Comunicaciones
TAXONOMIC VALUE OF THE METAFURCASTERNUM IN GEOMETRIDAE: A MORPHOLOGICAL COMPARISON APPROACH
VALOR TAXONÓMICO DEL METAFURCASTERNO EN GEOMETRIDAE: UN ENFOQUE MORFOLÓGICO COMPARATIVO
VALOR TAXONÔMICO DA META-FÚRCULA-ESTERNAL EM GEOMETRIDAE: UM ESTUDO MORFOLÓGICO COMPARATIVO
TAXONOMIC VALUE OF THE METAFURCASTERNUM IN GEOMETRIDAE: A MORPHOLOGICAL COMPARISON APPROACH
Interciencia, vol. 43, no. 3, pp. 202-207, 2018
Asociación Interciencia
Received: 26 June 2017
Accepted: 26 February 2018
Funding
Funding source: Dirección de Investigación, Universidad de Concepción
Contract number: VRID 214.113.087-1.0
Funding
Funding source: Universidad de Tarapacá
Contract number: UTA Mayor (9719-17)
Funding statement: This work was supported by grant Nº VRID 214.113.087-1.0 from the Dirección de Investigación, Universidad de Concepción and UTA Mayor (9719-17) from the Universidad de Tarapacá.
Abstract: In the more developed Pterygotas, such as Lepidoptera, the sternal apophyses are much closer and they tend to be held by a medium inflection of the sternum (the furca). In this way, the coxas (cx) become more ventral than in the original condition. Using the metafurcasternum as a morphological character is restricted as a phylogenetic relationship classification at family level. In the functional aspect, the furca is an indicator of flight capacity of Noctuid species. However, comparative morphology of the metafurcasternum between geometrid species has not been commonly used at a specific level in the Lepidoptera group. This report makes a comparison taking into account the taxonomic relationship of the metafurcasternun as a complex trait. The study was performed in ten species of Enomines of the Geometridae family when the morpho-functional variability of the different geometrid species could be related to the flight aerodynamic of the group, using this trait to help in future taxonomic classification. In the morphological field and regarding the taxonomic value of the metafurcasternum it can be said that the major differences between species, as to structure, are those in the relative shape of the anterior ventral plate, the other structures of the metafurcasternum remaining almost the same, with differences mainly in size and proportion.
Keywords: Ennominae, Geometridae, Metafurcasternum, Morphology, Wings.
Resumen: En los Pterigotos más desarrollados, como los lepidópteros, las apófisis esternales están mucho más cerca y tienden a estar sostenidas por una inflexión media del esternón (la furca). De esta manera, las coxas (cx) se vuelven más ventrales que en la condición original. El uso del metafurcasternum como carácter morfológico está restringido a la clasificación de relaciones filogenéticas a nivel de familias. En el aspecto funcional, la furca es un indicador de la capacidad de vuelo de las especies de Noctuidae. Sin embargo, la morfología comparada del metafurcasternum en geométridos no ha sido usada a nivel específico. Este estudio compara el metafurcaesternum en diferentes especies teniendo en cuenta la relación taxonómica de éste como un rasgo complejo. El estudio se realizó en diez especies de ennomines de la familia Geometridae, la variabilidad morfofuncional del metafurcaesterno se relaciona con la aerodinámica del vuelo del grupo, y se evalúa este rasgo para su uso en la clasificación taxonómica. En el campo morfológico y en relación con el valor taxonómico del metafurcasternum se encontró que las principales diferencias entre las especies están en la forma relativa de la placa ventral anterior. Las otras estructuras del metafurcasternum son muy similares, diferenciándose principalmente en tamaño y proporción.
