Comunicaciones
ROMPIMIENTO DE LA LATENCIA EN SEMILLA DE HIGUERILLA (Ricinus communis L.)
BREAKING DORMANCY IN CASTOR BEAN (Ricinus communis L.) SEEDS
QUEBRA DE DORMÊNCIA NA MAMONA (Ricinus communis L.)
ROMPIMIENTO DE LA LATENCIA EN SEMILLA DE HIGUERILLA (Ricinus communis L.)
Interciencia, vol. 43, núm. 12, pp. 858-863, 2018
Asociación Interciencia
Recepción: 15/03/2017
Corregido: 01/11/2018
Aprobación: 20/11/2018
Resumen: El biodiesel tiene ventajas ecológicas que han estimulado la investigación, tanto para la obtención de variedades mejoradas con alta productividad, como para la optimización del proceso de obtención del combustible, todo enfocado al desarrollo de tecnologías que permitan sustituir los combustibles fósiles. Uno de los cultivos utilizados con esta finalidad es la higuerilla (Ricinus communis) pero el mismo presenta gran diversidad fenotípica y problemas de germinación. En este estudio se utilizaron 12 tratamientos para el rompimiento de la latencia en la semilla de una población nativa de la región y otra derivada de ella, mutada con fines de mejoramiento. Se usó la refrigeración, ácido giberélico + nitrato de potasio y luz infrarroja. Los mejores resultados (tratamientos 5, 9 y 11) se obtuvieron con la combinación de giberelina + nitrato de potasio, resultando en más del 95% de germinación; algunos tratamientos (frío con luz infrarroja o lavado con metanol) no lograron romper la latencia. Una alta germinación y un buen establecimiento son cruciales para las siembras con fines comerciales. El estudio de la latencia en el cultivo tiene implicaciones importantes, tanto desde el punto de vista fisiológico, como de calidad de la semilla con el fin de que su producción sea rentable y de manejo fácil. En conclusión, fue posible lograr más del 85% de germinación en ambas poblaciones.
Palabras clave: Biodiesel , Giberelinas , Nitrato , Osmoacondicionamiento , Ricinus communis.
Abstract: Biodiesel has ecological advantages that have stimulated research, both in order to obtain improved varieties with high productivity, and to optimize the process of fuel obtaining, all focused on the development of technologies to replace fossil fuels. One of the crops used for this purpose is the castor bean (Ricinus communis) but it presents a large phenotypic diversity and germination problems. In this study, 12 treatments were used to break dormancy in seeds of a native population of the region and another derived from it, mutated for breeding purposes. Refrigeration, gibberellic acid + potassium nitrate and infrared light were used. The best results (treatments 5, 9 and 11) were obtained with the combination of gibberellin + potassium nitrate, with more than 95% germination, while some treatments (cold with infrared light or washing with methanol) failed to overcome dormancy. High germination rate and good sowing are crucial for commercial purpose planting. The study of latency has important implications both physiologically and for seed quality in order to make production cost-effective and easy to handle. In conclusion, it was possible to achieve more than 85% germination in both populations.
Resumo: Biodiesel tem vantagens ecológicas que estimularam a pesquisa, tanto para a obtenção de alta produtividade melhorada, bem como para otimizar o processo de obtenção de combustível, todos focalizados no desenvolvimento de tecnologias para substituir os combustíveis fósseis. Uma das culturas utilizadas para esta finalidade é a mamona (Ricinus communis) mas apresenta uma grande diversidade fenotípica e problemas de germinação. Neste estudo foram utilizados 12 tratamentos para a quebra de dormência nas sementes de uma população nativa da região e de outra derivada dessa, mutada com fins de reprodução. Foram usadas refrigeração, ácido giberélico + nitrato de potássio e luz infravermelha. Os melhores resultados de tratamentos (5, 9 e 11) foram obtidos com a combinação de giberelina + nitrato de potássio, com mais do que 95% de germinação; alguns tratamentos (luz infravermelha frio ou lavado com metanol) não conseguiram quebrar a dormência. A alta germinação e bom estabelecimento é crucial para o plantio comercial. O estudo da latência na cultura tem implicações importantes, tanto ponto de vista fisiológico como da qualidade das sementes, a fim de tornar rentável a produção e de fácil manuseio. Em conclusão, foi possível conseguir mais de 85% de germinação em ambas as populações.
