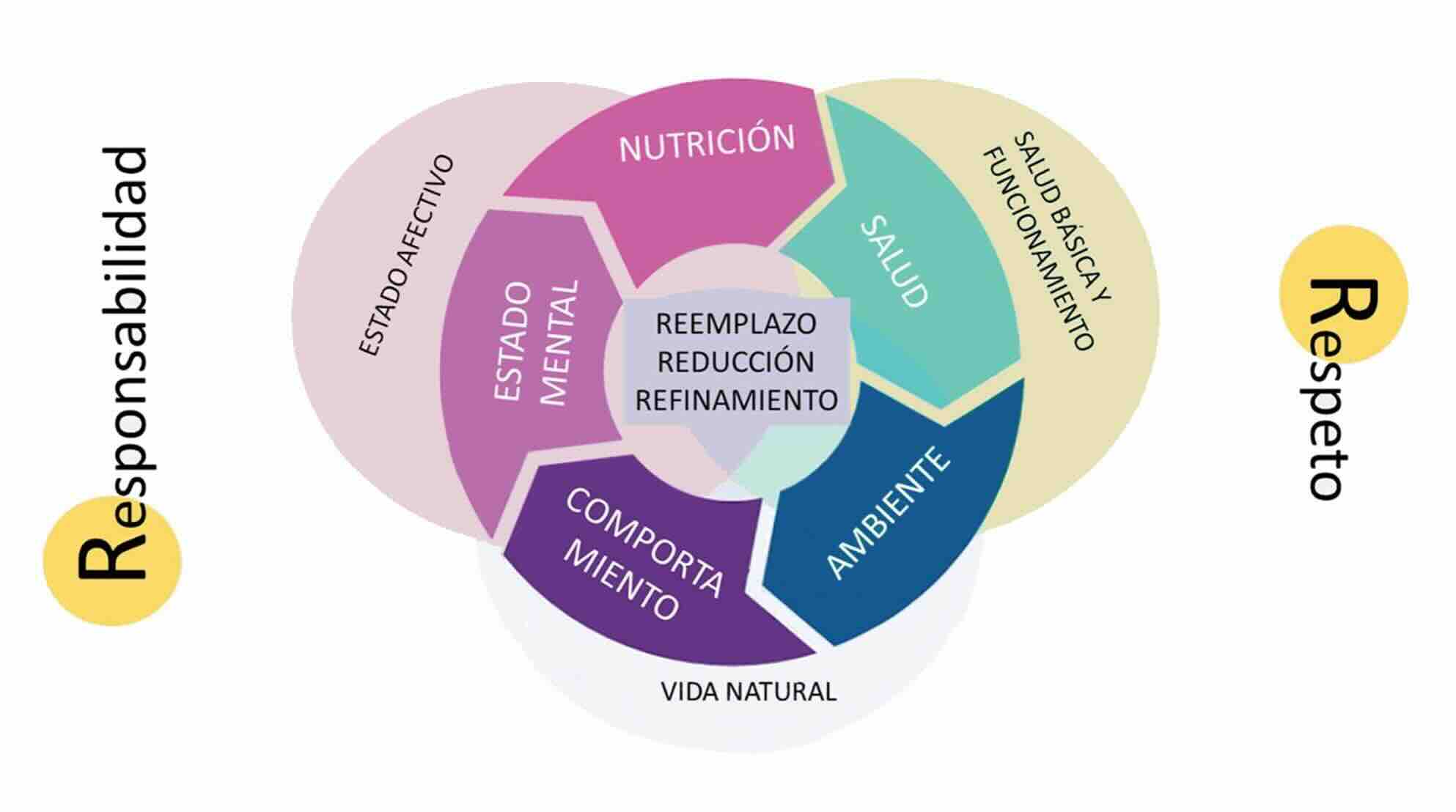
Figura 6
Intersección entre tres marcos conceptuales clave: los cinco dominios del bienestar animal (nutrición, ambiente, salud, comportamiento y estado mental), las tres concepciones filosóficas del bienestar (funcional, afectiva y naturalista), y los principios de acción ética (3Rs clásicas y las ampliadas 5Rs, que incluyen Respeto y Responsabilidad).
Esta superposición permite abordar el bienestar de insectos de forma multidimensional, reconociendo sus necesidades fisiológicas y comportamentales, y guiando la toma de decisiones éticas en contextos de investigación, producción y manejo (modificada de Crespi-Abril & Rubilar, 2024)
En el ámbito del bienestar animal, se han propuesto marcos conceptuales que identifican puntos relevantes a considerar. El postulado fundamental de las 3Rs (reemplazo, reducción y refinamiento) resulta insuficiente como base para la manipulación de insectos. Dicho principio fue ampliado a las 5Rs donde se incorporaron el Respeto y la Responsabilidad (Crespi-Abril & Rubilar, 2021, 2024). Sin embargo, otras propuestas indican que es necesario, en realidad, promover estados positivos que consideren concepciones funcionales (o fisiológicas vitales), afectivas (o estados mentales) y ambientales (desafíos del medio). Realmente, todos los enfoques propuestos que apuntan al bienestar de los individuos pueden ser unificados en un marco conceptual anidado (Fig. 6). Esto evidencia la complejidad de factores que deben ser considerados a la hora de trabajar responsablemente con cualquier grupo de animales.
En muchos casos, las biofábricas operan sin tener en cuenta el comportamiento y las necesidades particulares de cada especie. Sin embargo, está aumentando el número de trabajos que analizan el bienestar de los insectos en las biofábricas (Bear, 2021). Existen excelentes revisiones sobre grillos (Rowe et al., 2024), mosca soldado (Hermetia illucens, Barrett et al., 2023b), gusanos de seda (Bombyx mori L.,Rowe, 2021), gusano de la harina (Tenebrio molitor, Barrett et al., 2024a) y abeja melífera (Apis mellifera L., Garrido & Nanetti, 2019; Schukraft, 2019d; Formato et al., 2024), entre otras. Tomando el caso de Hermetia illucens como ejemplo, basta mencionar que persiste la afirmación errónea de que los adultos no se alimentan, a pesar de las evidencias que demuestran sus preferencias dietarias que afectan su reproducción y longevidad (Tomberlin et al., 2002; Nakamura et al., 2016; Ramano et al., 2020). Desafortunadamente para los insectos, la perpetuación de la colonia es posible aunque los adultos no se alimenten, y por cuestiones logísticas o de simple comodidad, raramente se les ofrece alimento en este estado (Barrett et al., 2023b), y los insectos mueren de hambre luego de una corta vida. Además, las altas densidades poblacionales pueden inducir comportamientos agresivos y canibalismo, lo que plantea serias dudas éticas (Erens et al., 2012).
5. Plagas
Los insectos afectan negativamente la vida humana de múltiples maneras, principalmente al competir con nosotros por los alimentos que producimos y al transmitir enfermedades (WHO (World Health Organization), 2020; Belluco et al., 2023; Grimaldi, 2023). Se consideran las plagas más perjudiciales y los esfuerzos por controlar sus poblaciones son intensos y sostenidos en el tiempo. La amplitud del tema del control de plagas nos obliga a ser concisos y a adoptar una perspectiva general para reflexionar éticamente sobre sus implicaciones. Empleamos una amplia variedad de estrategias, desde el uso de insecticidas convencionales hasta las formas más sofisticadas de manipulación genética. Esto plantea una pregunta central: ¿tenemos alguna obligación moral hacia los insectos que intentamos eliminar? Existe un continuo ético: cuanto mayor la escala del control, es decir cuando se apunta a la erradicación completa de la especie y no simplemente a la reducción numérica de las poblaciones perjudiciales, mayores son las implicancias morales (Draney, 1997). Técnicas como los métodos autocidas aplicados a gran escala geográfica y las estrategias de edición genética dirigidas mediante CRISPR (“gene drive”), podrían llevar a la extinción local o incluso global de especies que consideramos indeseables (Gantz & Akbari, 2018; Kaebnick et al., 2025). Estas formas de control de plagas plantean riesgos éticos que se discuten poco a pesar de su creciente desarrollo y aplicación. El control químico, por su parte, implica la disponibilidad de cientos de productos al alcance de cualquier usuario, con efectos colaterales repetidamente documentados sobre organismos no blanco, incluidos los seres humanos (Tostado & Bollmohr, 2022). Desde la perspectiva de la sintiencia de los insectos como individuos, tema en el que se centra el presente artículo, es fundamental que los entomólogos que diseñan o respaldan medidas de control se formulen críticamente las siguientes preguntas: 1- Existe un método no letal para protegernos del insecto? 2- Existe un método menos dañino que otro para lograr los mismos objetivos? (Durrant, 2024). Al hablar de menos dañino nos referimos al modo en que el insecto muere, la rapidez y la eficiencia del método a emplear, aspectos que serán abordados en la siguiente sección.