Resumo: Em Pterigotos mais desenvolvidos, como os lepidópteros, as apófises esternais estão muito mais perto e tendem a estar sustentadas por uma inflexão média do esterno (fúrcula). De esta forma, as coxas (cx) ficam mais ventrais que na condição original. O uso da meta-fúrcula-esternal como carácter morfológico está restrito à classificação de relações filogenéticas no nível de famílias. No aspecto funcional, a fúrcula é um indicador da capacidade de voo das espécies de Noctuidae. No entanto, a morfologia comparada da meta-fúrcula-esternal nos geometridae não tem sido usada no nível específico. Este estudo compara a meta-fúrcula-esternal em diferentes espécies levando em consideração a relação taxonômica deste como uma característica complexa. O estudo foi realizado em dez espécies de ennominae da família Geometridae, a variabilidade morfofuncional da meta-fúrcula-esternal se relaciona com a aerodinâmica de voo do grupo, e se avalia esta característica para seu uso na classificação taxonômica. No campo morfológico e em relação com o valor taxonômico da meta-fúrcula-esternal se observou que as principais diferenças entre as espécies estão na forma relativa da lâmina ventral anterior. As outras estruturas da meta-fúrcula-esternal são muito similares, diferenciando-se principalmente em tamanho e proporção.
Introduction
Insects have several adaptation characters that have lead them to evolutionary success. Many of these characters are linked to plasticity or morphological adaptation, where differences may have high behavioural repertoires or they may vary in performance (that is, the degree in which individuals fulfil the corresponding ecological functions) of specific behaviours (Stillwell and Davidowitz, 2010). In case of a discrete morphological variation, such differences may be obvious; for example, there are several insects that have non-winged forms (braquypterous) and winged forms (macropterous), and only the latter can fly (Zera and Denno, 1997; Berwaerts et al., 2002; Stillwell and Davidowitz, 2010).
For traits with continuous variation, relationship between shape and performance may be more subtle and require careful testing. Nevertheless, not only the performance variation can influence the variations in form, development, behaviour, since other history life traits also cause variation in fitness (Arnold, 1983; Berwaerts et al., 2002; Bonte and Van Dyck, 2009).
Undoubtedly, development of wings brought along countless adaptation advantages in butterflies, such as dispersion, migration, defence and sexual selection (e.g. courtship flight, territoriality and anti-predation) (Ellington, 1984; Benson et al., 1989; Srygley, 1994; Dudley, 2000; Breuker et al., 2007; Johansson et al., 2009), where development, shape and ratio of endoskeleton structures are fundamental to support wing muscles (endosternite) (Angulo et al., 1986).
The sternum in winged insects is characterized by the presence of a pair of apophyses rising from the sternal plates (Snodgrass, 1935; Kristensen, 1984). The sternal apophyses (as), called ‘furca arms’ in the superior Pterygota are held by a medium inflection of the sternum called furca (fu). The external margin of the sternal apophyses is attached to the internal margin of ‘pleural arms’ (blp) of the same segment through short muscle fibres or, in some cases, they become fused, forming an arc across the coxal cavity from the sternum to the pleura.
In the more primitive Pterygotas, the base of the sternal apophysis would connect through an internal transverse fold called costa-sternum (sc). In the more developed Pterygotas, such as Lepidoptera, the sternal apophyses are much closer and they tend to be held by a medium inflection of the sternum (the furca). In this way, the coxas (cx) become more ventral than in the original condition (Scoble, 1995; Dudley, 2000).
Using the metafurcasternum as a morphological character is restricted to the work of Brock (1971), only at family level, to establish phylogenetic relationships (Kitching, 1984). In the functional aspect, Angulo et al. (1986) state that the furca is an indicator of flight capacity of Noctuidae species. Howe ver, the study of these morphological structures has not been commonly used at a specific level in the Lepidoptera group.
This report analyzes and compares the morphological variation between ten geometrid species metafurcasternum as an intraspecific functional taxonomic trait.
Materials and Methods
The biologic material used consisted of ten species of Enomines of the Geometridae family: Leucolithodes paulina (Ureta); Catophoenissa dibapha Felder & Rogenhofer; Catophoenissa fuenzalidai Ureta; Mallomus antennata (Mabille); Mallomus clarkei (Sperry); Mallomus chilenaria (Felder & Rogenhofer); Mallomus penai (Rindge); Syncirsodes primata (Walker); Praentarctia indecisa Heimlich and Tetracis chilenaria Blanchard.
Specimens were treated with KOH at 10% w/v, after being dampened in ethyl alcohol at 70% w/v in order to make structures clearer and denude the specimen.
The thorax was prepared in faucet water inside a Petri capsule; then the metathorax was separated from the rest of the thorax to examine and clean it from the posterior area, so that the furcasternum could be visualized (Angulo et al. 1986).
