Artículos originales
Recepción: 10 Febrero 2021
Aprobación: 14 Julio 2021
Financiamiento
Fuente: Fondo Nacional de Desarrollo Científico y Tecnológico (Fondecyt) de la Agencia Nacional de Investigación y Desarrollo de Chile
Nº de contrato: proyecto Nº 1210719
Beneficiario: Edgar Vogel
Resumen: Se reportaron los resultados de un procedimiento de condicionamiento palpebral en humanos destinado a caracterizar distintos tipos de respuestas. Para ello, ocho participantes recibieron un estímulo condicionado auditivo emparejado con un estímulo incondicionado, soplo de aire en el ojo, mientras que un estímulo condicionado vibro-táctil no fue reforzado. Se demostró la existencia de cuatro repuestas. En los primeros 100 milisegundos (ms) desde la iniciación de los estímulos condicionados ocurren respuestas de pequeña amplitud, denominadas alfa, a las cuales le siguen otras respuestas de amplitud intermedia, denominadas voluntarias (200-300 ms) y condicionadas (300-400 ms). Finalmente, las respuestas incondicionadas ocurren en los 200 ms que siguen el inicio del estímulo incondicionado y son de gran amplitud. Tanto las respuestas voluntarias como condicionadas son más frecuentes en presencia del estímulo A que B. Discutimos la relevancia de identificar y distinguir estos tipos de respuestas en cualquier rutina de condicionamiento clásico.
Palabras clave: respuesta incondicionada, condicionamiento de parpadeo, respuesta condicionada, respuesta voluntaria.
Abstract: We report the results of a human eyeblink conditioning procedure aimed at characterizing different types of responses. To do this, eight participants received an auditory conditioned stimulus paired with an unconditioned stimulus -air puff in the eye- and a vibro-tactile stimulus non reinforced. We demonstrated the existence of four responses. In the first 100 milliseconds (ms) since the initiation of conditioned a small amplitude response, called alpha, appears, which is followed by intermediate amplitude responses, called voluntary (200-300 ms) and conditioned (300-400 ms). Finally, the largest-amplitude responses occur about 200 ms after unconditioned stimulus onset. Both voluntary and conditioned responses are more frequent in the presence of stimulus A that B. We discuss the relevance of identifying and distinguishing these types of responses in any classical conditioning routine.
Keywords: eyeblink conditioning, conditioned response, unconditioned response, voluntary response.
Introducción
Sucesivos emparejamientos de un estímulo conductualmente neutral (EN), también denominado estímulo condicionado (EC), con uno biológicamente significativo denominado estímulo incondicionado (EI), traen como consecuencia que el EN se convierta en un EC y adquiera la capacidad para generar una respuesta condicionada (RC) similar a la respuesta incondicionada (RI) del EI (Pavlov, 1927). Este proceso de aprendizaje se conoce como condicionamiento clásico o pavloviano y ha sido observado en numerosos procedimientos, tales como la salivación (Pavlov, 1927), el parpadeo reflejo (Gormezano, Schneiderman, Deux, & Fuentes, 1962; Hilgard & Campbell, 1936), reacciones de miedo (Estes & Skinner, 1941) y conducta alimenticia (Brown & Jenkins, 1968; García & Koelling, 1966).
La explicación predominante de este fenómeno es que los animales no humanos forman una asociación entre el EC y el EI, la cual es una función del grado en que el EI es predicho por todos los EC presentes en el ensayo (Rescorla & Wagner, 1972; Vogel, Brandon, & Wagner, 2003; Vogel, Ponce, & Wagner, 2019; Vogel & Ponce, 2011; Wagner & Vogel, 2008). Sin embargo, la investigación en humanos ha revelado que en los protocolos de condicionamiento existen también respuestas de carácter no asociativo que dificultan la medición de la RC. El condicionamiento clásico de la respuesta de parpadeo es un caso prototípico de esta problemática. En esta situación, los participantes son expuestos a un tono (EC) seguido de un breve soplo de aire en el ojo (EI) que provoca una RI de parpadeo. Luego de varios ensayos, emerge una RC de parpadeo al tono. Se podría decir aquí que las personas aprenden al menos dos cosas (Shanks, 2010; Weidemann, Tangen, Lovibond, & Mitchell, 2009): la primera es información declarativa acerca de la relación predictiva entre el tono y el soplo de aire (Bolaram Coe, Power, & Cheng, 2020; Lovibond, Liu, Weidemann, & Mitchell, 2011; Weidemann, Satkunarajah, & Lovibond, 2016). La segunda es información no declarativa y consiste en la emisión de una RC de parpadeo (Schade Powers, Coburn-Litvak, & Evinger, 2010). Esto es consistente con la creencia de que el aprendizaje y la memoria no son entidades únicas, sino más bien un conjunto de componentes que operan con formatos, leyes y sistemas cerebrales diferentes (Schacter & Tulving, 1994; Squire, 1994). Una de las distinciones más connotadas es entre el sistema no declarativo, de carácter automático e inconsciente, dependiente de estructuras subcorticales como el cerebelo; y el sistema declarativo, caracterizado por operar de manera consciente y depender del hipocampo (Clark, Manns, & Squire, 2002; Christian & Thompson, 2003).
Los investigadores del condicionamiento en humanos han sido desde el comienzo muy sensibles a las dificultades metodológicas que ofrece esta dualidad (Martin & Levey, 1969; Spence & Ross, 1959). Por una parte, el condicionamiento de parpadeo en humanos se ha caracterizado por la dificultad de identificar la verdadera RC dado los varios tipos de parpadeos que ocurren en el curso del condicionamiento (Martin & Levey, 1969). En los primeros estudios de condicionamiento palpebral en humanos, cualquier respuesta que ocurría entre el inicio del EC y el inicio del EI era identificado como RC (Spence & Ross, 1959). Este procedimiento abarcó un rango de respuestas que se comportaban de maneras muy diferentes durante el curso de la adquisición. Específicamente, además de las RC y RI, se han identificado parpadeos incondicionados al EC denominados respuestas alfa (Ra) y beta (Rb), las cuales tienden a ocurrir a una latencia de 100 milisegundos (ms) y en un rango de 120 a 240 ms luego de la presentación del EC, respectivamente (Grant, 1945; Grant & Norris, 1947; Grant, Norris, & Hornseth, 1948).