Formas responsables de sacrificar insectos
Cada vez que nos enfrentemos a la decisión de sacrificar un insecto, deberíamos detenernos a reflexionar sobre el modo en que lo haremos. El sacrificio de insectos raramente se hace con el objetivo de evitar sufrimiento. En la mayoría de los casos, se hace por conveniencia, sin cumplir con los criterios de la eutanasia ética, que implica un acto compasivo y considerado (Shelomi, 2021). Es importante destacar que no todos los métodos de anestesia y eutanasia son adecuados para todas las especies y que los estudios al respecto son limitados. Sin embargo, hay un principio que debería ser universal: la muerte debe ser lo más rápida posible, idealmente en fracciones de segundo, para minimizar cualquier posible sufrimiento. Los métodos más recomendados para sacrificar insectos consisten en una sobredosis de anestésicos o bien deberían incluir dos pasos. En el primer paso, el insecto es anestesiado y en el segundo, que debe aplicarse antes de que se recupere de la anestesia, se debe destruir el sistema nervioso, lo que garantiza la pérdida irreversible de la conciencia (AVMA (American Veterinary Medical Association), 2020; Heniff et al., 2023). La anestesia puede lograrse en insectos mediante temperaturas ligeramente inferiores a cero grados, que conduce a coma por congelación, o mediante compuestos volátiles como el dióxido de carbono y el isoflurano, que causan coma narcótico (MacMillan et al., 2017; Cooper et al., 2022). El segundo paso puede lograrse por métodos físicos, mecánicos y químicos. Entre estos métodos se encuentran el frío o calor excesivo, sumergir los insectos en agua hirviendo y la trituración mecánica. Los métodos químicos consisten en inyecciones de cloruro de potasio o ivermectina en el sistema nervioso o circulatorio (posible cuando el insecto tiene gran tamaño), ser sumergidos en diversos compuestos como eugenol, nitrógeno líquido, etc. (Murray, 2022; Zachariah, 2022; Lum & Keller, 2024). El uso de los métodos del segundo paso sin anestesia previa se considera un modo no humanitario para sacrificar insectos (AVMA, 2020). Así, por ejemplo, la muerte por temperaturas extremadamente bajas (freezado) se considera dolorosa debido a la formación de cristales de hielo dentro de los tejidos (AVMA, 2020).
En el contexto de las biofábricas, el dilema moral se intensifica, ya que mueren millones de individuos cada día. Los métodos de sacrificio más comúnmente empleados incluyen la exposición a temperaturas extremas, la inmersión en agua hirviendo o la trituración mecánica. Estas prácticas suelen ir precedidas de un periodo de inanición variable, cuyo objetivo es vaciar el tracto digestivo de los insectos antes del sacrificio. No obstante, determinar la duración adecuada de este ayuno es un reto, ya que debe ser lo suficientemente prolongado para cumplir su función, pero sin llegar a provocar comportamiento caníbal entre los individuos, lo que implica una necesidad constante de observación y ajuste. El sacrificio de los insectos debe ser lo más eficaz posible, es decir, debe afectar al mayor número posible de individuos de manera inmediata. En este sentido, incluso pequeñas modificaciones en el diseño de los dispositivos de trituración pueden marcar una diferencia significativa en términos de eficacia y velocidad del sacrificio (Barrett et al., 2024b). En la actualidad, los biofabricantes de insectos operan en una zona gris en términos legislativos en la mayoría de los países, lo que les otorga un amplio margen de libertad para decidir si adoptan o no prácticas orientadas al bienestar animal (Bear, 2019).
En el caso de que sea necesario eliminar colmenas completas de abejas, ya sea por infestación con parásitos o por representar un riesgo para la salud pública debido a la africanización, debe prestarse especial atención a minimizar el sufrimiento colectivo. Dado que una colonia puede albergar miles de individuos, el método empleado debería inducir una pérdida de conciencia rápida e irreversible, seguida de una muerte igualmente rápida para la mayoría de las abejas (Mutinelli, 2023). En este sentido, el uso de dióxido de azufre se considera el método más recomendable, debido a su rápida acción, bajo costo y facilidad de aplicación (Roy & Vidal-Naquet, 2022).
CONCLUSIONES
Los tiempos cambian y, con ellos, nuestras preguntas, certezas y responsabilidades. Hoy, la ciencia empieza a considerar seriamente la posibilidad de que los insectos puedan tener experiencias subjetivas, por lo que nosotros, como entomólogos, también estamos llamados a repensar la forma en que nos relacionamos con ellos. A pesar del interés que muchos tenemos por las cuestiones éticas, aún no contamos con espacios específicos donde discutir estos temas de forma regular y no siempre estamos adecuadamente preparados para incorporar consideraciones éticas en la toma de decisiones cotidianas. Sin embargo, como especie capaz de anticipar las consecuencias de nuestras acciones, nos corresponde asumir la responsabilidad de velar por el bienestar de los seres vivos con los que compartimos el planeta. No se trata de dejar de investigar, enseñar o manejar insectos, sino de hacerlo con responsabilidad, basada en evidencias y guiada por la ética profesional. Incorporar esta perspectiva puede enriquecernos, generar nuevas preguntas científicas y abrir espacios de diálogo con otros sectores de la sociedad. Tal vez nunca podamos saber con certeza qué sienten los insectos, pero sí existe la posibilidad real de que puedan sufrir, por lo que actuar con cuidado no solo es prudente, sino también profundamente humano.
REFERENCIAS
Adamo, S. A. (2019). Is it pain if it does not hurt? On the unlikelihood of insect pain. The Canadian Entomologist, 151, 685-695. https://doi.org/10.4039/tce.2019.49
Alem, S., Perry, C. J., Zhu, X., Loukola, O. J., Ingraham, T., Søvik, E., & Chittka, L. (2016). Associative mechanisms allow for social learning and cultural transmission of string pulling in an insect. PLoS biology, 14(10), e1002589. https://doi.org/10.1371/journal.pbio.1002589
Amiri, A., & Bandani, A. R. (2024). Encountering a dead or live conspecific affects the behavior and longevity of Callosobruchus. Journal of Stored Products Research, 108, 102387. https://doi.org/10.1016/j.jspr.2024.102387
Andrews, K. (2023). All animals are conscious: Shifting the null hypothesis in consciousness science. Mind & Language, 39(3), 415-433. https://doi.org/10.1111/mila.12498
Andrews, K., Birch, J., & Sebo, J. (2025). Evaluating animal consciousness. Science, 387(6736), 822-824. https://doi.org/10.1126/science.adp4990
Andrews, K., Birch, J., Sebo, J., & Sims, T. (2024). Background to the New York Declaration on Animal Consciousness. https://sites.google.com/nyu.edu/nydeclaration/declaration
AVMA (American Veterinary Medical Association). (2020). Guidelines for the Euthanasia of Animals. https://www.avma.org/sites/default/files/2020-01/2020-Euthanasia-Final-1-17-20.pdf
AWA (Animal Welfare Act). 1970. Public Law. 91-579, United States. https://www.congress.gov/91/statute/STATUTE-84/STATUTE-84-Pg1560.pdf
Baracchi, D., Lihoreau, M., & Giurfa, M. (2017). Do insects have emotions? Some insights from bumble bees. Frontiers in Behavioral Neuroscience, 11, 157. https://doi.org/10.3389/fnbeh.2017.00157
Barrett, M. (2024a). New frontiers in animal welfare science: an introduction to (farmed) insects. Veterinary Records, 194, 344-346. https://doi.org/10.1002/vetr.4229
Barrett, M. (2024b). Pain and Suffering in Farmed Animals: First Steps Towards Better Understanding and Management. Insects. OSF Preprints, 1-47. https://doi.org/10.31219/osf.io/qem7f
Barrett, M., & Adcock, S. J. J. (2023). Animal welfare science: an integral piece of sustainable insect agriculture. Journal of Insects as Food and Feed, 10(4), 517-531. https://doi.org/10.1163/23524588-20230126
Barrett, M., & Fischer, B. (2023). Challenges in farmed insect welfare: beyond the question of sentience. Animal Welfare, 32, e4. https://doi.org/10.1017/awf.2022.5
Barrett, M., & Fischer, B. (2024). I. The Era Beyond Eisemann et al. (1984): Insect pain in the 21st century. OSF Preprints. https://doi.org/osf.io/preprints/osf/ng7pu
Barrett, M., Fischer, B., & Buchmann, S. (2023a). Informing policy and practice on insect pollinator declines: tensions between conservation and animal welfare. Frontiers in Ecology and Evolution, 10, 1071251. https://doi.org/10.3389/fevo.2022.1071251
Barrett, M., Chia, S. Y., Fischer, B., & Tomberlin, J. K. (2023b). Welfare considerations for farming black soldier flies, Hermetia illucens (Diptera: Stratiomyidae): a model for the insects as food and feed industry. Journal of Insects as Food and Feed, 9(2), 119-148. https://doi.org/10.3920/JIFF2022.0041
Barrett, M., Godfrey, R. K., Schnell, A., & Fischer, B. (2024a). Farmed yellow mealworm (Tenebrio molitor; Coleoptera: Tenebrionidae) welfare: species-specific recommendations for a global industry. Journal of Insects as Food and Feed, 10(6), 903-948. https://dx.doi.org/10.1163/23524588-20230104