For this study, the metafurcasternum morphology was considered, which is an actual support for wing muscles of the second wing pair, depending on the degree of development. The index proposed by Angulo et al. (1986) was used for better interpretation of the status within the character series of the metasternal furca (metafurcasternum). This index includes measures, shown in Table I, which are considered in two relations, R1 and R2, as follows:
R1= LBS/LEPF
and
R2= LLPV/LEPF,
where LEPF: length of the postfurcal sclerite or furca stem, a constant structure within the different types of species studied; LBS: length of secondary furca arm; and LLPV: length of the thoracic-pleural line. The relations R1 and R2 reflect the measure of development of the metasternal furca. Significant differences between morphological variables in the ten species were analyzed by means of a non-parametric t test.
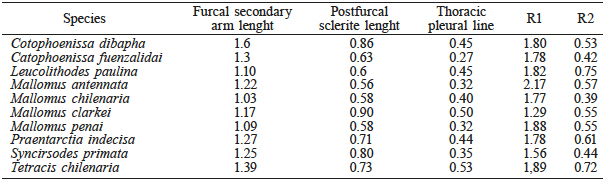
Values are in milimeters (mm).
Results
The morphological variables analysed and their relations R1 and R2 (Table I) showed significant differences in the ten species of Geometridae confirming the high taxonomic variability of the metafurcasternum (t= 22.462; p<0.0001). The morpho-taxonomic description is as follows:
Leucolithodes paulina (Ureta)
In the anterior view (Figure 1) the secondary arms of the furca (bsf) are in an elongated triangular shape, the external lateral edge of these has straight sides forming an angle at more than 90º; the furca body (cf) has the fork in a ‘v’ shape, with the apophyses (af) in 1/3 of the size of it and the apex of it is crowned by the apophysis membrane (ma) forming a saddle. In the side view (Figure 1) the forked condition of the furca apophysis is observed; the furca body in its ventral region and on the base of the apophysis ends acutely, and the ligulated anterior ventral plate (lva) hangs from here, running backwards to the subterminal region of the furca stem (tf) where it becomes curved.
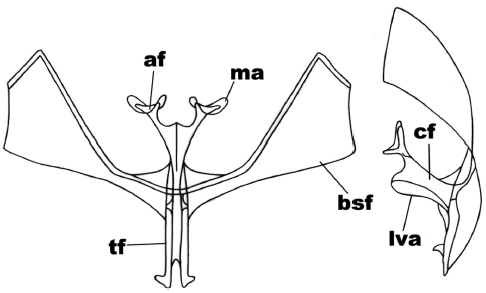
Figure 1
Anterior and side view of the metafurcasternum in Leucolithodes paulina (Ureta)
The abbreviated traits are: secondary arms of the furca (bsf), furca body (cf), apophyses (af), apophysis membrane (ma), ligulated anterior ventral plate (lva), furca stem (tf).
Cotophoenissa dibapha (Felder & Rogenhofer)
In the anterior view (Figure 2), the secondary arms are in a subtriangular shape with three of their sides almost identical; the furca body is forked in a ‘v’ shape, from where both apophyses emerge in subreniform shape. In the side view (Figure 2) the furca apophyses show their forked condition running towards the dorsal region; the furca body on the base of apophyses is wider than its medial region; the anterior ventral plate has its anterior edge twice wider than its posterior one.
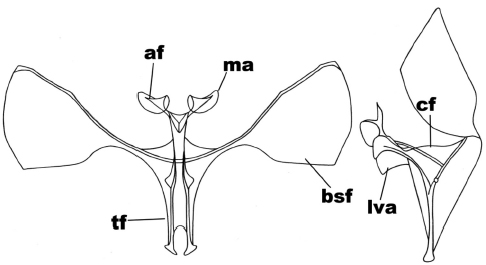
Figure 2
Anterior and side view of the metafurcasternum in Cotophoenissa dibapha (Felder & Rogenhofer)
The abbreviated traits are: secondary arms of the furca (bsf), furca body (cf), apophyses (af), apophysis membrane (ma), ligulated anterior ventral plate (lva), furca stem (tf).
Catophoenissa fuenzalidai Ureta
In the anterior view (Figure 3), the secondary furca arms are substraight, in subtriangular shape; the furca body is forked in a ‘v’ shape with the upper edge three times wider than its base; the furca apophyses are in triangular shape, surrounded by the subreniform apophysis membrane. In the side view (Figure 3) the forked apophyses go to the dorsal region, the furca body has parallel sides in all its length; the anterior ventral plate is triangular (all its sides in the same length) and it does not reach the furca stem.