Introducción
Los biocombustibles contienen metabolitos o derivados de la biomasa vegetal y pueden ser obtenidos a partir de cultivos tales como el maíz, caña de azúcar y remolacha, de los que se extraen azúcares para la producción de etanol. De las semillas oleaginosas se puede extraer aceite para la obtención de biodiesel. La producción de biocombustibles ha generado interés en todo el mundo con la finalidad de tener una fuente de energía renovable para el transporte, lo que permitirá reducir la dependencia del petróleo y bajar los índices de contaminación (González-Merino y Castañeda-Zavala, 2008). El cambio de uso de cultivos tradicionales dedicados a la producción de alimentos por cultivos para la generación de biocombustibles ocasiona un déficit en el suministro de granos y materias primas para consumo humano y ganadería, mientras que el cambio en el uso de suelo no apto para la producción de alimentos a fin de dedicarlo a los agrocombustibles es una alternativa que produciría mayores ganancias a los agricultores (Ruiz-Olivares et al., 2013). En la Unión Europea se acordó que para 2020 todos los combustibles utilizados en el transporte deberán contar con 10% de biocombustible, por lo que Europa tendrá que destinar 72% de la superficie agrícola a la producción de energía (Platas-Rosado et al., 2016).
La higuerilla (Ricinus communis L.) es una especie originaria de África que se cultiva con fines industriales y energéticos. Pertenece a la familia de las Euphorbiaceas y tiene una notable capacidad de adaptarse a diferentes ambientes (Garcillán y Rebman, 2016). En México, la investigación con esta especie se inició desde los años 60 en el norte del país, para la obtención de aceite. La implementación de procesos de producción basados en materias primas y su disponibilidad deben optimizarse, para así obtener un producto cuyo costo de producción sea competitivo y de una calidad adecuada (Cruz-Meza et al., 2012). La situación actual en el mundo exige la búsqueda de fuentes alternativas de energía para poder disminuir la contaminación y mantener el ritmo de crecimiento. La intervención y el giro que se está dando en el país a la inversión extranjera, que se está orientando hacia otro tipo de actividades, afecta la producción del campo y ello ocasiona el abandono de tierras de cultivo, las que pueden ser reusadas para la producción de bioenergéticos. Esta industria podría permitir reactivar el sector agrícola sin afectar directamente la producción alimentaria, al utilizar suelos no aptos para cultivos tradicionales (Platas-Rosado et al., 2016).
La higuerilla es considerada un cultivo alternativo de importancia económica y social para las condiciones climáticas de El Bajío, México, debido a su fácil manejo y resistencia a la sequía, y por tratarse de un generador potencial de actividades remunerativas para pequeños agricultores. Debido a su composición química, el aceite de higuerilla conserva su viscosidad a altas temperaturas y resiste las bajas sin congelarse, razón por la cual se le emplea para motores de altas revoluciones. Además, la torta proteica residual también tiene posibles usos en la alimentación del ganado (Silitonga et al., 2016).
En las plantas silvestres la latencia es una característica para la conservación y dispersión de la especie (Hernández et al., 2009). Finch-Savage et al. (2004) mencionan que esta es una propiedad innata que define las condiciones para la germinación; es dependiente de la constitución genética, con influencia ambiental significativa (Rajjou y Debeaujon, 2008). En algunas especies, el efecto inductor de la luz es importante y permite el mantenimiento de la latencia en las simientes enterradas en el suelo (Soriano et al., 2011). Conocer las características que influyen sobre la longevidad de las semillas tiene importancia ecológica, agronómica y económica (O-Melo et al., 2009; Finch-Savage y Bassel, 2016). Los mecanismos de reparación de la semilla, inducidos en condiciones subóptimas para la germinación, pueden mejorar el comportamiento posterior de ésta (Harrigan et al., 2007). Los tratamientos para ello, que se basan en la hidratación controlada de las semillas, se denominan ‘osmoacondicionamiento’ (osmopriming). Durante el tratamiento, las semillas mantienen tolerancia a la desecación debido a la hidratación incompleta y pueden ser secadas otra vez. En la búsqueda de tratamientos que mejoren el comportamiento de las semillas se reportó que al someterlas a la acción de microondas se incrementa la germinación, aumenta la tasa de imbibición y se promueve la redistribución del agua de las células (Anand et al., 2008). Debido a los problemas de germinación, en el caso de la higuerilla se colocan desde dos hasta cuatro semillas por golpe, para asegurar un número suficiente de plántulas (Barrios-Gómez et al., 2013). Por debajo de 20ºC la emergencia de las semillas puede durar de 15 a 20 días, mientras que por encima de 30ºC la emergencia puede ocurrir en 7 días (Barrios-Gómez et al., 2013).