Barrett, M., Miranda, C., Veloso, I. T., Flint, C., Perl, C. D., Martinez, A., & Tomberlin, J. K. (2024b). Grinding as a slaughter method for farmed black soldier fly (Hermetia illucens) larvae: Empirical recommendations to achieve instantaneous killing. Animal Welfare, 33, e16. https://doi.org/10.1017/awf.2024.10
Barrett, M., Drewery, M., & Fischer, B. (2025). Entomologists' knowledge of, and attitudes towards, insect welfare in research and education. Ecological Entomology, 50(3), 468-484. https://doi.org/10.1111/een.13415
Barron, A. B., & Klein, C. (2016). What insects can tell us about the origins of consciousness. Proceedings of the National Academy of Sciences, 113(18), 4900-4908. https://doi.org/10.1073/pnas.1520084113
Bateson, M., Desire, S., Gartside, S. E., & Wright, G. A. (2011). Agitated honeybees exhibit pessimistic cognitive biases. Current Biology, 21, 1070-1073. https://doi.org/10.1016/j.cub.2011.05.017
Bear, C. (2019). Approaching insect death: Understandings and practices of the UK’s edible insect farmers. Society & Animals, 27(7), 751-768. https://doi.org/10.1163/15685306-00001871
Bear, C. (2021). Making insects tick: Responsibility, attentiveness and care in edible insect farming. Environment and Planning E: Nature and Space, 4(3), 1010-1030. http://dx.doi.org/10.1177/2514848620945321
Belluco, S., Bertola, M., Montarsi, F., Di Martino, G., Granato, A., Stella, R., Martinello, M., Bordin, F., & Mutinelli, F. (2023). Insects and public health: an overview. Insects, 14(3), 240. https://doi.org/10.3390/insects14030240
Bermúdez-Serrano, I. M., Quirós-Blanco, A. M., & Acosta-Montoya, Ó. (2023). Producción de insectos comestibles: retos, oportunidades y perspectivas para Costa Rica. Agronomía Mesoamericana, 34(3), 53052. https://doi.org/10.15517/am.2023.53052
Birch, J. (2017a). Animal sentience and the precautionary principle. Animal sentience, 2(16), 1. https://doi.org/10.51291/2377-7478.1200
Birch, J. (2017b). Refining the precautionary framework. Animal Sentience, 2(16), 20. https://doi.org/10.51291/2377-7478.1279
Birch, J., Burn, C., Schnell, A., Browning, H., & Crump, A. (2021). Review of the evidence of sentience in cephalopod molluscs and decapod crustaceans. London School of Economics. The London School of Economics and Political Science. https://www.lse.ac.uk/News/News-Assets/PDFs/2021/Sentience-in-Cephalopod-Molluscs-and-Decapod-Crustaceans-Final-Report-November-2021.pdf.
Bierman, A., & Lloyd, M. (2024). Metabarcoding and eDNA for insect conservation. In J. S. Pryke, M. J. Samways, T. R. New, P. Cardoso, & R. Gaigher (Eds), Routledge Handbook of Insect Conservation (pp. 487-500). Routledge. https://doi.org/10.4324/9781003285793-43
Bjerge, K., Alison, J., Dyrmann, M., Frigaard, C. E., Mann, H. M., & Høye, T. T. (2023). Accurate detection and identification of insects from camera trap images with deep learning. PLOS Sustainability and Transformation, 2(3), e0000051. https://doi.org/10.1371/journal.pstr.0000051
Blackiston, D. J., Silva Casey, E., & Weiss, M. R. (2008). Retention of memory through metamorphosis: can a moth remember what it learned as a caterpillar?. PloS one,3(3), e1736. https://doi.org/10.1371/journal.pone.0001736
Brink, C. B., & Lewis, D. I. (2023). The 12 Rs Framework as a Comprehensive, Unifying Construct for Principles Guiding Animal Research Ethics. Animals, 13(7), 1128. https://doi.org/10.3390/ani13071128
Browning, H., & Birch, J. (2022). Animal sentience. Philosophy Compass, 17(5), e12822. https://doi.org/10.1111/phc3.12822
Buehlmann, C., Mangan, M., & Graham, P. (2020). Multimodal interactions in insect navigation. Animal Cognition, 23, 1129-1141. https://doi.org/10.1007/s10071-020-01383-2
Byrne, A. Q. (2023). Reimagining the future of natural history museums with compassionate collection. PLoS Biology, 21(5), e3002101. https://doi.org/10.1371/journal.pbio.3002101
Carey, J. R. (2001). Demographic mechanisms for the evolution of long life in social insects. Experimental Gerontology,36(4-6), 713-722. https://doi.org/10.1016/S0531-5565(00)00237-0
Caron, D. P., Rimniceanu, M., Scibelli, A. E., & Trimmer, B. A. (2020). Nociceptive neurons respond to multimodal stimuli in Manduca sexta. Journal of Experimental Biology, 223(3), jeb218859. https://doi.org/10.1242/jeb.218859
Carruthers, P. (2007). Invertebrate minds: A challenge for ethical theory. The Journal of Ethics, 11, 275-297. https://doi.org/10.1007/s10892-007-9015-6
Cattaneo, A., Meneguz, M., Dabbou, S., Tambone, F., & Scaglia, B. (2024). Local circular economy: BSF insect rearing in the Italian Agri-Food Industry. Waste Management, 179, 234-244. https://doi.org/10.3390/insects15100817
Chapman, J. W., Drake, V. A., & Reynolds, D. R. (2011). Recent insights from radar studies of insect flight. Annual Review of Entomology, 56(1), 337-356. https://doi.org/10.1146/annurev-ento-120709-144820
Chen, Z., Liu, Y. Q., Wu, Y. F., Lee, M., Li, H., & Cai, W. Z. (2022) A survey of the tribe Ectinoderini (Hemiptera: Reduviidae: Harpactorinae) of China, with the description of a new Ectinoderus species. Journal of Asia-Pacific Entomology, 25(4), 101974. https://doi.org/10.1016/j.aspen.2022.101974
Chittka, L. (2022). The Mind of a Bee. Princeton University Press. https://doi.org/10.1515/9780691236247
Chittka, L., & Rossi, N. (2023). Bees learn to dance. Experience yields precision in the waggle dance of honey bees. Science, 379, 985-986. https://doi.org/10.1e126/science.adg6020
Chua, P. Y. S., Bourlat, S. J., Ferguson, C., Korlevic, P., Zhao, L., Ekrem, T., Meier, R., & Lawniczak, M. K. N. (2023). Future of DNA-based insect monitoring. Trends in Genetics, 39(7), 531-544. https://doi.org/10.1016/j.tig.2023.02.012
Cooper, J. E., Pellett, S. & O´Brien, M. F. (2022). Insects. In G. A. Lewbart (Ed.), Invertebrate Medicine (pp 413-438). John Wiley & Sons Inc. https://doi.org/10.1002/9781119569831.ch18
Costello, M. J., Beard, K. H., Corlett, R. T., Cumming, G. S., Devictor, V., Loyola, R., Maas, B., Miller-Rushing, A. J., Pakeman, R., & Primack, R. B. (2016). Field work ethics in biological research. Biological Conservation, 203, 268-271. https://doi.org/10.1016/j.biocon.2016.10.008
Crespi-Abril, A. C., & Rubilar, T. (2021). Moving forward in the ethical consideration of invertebrates in experimentation: Beyond the Three R’s Principle. Revista de Biología Tropical, 69, 346-357. https://doi.org/10.15517/rbt.v69iSuppl.1.46366
Crespi-Abril, A. C., & Rubilar, T. (2024). Ethics and Welfare in invertebrates: a stepping-stone to research and animal production. Revista de Biología Tropical, 72, e58228. https://doi.org/10.15517/rev.biol.trop..v72iS1.58228
Crump, A., Gibbons, M., Barrett, M., Birch, J., & Chittka, L. (2023). Is it time for insect researchers to consider their subjects’ welfare?. PLoS biology, 21(6), e3002138. https://doi.org/10.1371/journal.pbio.3002138
Cuff, J. P., Barrett, M., Gray, H., Fox, C., Watt, A., & Aimé, E. (2024). The case for open research in entomology: reducing harm, refining reproducibility and advancing insect science. Agricultural and Forest Entomology, 26(3), 285-295. https://doi.org/10.1111/afe.12617
Curtis, V. (2013). Don't look, don't touch, don't eat: The science behind revulsion. University of Chicago Press. https://doi.org/10.7208/chicago/9780226089102.001.0001
Czaczkes, T. J. (2022). Advanced cognition in ants. Myrmecological News, 32, 51-64. https://doi.org/10.25849/myrmecol.news_032:051
Danchin, É., Nöbel, S., Pocheville, A., Dagaeff, A. C., Demay, L., Alphand, A. Ranty-Roby, S., van Renssen, L., Monier, M., Gazagne, E., Allain, M., & Isabel, G. (2018). Cultural Flies: Conformist Social Learning in Fruit Flies Predicts Long-Lasting Mate-Choice Traditions. Science, 362, 1025-1030. https://doi.org/10.1126/science.aat1590
Danchin, É., Guillaume, I., & Nöbel, S. (2024). Culture in Insects. In J. J. Tehrani, J. Kendal, & R. Kendal (Eds.), The Oxford Handbook of Cultural Evolution (337-388). Oxford Academic. https://doi.org/10.1093/oxfordhb/9780198869252.013.28
Das Chakraborty, S., & Sachse, S. (2021). Olfactory processing in the lateral horn of Drosophila. Cell and Tissue Research, 383, 113-123. https://doi.org/10.1007/s00441-020-03392-6
Dawkins, M. S. (2006). Through animal eyes: what behaviour tells us. Applied Animal Behaviour Science, 100(1-2), 4-10. https://doi.org/10.1016/j.applanim.2006.04.010
De Goede, D. M., Erens, J., Kapsomenou, E., & Peters, M. (2013). Large scale insect rearing and animal welfare. In H. Rocklinsberg & P. Sandin (Eds.), The Ethics of Consumption (pp. 236-242). Wageningen Academic. https://doi.org/10.3920/9789086867844_040
de Souza Valente, C. (2025). Rethinking Sentience: Invertebrates as Worthy of Moral Consideration. Journal of Agricultural and Environmental Ethics, 38(3), 1-19. https://doi.org/10.1007/s10806-024-09940-2
De Waal, F. B. (1999). Anthropomorphism and anthropodenial: consistency in our thinking about humans and other animals. Philosophical Topics,27(1), 255-280. http://www.jstor.org/stable/43154308
Delvendahl, N., Rumpold, B. A., & Langen, N. (2022). Edible insects as food-insect welfare and ethical aspects from a consumer perspective. Insects, 13(2), 121. https://doi.org/10.3390/insects13020121
Döring, T. F., & Chittka, L. (2011). How human are insects, and does it matter. Formosan Entomology, 31, 85-99.
