Comunicaciones
CULTIVO DE MANDIOCA (Manihot esculenta CRANTZ) EN INVERNADERO: EFECTOS SOBRE EL RENDIMIENTO Y LA CALIDAD DE RAÍCES TUBEROSAS
CASSAVA GROWING IN GREENHOUSE: EFFECTS ON YIELD AND QUALITY OF STORAGE ROOTS
CULTIVO DA MANDIOCA EM ESTUFA: EFEITOS SOBRE A PRODUÇÃO E QUALIDADE DE RAÍZES TUBEROSAS
CULTIVO DE MANDIOCA (Manihot esculenta CRANTZ) EN INVERNADERO: EFECTOS SOBRE EL RENDIMIENTO Y LA CALIDAD DE RAÍCES TUBEROSAS
Interciencia, vol. 42, núm. 8, pp. 515-521, 2017
Asociación Interciencia
Recepción: 04 Noviembre 2016
Aprobación: 22 Julio 2017
Resumen: Dada la fuerte interacción entre la temperatura ambiente y el rendimiento de raíces tuberosas (RT) de mandioca, en regiones subtropicales el cultivo bajo invernadero puede ser una tecnología de producción a considerar si se quiere incrementar el rendimiento y adelantar la cosecha. El objetivo de este trabajo fue evaluar cuantitativa y cualitativamente la producción de RT de mandioca bajo invernadero en un suelo arenoso de Corrientes, Argentina. Para ello se estudiaron variaciones temporales de componentes del rendimiento; índice de cosecha (IC); y contenidos de almidón, proteínas, fibra bruta y minerales de RT de tres cultivares (Blanca de Santa Catarina, Ramada Paso y Rocha). El experimento se condujo mediante un diseño en bloques completos al azar, con tres repeticiones. El peso fresco de RT por planta mostró una interacción genotipo-tiempo significativa. El número de RT se mantuvo en el tiempo en cada cultivar, aunque se presentaron diferencias entre genotipos. A los 193 días después de plantación, los IC superaron valores de 0,5. Los contenidos de almidón no difirieron entre genotipos, siendo más precoces Blanca de Santa Catarina y Rocha. Se encontraron correlaciones negativas entre tiempo y contenidos de proteína y fibra bruta, y una asociación muy alta entre éstos. No hubo diferencias estadísticas en contenidos de minerales en el tiempo. En conclusión, es factible producir mandioca bajo invernadero alcanzando rendimientos elevados y cosechas tempranas.
Palabras clave: Cultivo protegido, Mandioca, Manihot esculenta, Raíces tuberosas, Tapioca.
Abstract: Due to the strong interaction between ambient temperature and cassava yields, in subtropical regions greenhouse culture can be considered as an appropriate production technology in order to increase root yield and anticipate harvest. The aim of this work was to evaluate quantitatively and qualitatively the production of cassava storage roots (SR) in a sandy soil of Corrientes, Argentina, under greenhouse. For this purpose, temporal variations of yield components, harvest index (HI), contents of starch, protein, crude fiber and minerals in SR of three cassava cultivars (Blanca de Santa Catarina, Ramada Paso and Rocha) were studied. The experiment was conducted under a randomized complete block design with three replications. SR fresh weight per plant showed a significant genotype-time interaction. The number of SR significantly varied between cultivars and remained constant over time in each one of them. All IC obtained exceeded 0.5 after 193 days of plantation. The starch content did not differ between genotypes, while Blanca de Santa Catarina and Rocha yielded the earliest. There were negative relationships between time and protein content and crude fiber, and a very high association between the last ones. No statistical differences in mineral content with respect to time were recorded. It is feasible to produce cassava in greenhouses achieving high yields and early harvests.
Resumo: Dada a forte interação entre a temperatura ambiente e a produtividade de raízes tuberosas (RT) de mandioca, em regiões subtropicais o cultivo em estufas pode ser uma tecnologia de produção a considerar para aumentar o desempenho e antecipar a colheita. O objetivo deste trabalho foi avaliar quantitativamente e qualitativamente a produção de mandioca RT na estufa num solo arenoso de Corrientes, Argentina. Para este efeito, foram estudadas variações temporais de componentes de produção; índice de colheita (IC); teor de amido, proteínas, fibra bruta e minerais RT de três cultivares (Blanca de Santa Catarina, Ramada Paso e Rocha). O delineamento experimental foi em blocos completos casualizados com 3 repetições. O Peso fresco de RT por planta mostrou uma interação genótipo-tempo significativa. O número de RT foi mantido ao longo do tempo em cada cultivar, embora apresentasse diferenças genotípicas. Nos 193 dias após o plantio, os IC foram maiores que 0,5. Os teores de amido não diferiram entre os genótipos, sendo acumulação mais rápida em Branca de Santa Catarina e Rocha. Houve correlação negativa entre tempo e conteúdo de proteína e fibra bruta e uma associação muito grande entre eles. Não houve diferenças no conteúdo mineral ao longo do tempo. É viável a produção de mandioca em estufas para alcançar altos rendimentos e colheitas adiantadas.