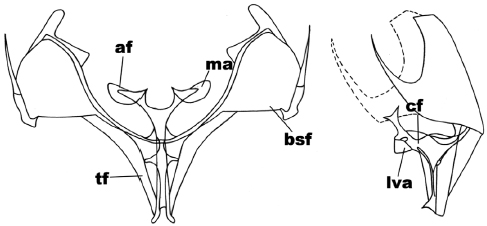
Figure 3
Anterior and side view of the metafurcasternum in Catophoenissa fuenzalidai Ureta
The abbreviated traits are: secondary arms of the furca (bsf), furca body (cf), apophyses (af), apophysis membrane (ma), ligulated anterior ventral plate (lva), furca stem (tf).
Mallomus antennata (Mabille)
In the anteror view (Figure 4), the secondary furca arms are subtriangular with straight sides and forming an angle at 90º in the ventral-lateral region, the dorsal margin has a low cut in the distal third of the arm; the furca body is forked in a ‘v’ shape, the apophyses are elongated and crowned by the membrane of the subreniform apophysis. In the side view (Figure 4), the furca body has parallel sides all along its length, in its anterior end and beneath the apophysis base it is slightly invaginated towards the anterior ventral plate; uniform apophysis; the anterior ventral plate is subequal in length to the furca body (plus the apophyses); this is subrectangular with round anterior edge strongly expanded.
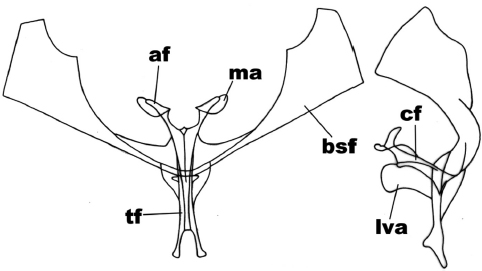
Figure 4
Anterior and side view of the metafurcasternum in Mallomus antennata (Mabille)
The abbreviated traits are: secondary arms of the furca (bsf), furca body (cf), apophyses (af), apophysis membrane (ma), ligulated anterior ventral plate (lva), furca stem (tf).
Mallomus chilenaria (Felder & Rogenhofer)
In the anterior view (Figure 5), the secondary furca arms are subtriangular (sides are similar) going towards the dorsal region, rounded external side edge; the furca body is forked in a ‘v’ shape, subequal in length to the furca stem; the furca apophyses are subtriangular in shape, with the apex crowned by the membrane of the subreniform apophysis. In the side view (Figure 5), the furca apophysis is simple; the furca body is subrectangular and under this there is a membrane in triangular shape curved towards the region where it becomes attached to the furca stem.
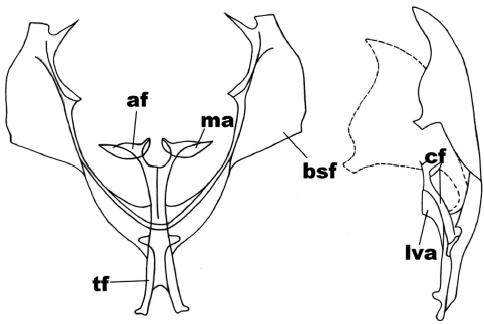
Figure 5
Anterior and side view of the metafurcasternum in Mallomus chilenaria (Felder & Rogenhofer)
The abbreviated traits are: secondary arms of the furca (bsf), furca body (cf), apophyses (af), apophysis membrane (ma), ligulated anterior ventral plate (lva), furca stem (tf).
Mallomus clarkei (Sperry)
In the anterior view (Figure 6), the secondary furca arms are triangular, the lateral and ventral margin of these are rounded; the furca body is forked in a ‘v’ shape; the furca apophyses are subtriangular, and tye apophysis membrane is subcircular. In the side view (Figure 6), the furca apophysis is forked but the anterior branch is four times longer than the posterior one; the furca body is subrectangular; the anterior ventral plate is subrectangular, with anterior margin 33% larger than posterior margin.