El objetivo de la presente investigación fue determinar el comportamiento germinativo de semillas de higuerilla y desarrollar protocolos de rompimiento de la latencia e inducción de la germinación tratando la semilla con frío, luz infrarroja, nitrato de potasio y ácido giberélico.
Materiales y Métodos
La semilla se obtuvo de una recolección de frutos de una población silvestre ubicada en el municipio de Apaseo el Alto, Guanajuato, México, y de una variedad mutante generada con etil-metano-sulfonato (EMS) en el Instituto Tecnológico de Roque. Las mutantes fueron generadas para tratar de incrementar el contenido de aceite (datos por publicar). La semilla nativa fue utilizada para establecer el tratamiento con la mejor respuesta y luego se comparó este en las dos variedades. La extracción de las semillas se realizó de forma manual, se lavaron al chorro de agua hasta obtener semillas limpias y se secaron a 17ºC durante 48h. Se evaluaron 12 tratamientos (Tabla I) mediante un diseño completamente al azar, con tres repeticiones. Antes de los tratamientos, las semillas se sumergieron en agua por 10min. Se evaluó el efecto de la luz infrarroja aplicándola durante 8h diarias por 3 días; el ácido giberélico (GA3) se aplicó en una mezcla con nitrato de potasio (0,1% de GA3 + 3% de nitrato de potasio), incubando las semillas por 12 y 24h; se hizo un lavado con metanol sacando la semilla inmediatamente; o se incubó durante 15min, luego de un pretratamiento a la semilla de 60h de frio (4ºC). El control se mantuvo a temperatura ambiente en la oscuridad. Después de los diferentes tratamientos las semillas fueron colocadas en papel germinador e incubadas en cámara de germinación (Control SystemLabs®) a 25ºC con 80% de humedad relativa en ausencia de luz, para evaluar el porcentaje de germinación durante 15 días (Moreno 2009).

Acidez titulable
Para cuantificar la acidez titulable (AT) se pesaron 2g de muestra de semillas y se diluyó dos veces su volumen con agua destilada; se añadieron unas gotas de solución de fenolftaleína y se tituló con NaOH 0,1N hasta la aparición de un color rosado que persistió de 15 a 30s. La acidez en la muestra expresada como equivalentes de ácido málico se calculó mediante la fórmula AT= (V×N×67)/M, donde V: volumen de solución de NaOH 0,1N gastado en la titulación de la muestra (ml), N: normalidad de la solución de NaOH, M: volumen de la muestra (ml), y 67: peso equivalente en gramos del ácido málico.
Conductividad eléctrica
Para medir la conductividad eléctrica (CE) se utilizaron 15 semillas, con tres repeticiones. Las semillas fueron pesadas en una balanza de precisión (0,0001g) y colocadas en 30ml de agua desionizada durante: 1, 2, 3, 4, 5, 6, y 12h, a 25ºC. Después de cada período, la CE fue medida (mS) empleando un conductímetro digital marca ACUAPRO.
Análisis estadístico
Para realizar el análisis de varianza (ANOVA) los datos fueron transformados mediante la función arcseno, a fin de satisfacer la condición de homogeneidad de la varianza, la cual se viola cuando los datoas presentan porcentajes de variación menores de 30%y mayores de 70% (Gómez y Gómez, 1984). La significancia estadística se determinó mediante la prueba DMS (P<0,05) utilizando el paquete SAS 9.4.
