Comunicaciones
CRECIMIENTO Y PRODUCCIÓN DE CHÍA (Salvia hispanica L.) EN FUNCIÓN DE LA IRRADIANCIA Y FERTILIZACIÓN ORGÁNICA
CRESCIMENTO E PRODUÇÃO DE CHIA (Salvia hispanica L.) EM FUNÇÃO DA IRRADIÂNCIA E FERTILIZAÇÃO ORGÂNICA
GROWTH AND YIELD OF CHIA (Salvia hispánica L.) AS A FUNCTION OF IRRADIANCE AND ORGANIC FERTILIZATION
CRECIMIENTO Y PRODUCCIÓN DE CHÍA (Salvia hispanica L.) EN FUNCIÓN DE LA IRRADIANCIA Y FERTILIZACIÓN ORGÁNICA
Interciencia, vol. 44, núm. 6, pp. 340-346, 2019
Asociación Interciencia
Recepción: 04 Octubre 2018
Corregido: 17 Junio 2019
Aprobación: 18 Junio 2019
Resumen: Se evaluó el crecimiento y rendimiento de chía (Salvia hispanica L.) cultivada bajo diferente irradiancia generada por malla sombra de color y nutrición orgánica, siendo la hipótesis de trabajo que la combinación de fertilizantes orgánicos y disminución en la irradiancia afecta el desarrollo del cultivo y rendimiento de la semilla. El estudio se realizó en campo, la siembra fue manual y se instalaron 12 tratamientos, combinación de las mallas, la aplicación de vermicompost y aplicación foliar de té de vermicompost. El cultivo se cubrió con mallas sombra de color azul, rojo y verde, con un testigo sin malla. El flujo fotónico fue de 691, 720, 758 y 1848µmol·m-2·s-1, respectivamente. El diseño experimental fue de bloques al azar con arreglo en parcelas divididas con cuatro repeticiones. En muestreos mensuales se registraron variables de crecimiento, y a la cosecha variables de rendimiento. El crecimiento de chía se modificó por los cambios de irradiancia y aplicación de vermicompost. El cultivo bajo malla verde (758µmol·m-2·s-1) presentó una biomasa y tasa de crecimiento absoluta más alta pero menor rendimiento de semilla. Con el uso de malla sombra se acorta el ciclo del cultivo y el rendimiento de semilla no se afecta. La combinación de vermicompost y té de vermicompost como fuente de nutrimentos incrementó la biomasa y el rendimiento de semilla.
Palabras clave: Biomasa Total, Irradiancia, Nutrición Orgánica, Rendimiento, Salvia hispanica L.
Resumo: Avaliou-se o crescimento e rendimento de chia (Salvia hispanica L.) cultivada sob particular irradiância gerada por malha de sombreamento colorida e nutrição orgânica, sendo a hipótese de trabalho que a combinação de fertilizantes orgânicos e diminuição na irradiância afeta o desenvolvimento do cultivo e rendimento da semente. O estudo foi realizado em campo, a plantação foi manual e instalados 12 tratamentos, combinações de malhas, aplicação de vermicompostagem e aplicação foliar de chá de vermicompostagem. O cultivo foi coberto com malhas de sombreamento na cor azul, vermelho e verde, com testemunha sem malha. O fluxo fotônico foi de 691, 720, 758 e 1848µmol·m-2·s-1, respectivamente. O desenho experimental foi de blocos aleatórios com arranjos em parcelas divididas com quatro repetições. Amostragens mensais permitiram registrar variáveis de crescimento e, na colheita, variáveis de rendimento. O crescimento de chia foi alterado devido às mudanças de irradiância e aplicação de vermicompostagem. O cultivo sob malha verde (758µmol·m-2·s-1) apresentou biomassa e taxa de crescimento absoluta mais alta, embora com menor rendimento de semente. Com o uso de malha de sombreamento, diminui o ciclo do cultivo e o rendimento de semente não é afetado. A combinação de vermicompostagem e chá de vermicompostagem como fonte de nutrientes incrementou a biomassa e o rendimento de semente.