Por otra parte, quizás la principal dificultad es la demostración de que un número sustancial de participantes, quienes, además de estar conscientes de las contingencias (Huang et al., 2014; Knuttinen, Power, Preston, & Disterhoft, 2001; Lovibond et al., 2011; Lovibond & Shanks, 2002; Weidemann, Best, Lee, & Lovibond, 2013; Weidemann & Antees, 2012), emiten las llamadas respuestas voluntarias (RV) (Goodrich, 1966; Gormezano, 1965; Rasmussen & Jirenhed, 2017; Spence & Ross, 1959). Específicamente, Spence y Ross (1959) mostraron que algunos sujetos dan una alta incidencia de respuestas que se caracterizan por un cierre fuerte, suave del párpado, con el ojo cerrado hasta después de que el soplo de aire es recibido. Este tipo de respuestas fue observado en aquellos sujetos instruidos a parpadear al EC (instrucción dada antes de iniciar el entrenamiento), especialmente cuando era administrado un fuerte soplo de aire (sobre 2 psi). Este tipo de parpadeos fueron denominados RV. El rango de latencia de la RV es entre los 200-300 ms, seguido del inicio del EC, usando un intervalo EC-EI de aproximadamente 500 ms. Las curvas de condicionamiento obtenidas de sujetos que parpadean voluntariamente comienzan con un mayor porcentaje de respuesta y tienden a alcanzar asíntota en fases más tempranas del condicionamiento que en participantes involuntarios (sujetos que reciben instrucciones neutrales; Spence & Ross, 1959). Otra característica de este tipo de curvas, es un súbito salto de cero al máximo de respuesta, mientras que los involuntarios exhiben un incremento gradual en el porcentaje de RC (curva negativamente acelerada). En respuesta a esto, se ha utilizado un número de estrategias, tales como la eliminación de los parpadeos que ocurren entre 200 a 300 ms luego del EC (Spence & Ross, 1959), la clasificación de respuestas según su topografía (Martin & Levey, 1969), la reducción del componente declarativo mediante técnicas de distracción o enmascaramiento (Clark et al., 2002; Clark & Squire, 1999; Manns, Clark, & Squire, 2000); y el uso de un EI inevitable, por ejemplo, la estimulación eléctrica del nervio trigémino (Schade Powers et al., 2010); sin que aún exista consenso respecto de cuál es la estrategia óptima o incluso si el conocimiento declarativo de la relación entre el EC y el EI es una condición necesaria para todas las instancias de condicionamiento clásico en humanos (McLaren et al., 2014; Mitchell, De Houwer, & Lovibond, 2009).
Esto demanda que los protocolos de condicionamiento establezcan distinciones entre los tipos de respuesta, ya que no todos los parpadeos que emiten las personas en el intervalo EC-EI son, en estricto rigor, RC. Es por ello que basados en los trabajos clásicos de Grant (Grant, 1945; Grant et al., 1948; Grant & Norris, 1947) y de Spence y Ross (1959), el objetivo de esta investigación fue implementar un sistema de análisis que permita caracterizar la topografía temporal de los tipos de respuestas de parpadeo (Ra, Rb, RV, RC y RI) emitidas por los participantes en ensayos de condicionamiento discriminativo. Para ello, los participantes fueron expuestos a dos estímulos, A (EC) y B (EN), donde uno de ellos fue sistemáticamente emparejado con el EI (A+) y el otro no lo fue (B-). Se espera que cualquier cambio en la respuesta de parpadeo que ocurra producto de la mera exposición a los EC y no a su emparejamiento con el EI, debiese ocurrir tanto para el estímulo A como B. Además, en caso de existir condicionamiento, esto debería reflejarse en una mayor frecuencia (o proporción) y amplitud de la RC provocada por el estímulo A que la provocada por el estímulo B.
Método
Participantes
Un total de ocho estudiantes de la Universidad de Talca participaron voluntariamente en el estudio; sin experiencia previa en experimentos similares, cuyas edades fluctuaron entre los 19 y 24 años (cuatro hombres y cuatro mujeres). Todos firmaron un consentimiento informado con la opción de renunciar a participar del experimento en el momento en que considerasen pertinente. Este documento en conjunto con el procedimiento del experimento fue aprobado previamente por el Comité Ético Científico (CEC) de la Universidad de Talca.
Aparatos
El experimento fue realizado en el laboratorio de psicología, en una habitación acústicamente aislada (2.5 x 2.8 x 2.4 metros), tenuemente iluminada mediante lámparas fluorescentes de 18 watts. Para configurar el experimento, la administración de estímulos y la medición de la respuesta palpebral se empleó el sistema Eyeblink Conditioning System (San Diego Instruments, 2022). Los participantes que ingresaron a la sala experimental se sentaron en un sillón a lo largo de toda la sesión, recibiendo como EI un estímulo de tipo táctil consistente en un soplo de aire en el contorno del ojo (9 Psi, 100 ms de duración) administrados a través de un cintillo con un tubo de aire. Por una parte, el EC designado como A, consistió en un tono de 1 kHz (65 dB, 500 ms de duración). El EC designado como B correspondió a una vibración en el respaldo del asiento de la silla experimental (30 Hz, 500 ms de duración).
La respuesta palpebral fue medida por medio de un sensor infrarrojo colocado frente al ojo derecho de los participantes y sujetado por un cintillo que permitía mantener fija la posición del sensor independientemente de los cambios posturales. La respuesta de parpadeo produce una interrupción en la emisión infrarroja, la que se traduce en cambios de voltaje transmitido por el sensor. Se registró el valor de voltaje con una frecuencia de una muestra por cada 1 ms.
Procedimiento
La sesión experimental duró 60 minutos aproximadamente y consideró tres etapas: preparación, adaptación y entrenamiento. En la etapa de preparación, los participantes leyeron y firmaron un consentimiento informado, donde se enfatizó sobre la posibilidad de consultar acerca de cualquier aspecto vinculado al experimento. A continuación, se procedió a colocar los audífonos, el cintillo con el dispositivo de sensor infrarrojo y el tubo que arrojaba el soplo de aire al contorno del ojo de los participantes. Posteriormente, se les solicitó a los sujetos experimentales que parpadeasen repetidamente con el fin de obtener una respuesta de parpadeo detectable con el software. Asimismo, se les dio la instrucción de prestar atención a la película The Gold Rush (Chaplin, 1925; sin audio), enfatizando que posteriormente a la sesión experimental se les preguntaría sobre esta misma. Esta es una tarea de distracción utilizada frecuentemente en la investigación de condicionamiento de parpadeo en humanos (Clark & Squire, 1999). En la etapa de adaptación, se cerró la cámara experimental y los participantes fueron sometidos a un período de cinco minutos sin estimulación con el fin de que se aclimatasen al contexto novedoso. Posteriormente a ello, se ejecutó la siguiente fase experimental desde la sala de control en la computadora principal. En la etapa de entrenamiento, los participantes fueron expuestos a un total de 132 ensayos, de los cuales 66 ensayos correspondían al estímulo A que co-terminaba con el EI de 100 ms de duración (A+), entremezclados con 66 ensayos del estímulo B presentado por sí sólo, es decir, sin el EI (B-). El intervalo entre ensayos (ITI) varió aleatoriamente entre 15, 20 y 25 segundos. Para todos los participantes el estímulo A fue el tono y el estímulo B la vibración. Una vez concluido el entrenamiento, se formularon las preguntas respectivas a la película a los participantes, bajo una rúbrica general.
Análisis estadísticos
Con el fin de obtener una medida de la amplitud respecto de una respuesta palpebral típica en términos de cambio en el voltaje de la señal, se llevó a cabo un ensayo de normalización con un voluntario. El voltaje máximo obtenido con este participante durante los 200 ms siguientes del estímulo fue considerado como una respuesta con amplitud igual a 100. Las respuestas de todos los participantes se expresaron en relación con este valor estandarizado.