Introducción
La mandioca (Manihot esculenta Crantz) constituye la cuarta fuente de calorías en la dieta de pobladores de Latinoamérica, África y Asia (Techeira et al., 2014), siendo las raíces tuberosas amiláceas el órgano de interés comercial por excelencia. Este cultivo tiene además importancia en términos de agricultura sustentable, con potencial para integrar sistemas productivos de alimentos de baja agresión al medio ambiente en climas tropicales y subtropicales, y suelos de baja fertilidad (Alves, 2002; Howeler, 2014). El rol más importante que asume este cultivo en muchos países en desarrollo es el de garantizar la seguridad alimentaria basado en su rusticidad (Burns et al., 2012).
Argentina es el más austral entre los países productores de mandioca de Latinoamérica. Se la cultiva en la región nordeste del país, en un estrato de pequeños productores asociados en cooperativas que alcanzan un ámbito de comercialización principalmente nacional, sin llegar a lograr satisfacer la demanda a nivel país, mostrando un valor comercial bastante escaso (Bongiovanni et al., 2012). Dado que las condiciones agroecológicas regionales son propicias para obtener altos rendimientos, la industrialización del rubro mejoraría la rentabilidad de la cadena agroindustrial y podría abastecer un mercado insatisfecho. Si en Argentina se considerase a las raíces de mandioca como un producto estratégico para el desarrollo de numerosas industrias y se le otorgara el tratamiento correspondiente en términos de inversiones, muy certeramente podría darse un vuelco en el desarrollo del sector agroalimentario e industrial regional, contribuyendo a la generación de riqueza y de empleo rural y urbano, con el valioso sello del valor agregado en origen de productos diversificados, originales e innovadores.
En términos agronómicos de cultivo, los más altos rendimientos de raíces y de almidón, se registran en climas tropicales, con temperaturas entre 25-29ºC (Alves, 2002), mientras que en regiones subtropicales, como es el caso del Nordeste argentino, se retrasa su crecimiento dado que las bajas temperaturas invernales ponen fin al ciclo de crecimiento anual. Esta respuesta se da porque el patrón de crecimiento y desarrollo fenológico de la mandioca se explica a través de la suma térmica acumulada durante el ciclo (Burgos et al., 2013). Bajo estas circunstancias, la planificación de cultivos protegidos de mandioca en invernaderos plásticos sería una alternativa para conseguir cosechas tempranas, con un elevado rendimiento de raíces.
Por ello, se planteó como objetivo evaluar el potencial de rendimiento de tres cultivares de mandioca bajo invernadero, a través del análisis de sus componentes, y analizar el contenido de almidón de las raíces frescas y la concentración de proteínas, fibra bruta y minerales (P, K, Ca, Mg y Na) de las harinas de raíces tuberosas, en función del tiempo de cultivo.
Materiales y Métodos
El experimento se llevó a cabo en la localidad de Corrientes, Argentina (27º28’ 27.23’’S; 58º47’00.6’’O; altitud de 50msnm), donde el clima se caracteriza por presentar precipitaciones promedio anuales de 1300mm, y temperaturas medias anuales de 21,6ºC. El período libre de heladas es de 340 a 360 días por año y la frecuencia de ocurrencia de las mismas es de 0,5. Tomando la clasificación de Köppen modificada, el clima en la región se clasifica como mesotermal húmedo, Cf w’a (h) (Murphy, 2008). El suelo es clasificado como Udipsamment árgico, familia mixta, hipertérmica, perteneciente a la Serie Ensenada Grande, presenta una granulometría gruesa en superficie, mediano a débilmente ácido en el horizonte A, arenoso (Soil Survey Staff, 1990). Son suelos de baja fertilidad, baja capacidad de intercambio catiónico, pero con buenas condiciones físicas asociadas a su textura arenosa.
El experimento se condujo en un invernadero de 25× 8×5m, en forma de arco redondeado con paredes verticales, cubierto con plástico de 100 y 150um de larga duración térmica y con ventilación lateral. La ubicación del invernadero y de los liños dentro del mismo sigue la transecta N-S. El riego fue suministrado por goteo superficial y las malezas se controlaron mediante carpidas manuales. Se evaluaron tres cultivares de mandioca de interés para Argentina: Blanca de Santa Catarina, Ramada Paso y Rocha.
El diseño experimental fue en bloques completos al azar, con tres repeticiones. La plantación de estacas se realizó el 1/08/2012 en un marco de 1×1m, lo que representa una densidad de 10.000 plantas/ha. Las plantas se cosecharon mensualmente entre diciembre/2012 y abril/2013. Para estudiar el rendimiento final de raíces frescas (Tn·ha1 ), se cuantificaron los componentes del mismo: el número de raíces tuberosas (Nº RT/planta) y el peso fresco individual de las mismas (PFiRT, kg). Posteriormente por cultivar se tomaron tres muestras de 3000g de raíces que luego se pesaban en agua y por el método de la gravedad específica se obtuvieron los valores de almidón (%) según Toro y Cañas (1983).
En cada muestreo, 10 raíces tuberosas de tamaño comercial (longitud ≥20cm y diámetro ≥5cm) de cada variedad fueron lavadas y cortadas en tres rodajas de ~1cm de espesor cada una que provenían de la parte apical, central y distal de cada raíz. Estas 30 rodajas con peridermis de cada variedad fueron secadas en estufa a 70-80ºC hasta peso constante (48-60h). Posteriormente las muestras fueron molidas en molinillo de martillo hasta obtener una harina fina que se tamizó con malla de 1mm y se sometió a los análisis químicos de laboratorio para determinar contenido de proteínas, fibra bruta, N, P, K, Ca, Mg y Na.