Dona, H. S. G., Solvi, C., Kowalewska, A., Mäkelä, K., MaBouDi, H., & Chittka, L. (2022). Do bumble bees play? Animal Behaviour, 194, 239-251. https://doi.org/10.1016/j.anbehav.2022.08.013
Dong, S., Lin, T., Nieh, J. C., & Tan, K. (2023). Social signal learning of the waggle dance in honey bees. Science, 379, 1015-1018. https://doi.org/10.1126/science.ade1702
Draney, M. L. (1997). Ethical obligations toward insect pests. Ethics and the Environment,1, 5-23. https://www.jstor.org/stable/27766028
D’Silva, J., Dalton, H., Boyland, N. K., & Turner, J. (2024). Animal sentience: The science and its implications, with particular reference to farmed animals. Animal Research and One Health, 2(2), 230-236. https://doi.org/10.1002/aro2.65
Durrant, R. (2024). Invertebrate Justice: Extending The Boundaries of Non-Speciesist Green Criminology. Springer Nature. https://doi.org/10.1007/978-3-031-64443-6
Eisemann, C. H., Jorgensen, W. K., Merritt, D. J., Rice, M. J., Cribb, B. W., Webb, P. D., & Zalucki, M. P. (1984). Do insects feel pain? A biological view. Experientia,40(2), 164-167. https://doi.org/10.1007/BF01963580
Elwood, R. W. (2011). Pain and suffering in invertebrates?. ILAR Journal, 52(2), 175-184. https://doi.org/10.1093/ilar.52.2.175
Elwood, R. W. (2023). Behavioural indicators of pain and suffering in arthropods and might pain bite back?. Animals, 13(16), 2602. https://doi.org/10.3390/ani13162602
Emberts, Z., St. Mary, C. M., Howard, C. C., Forthman, M., Bateman, P. W., Somjee, U., Hwang, W. S., Li, D., Kimball, R. T., & Miller, C. W. (2020). The evolution of autotomy in leaf-footed bugs. Evolution, 74, 897-910. https://doi.org/10.1111/evo.13948
Erens, J., van Es, S., Haverkort, F., Kapsomenou, E., & Luijben, A. (2012). A bug’s life. Large-Scale Insect Rearing in Relation to Animal Welfare. Wageningen University and Research. https://venik.nl/onewebmedia/Rapport-Large-scale-insect-rearing-in-relation-to-animal-welfare.pdf
Fenton, F. A. (1952). Field crop insects. Macmillan.
Ferrante, M., Howe, A. G., & Lövei, G. L. (2024). Experimental considerations support the use of artificial sentinel prey—a comment on Rodríguez-Campbell et al. Journal of Biogeography, 51, 2152-2155. https://doi.org/10.1111/jbi.14978
Fischer, B., & Larson, B. M. H. (2019). Collecting insects to conserve them: a call for ethical caution. Insect Conservation and Diversity, 12(3), 173-182. https://doi.org/10.1111/icad.12344
Fischer, B., Barrett, M., Adcock, S., Barron, A., Browning, H., Chittka, L., Crump, A., Drinkwater, E., Dunlap, A., Gibbons, M., & Perl, C. (2023). Guidelines for protecting and promoting insect welfare in research. Insect Welfare Research Society.
Fischer, B., Gottlieb, J., Schnell, A. K., & Barrett, M. (2025) Defending and refining the Birch et al. (2021) precautionary framework for animal sentience. Animal Welfare, 34, e28. https://doi.org/10.1017/awf.2025.27
Flindt, R. (2006). Amazing numbers in biology. Springer Science & Business Media. https://doi.org/10.1007/3-540-30147-X
Foley, B. R., Marjoram, P., & Nuzhdin, S. V. (2017). Basic reversal-learning capacity in flies suggests rudiments of complex cognition. Plos One, 12(8), e0181749. https://doi.org/10.1371/journal.pone.0181749
Fore, G. A., & Barrett, M. (2024). Reflecting on collecting: Ethical challenges in entomology education. American Entomologist,70(4), 22-24. https://doi.org/10.1093/ae/tmae066
Frank, E. T., Kesner, L., Liberti, J., Helleu, Q., LeBoeuf, A. C., Dascalu, A., Sponsler, D. B., Azuma, F., Economo, E. P., Waridel, P., Engel, P., Schmitt, T., & Keller, L. (2023). Targeted treatment of injured nestmates with antimicrobial compounds in an ant society. Nature Communications, 14(1), 8446. https://doi.org/10.1038/s41467-023-43885-w
Frank, E. T., Buffat, D., Liberti, J., Aibekova, L., Economo, E. P., & Keller, L. (2024). Wound-dependent leg amputations to combat infections in an ant society. Current Biology, 34(14), 3273-3278. https://doi.org/10.1016/j.cub.2024.06.021
Freelance, C. B. (2019). To regulate or not to regulate? The future of animal ethics in experimental research with insects. Science and Engineering Ethics,25(5), 1339-1355. https://doi.org/10.1007/s11948-018-0066-9
Formato, G., Giannottu, E., Roncoroni, C., Lorenzi, V., & Brajon, G. (2024). Apis mellifera welfare: definition and future directions. Frontiers in Animal Science, 5, 1486587. https://doi.org/10.3389/fanim.2024.1486587
Gantz, V. M., & Akbari, O. S. (2018). Gene editing technologies and applications for insects. Current Opinion in Insect Science, 28, 66-72. https://doi.org/10.1016/j.cois.2018.05.006
Garrido, C., & Nanetti, A. (2019). Welfare of managed honey bees. In C. Carere & J. Mather (Eds.), The Welfare of Invertebrate Animals (pp. 69-104). Springer. https://doi.org/10.1007/978-3-030-13947-6
Gibbons, M., Crump, A., Barrett, M., Sarlak, S., Birch, J., & Chittka, L. (2022a). Can insects feel pain? A review of the neural and behavioural evidence. Advances in Insect Physiology, 63, 155-229. https://doi.org/10.1016/bs.aiip.2022.10.001
Gibbons, M., Versace, E., Crump, A., Baran, B., & Chittka, L. (2022b). Motivational trade-offs and modulation of nociception in bumblebees. Proceedings of the National Academy of Sciences, 119(31), e2205821119. https://doi.org/10.1073/pnas.2205821119
Gibbons, M., Pasquini, E., Kowalewska, A., Read, E., Gibson, S., Crump, A., Solvi, C., Versace, E., & Chittka, L. (2024). Noxious stimulation induces self-protective behavior in bumblebees. Iscience, 27(8), 110440. https://doi.org/10.1016/j.isci.2024.110440
Giurfa, M. (2013). Cognition with few neurons: higher-order learning in insects. Trends in Neurosciences, 36(5), 285-294. https://doi.org/10.1016/j.tins.2012.12.011
Giurfa, M. (2015). Learning and cognition in insects. Wiley Interdisciplinary Reviews: Cognitive Science,6(4), 383-395. https://doi.org/10.1002/wcs.1348
González, V. H., Osborn, A. L., Brown, E. R., Pavlick, C. R., Enríquez, E., Tscheulin, T., Petanidou, T., Hranitz, J. M., & Barthell, J. F. (2020). Effect of pan trap size on the diversity of sampled bees and abundance of bycatch. Journal of Insect Conservation,24(3), 409-420. https://doi.org/10.1007/s10841-020-00224-4