La Figura 1 describe los distintos eventos involucrados en un ensayo y durante el cual se midió la respuesta palpebral de acuerdo con el procedimiento descrito. Tal como se aprecia, los segmentos temporales para la medición de cada respuesta en ms fueron representados por medio de líneas discontinuas; en cambio la duración de los estímulos EC y EI, por medio de formas rectangulares. De lo anterior, se observa que la línea base (LB) o período de adaptación sin estimulación correspondió a los primeros 200 ms del ensayo. Las repuestas de parpadeo durante este periodo de LB fueron utilizadas como criterio para definir si un ensayo era considerado como válido. Específicamente, un ensayo se consideró válido si la respuesta palpebral no superó una amplitud de 20 unidades estandarizadas durante la LB. Una vez que un ensayo es considerado válido, cualquier respuesta emitida a partir del inicio del EC y que supere cinco unidades estandarizadas se consideró como una respuesta de parpadeo. El intervalo correspondiente a las Ra abarcó desde los 200 a 300 ms; las RV por su parte fueron ubicadas dentro del intervalo de 400 a 500 ms; las RC fueron registradas desde los 500 ms, concluyendo con la presentación del EI en los 600 ms subsiguientes. Finalmente, la RI fue medida a partir de la aparición del EI (600 ms) hasta los 800 ms del mismo ensayo.

Figura 1.
Parámetros de medición según amplitud y latencia de los distintos tipos de respuestas de parpadeo en un ensayo de condicionamiento.
Cada una de las respuestas definidas por su intervalo de ocurrencia fueron caracterizadas en términos de su amplitud promedio, duración, latencia y latencia para lanzar su máxima amplitud (peak). Cada una de estas variables fue analizada a través de un ANOVA de 4 medidas repetidas (respuestas alfa, voluntarias, condicionadas e incondicionadas).
Para analizar los efectos del entrenamiento discriminativo con los dos tipos de estímulos (A+ y B-), se agruparon los ensayos en tres bloques de 22 ensayos cada uno y se calculó el porcentaje de Rα, RV, RC y RI. La significancia estadística de estos efectos se examinó a través de un ANOVA de medidas repetidas de 2 (tipo de ensayo: A+ y B-) x 3 (bloque de entrenamiento: bloque 1 - bloque 3) para cada una de las respuestas descritas. Cuando la interacción ensayo x bloque resultó significativa, se examinaron los efectos simples de la variable estímulo en cada bloque.
Resultados
La presentación de los resultados fue dividida en dos secciones: 1) análisis de la forma predominante de las respuestas palpebrales y 2) frecuencias de los distintos tipos de respuestas a lo largo de entrenamiento discriminativo.
Análisis de la forma predominante de las respuestas palpebrales emitidas en condicionamiento discriminativo
En la Figura 2 se presentan ejemplos de la forma de los cuatro tipos de respuesta al estímulo A obtenidos en la presente investigación. Las Rα son caracterizadas como un parpadeo de baja amplitud; las RV se caracterizan principalmente por una amplitud notoriamente mayor; las RC son manifestadas en los ensayos con una amplitud inicialmente menor registrada momentos antes de la respuesta incondicionada hasta finalmente fusionarse con esta; finalmente, las RI son de gran amplitud.
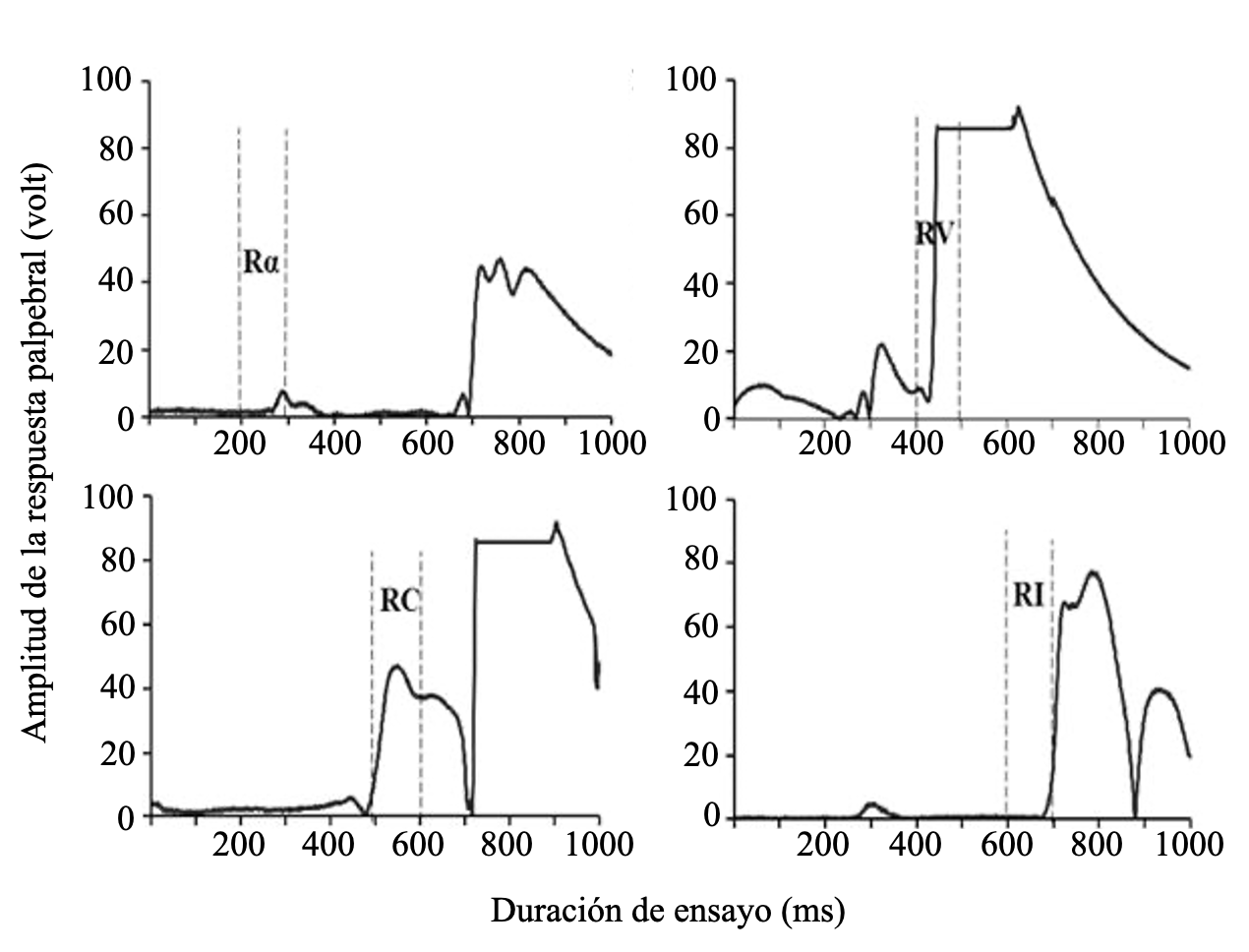
Figura 2.
Ejemplos de distintos tipos de respuestas palpebrales según su amplitud y latencia en ensayos de condicionamiento (A+).
Por supuesto, también existió variabilidad en la forma de las respuestas, las cuales se desviaron en mayor o menor grado de las formas características desplegadas en la Figura 2. Por ello, se clasificaron las respuestas en términos del momento en el cual se iniciaron dentro del ensayo (Figura 1). A partir de este encuadre, se calcularon los promedios de la amplitud, latencia, latencia de su máxima amplitud y duración de las respuestas que ocurrieron dentro de cada uno de los intervalos temporales.