En cada uno de los cinco muestreos las variables analizadas fueron: i) contenido de almidón, por el método de la gravedad específica (Toro y Cañas, 1983); ii) fibra bruta (Pearson, 1993), iii) proteínas (multiplicando el valor de N determinado por micro Kjeldahl por el factor de conversión 3,24 citado para mandioca (El-Sharkawy, 2012), y iv) contenido de P, K, Ca, Mg y Na siguiendo los protocolos descriptos en AOAC (2012). Se realizó análisis de varianza previa transformación de datos, test de comparaciones múltiples de Duncan (p≤0,05), análisis de correlación de Pearson y regresiones lineales, mediante el programa InfoStat v. 2008 (Di Rienzo et al., 2008). Todos los análisis se realizaron por triplicado y están referidos en % en base seca, con excepción del porcentaje de almidón.
Los datos de temperatura fueron registrados diariamente y por hora, con una mini estación meteorológica ‘Meteo’ (Cavadevices, Buenos Aires, Argentina), instalada en el invernadero durante el tiempo que duró el ensayo. Para determinar el tiempo térmico acumulado (TTA, ºC/día) en cada momento de cosecha (126, 160, 193, 223 y 255 días después de la plantación o días cronológicos) se empleó la ecuación general del método residual (Pascale y Damario, 2004), cuya fórmula es TTA= ∑ TT1 + TT2 + … + TTn; donde TT: temperatura media diaria – temperatura base; 1, 2 …n: días transcurridos entre plantación y momento de cosecha, y se expresó en ºC/día. Estudios realizados en Brasil, demostraron que la temperatura por debajo de la cual hay ausencia de crecimiento de plantas de mandioca es 16ºC (Ternes, 2002), por lo que se la consideró como la temperatura base (Tb) para el cálculo del tiempo térmico.
Resultados y Discusión
La distribución de fotoasimilados a las partes económicamente útiles de la planta de mandioca es medida por el índice de cosecha (IC); asimismo, el número de raíces tuberosas (RT) de mandioca y su peso medio son componentes del rendimiento que determinan la capacidad del destino o sumidero metabólico (Alves, 2002). De acuerdo a Hershey (1991) los máximos rendimientos se obtienen con plantas que alcanzan un valor de IC entre 0,5 y 0,65, siendo un carácter altamente heredable. Los resultados registrados en el presente trabajo demuestran el potencial que poseen estos genotipos, que sometidos a condiciones protegidas alcanzaron tempranamente valores de IC considerados óptimos; incluso se evidenció IC de 0,5 en dos de los clones a partir de los 160 días después de la plantación (ddp; Figura 1a). De todos modos, los IC variaron significativamente en función del tiempo y no respecto de los cultivares ni su interacción.
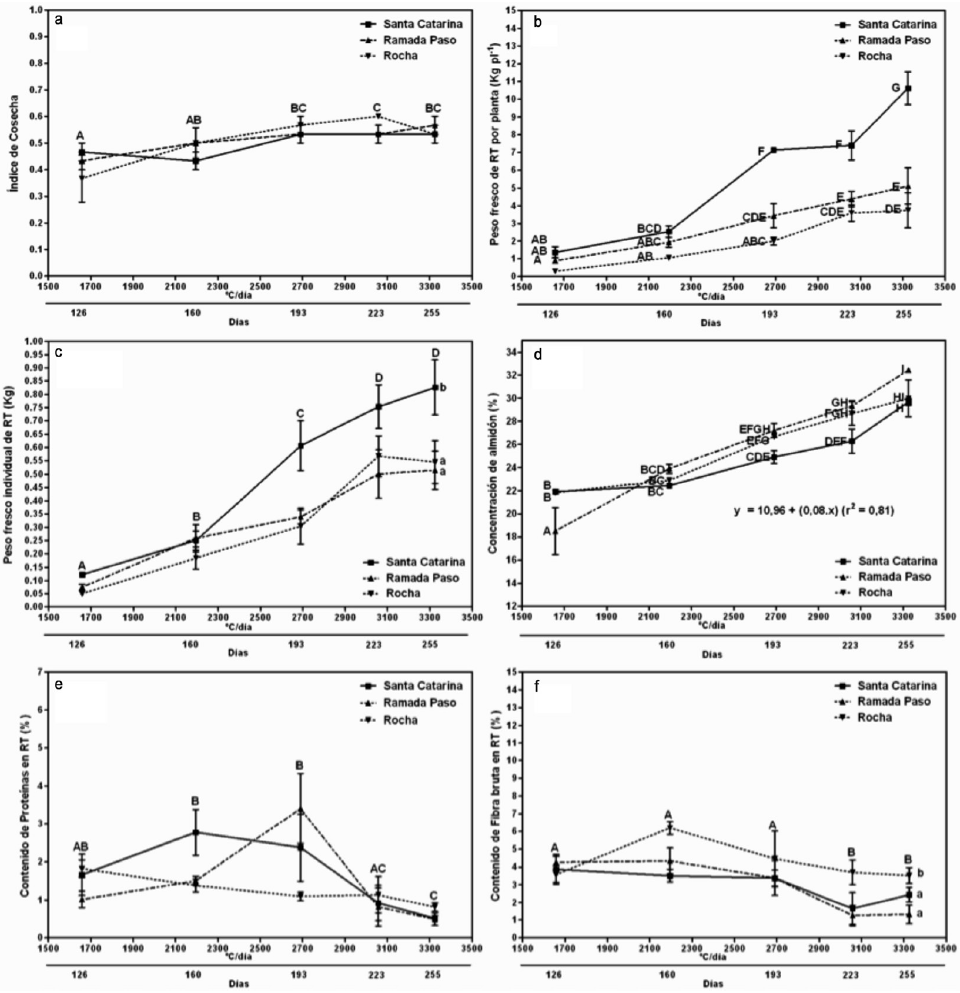
Figura 1
Evolución del índice de cosecha (a), el rendimiento de raíces tuberosas por planta (b), el peso fresco individual de raíces tuberosas (c), el contenido de almidón (d), proteínas (e) y fibra bruta (f) de raíces tuberosas de tres genotipos de mandioca cultivados bajo invernadero, en Corrientes, Argentina (2012/2013) en función del tiempo térmico acumulado expresado en ºC/día (TTA= S TT1+TT2+…+TTn; donde TT: temperatura media diaria - temperatura base; 1, 2 … n: días transcurridos entre plantación y momento de cosecha) y del tiempo en días. Letras mayúsculas distintas en a, c, e y f indican diferencias significativas con respecto al tiempo; letras minúsculas diferentes en c y f indican diferencias significativas con respecto al genotipo y letras mayúsculas distintas en b y d denotan diferencias estadísticas debidas a la interacción entre genotipo vs. tiempo según el test de comparaciones múltiples de Duncan (P£0,05).