Gowda, S. B. M., Paranjpe, P. D., Reddy, O. V., Thiagarajan, D., Palliyil, S., Reichert, H., & VijayRaghavan, K. (2018). GABAergic inhibition of leg motoneurons is required for normal walking behavior in freely moving Drosophila. Proceedings of the National Academy of Sciences, 115(9), E2115-E2124. https://doi.org/10.1073/pnas.1713869115
Grimaldi, D. A. (2023). The Complete Insect: Anatomy, Physiology, Evolution, and Ecology. Princeton University Press. https://doi.org/10.1515/9780691243115
Gritsai, O. B., Dubynin, V. A., Pilipenko, V. E., & Petrov, O. P. (2004). Effects of peptide and non-peptide opioids on protective reaction of the cockroach Periplaneta americana in the “hot camera”. Journal of Evolutionary Biochemistry and Physiology, 40, 153-160. https://doi.org/10.1023/B:JOEY.0000033806.85565.3e
Groening, J., Venini, D., & Srinivasan, M. V. (2017). In search of evidence for the experience of pain in honeybees: A self-administration study. Scientific reports, 7(1), 45825. https://doi.org/10.1038/srep45825
Guo, X., Ma, Z., & Kang, L. (2015). Serotonin enhances solitariness in phase transition of the migratory locust. Frontiers in Behavioral Neuroscience, 9, 129. https://doi.org/10.3389/fnbeh.2015.00129
Harris, J. T., & Eisenstein, E. M. (1999). Transfer of learned information between ganglia in the insect ventral nerve cord. Behavioural Brain Research, 103(2), 211-217. https://doi.org/10.1016/s0166-4328(99)00039-x
Harrison, M. A., & Hall, A. E. (2010). Anthropomorphism, empathy, and perceived communicative ability vary with phylogenetic relatedness to humans. Journal of Social, Evolutionary, and Cultural Psychology,4(1), 34-48. https://doi.org/10.1037/h0099303
Heniff, A. C., Gorges, M. A., Lewbart, G. A., & Balko, J. A. (2023). Evaluation of targeted injections of ivermectin or potassium chloride for euthanasia of anesthetized thorny devil stick insects (Eurycantha calcarata). Journal of Zoological and Botanical Gardens, 4(1), 231-239. https://doi.org/10.3390/jzbg4010020
Ings, T. C., & Chittka, L. (2008). Speed-accuracy tradeoffs and false alarms in bee responses to cryptic predators. Current Biology, 18, 1520-1524. https://doi.org/10.1016/j.cub.2008.07.074
Jirkof, P., Rudeck, J., & Lewejohann, L. (2019). Assessing affective state in laboratory rodents to promote animal welfare—what is the progress in applied refinement research?. Animals,9(12), 1026. https://doi.org/10.3390/ani9121026
Kaebnick, G. E., Collins, J. P., Jayaram, A., Tiernan, R. G., Barnhill, K., Carter, L., Chemhuru, M., Fresia, P., Jennings, B., Meine, C., Ndebele, P., Palmer, C., Preston, C., Redford, K. H., Rohwer, Y., Sandler, R., Scott, M. J., Taitingfong, R., & Zoloth, L. (2025). Deliberate extinction by genome modification: An ethical challenge. Science, 388(6748), 707-709. https://doi.org/10.1126/science.adv4045
Kartsev, V. M. (2014). Phenoptosis in arthropods and immortality of social insects. Biochemistry, 79, 1032-1048. https://doi.org/10.1134/S0006297914100058
Kellert, S. R. (1993). Values and perceptions of invertebrates. Conservation Biology, 7(4), 845-855. https://doi.org/10.1046/j.1523-1739.1993.740845.x
Keyghobadi, N., Crawford, L. A., & Desjardins, S. (2021). High genetic drift in endangered northern peripheral populations of the Behr's hairstreak butterfly (Satyrium behrii). Insect Conservation and Diversity, 14(3), 403-411. https://doi.org/10.1111/icad.12469
Khuong, T. M., Wang, Q. P., Manion, J., Oyston, L. J., Lau, M. T., Towler, H., Lin, Y. Q., & Neely, G. G. (2019). Nerve injury drives a heightened state of vigilance and neuropathic sensitization in Drosophila. Science Advances, 5, eaaw4099. https://doi.org/10.1126/sciadv.aaw4099
Klein, B. A. (2011). The curious connection between insects and dreams. Insects, 3(1), 1-17. https://doi.org/10.3390/insects3010001
Klein, C., & Barron, A. B. (2016). Crawling around the hard problem of consciousness. Proceedings of the National Academy of Sciences, 113(27), E3814-E3815. https://doi.org/10.1073/pnas.1607409113
Klobučar, T., & Fisher, D. N. (2023). When do we start caring about insect welfare?. Neotropical Entomology, 52(1), 5-10. https://doi.org/10.1007/s13744-022-01023-z
Knobe, J. (2003). Intentional action and side effects in ordinary language. Analysis, 63(3),190-194. https://doi.org/10.1093/analys/63.279.190
Kohlberg, A. B., Myers, C. R., & Figueroa, L. L. (2024). From buzzes to bytes: a systematic review of automated bioacoustics models used to detect, classify and monitor insects. Journal of Applied Ecology, 61(6), 1199-1211. https://doi.org/10.1111/1365-2664.14630
Krashes, M. J., Keene, A. C., Leung, B., Armstrong, J. D., & Waddell, S. (2009). A neural circuit mechanism integrating motivational state with memory expression in Drosophila. Cell, 139(2), 416-427. https://doi.org/10.1016/j.cell.2009.08.035
Kriebel, D., Tickner, J., Epstein, P., Lemons, J., Levins, R., Loechler, E. L., Quinn, M., Rudel, R., Schettler, T., & Stoto, M. (2001). The precautionary principle in environmental science. Environmental Health Perspectives, 109(9), 871-876. https://doi.org/10.1289/ehp.01109871
Lähteenmäki-Uutela, A., Marimuthu, S. B., & Meijer, N. (2021). Regulations on insects as food and feed: a global comparison. Journal of Insects as Food and Feed, 7(5), 849-856. https://doi.org/10.3920/JIFF2020.0066
Lambert, H., Moccia, J., Bjorkbom, C., & Cochet, M. (2022). Insect Farming and sustainable food systems: the precautionary principle. Eurogroup for Animals. https://www.eurogroupforanimals.org/files/eurogroupforanimals/2022-07/1813%20Insect%20report_ONLINE_0.pdf
Leadbeater, E., & Chittka, L. (2007). The dynamics of social learning in an insect model, the bumblebee (Bombus terrestris). Behavioral Ecology and Sociobiology, 61, 1789-1796. https://doi.org/10.1007/s00265-007-0412-4
Li, W., Wang, Z., Syed, S., Lyu, C., Lincoln, S., O ’ Neil, J., Nguyen, A. D., Feng, I., & Young, M. W. (2021). Chronic social isolation signals starvation and reduces sleep in Drosophila. Nature, 597(7875), 239-244. https://doi.org/10.1038/s41586-021-03837-0
Lockwood, J. (2013). The infested mind: Why humans fear, loathe, and love insects. Oxford University Press.