En la Figura 3 se muestra tanto el promedio de amplitud (panel A), latencia (panel B), latencia para alcanzar la máxima amplitud o peak (panel C) así como duración de cada tipo de respuesta (panel D) al estímulo A. Es posible visualizar en el panel A que la amplitud tiende a aumentar linealmente, desde la Rα (la de menor amplitud) hasta la RI (la de mayor amplitud). Esta tendencia se confirma parcialmente dado que el ANOVA de medidas repetidas arrojó un efecto principal para el tipo de respuestas F(3, 21) = 4.510, p = .013, np2 = .395. Sin embargo, las pruebas post hoc indicaron que solo hay diferencias significativas entre la amplitud de las RI y Rα (p = 0.010). Lo anterior, probablemente debido a que una gran cantidad de ensayos de condicionamiento produjo RC y RV de alta amplitud, así como RI que disminuyen su amplitud como consecuencia de la repetición del EI, es decir, en proceso de habituación.
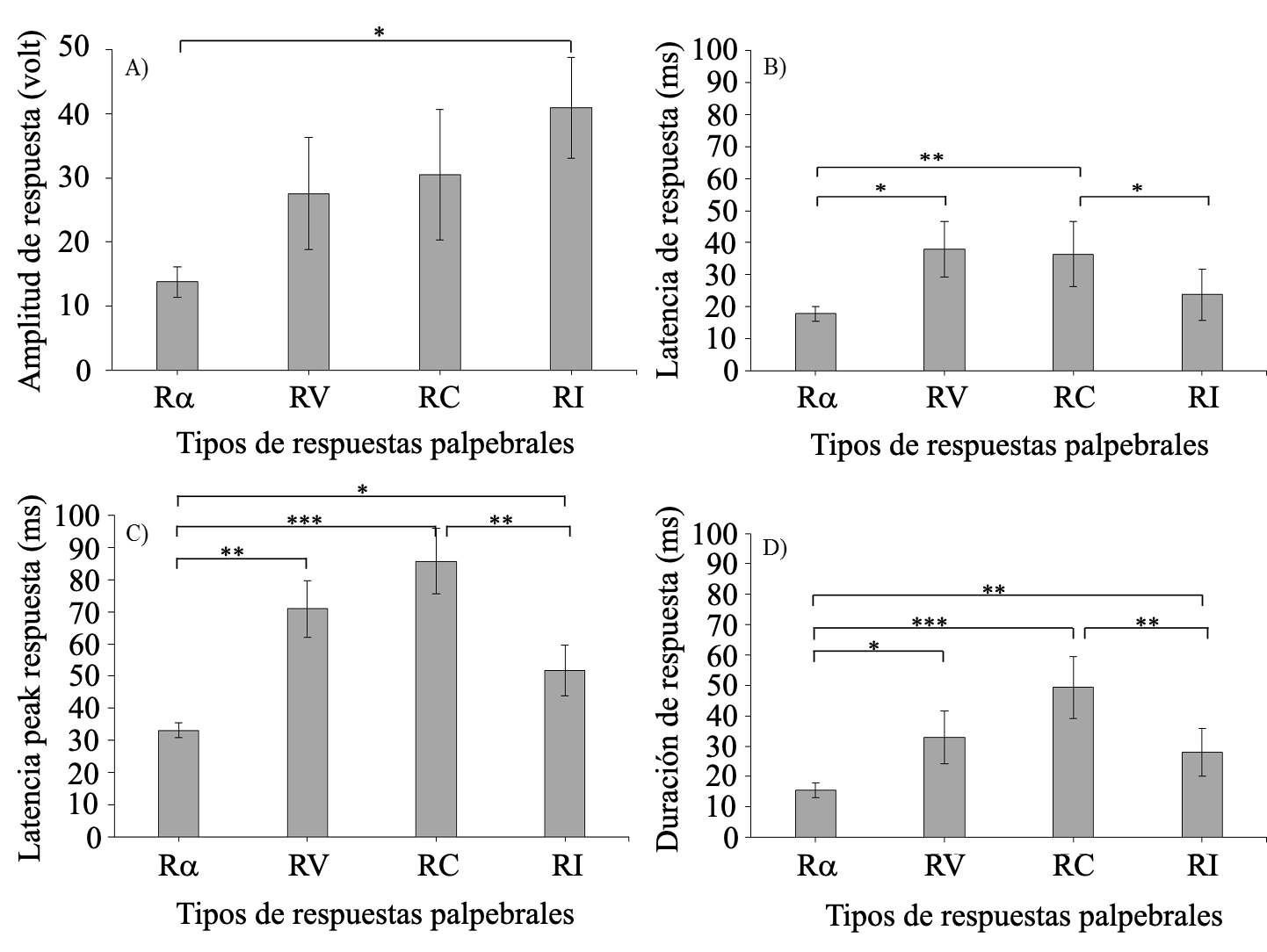
Figura 3.
Promedios de amplitud (panel A), latencia (panel B), latencia máxima o peak (Panel C) y duración (panel D) de la respuesta en ensayos del tipo A+.
Respecto a la latencia, en el panel B de la Figura 3 se observa que las Rα y RI ocurren tempranamente, mientras que las RV y RC son comparativamente más tardías. El ANOVA de medidas repetidas confirmó esta observación arrojando un efecto principal significativo F(3, 21) = 3.403, p = .037, np2 = .327. Las comparaciones post hoc indicaron que las RV y RC no difieren significativamente entre sí (p = .825) pero ambas son significativamente más tardías que las Rα (p = .036 y p = .008, respectivamente). Adicionalmente, las RC son significativamente más tardías que las RI (p = .049), diferencia que no alcanza la significancia para el caso de las RV (p = .245). Por otra parte, las Rα y RI no difieren significativamente en su latencia (p = .429).
En cuanto a la latencia para alcanzar el máximo dentro de sus respectivos intervalos, panel C de la Figura 3, se aprecia una tendencia similar a la latencia; donde las Rα parecieran ser aquellas que alcanzan su máxima amplitud (latencia peak) con mayor rapidez, seguidas por las RI, RV y RC. El ANOVA confirma esta tendencia ya que arrojó un efecto principal F(3, 21) = 12.005, p < .001, np2 = .632. Específicamente, las Rα presentan una latencia peak significativamente menor que las RI (p = .042), RV (p = .004) y RC (p < .001). Por otra parte, las RI también alcanzaron la máxima amplitud con mayor rapidez, aunque esta no difirió significativamente de las RV (p = 0.189), pero fue significativamente más rápido que las RC (p = .003); mientras que las RC y RV no difirieron significativamente entre sí (p = .225).
Finalmente, con respecto a la duración de las respuestas (Panel D), se calculó como la diferencia entre latencia para alcanzar la máxima amplitud y latencia. Nuevamente las respuestas mostraron diferencias significativas entre sí F(3, 21) = 12.664, p < .001, np2 = .644. Específicamente, las Rα fueron más breves, difiriendo significativamente de las RI (p = .007), RV (p = .013) y RC (p < .001). La duración de las RI fue significativamente menor que las RC (p = .008) y menor, aunque no significativamente, que las RV (p = 0.377). Nuevamente se encontró que las RV y RC no difieren significativamente entre sí (p = .065).
En resumen, de esta primera parte se puede extraer que los datos de la Figura 3 y su correspondiente análisis estadístico indicaron que las Rα y RI tienden a coincidir respecto de sus parámetros temporales (son más rápidos y de menor duración), pero difieren en su amplitud (Ra bajas y RI altas). Por otra parte, las RV y RC son más lentas y duraderas que las anteriores.