Resultados y Discusión
En la Tabla I se presentan los resultados de la germinación para la higuerilla. Los tratamientos con mayor respuesta fueron T2, T5, T9 y T11, 93 a 100%. Otros tratamientos, como T8, T10 y T12 fueron menos efectivos para inducir la germinación, y en el resto de los tratamientos, así como en el testigo, la germinación fue de cero. Los tratamientos que resultaron efectivos incluyeron el ácido giberélico, lo cual sugiere que esta fitohormona es importante para romper la dormancia en la higuerilla, sin dejar de considerar el tratamiento con frío. Este resultado concuerda con los hallados por Song et al. (2011), quienes señalan que la síntesis de giberelinas activas es necesaria para la germinación. En semilla seca se han detectado mRNAs de proteínas que intervienen en el metabolismo de las giberelinas, que podrían convertir precursores inactivos en giberelinas activas (Song et al., 2011; Ventura et al., 2012). En semillas de jaúl (Almus acuminata), que requieren de luz para germinar, ésta promueve la síntesis de AG1 (Araya et al., 2000).
Los resultados del análisis de varianza se observan en el Tabla II. Hubo diferencias significativas y al menos una de las líneas difiere entre los tratamientos evaluados. El coeficiente de variación en germinación fue adecuado para esta variable, por lo que se tiene confiabilidad de los resultados.
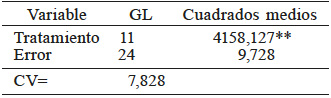
** Significancia estadística al nivel 0,01 de probabilidad.
En la Tabla III se muestran los resultados de la comparación de medias mediante la prueba DMS. Los tratamientos 5, 9 y 11 fueron los mejores, y con los tratamientos 7, 1,6, 3 y 4 no se logró romper la latencia. La combinación de giberelinas más nitrato de potasio resultó ser esencial para el rompimiento de la latencia en semilla de higuerilla bajo las condiciones en que se realizó este experimento. Este resultado concuerda con los obtenidos por Lara et al., (2014) quienes señalan que la nitrato reductasa produce nitritos y óxido nítrico, que interrumpe la dormancia y promueve la germinación.
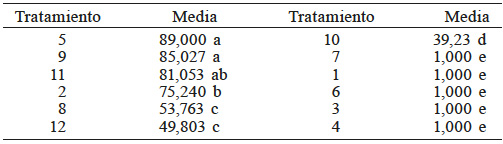
Medias con la misma letra son estadísticamente iguales (DMS, p<0,05).
En la Tabla IV se muestran los resultados de la evaluación de tratamientos para la variable acidez titulable. Hubo diferencias altamente significativas entre tratamientos, por lo que al menos un tratamiento fue efectivo para inducir un cambio de pH en la semilla. Esto es positivo, pues indica actividad o reacomodo metabólico que, aparentemente, le permite a la semilla una mayor germinación. Existe confiabilidad en este resultado ya que el coeficiente de variación fue de 13,4. El modelo propuesto con este análisis explica el 95% de la variación. La interacción línea × tratamiento no fue significativa, lo que indica que estos factores, bajo las condiciones en que se realizó este experimento, fueron independientes. El ácido málico, cítrico y otras moléculas de masa molecular pequeña, pero abundantes, pueden influir de manera importante sobre el comportamiento de la semilla, regulando el pH o, incluso, permitiendo la acumulación o disolución de otros metabolitos (Choi et al., 2011). Se ha reportado que durante los tratamientos pregerminativos cambian los niveles de GTP (guanosin trifosfato) y CTP (citidin trifosfato), incrementándose hasta cinco veces los de UDP-glucosa (glucosa uridin difosfato); estos UDP-azucares son importantes para formar carbohidratos más complejos y de mayor tamaño (Decker y Kleczkowski, 2017) y son necesarios para la síntesis de la pared celular. Otros metabolitos, como los ácidos orgánicos de cadena corta, son metabolizados mediante la respiración, la gluconeogénesis, la fermentación a etanol, la síntesis e interconversión de aminoácidos para la producción de metabolitos secundarios, tales como pigmentos (Famiani et al., 2015). De acuerdo con Sveinsdottir et al. (2009) la H+ATPasa, está activa a las 2h después de la imbibición.
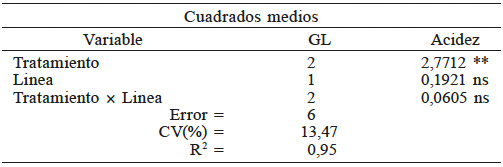
** Significancia estadística al nivel 0,01 de probabilidad; n: no significativo.
En la comparación de medias (Tabla V) se observa que el tratamiento 2 (60h de frío) fue estadísticamente igual al testigo y el tratamiento GA+ KNO3 fue distinto del testigo. Se observó que un decremento en la acidez titulable mejora la germinación de las semillas. Se ha reportado que la cantidad de proteínas solubles y aminoácidos libres es mayor en semillas osmoacondicionadas, lo que podría explicar el efecto sobre el pH (Zhang et al., 2015). Se ha encontrado que durante la rehidratación el agua, así como moléculas anfifílicas, se redistribuyen en la semilla (Anand et al., 2008; Ventura et al., 2012).