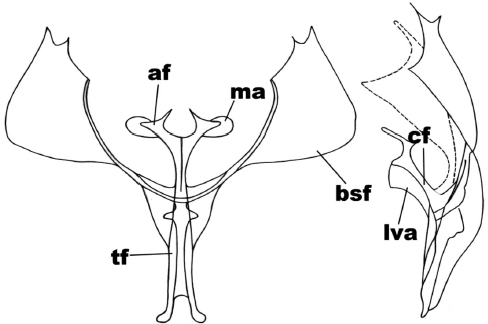
Figure 6
Anterior and side view of the metafurcasternum in Mallomus clarkei (Sperry)
The abbreviated traits are: secondary arms of the furca (bsf), furca body (cf), apophyses (af), apophysis membrane (ma), ligulated anterior ventral plate (lva), furca stem (tf).
Mallomus penai (Rindge)
In the anterior view (Figure 7), the secondary furca arms are triangular; the furca body is forked in a ‘v’ shape; the furca apophysis is subtriangular, the apophysis membrane has triangular shape. In the side view (Figure 7), the furca apophysis is triangular; the furca body is subrectangular and curved; the anterior ventral plate is unciform (thickened).
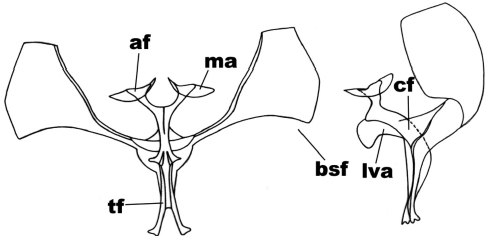
Figure 7
Anterior and side view of the metafurcasternum in Mallomus penai (Rindge)
The abbreviated traits are: secondary arms of the furca (bsf), furca body (cf), apophyses (af), apophysis membrane (ma), ligulated anterior ventral plate (lva), furca stem (tf)
Syncirsodes primata (Walker)
In the anterior view (Figure 8), the secondary furca arms are subcircular; the furca body is forked in a ‘v’ shape; the furca apophyses are small and subtriangular, in their apex they have the apophysis membrane in subcircular shape. In the side view (Figure 8), the apophysis is simple and curved forward, forming a right angle; the furca body is subrectangular; the anterior ventral plate is triangular (isosceles triangle).
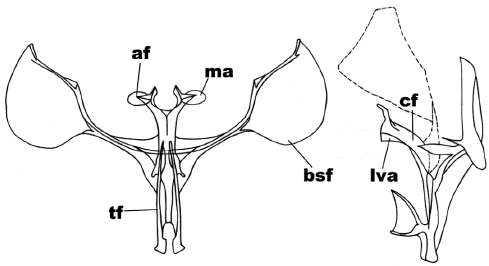
Figure 8
Anterior and side view of the metafurcasternum in Syncirsodes primata (Walker)
The abbreviated traits are: secondary arms of the furca (bsf), furca body (cf), apophyses (af), apophysis membrane (ma), ligulated anterior ventral plate (lva), furca stem (tf).
Praentarctia indecisa Heimlich
In the anterior view (Figure 9), the secondary furca arms are subtriangular, their ventral and closest lateral margins form a right angle; the furca body is forked in a ‘v’ shape; the apophyses are triangular in shape, in their apex they have the apophysis membrane in a saddle shape. In the side view (Figure 9), the furca body shows its forked condition; the apophyses are slightly separated, the anterior ventral plate is bilobed, with the anterior lobe longer and narrower than the posterior one.
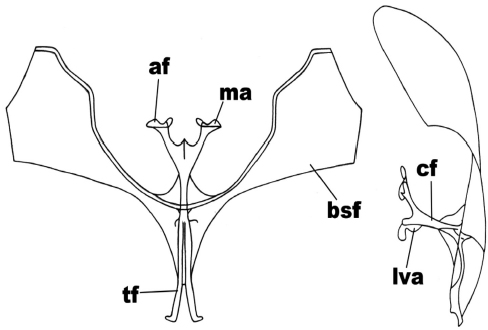
Figure 9
Anterior and side view of the metafurcasternum in Praeantarctia indecisa Heimlich
The abbreviated traits are: secondary arms of the furca (bsf), furca body (cf), apophyses (af), apophysis membrane (ma), ligulated anterior ventral plate (lva), furca stem (tf).