Análisis de las frecuencias de los distintos tipos de respuestas a lo largo del entrenamiento discriminativo
Es posible detectar las diferencias en las respuestas palpebrales de distinto tipo emitidas durante los ensayos, tanto para EC reforzados (A+) como no reforzados (B-). Para cada uno de estos casos, la variable dependiente se encuentra asociada a la proporción de respuestas bajo un análisis de ANOVA de medidas repetidas de 2 (tipo ensayo: A+ y B-) x 3 (bloques de entrenamiento: 1-3).
Respuestas Alfa.
En la Figura 4 se muestra la proporción de Ra a lo largo de los tres bloques de entrenamiento discriminativo para los ensayos A+ y B-. Los datos de la figura indican que al comienzo del entrenamiento existió un mayor número de Ra en los ensayos A+, las cuales disminuyeron con el entrenamiento. Consistente con ello, el ANOVA no arrojó efectos principales significativos de tipo de estímulos F(1, 14) = 3.987, p = .086, np2 = .363, ni de bloque F(2, 14) = 2.151, p = .153, np2 = .235, pero sí una interacción significativa entre tipo de ensayo y bloque F(2, 14) = 4.738, p = .027, np2 = .404. Consecuente con esta interacción, los efectos simples revelaron un efecto bloque significativo para los ensayos A+ (entre los bloques 1 y 2, p = .05), pero no en los ensayos B- (p > .807). El efecto nulo de la variable tipo de estímulo confirma que no hay diferencias en la frecuencia de las Ra emitidas en los dos tipos de ensayos (A+ y B-). Por otra parte, el efecto nulo de la variable bloque indica que no existe una disminución global significativa de las Ra a lo largo del entrenamiento. Sin embargo, la presencia de interacción significativa hace este análisis un tanto más complejo, pudiéndose especular que las Ra emitidas en presencia del estímulo no reforzado (B-) son de muy baja frecuencia (quizá a nivel de un efecto suelo), y que Ra emitidas en presencia de A+ disminuyan con el entrenamiento, como lo revelan las diferencias significativas entre los bloques 1 y 2.
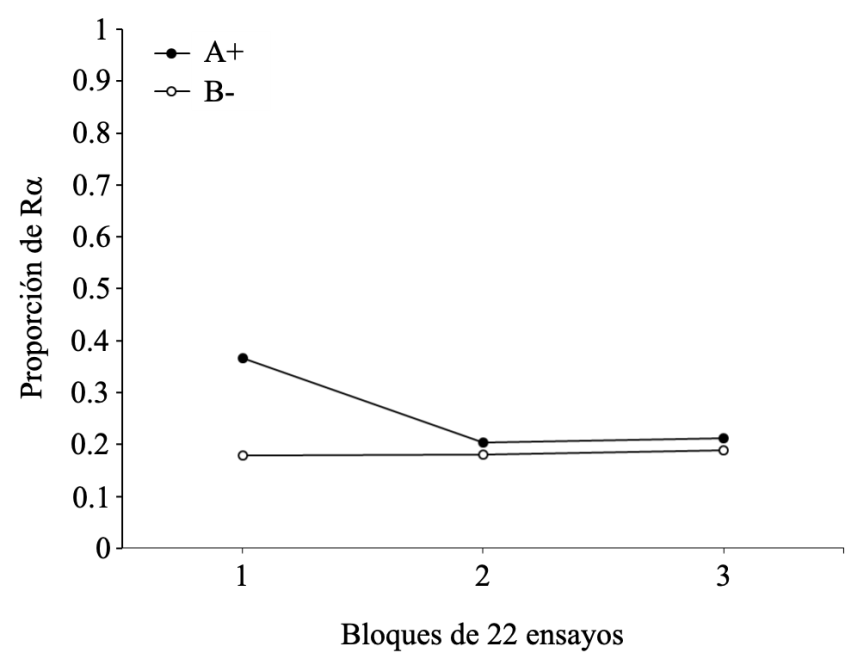
Figura 4
Proporción de Ra para los estímulos A+ y B- según los bloques de ensayos del entrenamiento discriminativo
Respuestas Condicionadas
En la Figura 5 se reporta la proporción de RC a lo largo de los tres bloques de entrenamiento discriminativo para los ensayos A+ y B-. La RC emitida ante el estímulo A+ presentó un progresivo aumento a lo largo de los tres bloques de entrenamiento. Por el contrario, las RC al estímulo B- son muy bajas a lo largo de los tres bloques, indicando la adquisición de discriminación por parte de los participantes. Es importante destacar que pese a que las respuestas a B- tienen una baja proporción, esta varía a lo largo del entrenamiento con un aumento del bloque 1 al bloque 2 y un posterior descenso a un nivel más bajo en el bloque 3. Esto reporta un hallazgo típico asociado a situaciones de discriminación, donde se visualiza un aumento inicial de la RC al estímulo no reforzado producto de la generalización entre los estímulos A y B. El ANOVA confirma estas observaciones puesto que arrojó un efecto principal de tipo de ensayos F(1, 14) = 5.545, p = .050, np2= .442, un efecto principal de bloque F(2, 14) = 4.713, p = .027, np2 = .402 y una ausencia de interacción significativa entre tipo de ensayo y bloque F(2, 14) = 3.260, p = .068, np2 = .318. Pese a que la interacción resultó no significativa, se examinaron los efectos simples, los cuales revelan que las diferencias en la frecuencia de RC a los estímulos A y B no son significativas en el bloque 1 (p = .082) y en el bloque 2 (p = .222), pero significativas en el bloque 3 (p = .015). En conclusión, el efecto principal del tipo de ensayo confirma la existencia de diferencias significativas en la frecuencia de RC emitidas en los ensayos con el EC reforzado (A+) así como con el EC no reforzado (B-); asimismo el efecto principal de bloque confirma la existencia de un aumento significativo en la frecuencia de RC emitidas al EC reforzado (A+) a lo largo de los ensayos de entrenamiento discriminativo.
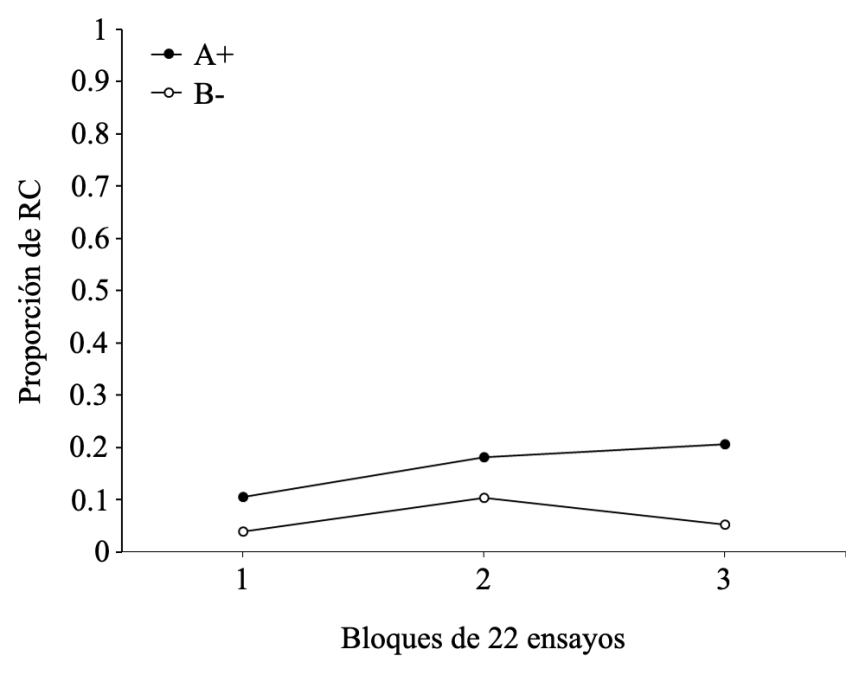
Figura 5.