En términos de rendimiento de RT de mandioca fresca en el invernadero (Figura 1b), los valores más altos alcanzados en este experimento, se observaron en el cv. Blanca de Santa Catarina, comprendidos entre 7 y 11kg/planta a los 193 y 255 ddp, respectivamente (2689,6ºC día y 3324,7ºC día), lo que representaría una producción estimada de 70 y 110Tn·ha-1 bajo las condiciones del invernadero. Estos valores son particularmente elevados respecto a los reportados para la región Nordeste argentino (Cenóz et al., 2001; Burgos et al., 2005, 2009) y a los rendimientos potenciales previstos para el año 2020, que de acuerdo a Howeler (2014) serán de 28,7Tn·ha-1 en India y 15,7Tn·ha-1 en América Latina y el Caribe. Los resultados son claramente alentadores, más aún cuando bajo condiciones experimentales en Colombia a partir de materiales selectos se registraron rendimientos de 90Tn·ha-1 de raíces frescas, considerándose este valor como el máximo potencial productivo reportado para la especie (Tofiño et al., 2008).
A los 255 ddp (3324,7ºC día), los cv. Ramada Paso y Rocha alcanzaron 4,8 y 3,1kg/planta, respectivamente, mostrando una interacción genotipo×tiempo significativa (Figura 1b). Sin embargo, a pesar de que el rendimiento fue significativamente menor en comparación con el cv. Blanca de Santa Catarina, éstos fueron superiores a los promedios citados para el continente americano, que no sobrepasan los 12,8Tn·ha-1 (Howeler, 2014).
Particularizando el estudio desde el análisis de los componentes del rendimiento, el Nº RT/planta varió significativamente entre cultivares, alcanzando valores de 6 RT/planta en el cv. Rocha y entre 10 y 11 RT/planta en los cv. Ramada Paso y Blanca de Santa Catarina, respectivamente (Tabla I). Con respecto al tiempo, si bien hubo variaciones las mismas no fueron significativas (F= 1,75; P=0,1661), ni en la interacción genotipo×tiempo (F=1,43; P=0,2261). Según Hershey (1991), la planta define el número de raíces durante el primer período de su desarrollo, posiblemente como respuesta a una mayor cantidad de fotoasimilados, producto de una actividad fotosintética más eficiente. En coincidencia, podría afirmarse que ante los resultados obtenidos (Tabla I), el Nº RT/planta ya estaba determinado antes del primer muestreo realizado (126 ddp) y por ello no se modificó a lo largo de los sucesivos muestreos. Da Silva et al. (2002) señalan que la determinación del mismo está más influenciado por el clon en si mismo, que por el ambiente; lo cual también coincide con los resultados hallados en este experimento.

* Días después de la plantación (ddp)** media aritmética ±error estándar*** letras mayúsculas distintas indican diferencias significativas según el test de comparaciones múltiples de Duncan (P£0,05)
El otro componente del rendimiento, el peso fresco individual de raíces tuberosas (Figura 1c), varió significativamente en función del tiempo y entre los cultivares. Por lo que estando en igualdad de número de raíces los cv. Blanca de Santa Catarina y Ramada Paso (Tabla I), el componente que determinó el rendimiento fue el peso individual de las mismas. Las diferencias en las ganancias de peso fresco individual de las raíces tuberosas del cv. Blanca de Santa Catarina con respecto a los demás cultivares, se hicieron evidentes a partir de los 193 ddp (2689,6ºC día). El cv. Ramada Paso, teniendo casi el doble de raíces tuberosas (Nº RT/planta) que el cv. Rocha, no se diferenció estadísticamente en términos de rendimiento (kg RT/planta), debido a la menor capacidad de ganancia de peso individual de las mismas (Figura 1c).