Tetracis chilenaria (Blanchard)
In the anterior view (Figure 10), the secondary furca arms are subtriangular; the furca body is forked in a ‘v’ shape, the apophysis is caligiform, with apophysis membrane in triangle shape and curved towards the medial region of the furca. In the side view (Figure 10), the furca body shows its forked condition, the apophysis is simple and digitiform; the anterior ventral plate is unciform.
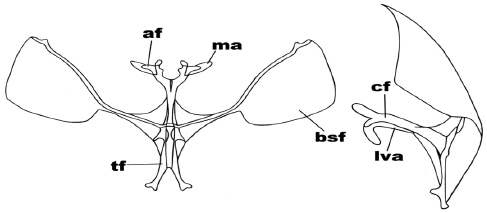
Figure 10
Anterior and side view of the metafurcasternum in Tetracis chilenaria (Blanchard)
The abbreviated traits are: secondary arms of the furca (bsf), furca body (cf), apophyses (af), apophysis membrane (ma), ligulated anterior ventral plate (lva), furca stem (tf).
Discussion
The traditional morphometrics results are an apprpriate tool to analyse the morphological variation of the metafurcasternun, revealing clear differences between species and corroborating the taxonomic importance of the metafurcasternun as a trait to differentiate species in the Geometridae family.
In Lepidoptera anatomy, has been stated that the metafurcasternum is a trait which is directly related to the flight capacity (Angulo et al., 1986; Scoble, 1995; Dudley, 2000). Therefore, the related measurements at the studied indexes were used to indicate the flight capacity previous between the different species. results show that the species with highest flight capacity are Leucolithodes paulina and Tetracis chilenaria; the species with lowest flight capacity are Praeentarctia indecisa, Catophoenissa dibapha and Mallomus penai; the species with lower flight capacity are Catophoenissa fuenzalidai and Mallomus chilenaria; the following are bad at flying: Mallomus clarkei and Syncirsodes primata; Mallomus antennata is bad at flying but good at gliding (Parra and Henriquez-Rodriguez, 1993).
In the morphological field and regarding the taxonomic value of the metafurcasternum it can be stated that the major differences between species, as to structure, are those in the relative shape of the anterior ventral plate. The other structures of the metafurcasternum are almost the same, with differences mainly in size and proportion.
From the species with better flight capacity, L. paulina is the one with a more aerodynamic body. T. chilenaria is another species that is good at flying in spite of its wide wings. Its higher flight capacity is due to an increase (compared to the other species under study) in the length of the furca body (thoracic-pleural line) (Figure 10). M. clarkei and Syncirsodes primata are bad at flying and they have in general a less aerodynamic appearance: very expanded wings and rather thick body (the latter is more evident in M. clarkei)
Based on its relationships R1 and R2 (Table I), M. antennata is the ‘best gliding’ species, since proportionally it has, among all the species, increased length of the secondary furcal arm. Therefore, the wings seem to be more separated from one another, preventing good wing flapping; conversely, this larger separation of the wings provides a better lift in the aerial environment. The latter observation is proven by the wind intensity existing in the south of the Chilean distribution range. With strong winds it would let itself be swept and glide in the aerial environment, flapping would be minimum.
The other species are in an intermediate position, their condition of being better or worse at flying is determined by the R1 and R2 ratios, and these by the length of the different structures considered (that is, LBS, LEPF, LLPV).
The scarce variability of morphological characters observed in the metafurcasternum is possibly due to the body nature of Geometridae in contrast to other families, such as Noctuidae, which need a good support base for wing muscles, and these must withstand a higher number of contractions to lift a thicker and heavier body (Heimlich, 1987). In general, the fact that Geometridae have large wings and lower body weight allows them to stay in the air with a low frequency of wing flaps (Casey and Joos 1983). In this sense, the two species that are best at flying would have a low performance in these two patterns: L. paulina, with a thicker body, needs more flapping to fly and hence higher energy for this task; T. chilenaria would be closer to behave as a Geometridae in flying, it has large wings and a rather small body (less weight), therefore it needs less flapping and lower energy for this purpose.
The report describes for the first time multiple comparison of a single functional trait using traditional morphometrics. The metafurcasternum in Geometridae is a trait that has an exclusive relationship with flight aerodynamics in Lepidoptera, helping to improve the taxonomic information of description of complex trait for classification of species, combining functional classifications and opening the door for further research using functional traits for taxonomic differentiation.