Lövei, G. L., & Ferrante, M. (2024). The use and prospects of nonlethal methods in entomology. Annual Review of Entomology, 69(1), 183-198. https://doi.org/10.1146/annurev-ento-120220-024402
Lövei, G. L., Ferrante, M., Möller, D., Möller, G., & Vincze, É. (2023). The need for a (non-destructive) method revolution in entomology. Biological Conservation, 282, 110075. https://doi.org/10.1016/j.biocon.2023.110075
Lum, A. M., & Keller, K. A. (2024). Anesthesia and humane euthanasia methods for king worm larvae (Zophobas morio). Zoo Biology, 43(2), 164-168. https://doi.org/10.1002/zoo.21821
MacMillan, H. A., Nørgård, M., MacLean, H. J., Overgaard, J., & Williams, C. J. (2017). A critical test of Drosophila anaesthetics: Isoflurane and sevoflurane are benign alternatives to cold and CO2. Journal of Insect Physiology, 101, 97-106. http://dx.doi.org/10.1016/j.jinsphys.2017.07.005
Magalhães-Sant’Ana, M. (2009). Consciência animal: para além dos vertebrados. Jornal de Ciências Cognitivas. https://www.researchgate.net/publication/307167210
Makarova, A. A., Polilov, A. A., & Chklovskii, D. B. (2021). Small brains for big science. Current Opinion in Neurobiology, 71, 77-83. https://doi.org/10.1016/j.conb.2021.09.007
Mancini, N., Hranova, S., Weber, J., Weiglein, A., Schleyer, M., Weber, D., Thum, A. S., & Gerber, B. (2019). Reversal learning in Drosophila larvae. Learning & Memory, 26(11), 424-435. http://doi.org/10.1101/lm.049510.119
Mather, J. A. (2019). Ethics and care: For animals, not just mammals. Animals, 9(12), 1018. http://dx.doi.org/10.3390/ani9121018
Mattila, H. R., Otis, G. W., Nguyen, L. T., Pham, H. D., Knight, O. M., & Phan, N. T. (2020). Honey bees (Apis cerana) use animal feces as a tool to defend colonies against group attack by giant hornets (Vespa soror). PLoS One, 15(12), e0242668. https://doi.org/10.1371/journal.pone.0242668
Mikhalevich, I., & Powell, R. (2020). Minds without spines: Evolutionarily inclusive animal ethics. Animal sentience, 29(1), 1-25. https://www.wellbeingintlstudiesrepository.org/cgi/viewcontent.cgi?article=1527&context=animsent
Mollá-Albaladejo, R., & Sánchez-Alcañiz, J. A. (2021). Behavior individuality: a focus on Drosophila melanogaster. Frontiers in Physiology, 12, 719038. https://doi.org/10.3389/fphys.2021.719038
Montero‐Castaño, A., Koch, J. B. U., Lindsay, T. T. T., Love, B., Mola, J. M., Newman, K., & Sharkey, J. K. (2022). Pursuing best practices for minimizing wild bee captures to support biological research. Conservation Science and Practice, 4(7), e12734. https://doi.org/10.1111/csp2.12734
Moruzzo, R., Riccioli, F., Espinosa Diaz, S., Secci, C., Poli, G., & Mancini, S. (2021). Mealworm (Tenebrio molitor): potential and challenges to promote circular economy. Animals, 11(9), 2568. https://doi.org/10.3390/ani11092568
Murray, M. J. (2022). Euthanasia. In F. A. Lewbart (Ed.), Invertebrate Medicine (pp. 645-650). John Wiley & Sons, Inc. https://doi.org/10.1002/9781119569831.ch27
Mutinelli, F. (2023). Euthanasia and welfare of managed honey bee colonies. Journal of Apicultural Research, 62(1), 2-10. https://doi.org/10.1080/00218839.2021.1895569
Nagel, T. (1974). What is it like to be a bat? Philosophical Review, 83(4), 435-50. https://doi.org/10.2307/2183914
Nakamura, S., Ichiki, R. T., Shimoda, M., & Morioka, S. (2016). Small-scale rearing of the black soldier fly, Hermetia illucens (Diptera: Stratiomyidae), in the laboratory: low-cost and year-round rearing. Applied Entomology and Zoology, 51, 161-166. https://doi.org/10.1007/s13355-015-0376-1
Nieberding, C. M., Van Dyck, H., & Chittka, L. (2018). Adaptive learning in non-social insects: from theory to field work, and back. Current Opinion in Insect Science, 27, 75-81. http://dx.doi.org/10.1016/j.cois.2018.03.008
Núñez, J., Maldonado, H., Miralto, A., & Balderrama, N. (1983). The stinging response of the honeybee: effects of morphine, naloxone and some opioid peptides. Pharmacology Biochemistry and Behavior, 19(6), 921-924. https://doi.org/10.1016/0091-3057(83)90391-X
ONU (Organización de las Naciones Unidas). (1992). Declaración de Río Sobre el Medio Ambiente y el Desarrollo. https://www.un.org/spanish/esa/sustdev/agenda21/riodeclaration.htm
Ožana, S., Pyszko, P., & Dolný, A. (2020). Determination of suitable insect part for non‐lethal DNA sampling: case study of DNA quality and regeneration capability of dragonflies. Insect Conservation and Diversity, 13(4), 319-327. https://doi.org/10.1111/icad.12400
Pali-Schöll, I., Binder, R., Moens, Y., Polesny, F., & Monsó, S. (2019). Edible insects-defining knowledge gaps in biological and ethical considerations of entomophagy. Critical Reviews in Food Science and Nutrition, 59(17), 2760-2771. https://doi.org/10.1080/10408398.2018.1468731
Pasquini, E., Brouwer, J., Di Rollo, V., Baracchi, D., Messina, A., & Frasnelli, E. (2025). Central GABAergic neuromodulation of nocifensive behaviours in bumble bees. iScience, 28(3), 112024. https://doi.org/10.1016/j.isci.2025.112024
Pérez Claudio, E., Rodríguez-Cruz, Y., Arslan, O. C., Giray, T., Agosto Rivera, J. L., Kence, M., Wells, H., & Abramson, C. I. (2018). Appetitive reversal learning differences of two honey bee subspecies with different foraging behaviors. PeerJ,6, e5918. https://doi.org/10.7717/peerj.5918
Perry, C. J., & Baciadonna, L. (2017). Studying emotion in invertebrates: what has been done, what can be measured and what they can provide. Journal of Experimental Biology, 220(21), 3856-3868. https://doi.org/10.1242/jeb.151308
Perry, C. J., Baciadonna, L., & Chittka, L. (2016). Unexpected rewards induce dopamine-dependent positive emotion-like state changes in bumblebees. Science, 353(6307), 1529-1531. https://doi.org/10.1126/science.aaf4454
Perry, C. J., Barron, A. B., & Chittka, L. (2017). The frontiers of insect cognition. Current Opinion in Behavioral Sciences, 16, 111-118. http://dx.doi.org/10.1016/j.cobeha.2017.05.011
Pierce Jr, J. D. (1986). A review of tool use in insects. Florida Entomologist, 69(1), 95-104.
Pitman, J. L., DasGupta, S., Krashes, M. J., Leung, B., Perrat, P. N., & Waddell, S. (2009). There are many ways to train a fly. Fly, 3, 3-9 https://doi.org/10.4161/fly.3.1.7726
Porter, S. D. & C.D. Jorgensen. (1988). Longevity of harvester ant colonies in southern Idaho. Rangeland Ecology & Management/Journal of Range Management Archives, 41, 104-107 https://doi.org/10.2307/3898942
Prudic, K. L. (2024). Perspective Chapter: Insect Conservation, Machine Learning, and Citizen Science Take Flight. In J. Mato, I. Garrido, & F. Ramil (Eds.), Ecosystems and Biodiversity (pp. 1-13). Intechopen. https://doi.org/10.5772/intechopen.115302
Punzo, F. (1980). Neurochemical changes associated with learning in Schistocerca americana (Orthoptera: Acrididae). Journal of the Kansas Entomological Society, 1, 787-796. https://www.jstor.org/stable/25084110
Raffensperger, C., & Tickner, J. (1999). Protecting Public Health and the Environment. Implementing the Precautionary Principle. Island Press.
Raja, S. N., Carr, D. B., Cohen, M., Finnerup, N. B., Flor, H., Gibson, S., Keefe, F. J., Mogil, J. S., Ringkamp, M., Sluka, K. A., Song, X., Stevens, B., Sullivan, M. D., Tutelman, P. R., Ushida, T., & Vader, K. (2020). The revised International Association for the Study of Pain. Definition of pain: concepts, challenges, and compromises. Pain,161(9), 1976-1982. https://doi.org/10.1097/j.pain.0000000000001939
Ramano, N., Fischer, H., & Egnew, N. (2020). Color and sugar preferences of adult black soldier fly (Hermetia illucens) (Diptera: Stratiomyidae) for feeding and oviposition. Journal of Environmental Biology, 41, 1132-1137. https://doi.org/10.22438/jeb/41/5(SI)/MS_03
Rhodes, M. W., Bennie, J. J., Spalding, A., ffrench‐Constant, R. H., & Maclean, I. M. (2022). Recent advances in the remote sensing of insects. Biological Reviews, 97(1), 343-360. https://doi.org/10.1111/brv.12802
Rillich, J., & Stevenson, P. A. (2015). Releasing stimuli and aggression in crickets: octopamine promotes escalation and maintenance but not initiation. Frontiers in Behavioral Neurosciences, 9, 95. https://doi.org/10.3389/fnbeh.2015.00095
Rodrigues, D., Goodner, B. W., & Weiss, M. R. (2010). Reversal learning and risk‐averse foraging behavior in the monarch butterfly, Danaus plexippus (Lepidoptera: Nymphalidae). Ethology, 116(3), 270-280. https://doi.org/10.1111/j.1439-0310.2009.01737.x
Rodríguez-Rodríguez, D. (2024). The Great Family of Life: Rethinking the place of Homo sapiens in the Biosphere. Springer.