Proporción de RC para los estímulos A+ y B- según los bloques de ensayos del entrenamiento discriminativo
Respuestas Voluntarias
En la Figura 6 se muestra la proporción de RV a lo largo de los tres bloques de entrenamiento discriminativo para los ensayos A+ y B-. Como se puede observar, las RV continúan un patrón similar a las RC descritas en el apartado anterior. Es decir, las respuestas al estímulo A+ aumentan progresivamente a lo largo de los tres bloques de entrenamiento y son siempre superiores a las repuestas al estímulo B-, lo cual es evidencia de discriminación. En este caso, las respuestas a B- son uniformemente más bajas que el caso de A+, probablemente debido al hecho de que, al ser RV y no automáticas, la discriminación se aprende más rápidamente. Consistente con lo anterior, el ANOVA arrojó un efecto principal del tipo de ensayos F(1, 14) = 9.300, p = .019, np2 = 0.571, un efecto principal de bloque F(2, 14) = 12.637, p = .001, np2 = 0.644, y una interacción no significativa entre tipo de ensayo y bloque F(2, 14) = 3.300, p = 0.067, np2 = 0.320. Al igual que en el caso de la RC, en esta oportunidad se examinaron los efectos simples, encontrando que la superioridad de A+ sobre B- se manifiesta tanto en el bloque 1 (p = .004) como en el bloque 2 (p < .001) y bloque 3 (p <.001). De este modo, el efecto principal de tipo de ensayo confirma la existencia de diferencias significativas en la frecuencia de RV emitidas en los ensayos con el EC reforzado (A+) así como con el EC no reforzado (B-); donde el efecto principal de bloque confirma la existencia de un aumento significativo en la frecuencia de RV emitidas al EC reforzado (A+) a lo largo de los ensayos de entrenamiento discriminativo.
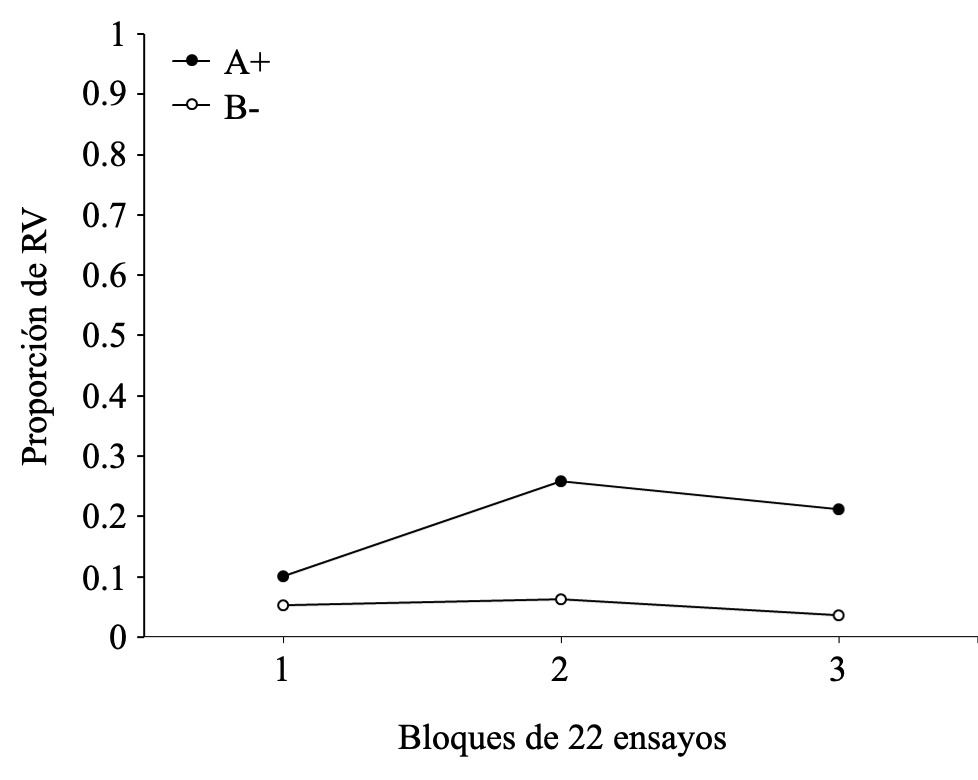
Figura 6
Proporción de RV para los estímulos A+ y B- según los bloques de ensayos del entrenamiento discriminativo.
Respuestas Incondicionadas
En la Figura 7 se reporta la proporción de RI a lo largo de los tres bloques de entrenamiento discriminativo para los ensayos A+ y B-. Como se puede observar, existe una gran diferencia en la frecuencia de respuestas a ambos estímulos con una mayor proporción de respuestas al estímulo A+. Dado que el estímulo B- no recibe el EI, la proporción de respuestas que pueden observarse podrían ser explicadas por un remanente de RC y RV hacia este estímulo. Como era esperable, el ANOVA arrojó solo un efecto principal significativo de tipo de ensayos F(1, 14) = 63.166, p < .001, np2 = .9. El hecho de que no exista un efecto principal de bloque F(2, 14) = 2.724, p = .100, np2 = .280, ni interacción entre tipo de ensayo y bloque F(2, 14) = 0.218, p = .807, np2 = .030 indica la ausencia de una disminución significativa en el porcentaje de RI a lo largo de los ensayos de entrenamiento discriminativo.