Las dos características del cultivo de mandioca que pueden influir sobre la calidad de la raíz son: el hecho de tratarse de un cultivo de ciclo largo (7 a 18 meses) y que no tiene un período definido de maduración para su cosecha; en consecuencia, es posible obtener variaciones de la calidad de la raíces tuberosas para consumo de acuerdo al momento de la cosecha (Hershey, 1991). No obstante, también influyen otros factores, tales como el genotipo y las condiciones de crecimiento (Odebunmi et al., 2007).
Entre los componentes químicos de mayor importancia presentes en las raíces de mandioca, se destaca el almidón, responsable de la mayoría de las propiedades funcionales que determinan su utilización en diferentes productos alimenticios (Techeira et al., 2014). En las condiciones de este ensayo, el contenido de almidón (Figura 1d) no difiere estadísticamente entre genotipos (29% en promedio), aumentando constante y significativamente en los tres cultivares con el tiempo (r2 = 0,81), a una tasa de acumulación diferencial dependiente del genotipo. Los cv. Blanca de Santa Catarina y Rocha fueron más precoces (Figura 1d), ya que desde los 4 meses después de la plantación ya alcanzaban contenidos de 22% de almidón, valor que normalmente se alcanzan a los 9-12 meses en condiciones convencionales de campo (Toro y Cañas, 1983). El cv. Ramada Paso, si bien a los 126 ddp presentó un porcentaje de almidón más bajo que los otros dos cultivares, finalizó el ciclo con los mayores porcentajes (Figura 1d).
En este sentido, según Burgos et al. (2013) en estudios realizados en el mismo sitio de experimentación, el tiempo térmico requerido para la plenitud de la fenofase de maduración de RT se requiere de 1500 a 1600ºC día, alcanzados a los 150-170 ddp. Para la plenitud de la fenofase de maduración de RT se requeire de 1500 a 1600ºC día, alcanzados al aire libre a los 160- 190 ddp. En el presente ensayo, en la primera cosecha de plantas de invernadero, en diciembre/2012 (126 ddp), la suma térmica fue de 1657,3ºC día (Tb= 16ºC), acumulando en abril/2013 (255 ddp) unos 3324,7ºC día. Esto explicaría porqué las raíces tuberosas cosechadas a los 126 ddp contenían una concentración de almidón entre 19 y 22% (Figura 1d), cercana a valores promedio registrados para mandioca cultivada por 270 ddp en condiciones de campo, alcanzando valores de 30-32% de almidón a los 255 ddp (Figura 1d), los cuales superan a los que se informan para cultivos bianuales en la región Nordeste de Argentina (De Bernardi, 2011).
Si bien el principal aporte nutricional de las raíces de mandioca son los carbohidratos, estos van acompañados de una fracción proteica que podría variar entre los cultivares, la edad a cosecha y las condiciones ambientales (Odebunmi et al., 2007). En este experimento se cuantificó su contenido en las harinas de diferentes cultivares y su evolución en el tiempo bajo condiciones protegidas. En la Figura 1e se observa que bajo invernadero el contenido de proteínas aumentó hasta los 193 ddp (2689,6ºC día) y posteriormente disminuyó significativamente. De acuerdo a Cabral et al. (2000), existe una estrecha relación entre la evolución ontogénica de las raíces tuberosas, el aumento de su diámetro y la reducción del contenido proteico, como consecuencia del aumento del número de las células del parénquima de reserva que acumulan una cantidad creciente de almidón; diluyendo el contenido de proteínas de los tejidos radicales. Asimismo, Gil y Buitrago (2002) determinaron que altos niveles de proteína se asocian a cosechas muy tempranas. En el experimento actual, el cv. Ramada Paso presentó a los 193 ddp los mayores porcentajes tanto de almidón (Figura 1d) como de proteínas (Figura 1e), alcanzando esta última valores próximos a los reportados por Gil y Buitrago (2002), Odebunmi et al. (2007), Rojas et al. (2007), Emmanuel et al. (2012) y Richardson (2013), quienes informaron contenidos entre 1,2 y 3,5%. Asimismo, Techeira et al. (2014) reportan porcentajes de proteína en harinas de mandioca del orden de 1,86% y Offor et al. (2012) de 0,96%, que en ambos casos se corresponden con los resultados obtenidos en este experimento al fin del ciclo, por lo que las condiciones del invernadero no perjudicarían su expresión. Ceballos et al. (2006), estudiando más de 140 clones durante 10 años hallaron variaciones extremas que van desde 0,95 a 6,42%, lo que denota la gran diversidad genética de la especie y la posibilidad de mejoramiento de los niveles proteicos mediante hibridaciones de la mandioca con parientes salvajes que cohabiten la misma región.