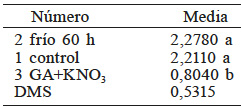
Medias con la misma letra son estadísticamente iguales (DMS, p<0,05).
Las variaciones observadas en la germinación se pueden deber a variaciones en el grado de desarrollo de la semilla en general, del embrión, de la presencia de ciertas sustancias como el ácido abscísico, y la ausencia o baja concentración de giberelinas. La dormancia puede deberse a aspectos físicos, como que la testa sea gruesa e impermeable. La acumulación de ácido abscísico también impide la germinación y puede degradarse mediante estratificación. Además, la impermeabilidad de la testa puede superarse mediante escarificación (Née et al., 2017).
En la Tabla VI se observa que hubo diferencias altamente significativas entre tratamientos para la variable conductividad eléctrica en los tiempos C1, C2 y C3; después de los cuales no se observaron diferencias. En la Tabla VII se observa que el tratamiento con frío fue estadísticamente igual al testigo; nuevamente, el tratamiento con AG3+KNO3 mostró una menor conductividad eléctrica, lo que significa que la semilla tuvo cambios a nivel de membranas que impiden la fuga de solutos y esto mejora la germinación. De acuerdo con Yu et al. (2015) durante la rehidratación las membranas de la semilla deben reorganizarse rápidamente para evitar la fuga de solutos, lo que parece estar sucediendo con los tratamientos favorables a la germinación, dado los efectos benéficos de las giberelinas y el nitrato de potasio sobre esta (Neé et al., 2017).
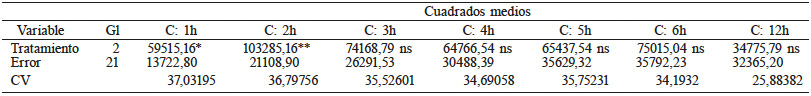
C: tiempo de incubación. * y **: Significancia estadística al nivel 0,01 y 0,05 de probabilidad.
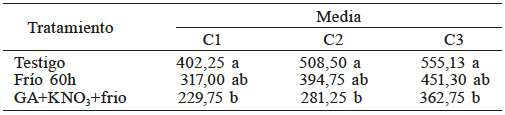
C1: incubación 1h, C2: incubación 2h, C3: incubación 3h. Medias con la misma letra son estadísticamente iguales (DMS, p<0,05).
La medición de la conductividad eléctrica mostró que hay diferencias en el comportamiento de las mismas; el osmoacondicionamiento previene la lixiviación y por lo tanto incrementa el porcentaje de germinación (Tabla III). De acuerdo con Yu et al. (2015) la imbibición provoca un cambio de fase en las membranas, de fase hexagonal a fase lamelar. La conservación de solutos como sacarosa y rafinosa permiten que la semillas mantengan el estado vítreo, que retarda la difusión molecular (Pereira-Lima et al., 2017).
En relación a la prueba de conductividad eléctrica, las mediciones realizadas permitieron diferenciar las poblaciones estudiadas. La integridad de las membranas celulares, determinada por los cambios bioquímicos que deterioran la semilla y disminuyen la capacidad para reorganizar y reparar daños, es la causa fundamental de las diferencias en el vigor de las semillas, que son medidas en forma indirecta a través de la lixiviación de electrolitos durante la prueba de conductividad eléctrica (Plazek et al., 2018). Los lotes de semillas que muestran una elevada germinación en laboratorio, pero liberan grandes cantidades de electrolitos luego de la inmersión en agua, son considerados de bajo vigor, presentando un bajo desempeño en condiciones de estrés (Plazek et al., 2018). Contrariamente, lotes con una alta germinación y baja liberación de electrolitos son considerados de alto vigor y con mejor capacidad para soportar condiciones de estrés. Anand et al. (2008) encontraron que al tratar las semillas con microondas se incrementa la conductividad y se mejora la germinación; por su parte Mei y Song (2008) señalan que la germinación incluye la salida de solutos desde la semilla, que dependiendo de las condiciones externas, pueden ser reabsorbidos. La salida de solutos permite la pérdida de moléculas inhibitorias de la germinación, aunque también puede ser indicativa de daño de las membranas (Ventura et al., 2012).