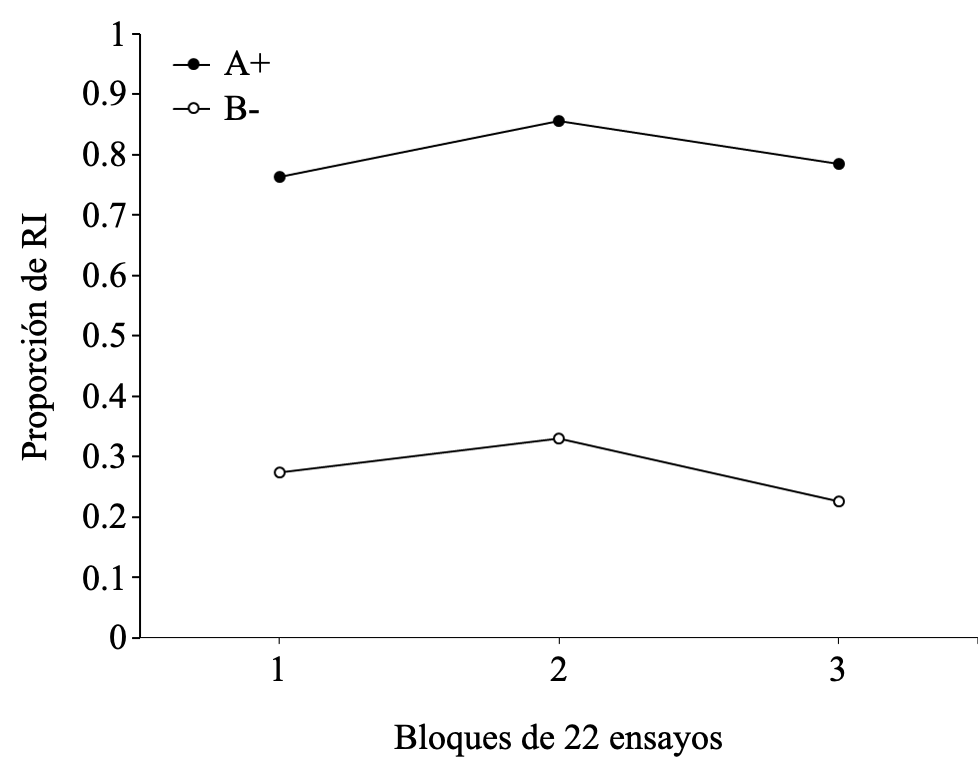
Figura 7
Proporción de RI para los estímulos A+ y B- según los bloques de ensayos del entrenamiento discriminativo
Discusión
La presente investigación demostró que es posible implementar un sistema para detectar los distintos tipos de respuestas de parpadeo que emiten las personas durante el condicionamiento discriminativo. Los resultados indicaron que las Ra y RI difieren en amplitud y duración, siendo las primeras de menor tamaño y menor duración. A su vez, la RI resultó ser también de mayor amplitud (aunque las diferencias no alcanzaron la significancia estadística) y menor duración que las RV y RC. Esto fue consistente con la idea de que las Rα y RI son respuestas reflejas a los EC y EI, respectivamente y que por ello ocurren con mayor rapidez que sus contrapartes RV y RC. Para el caso de las RV y RC, estas fueron similares entre sí en términos de amplitud y duración. Estableciendo, por consiguiente, velocidades de ocurrencia más tardías e intervalos de duración de mayor extensión que las propias Rα y RI.
Por otro lado, se examinó el efecto del entrenamiento discriminativo en los tipos de respuesta de parpadeo. En el caso de las Rα, se observó que al inicio del entrenamiento existe una mayor tasa de este tipo de respuestas emitidas para ensayos con A+, las cuales disminuyen con el paso del tiempo. Sin embargo, no se observó ningún cambio para el estímulo B-. En el caso de las RC al estímulo A+, estas presentan un progresivo aumento a lo largo de los tres bloques de entrenamiento, lo cual es un reflejo de aprendizaje. Por el contrario, en el caso del estímulo B-, se presentan tasas de respuesta muy bajas a lo largo de los tres bloques, sobre todo hacia el bloque 3. En el caso de las RV es posible observar cómo estas siguen un patrón muy similar al de las RC, es decir, aumentan progresivamente a lo largo del entrenamiento en el caso de los ensayos A+ pero no en los ensayos B-. Finalmente, para el caso de las RI, al existir un estímulo que produce una respuesta refleja, se establece una discriminación temprana entre A+ y B-.
A pesar del potencial de los presentes resultados, por su contribución a la detección y medición de la RC de parpadeo en humanos, debe considerarse que el presente estudio tiene las siguientes limitaciones o dificultades metodológicas: a) el procedimiento de agrupación de ensayos en tres bloques (22 ensayos por bloque) podría oscurecer cambios sensibles en las respuestas en los primeros ensayos; b) la no realización de contrabalanceos entre los estímulos A y B; c) en el condicionamiento discriminativo, el EC- está explícitamente no emparejado con el EI, es decir, se introduce una nueva contingencia donde el EC- puede ser visto como una señal de la ausencia del EI (Rescorla, 1967); d) el empleo del vibrador en la espalda podía ser evitado en su totalidad si el participante cambiaba su posición en el sillón masajeador. Además de solucionar las limitaciones metodológicas señaladas, futuras investigaciones podrían replicar los análisis presentados en esta investigación utilizando un mayor tamaño muestral; un repertorio más amplio de EC tales como ruidos, luces y el uso de un EI inevitable, tal como una estimulación eléctrica de baja intensidad, lo cual probablemente podría reducir la incidencia de RV.
En síntesis, para investigar condicionamiento en humanos sin duda se requiere de cuidadosos análisis y descripción de la topografía de las RC y su despliegue temporal en relación con la RI, en condiciones donde se minimice la posibilidad que las personas hagan inferencias acerca de la relación entre los estímulos (por ejemplo, introduciendo carga cognitiva). Esto podría arrojar claves para evaluar con exactitud cuando una respuesta es el producto de un proceso declarativo o no declarativo.
Agradecimientos
El presente estudio fue financiado por el Fondo Nacional de Desarrollo Científico y Tecnológico (Fondecyt) de la Agencia Nacional de Investigación y Desarrollo de Chile (proyecto Nº 1210719, investigador responsable Edgar Vogel).
Referencias
Bolaram, A., Coe, T. E., Power, J. M., & Cheng, D. T. (2020). Awareness and differential eyeblink conditioning: effects of manipulating auditory CS frequencies. Learning & Memory, 27(2), 78-82. doi: 10.1101/lm.050146.119
Brown, P., & Jenkins, H. (1968). Auto-shaping the pigeon’s key-peck. Journal of the Experimental Analysis of Behavior, 11(1), 1-8. doi: 10.1901/jeab.1968.11-1
Chaplin, C. (1925). The Gold Rush [Película]. Estados Unidos: Charles Chaplin Productions.
Christian, K. M., & Thompson, R. F. (2003). Neural substrates of eyeblink conditioning: acquisition and retention. Learning & Memory, 10(6), 427-455. doi: 10.1101/lm.59603
Clark, R. E., Manns, J. R., & Squire, L. R. (2002). Classical conditioning, awareness, and brain systems. Trends in Cognitive Sciences, 6(12), 524-531. doi: 10.1016/S1364-6613(02)02041-7
Clark, R. E., & Squire, L. R. (1999). Human eyeblink classical conditioning: Effects of manipulating awareness of the stimulus contingencies. Psychological Science, 10(1), 14–18. doi: 10.1111/1467-9280.00099
Estes, W., & Skinner, B. (1941). Some quantitative properties of anxiety. Journal of Experimental Psychology, 29(5), 390-400. doi: 10.1037/h0062283
García, J., & Koelling, R. (1966). Relation of cue to consequence in avoidance learning. Psychonomic Science, 4(1), 123-124. doi: 10.3758/BF03342209
Goodrich, K. (1966). Experimental analysis of response slopes and latency as criteria for characterizing voluntary and nonvoluntary responses in eyeblink conditioning. Psychological Monographs: General and Applied, 80(14), 1-34. doi: 10.1037/h0093904
Gormezano, I. (1965). Yoked comparisons of classical and instrumental conditioning of the eyelid response: An addendum on “voluntary responders”. En W. F. Prokasy (Ed). Classical conditioning (pp. 47-70). New York: Appleton-Century-Croffts.