Abstract: The growth and yield of chia (Salvia hispanica L.) was evaluated. Cultivated under different irradiances generated by color shade meshes and organic nutrition, the working hypothesis being that the combination of organic fertilizers and decrease in irradiance affects crop development and seed yield. The study was conducted in the field, planting was manual and 12 treatments were installed: combination of the meshes, application of vermicompost and foliar application of vermicompost tea. The crop is covered with blue, red and green mesh shades, with a mesh-less control. The photonic flux was 691, 720, 758 and 1848μmol·m-2·s-1, respectively. The experimental design was of randomized blocks in divided plots, with four repetitions. Growth and harvest yield variables were registered in monthly samplings. Chia growth was modified by the changes of irradiance and application of vermicompost. The crop under green mesh (758μmol·m-2·s-1) presented higher biomass and absolute growth rate but lower seed yield. With the use of shadow mesh, the cultivation cycle is shortened and seed yield is not affected. The combination of vermicompost and vermicompost tea as a source of nutrients increased the biomass and seed yield.
Introducción
La chía (Salvia hispanica L.) es un grano de consumo ancestral con propiedades funcionales que se recomienda incorporar a la dieta básica por ser fuente de ácidos grasos omega-6 y omega-3, y presentar compuestos antioxidantes (Di Sapio et al., 2012), fibra dietética y proteínas (Mohd et al., 2012). Dentro de las estrategias para favorecer el desarrollo de los cultivos e incrementar la producción está el uso de mallas foto-selectivas, que al cambiar la intensidad y la calidad de la luz alteran el microclima y el consumo de energía (Milenković et al., 2012). Por ejemplo, el sustituir la malla negra por mallas de otros colores en la producción de pimientos y tomates cambia la cantidad y calidad de luz, lo que se refleja en el incremento en rendimiento y calidad de los productos (Ben-Yakir et al., 2012). Las coberturas de colores o filtros fotoselectivos es una técnica agrícola que proporciona al agricultor alternativas reales de manejo en el cultivo (Casierra y Peña, 2015).
El vermicompost es rico en bacterias fijadoras de nitrógeno, solubilizadoras de fosfato, actinomicetos y hormonas de crecimiento como auxinas, citoquininas y giberelinas (Adhikary, 2012; Kenyangi y Blok, 2012) que ayudan a llevar a cabo el reciclaje eficiente de nutrientes y contribuyen al desarrollo de las plantas y mejoran la calidad del suelo (Sharma et al., 2005). Su aplicación al suelo y hojas (como té de vermicompost) aumenta el rendimiento y la calidad del pepino (Cucumis sativus L; Ozdamar et al., 2011).
El manejo de la luz y de la nutrición orgánica son técnicas que podrían ser útiles para mejorar el crecimiento y la producción de chía. Para el análisis de crecimiento del cultivo, se utilizan índices como la razón de área foliar (RAF), la razón del peso foliar (RPF), el área foliar específica (AFE) y la tasa de crecimiento absoluto (TCA). La aplicación de estos índices permite determinar los factores del ambiente y de manejo del cultivo que influyen sobre cada estructura de la planta y en consecuencia en su crecimiento y rendimiento (Gardner et al., 1990; Hunt et al., 2002; Villar et al., 2004;Nassi et al., 2011). El objetivo del presente estudio fue determinar el efecto de la variación en la irradiación y de la nutrición sobre el crecimiento y rendimiento de la chía cultivada bajo condiciones de campo, bajo la hipótesis que la combinación de fertilizantes orgánicos y los cambios en la irradiancia modifican el crecimiento y rendimiento de la semilla del cultivo.
Materiales y Métodos
El estudio se realizó en el periodo de lluvias estacionales en el verano de 2015, en el Colegio de Postgraduados, Montecillo, México (19°29´N; 98°54´O; 2250msnm), de clima templado (Cw) (García, 2004). El suelo es de textura franco limosa; pH de 7,8; CE de 2,93dS·m-1; 2% de materia orgánica y densidad aparente de 1,2g·cm-3. La semilla de chía, adquirida localmente a un productor, fue de la región de Texcoco, del ciclo anterior. Se establecieron 12 tratamientos, como resultado de la combinación de dos factores: 1) cuatro niveles de irradiancia producto de mallas sombra (M) de color azul (MA), roja (MR), verde (MV) y testigo sin malla (SM); y tres niveles de fertilización orgánica (F): vermicompost (FV), vermicompost más aplicación foliar de té de vermicompost (FVT) y testigo sin fertilización (SF).