Acknowledgements
The authors thank Maria Raquel Lazo de la Vega for help in the English of the manuscript. This work was supported by grant Nº VRID 214.113.087-1.0 from the Dirección de Investigación, Universidad de Concepción and UTA Mayor (9719-17) from the Universidad de Tarapacá.
References
Angulo AO, Jana-Saenz C, Parra LE (1986) Estudio del metafurcaesterno en algunas especies de noctuidos; valor morfofuncional (Lepidoptera: Noctuidae). Ann. Soc. Entomol. Fr. 22: 369-374.
Arnold SJ (1983) Morphology, performance and fitness. Am. Zool. 23: 347-361.
Benson WW, Haddad CF, Zikán M (1989) Territorial behavior and dominance in some heliconiine butterflies (Nymphalidae). J. Lepid. Soc. 43: 33-49.
Berwaerts K, Van Dyck H, Aerts P (2002) Does flight morphology relate to flight performance? An experimental test with the butterfly Pararge aegeria. Funct. Ecol. 16: 484-491.
Breuker C, Brakefield P, Gibbs M (2007) The associations between wing morphology and dispersal is sex-limited in the Glanville fritillary butterfly Melitaea conxia (Lepidoptera: Nymphalidae). Eur. J. Entomol. 104: 445-452.
Bonte D, Van Dyck H (2009) Mate-locating behaviour, habitat-use, and flight morphology relative to rainforest disturbance in an Afrotropical butterfly. Biol. J. Linn. Soc. 96: 830-839.
Brock JP (1971) A contribution towards an understanding of the morphology and phylogeny of the Ditrysian Lepidoptera. J. Nat. Hist. 5: 29-102.
Casey TM, Joos BA (1983) Morphometrics, conductance, thoracic temperature, and flight energetics of noctuid and geometrid moths. Physiol. Zool. 56: 160-173.
Dudley R (2000) The Biomechanics of Insect Flight: Form, Function, Evolution. Princeton University Press, Princeton, NJ, USA. 498 pp.
Ellington CP (1984) The aerodynamics of hovering insect flight. II. Morphological parameters. Phil. T. Roy. Soc. B 305: 17-40.
Johansson F, Soderquist M, Bokma F (2009) Insect wing shape evolution: independent effects of migratory and mate guarding flight on dragonfly wings. Biol. J. Linn. Soc. 97: 362-372.
Kitching IJ (1984) An historical review of the higher classification of Noctuidae (Lepidoptera) Bull. Br. Mus. Hist. (Entomol.) 49(3): 153-234.
Kristensen NP (1984) Studies on the morphology and systematics of primitive Lepidoptera (Insecta). Steenstrupia 10(5): 141-191.
Heinrich B (1987) Thermoregulation in winter moths. Sci. Am. 256(3): 82-89.
Miki T, Jacket S (2010) Indirect interactions in the microbial world: specificities and similarities to plant-insect systems. Popul. Ecol. 52: 475-483.
Parra LE, Henriquez-Rodriguez JL (1993) Aportes al conocimiento de las polillas del genero Mallomus Blanchard, 1852 (Geometridae, Nacophorini). Bol. Soc. Biol. Concepción 64: 171-187.
Scoble M (1995) The Lepidoptera: Form Function and Diversity. Oxford University Press, Oxford, UK. 404 pp.
Snodgras RE (1935) Principles of the Insect Morphology. McGraw-Hill. 166-172 pp.
Srygley RB (1994) Locomotor mimicry in butterflies? The associations of positions of centres of mass among groups of mimetic, unprofitable prey. Phil. Trans. Roy. Soc. B 343: 145-155.
Stillwell RC, Davidowitz G (2010) Developmental perspective on the evolution of sexual size dimorphism of a moth. Proc. Roy. Soc. Lond. B Biol. 277: 2069-2074.
Zera AJ, Denno RF (1997) Physiology and ecology of dispersal polymorphism in insects. Annu. Rev. Entomol. 42: 207-230.
Author notes
Alternative link
https://www.interciencia.net/wp-content/uploads/2018/03/202-6275-BENITEZ-43_3.pdf (pdf)