Con respecto al contenido de fibra bruta, la misma se mantuvo hasta los 193 ddp (2689,6ºC día), para luego disminuir significativamente (Figura 1f), lo que sugiere que estos valores también derivarían del efecto de dilución que provoca el aumento masivo del parénquima de reserva y su contenido de almidón durante el tiempo de realización del ensayo, coincidiendo con los valores reportados por Odebunmi et al. (2007) a partir de los 220 ddp. Según Gil y Buitrago (2002), Emmanuel et al. (2012) y Offor et al. (2012), el nivel de fibra es bastante estable, no supera 4% en las harinas de mandioca y, a diferencia del presente estudio, pocas diferencias se hallan entre cultivares. Como valores más extremos, Rojas et al. (2007) encontraron contenidos de fibra entre 7,4 y 8,5%; mientras Techeira et al. (2014) reportaron valores de 10,61%. En el presente experimento se presentaron evidentes diferencias entre variedades (Figura 1f), siendo máximo el porcentaje de fibra bruta en la harina del cv. Rocha a los 160 ddp (6,2%), para luego mantenerse en valores cercanos a 4%.
Si bien tanto los valores de proteínas y fibra bruta disminuyeron significativamente con el tiempo (Figuras 1e y 1f), no lo hicieron de modo lineal, registrándose correlaciones negativas significativas entre el tiempo y los contenidos de proteína (-0,49; P<0,0006) y fibra bruta (-0,48; P<0,0008). Además, se evidenció una asociación muy alta entre los contenidos de proteínas y fibra bruta (r= 0,98; P<0,0001), generando una recta de regresión con un r2 = 0,97 [y= 0,8+(0,29x)].<0,0006) y fibra bruta (-0,48; P<0,0008). Además, se evidenció una asociación muy alta entre los contenidos de proteínas y fibra bruta (r= 0,98; P<0,0001), generando una recta de regresión con un r2 = 0,97 [y= 0,8+(0,29x)].><0,0001), generando una recta de regresión con un r2 = 0,97 [y= 0,8+(0,29x)].
El contenido de almidón de las raíces frescas no tuvo una relación lineal con los contenidos de proteínas (r2 = 0,35) y fibra bruta (r2= 0,34) de las harinas.
En cuanto a la composición mineral de las harinas derivadas de raíces tuberosas obtenidas en condiciones de campo, en general la bibliografía informa amplios y diversos contenidos. Los resultados que se obtuvieron en este trabajo resultan originales en los términos de las condiciones de crecimiento en que se sometió al cultivo.
No se registraron diferencias significativas en los contenidos de P (0,04 ±0,0%), K (1,32 ±0,08%), Ca (0,32 ±0,02%), Mg (0,24 ±0,04%) y Na (0,02 ±0,0%), entre los cultivares, ni respecto al momento de cosecha. La caracterización de los valores promedio de cada uno de los contenidos de nutrientes minerales, expresados en base seca (%), se presentan en la Tabla II.
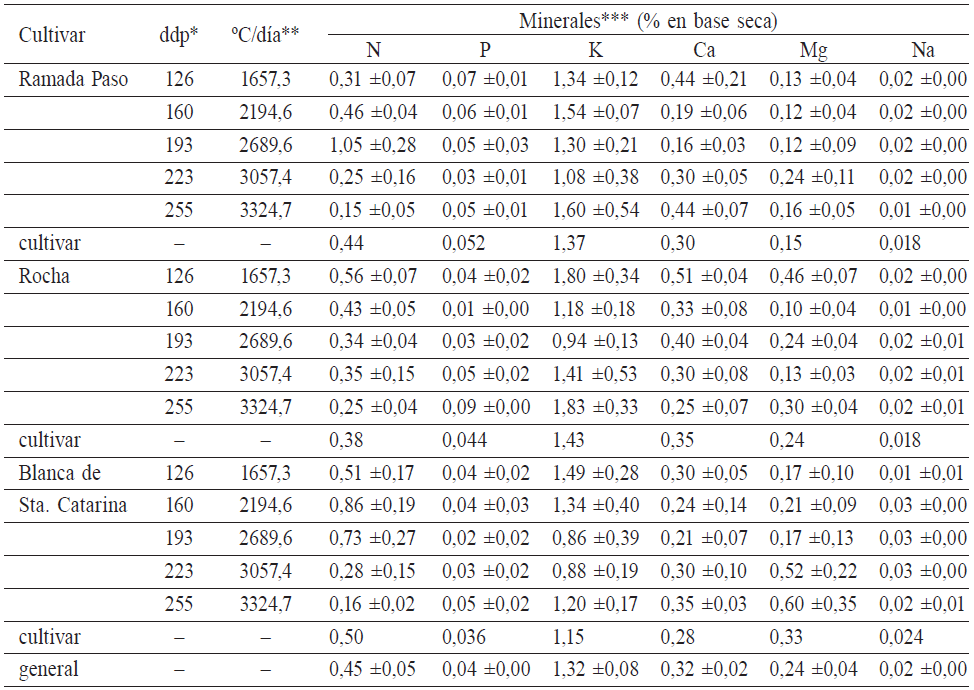
* Días después de la plantación (ddp)** tiempo térmico acumulado (ºC/día)*** media aritmética ±error estándar
En la Tabla II se pude observar que el P y Na fueron los minerales que se hallaron en menor porcentaje. El porcentaje de P resultó en muy baja cantidad respecto a lo reportado por Montaldo (1979) y Gil y Buitrago (2002). El porcentaje de Na se encontró dentro de los valores informados por Gil y Buitrago (2002) y Richardson (2013). Por otra parte, el porcentaje de Ca triplicó los contenidos publicados por Montaldo (1979) y fue semejante a los informados por Rojas et al. (2007). El porcentaje de K fue notoriamente elevado respecto a los resultados citados por Montaldo (1979) y Gil y Buitrago (2002) y similares a los publicados por Richardson (2013), siendo el mineral de mayor concentración en las harinas de mandioca (Howeler, 2014). El porcentaje de Mg alcanzó valores superiores a los reportados por Gil y Buitrago (2002), que contrastan con los de Montaldo (1979) y que son próximos a los hallados en este experimento, resultando muy favorable.