Finalmente, se determinó la acidez titulable. En la Tabla III se observa que hay significancia al nivel 0,01 de probabilidad, lo cual indica que en al menos uno de los tratamientos propuestos se observa un cambio de acidez. Aquellas semillas tratadas con el osmoacondicionamiento tienen menor acidez y esto podría estar relacionado con la liberación de iones. El estado vítreo de la semilla retarda la difusión molecular y las reacciones que provocan deterioro. El incremento en la movilidad molecular se correlaciona con un declive en la viabilidad (Gurusinghe y Bradford, 2001). El cambio en la acidez titulable sugiere este cambio en el estado vítreo de la semilla y la movilidad de moléculas tales como los ácidos orgánicos. En las semillas osmoacondicionadas se incrementa la poza de proteínas solubles y aminoácidos libres, lo que sin duda incide en la acidez titulable (Zhang et al., 2015).
La comparación de los efectos de los tratamientos en las semillas nativas (silvestres) y en las mutantes obtenidas en el laboratorio mostró (Figura 1) que en ambos casos las semillas tratadas con GA3+KNO3+ frío tuvieron el mayor porcentaje de germinación, y que estos tratamientos son los que presentan una menor acidez titulable (Figura 2). La mutante germina un 9% más que la nativa sin tratamiento y con tratamiento de frío; cuando se aplicó tratamiento de giberelinas, frío y nitrato la nativa alcanzó 100% de germinación y la mutante 85%.
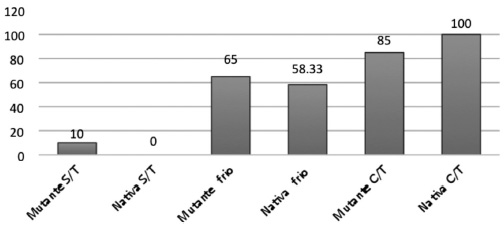
Figura 1
Germinación de semilla de higuerilla mutante y nativa sin tratamiento (S/T)
Con frío 60h y con tratamiento (C/T) de Giberelina +Nitrato de Potasio 24h + luz infrarroja + frio 60h. DMS= 4,19.
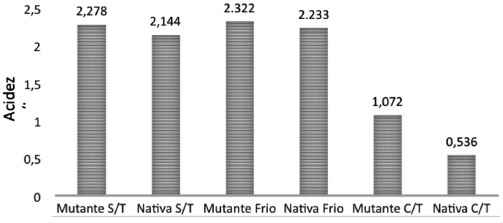
Figura 2
Acidez titulable en semilla de higuerilla mutante y nativa sin tratamiento (S/T)
Con frío 60h y con tratamiento (C/T) de Giberelina +Nitrato de Potasio 24h + luz infrarroja + frio 60. DMS= 0,411.
Conclusiones
Los tratamientos aplicados permitieron el rompimiento de la latencia en la semilla de higuerilla. Los mejores tratamientos fueron el 5, 9, 11 y 2, que incluyeron giberelinas más nitrato de potasio. Se encontraron diferencias en acidez titulable, el pH de las semillas con osmoacondicionamiento se incrementa ligera pero significativamente y son las que presentan mayor porcentaje de germinación. La conductividad eléctrica se incrementa conforme avanza el tiempo de incubación pero en las semillas osmoacondicionadas hay diferencias significativas entre los valores medidos al inicio de la imbibición y aquellos medidos al final. En el testigo no hay diferencias estadísticas significativas entre los valores iniciales y finales.
REFERENCIAS
Anand ANS, Joshi DK, Verma APS, Kar A (2009) Microwave seed treatment reduce hardseedness in Stylosanthes seabrana and promotes redistribution of cellular waters as studied by NMR relaxation measuresments. Seed Sci. Technol. 37: 88-97. doi: 10.15258/sst.2009.37.1.11
Araya EGL, Hidalgo N, Valverde R (2000) Efecto de la luz y del ácido giberélico sobre la germinación in vitro de jaul (Alnus acuminata). Agron. Costarric. 24: 75-80.