Roger, F., Ghanavi, H. R., Danielsson, N., Wahlberg, N., Löndahl, J., Pettersson, L. B., Andersson, G. K. S., Olén, N. B., & Clough, Y. (2022). Airborne environmental DNA metabarcoding for the monitoring of terrestrial insects—A proof of concept from the field. Environmental DNA, 4(4), 790-807. https://doi.org/10.1002/edn3.290
Rowe, A. (2020a). Insects raised for food and feed — global scale, practices, and policy. Rethink Priorities. https://rethinkpriorities.org/publications/insects-raised-for-food-and-feed
Rowe, A. (2020b). The Scale of Direct Human Impact on Invertebrates. Rethink Priorities. https://doi.org/10.31219/osf.io/psvk2
Rowe, A. (2021). Silk production: global scale and animal welfare issues. Rethink Priorities. https://rethinkpriorities.org/publications/silk-production
Rowe, E., López, K. R., Robinson, K. M., Baudier, K. M., & Barrett, M. (2024). Farmed cricket (Acheta domesticus, Gryllus assimilis, and Gryllodes sigillatus; Orthoptera) welfare considerations: Recommendations for improving global practice. Journal of Insects as Food and Feed, 10(8), 1253-1311. https://doi.org/10.1163/23524588-00001087
Roy, C., & Vidal-Naquet, N. (2022). Euthanasia of honey-bee colonies: Proposal of a standard method. The Canadian Veterinary Journal, 63(5), 541-544
Roy, D. B., Alison, J., August, T. A., Bélisle, M., Bjerge, K., Bowden, J. J., Bunsen, M. J., Cunha, F., Geissmann, Q., Goldmann, K., Gomez-Segura, A., Jain, A., Huijbers, C., Larrivée, M., Lawson, J. L., Mann, H. M., Mazerolle, M. J., McFarland, K. P., Pasi, L., Peters, S., Pinoy, N., Rolnick, D., Skinner, G. L., Strickson, O. T., Svenning, A., Teagle, S. & Høye, T. T. (2024). Towards a standardized framework for AI-assisted, image-based monitoring of nocturnal insects. Philosophical Transactions of the Royal Society B, 379(1904), 20230108. https://doi.org/10.1098/rstb.2023.0108
Russo, L. (2025). Roadside Surveys as a Method for Nonlethal Insect Monitoring. Psyche: A Journal of Entomology,1, 8881730. https://doi.org/10.1155/psyc/8881730
Safavi, A., Thrastardottir, R., Thorarinsdottir, R. I., & Unnthorsson, R. (2024). Insect Production: A Circular Economy Strategy in Iceland. Sustainability, 16(20), 9063. https://doi.org/10.3390/su16209063
Salman, I. N., Ferrante, M., Möller, D. M., Gavish-Regev, E., & Lubin, Y. (2020). Trunk refugia: A simple, inexpensive method for sampling tree trunk arthropods. Journal of Insect Science, 20(2), 5. https://doi.org/10.1093/jisesa/ieaa012
Sanabria, F., Santos, C., Rajagopal, S., & Sasaki, T. (2024). Reversal learning in ant colonies. International Journal of Comparative Psychology, 37(1). https://doi.org/10.46867/ijcp.20778
Sandhi, R. K., Pickens, V., Bello, E., Elzay, S., Salgado, S., Hauri, K. C., Ternest, J. J., Constancio, N., Gula, S., Gearner, O. M., Anderson, M., Edeburn, M., Hall, B., Maille, J., Toth, M., Khadka, A., Doherty, E., Musgrove, T., Silva, T., Desoto, A., Rampone, E., Jocson, D., Luppino, M., Pautzke, K., Wagstaff, C., & Wagstaff, C. (2023). Entomology beyond research and education: 2022 student debates. Journal of Insect Science, 23(3), 11. https://doi.org/10.1093/jisesa/iead036
Sanes, J. R., & Zipursky, S. L. (2010). Design principles of insect and vertebrate visual systems. Neuron, 66(1), 15-36. https://doi.org/10.1016/j.neuron.2010.01.018
Santos-Silva, T., Baeta, L. C. F., Soares, G. J. D., Berti, V. F., Lima, R. T. R., Araujo, N. L., & Gama, D. I. D. (2022). Classical analgesic drugs modulate nociceptive-like escape behavior in Drosophila melanogaster larvae. Research Results in Pharmacology, 8(4), 185-196. https://doi.org/10.3897/rrpharmacology.8.91390
Schukraft, J. (2019a). Opinion: Estimating invertebrate sentience. Rethink Priorities. https://rethinkpriorities.org/publications/opinion-estimating-invertebrate-sentience
Schukraft, J. (2019b). Features relevant to invertebrate sentience, part 2. Rethink Priorities. https://rethinkpriorities.org/publications/features-relevant-to-invertebrate-sentience-part-2
Schukraft, J. (2019c). Features relevant to invertebrate sentience, part 3. Rethink Priorities. https://rethinkpriorities.org/publications/features-relevant-to-invertebrate-sentience-part-3
Schukraft, J. (2019d). Managed honey bee welfare: problems and potential interventions. Rethink priorities. https://rethinkpriorities.org/publications/managed-honey-bee-welfare-problems-and-potentialinterventions.
Scotto, C. (2024). El sesgo antropocéntrico en la cognición animal. Artefactos, 13, 85-116. https://doi.org/10.14201/art2024.31800
Serpell, J. A. (2004). Factors influencing human attitudes to animals and their welfare. Animal Welfare, 13(1), 145-151. https://doi.org/10.1017/S0962728600005222
Shelomi, M. (2021). Pain, Suffering, and Euthanasia in Insects. International Journal of Applied Philosophy,35(1), 31-43. https://doi.org/10.5840/ijap2021129156
Short, A. E. Z., Dikow, T., & Moreau, C. S. (2018). Entomological collections in the age of big data. Annual Review of Entomology, 63, 513-530. https://doi.org/10.1146/annurev-ento-031616-035536
Simons, M., & Tibbetts, E. (2019). Insects as models for studying the evolution of animal cognition. Current Opinion in Insect Science, 34, 117-122. https://doi.org/10.1016/j.cois.2019.05.009
Sitaraman, D., Zars, M., LaFerriere, H., Chen, Y.-C., Sable-Smith, A., Kitamoto, T., Rottinghaus, G. E., & Zars, T. (2008). Serotonin is necessary for place memory in Drosophila. Proceedings of the National Academy of Sciences, 105(14), 5579-5584. https://doi.org/10.1073/pnas.0710168105
Spears, L. & Ramirez, R. A. (2015). Learning to love leftovers: using by-catch to expand our knowledge in Entomology. American Entomologist, 61, 168-173. https://doi.org/10.1093/ae/tmv046
Staab, M., Ohl, M., Zhu, C. D., & Klein, A. M. (2014). A unique nest-protection strategy in a new species of spider wasp. PLoS ONE, 9, e101592. https://doi.org/10.1371/journal.pone.0101592
Stack Whitney, K., & Whitney, K. (2024). Squished Bugs: Teaching and Learning Reflexivity in Ecology. Environmental Humanities, 16(1), 211-229. https://doi.org/10.1215/22011919-10943169
Steel, D. (2015). Philosophy and the Precautionary Principle. Cambridge University Press. https://doi.org/10.1017/CBO9781139939652
Stewart, R. B. (2002). Environmental regulatory decision making under uncertainty. In T. Swanson (Ed.), An Introduction to the Law and Economics of Environmental Policy: Issues in Institutional Design (pp. 71-126). Emerald Group Publishing Limited. https://doi.org/10.1016/S0193-5895(02)20005-6
Strang, C. & Sherry, D. (2014). Serial reversal learning in bumblebees (Bombus impatiens). Animal Cognition, 17(3), 723-734. https://doi.org/10.1007/s10071-013-0704-1
Sunstein, C. R. (2002). Beyond the precautionary principle. University of Pensylvania Law Review, 151, 1003.