Corrientes es una de las provincias con mayor concentración de invernaderos en Argentina y por lo tanto con gran tradición hortícola en cultivos protegidos, caracterizándose por la producción temprana de hortalizas para consumo fresco. De acuerdo a los resultados obtenidos en el presente estudio, es posible cultivar mandioca en condiciones de invernadero logrando altos rendimientos y cosechas tempranas, con raíces tuberosas aptas tanto para el consumo en fresco como para la provisión de materia prima en la industria alimenticia.
En relación al contenido de almidón, se manifiesta un comportamiento genotípico diferencial, donde se presentan unas variedades más tardías y otras más precoces. Si bien ya se ha enfatizado que el valor nutricional de las raíces de mandioca depende de múltiples factores, entre ellos, el genotipo y la edad de la planta a la cual es cosechada (Odebunmi et al., 2007), este es el primer trabajo en donde se informan los contenidos de nutrientes de raíces tuberosas de mandioca derivadas de plantas cultivadas en condiciones de invernadero, donde particularmente se vislumbraría un efecto acelerador de la suma térmica sobre la determinación del rendimiento, la síntesis y almacenamiento de almidón.
Las diferencias halladas permiten caracterizar las variaciones entre materiales genéticos, constituyendo la base para el desarrollo de planes de mejoramiento genético. Más aún, la importancia de las condiciones ambientales locales en las que se desarrolló el experimento deberán también ser tomadas en cuenta, dado que el componente ambiental modificado por el invernadero plástico interacciona con la base genética de la planta para determinar la calidad de la raíz en lo que se refiere a determinados componentes químicos, tanto como en diversos componentes morfofisiológicos y del rendimiento a los que la mandioca resulta altamente sensible (Ntawuruhunga y Dixon, 2010).
Conclusiones
En este trabajo se demuestra que es factible realizar el cultivo de mandioca en condiciones de invernadero, alcanzando rendimientos muy elevados y cosechas tempranas, con raíces tuberosas con alta concentración de almidón, y contenidos de proteína y fibra aceptables. La composición mineral no presenta un patrón de comportamiento que responda al efecto del genotipo ni al tiempo, aunque los contenidos elevados de K y Mg registrados en las raíces tuberosas derivadas de invernadero constituyen un valor nutricional potencial a ser considerado.
Agradecimientos
Los autores agradecen a la Agencia Nacional de Promoción Científica y Tecnológica (PICT 2309-2012), a la Secretaría General de Ciencia y Técnica, UNNE (PI A006/10, PI A002/11 y PI A00/14) y al Consejo Nacional de Investigaciones Científicas y Técnicas por financiar este trabajo.
REFERENCIAS
Alves A (2002) Cassava Botany and Physiology. En Hillocks R, Thresh J, Belloti A (Eds.) Cassava: Biology, Production and Utilization. CABI. Nueva York, EEUU. pp. 67-89.
AOAC (2012) Official Methods of Analysis. 19th ed. Association of Official Analytical Chemists. Gaithersburg, MD, EEUU. 2610 pp.
Bongiovanni R, Morandi J, Troilo L (2012) Competitividad y Calidad de los Cultivos Industriales: Caña de Azúcar, Mandioca, Maní, Tabaco, Té y Yerba Mate. INTA. Córdoba, Argentina. 108 pp.
Burgos AM, Cenóz PJ, López A, Rodríguez SC (2005) Efecto de podas y del sistema de almacenamiento sobre factores de calidad de raíces de mandioca (Manihot esculenta Crantz). Agrotecnia 15: 17-21.
Burgos AM, Cenóz PJ, Prause J (2009) Efecto de la aplicación de auxinas sobre el proceso de enraizamiento de estacas de dos cultivares de mandioca (Manihot esculenta Crantz). UDO Agríc. 9: 539-546.
Burgos A, Prause J, Argüello J, Cenóz P (2013) Fenología de los estados vegetativos de mandioca (Manihot esculenta Crantz) en términos de tiempo térmico. Rev. Fac. Cs. Agr. UNCuyo 45: 43-52.
Burns A, Gleadow R, Zacarias A, Cuambe C, Miller R, Cavagnaro T (2012) Variations in the chemical composition of cassava (Manihot esculenta Crantz) leaves and roots as affected by genotypic and environmental variation. J. Agric. Food Chem. 60: 4946-4956.
Cabral G, Carvalho L, Schaal B (2000) The formation of storage root in cassava. En Carvalho L, Thro A, Duarte Vilarinhos A (Eds.) Proc. 4th Int. Scientific Meeting of Cassava Biotechnology Network. Embrapa. Brasilia, Brazil. pp. 345-356.