Gormezano, I., Schneiderman, N., Deux, E., & Fuentes, I. (1962). Nictitating membrane: Classical conditioning and extinction in the albino rabbit. Science, 138(3536), 33-34. doi: 10.1126/science.138.3536.33
Grant, D. A. (1945). A sensitized eyelid reaction related to the conditioned eyelid response. Journal of Experimental Psychology, 35(5), 393–402. doi: 10.1037/h0057865
Grant, D. A., & Norris, E. B. (1947). Eyelid conditioning as influenced by the presence of sensitized Beta-responses. Journal of Experimental Psychology, 37(5), 423–433. doi: 10.1037/h0054090
Grant, D., Norris, E., & Hornseth, J. (1948). Sensitization of the Beta response under verbally induced inhibitory set and counter-reinforcement. American Journal of Psychology, 61(1), 66-72. doi: 10.2307/1417293
Hilgard, E., & Campbell, A. (1936). The course of acquisition and retention of conditioned eyelid responses in man. Journal of Experimental Psychology, 19(2), 227-247. doi: 10.1037/h0055600
Huang, H., Wu, B., Li, Q., Yao, J., Li, X., Yang, Y., Wu, G.-Y., & Sui, J. F. (2014). Awareness is essential for differential delay eyeblink conditioning with soft-tone but not loud-tone conditioned stimuli. Neuroscience Bulletin, 30(3), 433-440. doi: 10.1007/s12264-013-1400-5
Knuttinen, M.-G., Power, J. M., Preston, A. R., & Disterhoft, J. F. (2001). Awareness in classical differential eyeblink conditioning in young and aging humans. Behavioral Neuroscience, 115(4), 747–757. doi: 10.1037/0735-7044.115.4.747
Lovibond, P. F., Liu, J. C., Weidemann, G., & Mitchell, C. J. (2011). Awareness is necessary for differential trace and delay eyeblink conditioning in humans. Biological Psychology, 87(3), 393-400. doi: 10.1016/j.biopsycho.2011.05.002
Lovibond, P. F., & Shanks, D. R. (2002). The role of awareness in Pavlovian conditioning: Empirical evidence and theoretical implications. Journal of Experimental Psychology: Animal Behavior Processes, 28(1), 3–26. doi: 10.1037/0097-7403.28.1.3
Manns, J. R., Clark, R. E., & Squire, L. R. (2000). Awareness predicts the magnitude of single‐cue trace eyeblink conditioning. Hippocampus, 10(2), 181-186. doi: 10.1002/(SICI)1098-1063(2000)10:2<181::AID-HIPO7>3.0.CO;2-V
Martin, I., & Levey, A. (1969). The genesis of the classical conditioned response. New York: Pergamon Press.
McLaren, I. P., Forrest, C. L. D., McLaren, R. P., Jones, F. W., Aitken, M. R. F., & Mackintosh, N. J. (2014). Associations and propositions: The case for a dual-process account of learning in humans. Neurobiology of Learning and Memory, 108, 185-195. doi: 10.1016/j.nlm.2013.09.014
Mitchell, C. J., De Houwer, J., & Lovibond, P. F. (2009). The propositional nature of human associative learning. Behavioral and Brain Sciences, 32(2), 183–246. doi: 10.1017/S0140525X09000855
Pavlov, I. P. (1927). Conditioned reflexes. Oxford: Oxford University Press.
Rasmussen, A., & Jirenhed, D. A. (2017). Learning and timing of voluntary blink responses match eyeblink conditioning. Scientific Reports, 7, 3404. doi: 10.1038/s41598-017-03343-2
Rescorla, R. A. (1967). Pavlovian conditioning and its proper control procedures. Psychological Review, 74(1), 71–80. doi: 10.1037/h0024109
Rescorla, R. A., & Wagner, A. R. (1972). A theory of Pavlovian conditioning: Variations in the effectiveness of reinforcement and nonreinforcement. En A. H. Black & W. F. Prokasy (Eds.), Classical conditioning II: Current theory and research (pp. 64–99). New York: Appleton Century Crofts.
San Diego Instruments (equipos y software, 2022). Eyeblink Conditioning System. San Diego: San Diego Instruments.
Schacter, D. L., & Tulving, E. (1994). What are the memory systems of 1994? En D. L. Schacter & E. Tulving (Eds.), Memory systems 1994 (pp. 1–38). Cambridge: The MIT Press.
Schade Powers, A., Coburn-Litvak, P., & Evinger, C. (2010). Conditioned eyelid movement is not a blink. Journal of Neurophysiology, 103(2), 641–647. doi: 10.1152/jn.00631.2009
Shanks, D. R. (2010). Learning: From association to cognition. Annual Review of Psychology, 61(1), 273-301. doi: 10.1146/annurev.psych.093008.100519
Spence, K. W., & Ross, L. E. (1959). A methodological study of the form and latency of eyelid responses in conditioning. Journal of Experimental Psychology, 58(5), 376–381. doi: 10.1037/h0045837
Squire, L. R. (1994). Declarative and nondeclarative memory: Multiple brain systems supporting learning and memory. En D. L. Schacter & E. Tulving (Eds.), Memory systems 1994 (pp. 203–231). Cambridge: The MIT Press.
Vogel, E., Brandon, S., & Wagner, A. (2003). Stimulus representation in SOP: - II. An application to inhibition of delay. BehaviouralProcesses, 62(1-3), 27-48. doi: 10.1016/S0376-6357(03)00050-0.
Vogel, E., & Ponce, F. (2011). Empirical Issues and Theoretical Mechanisms of Pavlovian Conditioning. En E. Alonso & E. Mondragón (Eds.), Computational Neuroscience for Advancing Artificial Intelligence: Models, Methods and Applications (pp. 81-110). New York: Medical Information Science Reference IGI Global.
Vogel, E. H., Ponce, F. P., & Wagner, A. R. (2019). The development and present status of the SOP model of associative learning. Quarterly Journal of Experimental Psychology, 72(2), 346-374. doi: 10.1177/1747021818777074
Wagner, A. R., & Vogel, E. H. (2008). Configural and elemental processing in associative learning: Commentary on Melchers, Shanks and Lachnit. BehaviouralProcesses, 77(3), 446-450. doi: 10.1016/j.beproc.2007.09.011
Weidemann, G., & Antees, C. (2012). Parallel acquisition of awareness and differential delay eyeblink conditioning. Learning & Memory, 19(5), 201-210. doi: 10.1101/lm.024851.111
Weidemann, G., Best, E., Lee, J. C., & Lovibond, P. F. (2013). The role of contingency awareness in single-cue human eyeblink conditioning. Learning & Memory, 20(7), 363-366. doi: 10.1101/lm.029975.112
Weidemann, G., Satkunarajah, M., & Lovibond, P. F. (2016). I think, therefore eyeblink: The importance of contingency awareness in conditioning. Psychological Science, 27(4), 467-475. doi: 10.1177/0956797615625973
Weidemann, G., Tangen, J. M., Lovibond, P. F., & Mitchell, C. J. (2009). Is Perruchet’s dissociation between eyeblink conditioned responding and outcome expectancy evidence for two learning systems? Journal of Experimental Psychology: Animal Behavior Processes, 35(2), 169–176. doi: 10.1037/a0013294
Notas de autor
Enviar correspondencia a: Pinto, J. A. E-mail: jpinto11@santotomas.cl
Información adicional
Citar este artículo como: Tapia, L. H., Becerra, S. A. y Pinto, J. A. (2022). Caracterización de los tipos de respuesta de parpadeo en un procedimiento de condicionamiento discriminativo en humanos. Revista Argentina de Ciencias del Comportamiento, 14(3), 93-103
Enlace alternativo
https://revistas.unc.edu.ar/index.php/racc/article/view/32109/40193 (pdf)