Svenningsen, C. S., & Schigel, D. (2024). Sharing insect data through GBIF: novel monitoring methods, opportunities and standards. Philosophical Transactions of the Royal Society B, 379(1904), 20230104. https://doi.org/10.1098/rstb.2023.0104
Sweeney, B. W. & Vannote, R. L. (1982). Population synchrony in mayflies: a predator satiation hypothesis. Evolution, 36, 810-821 https://doi.org/10.1111/j.1558-5646.1982.tb05447.x
Thiagarajan, D., & Sachse, S. (2022). Multimodal information processing and associative learning in the insect brain. Insects, 13(4), 332. https://doi.org/10.3390/insects13040332
Tibbetts, E. A., Pardo-Sanchez, J., Ramirez-Matias, J., & Avarguès-Weber, A. (2021). Individual recognition is associated with holistic face processing in Polistes paper wasps in a species-specific way. Proceedings of the Royal Society B, 288(1943), 20203010. https://doi.org/10.1098/rspb.2020.3010
Tiwasing, P., & Pate, L. (2024). Exploring edible insects as feed in the UK: current challenges and future prospects. Journal of Insects as Food and Feed,11(1), 139-155. https://dx.doi.org/10.1163/23524588-00001264
Tomberlin, J. K., Sheppard, D. C., & Joyce, J. A. (2002). Selected life-history traits of black soldier flies (Diptera: Stratiomyidae) reared on three artificial diets. Annals of the Entomological Society of America, 95(3), 379-386. https://dx.doi.org/10.1603/0013-8746(2002)095[0379:SLHTOB]2.0.CO;2
Tostado, L., & Bollmohr, S. (2022). Pesticide Atlas 2022. Pesticides and Agriculture: Dangerous Substances. https://eu.boell.org/en/PesticideAtlas-PDF
Tracey, W. D., Wilson, R. I., Laurent, G., & Benzer, S. (2003). Painless, a Drosophila gene essential for nociception. Cell, 113(2), 261-273. https://doi.org/10.1016/S0092-8674(03)00272-1
Traniello, J. F., & Avergués Weber, A. (2023). Individual and collective cognition in social insects: what’s in a name?. Behavioral Ecology and Sociobiology, 77(11), 119. https://doi.org/10.1007/s00265-023-03392-w
Trietsch, C., & Deans, A. R. (2018). The insect collectors’ code. American Entomologist, 64(3), 156-158. https://doi.org/10.1093/ae/tmy035
Trout, R. T., Minteer, C. R., Pallipparambil, G. R., Magnus, R. M., & Wiedenmann, R. N. (2010). A perspective on education in research ethics for entomology graduate students. American Entomologist, 56(4), 198-205.
van Huis, A (2019). Welfare of farmed insects. Journal of Insects as Food and Feed,5(3), 159-162. https://doi.org/10.3920/JIFF2020.0061
van Huis, A. (2020). Insects as food and feed, a new emerging agricultural sector: a review. Journal of Insects as Food and Feed, 6(1), 27-44. https://doi.org/10.3920/JIFF2019.0017
van Huis, A. (2021). Prospects of insects as food and feed. Organic Agriculture, 11(2), 301-308. https://doi.org/10.1007/s13165-020-00290-7
van Klink, R., August, T., Bas, Y., Bodesheim, P., Bonn, A., Fossøy, F., Høye, T. T., Jongejans, E., Menz, M. H. M., Miraldo, A., Roslin, T., Roy, H. E., Ruczyński, I., Schigel, D., Schäffler, L., Sheard, J. K., Svenningsen, C., Tschan, G. F., Wäldchen, J., Zizka, V. M. A., Åström, J., & Bowler, D. E. (2022). Emerging technologies revolutionise insect ecology and monitoring. Trends in Ecology & Evolution, 37(10), 872-885. https://doi.org/10.1016/j.tree.2022.06.001
Vicente-Sáez, R., & Martinez-Fuentes, C. (2018). Open Science now: a systematic literature review for an integrated definition. Journal of Business Research, 88, 428-436. https://doi.org/10.1016/j.jbusres.2017.12.043
Villamor Iglesias, A. (2021). The suffering of invertebrates: an approach from animal ethics. Tópicos, 61, 403-420. http://doi.org/10.21555/top.v0i61.1197
Walters, E. T., Illich, P. A., Weeks, J. C., & Lewin, M. R. (2001). Defensive responses of larval Manduca sexta and their sensitization by noxious stimuli in the laboratory and field. Journal of Experimental Biology, 204(3), 457-469. http://doi.org/10.1242/jeb.204.3.457
Weber, C., Noël, G., Sickel, W., Monaghan, M. T., Bonn, A., & Lokatis, S. (2024). Urban pavements as a novel habitat for wild bees and other ground-nesting insects. Urban Ecosystems, 27(6), 2453-2467. https://doi.org/10.1007/s11252-024-01569-3
Webster, J. (1994). Assessment of animal welfare: The five freedoms. In J. Webster (ed.), Animal Welfare: A Cool Eye Towards Eden (pp. 10-14). Blackwell Science.
Weidhaas, D. E., & Labrecque, G. C. (1970). Studies on the population dynamics of the housefly, Musca domestica L. Bulletin of the World Health Organization, 43(5), 721-725.
Wen, C., Shen, L. M., Chen, J., Zhang, J. L., Feng, Y., Wang, Z., Chen, X., Cai, J., Wang, L., He, Y., Wen, X., Ma, T., & Wang, C. (2022). Red imported fire ants cover the insecticide treated surfaces with particles to reduce contact toxicity. Journal of Pest Science, 95, 1135-1150. https://doi.org/10.1007/s10340-021-01474-0
Wen, C., Wang, C., Guo, X., Li, H., Xiao, H., Wen, J., & Dong, S. (2024a). Object use in insects. Insect Science, 31(4), 1001-1014. http://doi.org/10.1111/1744-7917.13275
Wen, C., Lu, Y., Solvi, C., Dong, S., Wang, C., Wen, X., Xiao, H., Dong, S., Wen, J., Peng, F., & Chittka, L. (2024b). Does bumblebee preference of continuous over interrupted strings in string-pulling tasks indicate means-end comprehension?. Elife, 13, RP97018. https://doi.org/10.7554/eLife.97018
Wilkinson, M. D., Dumontier, M., Aalbersberg, I. J., Appleton, G., Axton, M., Baak, A., Blomberg, N., Boiten, J.-W., Bonino da Silva Santos, L., Bourne, P. E., Bouwman, J., Brookes, A. J., Clark, T., Crosas, M., Dillo, I., Dumon, O., Edmunds, S., Evelo, C. T., Finkers, R., Gonzalez-Beltran, A., Gray, A. J. G., Groth, P., Goble, C., Grethe, J. S., Heringa, J., ‘t Hoen, P. A. C., Hooft, R., Kuhn, T., Kok, R., Kok, J., Lusher, S. J., Martone, M. E., Mons, A., Packer, A. L., Persson, B., Rocca-Serra, P., Roos, M., van Schaik, R., Sansone, S.-A., Schultes, E., Sengstag, T., Slater, T., Strawn, G., Swertz, M. A., Thompson, M., van der Lei, J., van Mulligen, E., Velterop, J., Waagmeester, A., Wittenburg, P., Wolstencroft, K., Zhao, J., & Mons, B. (2016). The FAIR Guiding Principles for scientific data management and stewardship. Scientific Data, 3(1), 1-9. http://doi.org/10.1038/sdata.2016.18
Wittman, J. T., & Aukema, B. H. (2020). A guide and toolbox to replicability and open science in entomology. Journal of Insect Science, 20(3), 6. http://doi.org/10.1093/jisesa/ieaa036
WHO (World Health Organization) (2020). Vector-borne diseases. https://www.who.int/news-room/fact-sheets/detail/vector-borne-diseases
Zachariah, T. T. (2022). Invertebrate Animal Welfare. In G. A. Lewbart (Ed.), Invertebrate Medicine (pp. 669-675). Wiley‐Blackwell. http://doi.org/10.1002/9781119569831.ch29
Zeuss, D., Bald, L., Gottwald, J., Becker, M., Bellafkir, H., Bendix, J., Bengel, P., Beumer, L. T., Brandl, R., Brändle, M., Dahlke, S., Farwig, N., Freisleben, B., Friess, N., Heidrich, L., Heuer, S., Höchst, J., Holzmann, H., Lampe, P., . . . Nauss, T. (2024). Nature 4.0: A networked sensor system for integrated biodiversity monitoring. Global Change Biology,30(1), e17056. http://doi.org/10.1111/gcb.17056
Zylinski, S. (2015). Fun and play in invertebrates. Current Biology, 25(1), 10-12.
Burke, C. J., Huetteroth, W., Owald, D., Perisse, E., Krashes, M. J., Das, G., Gohl, G., Gohl, D., Silies, M., Certel, S., & Waddell, S. (2012). Layered reward signalling through octopamine and dopamine in Drosophila. Nature, 492(7429), 433-437. https://doi.org/10.1038/nature11614
Notas
Notas de autor
asalvo@unc.edu.ar
Información adicional
redalyc-journal-id: 3220