Barrios-Gómez EJ, Canul-Ku AZCJ, Hernández-Arenas MG, Alarcón-Cruz N, Chepetla-Calderón VH (2013) Evaluación de materiales elite de higuerilla (Ricinus communis L.) en Morelos. Rev. Cienc. Tecnol. Agropec. Méx. 1: 27-32.
Choi YHC, SJ Dai Y, Verberne M, Hollmann F, Arends IWCE, Witkamp GJ, Verpoorte R (2011) Are Natural Deep Eutectic Solvents the Missing Link in Understanding Cellular Metabolism and Physiology? Plant Physiol. 156: 1701-1705. doi: 10.1104/pp.111.178426
Cruz-Meza P, Márquez-Rosano C, López-Canteñs G (2012) Determinación de las propiedades físico-mecánicas para el descascarado de las capsulas de la higuerilla (Ricinus communis L.). Rev. Mex. Cienc. Agríc. 3: 644-649.
Decker D, Kleczkowski LA (2017) Substrate specificity and inhibitor sensitivity of plant UDP-sugar producing pyrophosphorylases. Front. Plant Sci. 8:1610. doi: 10.3389/fpls.2017.01610
De O-Melo ZL, Goncalves JFC, Mazzafera P, dos Santos DYAC (2009) Mobilization of seed reserves during germination of four tropical species of the amazon rain forest. Seed Science and Technology 37(3): 597-607 doi:10.15258/sst.2009.37.3.09
Famiani F, Battistelli A, Moscatello S, Cruz-Castillo JG, Walker RP (2015) The organic acids that are accumulated in the flesh of fruits: occurrence, metabolism and factors affecting their contents - a review. Rev. Chapingo 21: 97-128. doi: 10.5154/r.rchsh. 2015.01.004
Finch-Savage WE, Bassel GW (2016) Seed vigour and crop establishment: extending performance beyond adaptation. J. Exp. Bot. 67: 567-591. doi: 10.1093/jxb/erv490
Finch-Savage WE, Dent KC, Clark LJ (2004) Soak conditions and temperature following sowing influence the response of maize (Zea mays L.) seeds to on-farm priming (pre-sowing seed soak). Fields Crops Res. 90: 361-374. doi: 10.1016/j.fcr.2004.04.006
Garcillán PP, Rebman JP (2016) Nota sobre los primeros registros históricos de dos plantas invasoras en la Peninsula de Baja California: crónica del misionero jesuita Miguel del Barco. Acta Bot. Mex. 115: 43-49. doi: 10.21829/abm115.2016.1110
Gomez KA, Gomez AA (1984) Statistical Procedures for Agricultural Research. 2a ed. Wiley. Nueva York, EEUU. 680 pp.
González-Merino A, Castañeda-Zavala Y (2008) Biocombustibles, biotecnología y alimentos: Impactos sociales para México. Argumentos 21(57): 55-83.
Gurusinghe S, Bradford KJ (2001) Galactosyl-sucrose oligosaccharides and potential longevity of primed seed. Seed Sci. 11: 121-133. doi: 10.1079/SSR200167
Harrigan GG, Susan GR, Reynolds TI, William PR, James DM, Susan M, Steven C Ha, Orth R, Smith RG, Wen L, Wayne EB, Welsch M, Riley R, Mcfarland D, Pandravada A, Glenn KC (2007) Impact of Genetics and Environment on Nutritional and Metabolite Components of Maize Grain. J. Agric. Food Chem. 55: 6177-6185. doi: 10.10 21/jf070494k.
Hernández P, Lobo A, Medina C, Cartagena V, Delgado P (2009) Comportamiento de la germinación y categorización de la latencia en semillas de mortiño (Vaccinium meridionale Swartz). Agron. Colomb. 27: 15-23.
Lara TS, Lira JMS, Rodríguez A, Rakocevic M, Alvarenga A (2014) Potassium nitrate priming affects the activity of nitrate reductase and antioxidants enzymes in tomato germination. J. Agric. Sci. 6: 72-80. doi: 10.5539/jas.v6n2p72
Mei YQ, Song SQ (2008) Early morphological and physiological events occurring during germination of maize seeds. Agric. Sci. China 7: 950-957. doi: 10.1016/S1671-2927(08)60134-0
Melo ZLdeO, Goncalves JFdeC, Mazzafera P, Dossantos DAC (2009) Mobilization of seed reserves during germination of four tropical species of the amazon rain forest. Seed Sci. Technol. 37: 597-607.