Ceballos H, Sánchez T, Chávez A, Iglesias C, Debouck D, Mafla G, Tohme J (2006) Variation in crude protein content in cassava (Manihot esculenta Crantz) roots. J. Food Comp. Anal. 19: 589-593.
Cenóz PJ, López A, Burgos AM (2001) Efecto de los macronutrientes en el desarrollo y rendimiento de mandioca (Manihot esculenta Crantz). Agrotecnia 7: 9-12.
Da Silva R, Fernández Faraldo M, Ando A, Veasey E (2002) Variabilidade genética de etnovariedades de mandioca. En Cereda M (Ed.) Culturas de Tuberosas Amiláceas Latino Americanas 2. Fundação Cargill. San Pablo, Brazil. pp. 207-241.
De Bernardi L (2011) Mandioca. Alim. Argent. 51: 49-52.
Di Rienzo J, Casanoves F, Balzarini M, Gonzalez L, Tablada M, Robledo C (2008) InfoStat v. 2008. Grupo InfoStat, Universidad Nacional de Córdoba. Córdoba, Argentina. 336 pp.
El-Sharkawy MA (2012) Stress-tolerant cassava: the role of integrative ecophysiology-breeding research in crop improvement. Open J. Soil Sci. 2: 162-186.
Emmanuel O, Clement A, Agnes S, Chiwona-Karltun L, Drinah B (2012) Chemical composition and cyanogenic potential of traditional and high yielding CMD resistant cassava (Manihot esculenta Crantz) varieties. Int. Food Res. J. 19: 175-181.
Gil J, Buitrago J (2002) La yuca en la alimentación animal. En Ospina B, Ceballos H (Eds.) La Yuca en el Tercer Milenio. Sistemas Modernos de Producción, Procesamiento, Utilización y Comercialización. Centro Internacional de Agricultura Tropical. Cali, Colombia. pp. 527-569.
Hershey C (1991) Mejoramiento Genético de la Yuca en América Latina. Centro Internacional de Agricultura Tropical. Cali, Colombia. 426 pp.
Howeler R (2014) Sustainable Soil and Crop Management of Cassava in Asia. Centro Internacional de Agricultura Tropical. Cali, Colombia. 280 pp.
Montaldo A (1979) La Yuca o Mandioca. Instituto Interamericano de Ciencias Agrícolas. San José, Costa Rica. 386 pp.
Murphy GM (Ed.) (2008) Atlas Agroclimático de la Argentina. Facultad de Agronomía, Universidad de Buenos Aires. Argentina. 130 pp.
Ntawuruhunga P, Dixon A (2010) Quantitative variation and interrelationship between factors influencing cassava yield. J. Appl. Biosci. 26: 1594-1602.
Offor C, Igwenyi I, Afiukwa C, Uruaku A., Aloh G, Aja P, Onwe F (2012) The comparative proximate compositions of staple food crops in Ezzangbo (Ohaukwu L.G.A.) and Iboko (Izzi L.G.A) of Ebonyi State, Nigeria. Cont. J. Pharmacol. Toxicol. Res. 5: 23-27.
Odebunmi E, Oluwaniyi O, Sanda A, Kolade B (2007) Nutritional compositions of selected tubers and root crops used in Nigerian food preparations. Int. J. Chem. 17: 37-43.
Pascale AJ, Damario EA (2004) Bioclimatología Agrícola y Agroclimatología. Facultad de Agronomía, Universidad de Buenos Aires. Argentina. 550 pp.
Pearson D (1993) Técnicas de Laboratorio para Análisis de Alimentos. Acribia. Zaragoza, España. 331 pp.
Richardson K (2013) Quality Characteristics, Root Yield and Nutrient Composition of Six Cassava ( Manihot esculenta Crantz) Varieties. GRAC Crop Research Report Nº 18, Department of Agriculture. Nasáu, Bahamas. 13 pp.
Rojas C, Nair B, Herbas A, Bergentahl B (2007) Proximal composition and mineral contents of six varieties of cassava (Manihot esculenta Crantz) from Bolivia. Rev. Bol. Quím. 24: 71-76.
Soil Survey Staff (1990) Kays to Soil Taxonomy. SMSS Technical Monograph Nº 6, 4a ed. Virginia, EE.UU. 422 pp.
Techeira N, Sívoli L, Perdomo B, Ramirez A, Sosa F (2014) Caracterización fisicoquímica, funcional y nutricional de harinas crudas obtenidas a partir de diferentes variedades de yuca (Manihot esculenta Crantz.), batata (Ipomoea batatas Lam.) y ñame (Dioscorea alata), cultivadas en Venezuela. Interciencia 39: 191-197.
Ternes M (2002) Fisiología da planta. En Cereda MP (Ed.) Agricultura: Tuberosas Amiláceas Latino Americanas. Fundación Cargill. San Pablo, Brazil. pp. 66-82.
Tofiño A, Ceballos H, Romero H (2008) Posibilidades de expansión del cultivo de yuca (Manihot esculenta Crantz) en el Caribe seco colombiano a partir de investigación multidisciplinaria. Actual. Biol. 30: 15-27.
Toro J, Cañas A (1983) Determinación del contenido de materia seca y almidón en yuca por el sistema de gravedad específica. En Domínguez C (Ed.) Yuca: Investigación, Producción y Utilización. PNUD/CIAT. Cali, Colombia. pp. 567-575.
Notas de autor