Moreno EM (2009) Análisis Físico y Biológico de Semillas Agrícolas. Instituto de Biología, UNAM. México. 393 pp.
Née G, Xiang Y, Soppe WJ (2017) The release of dormancy, a wake-up call for seeds to germinate. Curr. Opin. Plant Biol. 35: 8-14. doi: 10.1016/jpbi.2016.09.002
Pereira-Lima JJ, Buitink J, Lalanne D, Falopa-Rossi R, Pelletier S, Amaral da Silva EA, Leprince O (2017) Molecular characterization of the acquisition of longevity during seed maturationin soy bean. PlosOne 12(7):e0180282. doi: 10.1371/journal.pone.0180282
Platas-Rosado DE, Zetina-Córdoba P, Vilaboa-Arroniz J, Martínez-Hernández R (2016) Adaptación y mitigación del cambio climático con la producción de bioenergéticos en suelos marginales. Rev. Mex. Cienc. Agríc. (Publ. esp.) 14: 2857-2866.
Plazek A, Dubert F, Kopec P, Dziurka M, Kalandyk A, Pastuszak J, Wolko B (2018) Seed hydropriming and smoke water significantly improve low-temperature germination of Lupinus angustifolius L. Int. J. Mol. Sci. 19: 992. doi: 10.3390/ijms19040992
Rajjou Loïc DI (2008) Seed longevity: Survival and maintenance of high germination ability of dry seeds. Comp. Rend. Biol. 331: 796805. doi: /10.1016/j.crvi.2008.07.021
Ruiz-Olivares A, González-Chávez MC, Soto-Hernández RM (2013) Potential of castor bean (Ricinus communis L.) for phytoremediation of mine tailings and oil production. J. Environ. Manag. 114: 316-323. doi: 10.1016/j.jenvman.2012.10.023
Silitonga AS, Ong HC, Yusaf T, Kasumo F, Mahlia TMI (2016) Synthesis and optimization of Hevea brasiliensis and Ricinus communis as feedstock for biodiesel production: A comparative study. Indust. Crops Prod. 85: 279-286. doi: 10.10 16/j.indcrop.2016.03.017.
Song J, Song F, Peng H, Yao Y, Zhang Y, Sun ZN (2011) Genome-wide identification of gibberellins metabolic enzyme genes expression profiling analysis during seed germination in maize. Gene 482: 34-42. doi: 10.1016/j.gene.2011.05.008.
Soriano D, Márquez-Guzman J, Kitajima K, Gamboa de Buen A, Huante P (2011) Seed reserve composition in 19 tree species of a tropical deciduous forest in México and its relationship to seed germination and seedling growth. Ann. Bot. 107: 939-951. doi: 10.1093/aob/mcr041
Sveinsdottir H, Yan F, Zhu Y, Peiter-Voik T, Schubert S (2009) Seed ageing-induced imbibition of germination and post-germination root growth is related to lower activity of plasma membrane H+-ATPase in maize roots. J. Plant Physiol. 166: 128-135. doi: 10.1016/j.jplph. 2008.01.012.
Ventura L, Donà M, Macovei A, Carbonera D, Buttafara A, Mondoni A, Rossi G, Balestrazzi A (2012) Understanding the molecular pathways associated with seed vigor. Plant Physiol. Biochem. 60: 196-206. doi: 10.10 16/j.plaphy. 2012.07.031.
Yu X, Li A, Li W (2015) How membranes organize during seed germination: three patterns of dynamic lipid remodeling define chilling resistance and affect plastid biogenesis. Plant Cell Environ. 38: 1391-1403. doi: 10.1111/pce.12494.
Zhang F, Johnston CR, Wang Y, Zhu K, Lu F, Zhang Z, Zhou J (2015) Seed priming with polyethylene glycol Induces physiological changes in sorghum (Sorghum bicolor L. Moench) seedlings under suboptimal soil moisture environments. PLoS ONE 10(10), e0140620. doi: 10. 1371/journal.pone.0140620
Notas de autor
Autor de correspondencia: Juan Carlos Raya Pérez. e-mail: juraya@itroque.edu.mx
Enlace alternativo
https://www.interciencia.net/wp-content/uploads/2018/12/858-RAYA-43_12.pdf (pdf)