El diseño experimental fue en parcelas divididas con cuatro repeticiones. La parcela mayor correspondió al factor malla y la parcela menor al factor fertilización. La unidad experimental fue de cinco surcos de 5m de longitud por 0,5m de ancho. Las mallas sombra utilizadas fueron de polietileno de alta densidad con 80% de sombra, colocadas al momento de la siembra a una altura de 1,5m. En condiciones de cielo despejado, con un sensor lineal de quantum modelo LI-191, se registró el flujo de fotones fotosintéticos de la radiación fotosintéticamente activa (RFA), el cuál fue de 1848µmol·m-2·s-1 para el testigo (SM) y de 691µmol·m-2·s-1 con MA, 720µmol·m-2·s-1 con MR y 758µmol·m-2·s-1 con MV, lo que generó una reducción de fotones de 86, 61 y 59%, respecto al testigo. El análisis químico del vermicompost indicó un pH de 8,5; materia orgánica 19,56%; CIC 25,2cmol(+)·kg-1, CE 6,76dS·m-1, N 1,47% y P, K, Ca, Mg y Na con 23, 5939, 900, 73 y 1283mg·kg-1, respectivamente (Romero et al., 2017). La fertilización consistió en la aplicación de vermicompost al suelo a razón de 12Mg·ha-1 a los 30, 60, 90 y 120 días después de la siembra (dds). El té de vermicompost se obtuvo solubilizando en agua el vermicompost (González et al., 2013). Las aplicaciones foliares fueron realizadas cada 30 días, para lo cual el té se diluyó con agua hasta obtener una CE de 0,5dS·m-1 y se ajustó a un pH de 5,5 con H2SO4 con la utilización de un medidor portátil (Conductronic PC18). Durante el ciclo de cultivo se midió los días a la ocurrencia de las fases fenológicas, cuando en más del 50% de las plantas se presentó: emergencia (E), aparición de primordio floral (R1), el inicio de floración (R2), inicio de fructificación (R3) y la madurez fisiológica (MF). Para evaluar el crecimiento del cultivo se realizaron muestreos destructivos mensuales en los que se midió la altura de planta (ALT) con una regla graduada, el diámetro de tallo (DT) con un vernier digital y el área foliar (AF) con un integrador electrónico (Li-Cor™ 3100). Se calculó la razón de área foliar (RAF) mediante la expresión RAF= AF/W, donde AF: área foliar y W: peso seco total; la razón de peso foliar (RPF) con la expresión RP= PH/W, donde PH: peso de las hojas; y el área foliar específica (AFE) con AFE= AF/PH. Para calcular la tasa media de crecimiento absoluto (TCA) se utilizó la ecuación TCA= [(PS2-PS1) / (T2-T1)] donde: PS2 y PS1: peso de la materia seca de la planta en los tiempos T2 y T1, respectivamente (Hunt, 1990; Escalante y Kohashi, 1993). A la cosecha se evaluó número de nudos y brotes, número y peso de espigas, el rendimiento de semilla (RS; g·m-2 al 12% de humedad) y la biomasa total (BT; kg·m-2) que representa la suma de la masa seca (MS) de todos los órganos de la planta en cada muestreo. La MS se obtuvo mediante el secado del material vegetal una estufa de aire forzado a 72°C por 72h. A las variables de estudio se les aplicó pruebas de normalidad. Después, se procedió al análisis de varianza con (α=0,05), prueba de homocedasticidad e independencia en los residuos. Finalmente, se realizaron pruebas de comparación de medias de Tukey con (α=0,05). Los análisis se realizaron con el paquete estadístico SAS V9.4 (SAS, 2015).
Resultados y Discusión
Fenología del cultivo
El sombreado generado por la malla de color afectó los días a ocurrencia de las fases fenológicas (Tabla I). Como se explicó en la metodología, al tener al menos el 50% de las plantas en estudio en una etapa fenológica se encontró que: la emergencia se presentó dos días antes con la malla de color azul (691µmol·m-2·s-1) y tres días antes con la de color rojo (720µmol·m-2·s-1) respecto a la malla verde (758µmol·m-2·s-1) y el testigo (SM; 1848µmol·m-2·s-1). La aparición del primordio floral, la floración y el llenado de frutos, se presentó ocho y diez días después en las plantas bajo la malla verde y el testigo, respectivamente, por lo cual el ciclo de cultivo se retrasó 12 y 13 días con respecto a las mallas azul y roja.
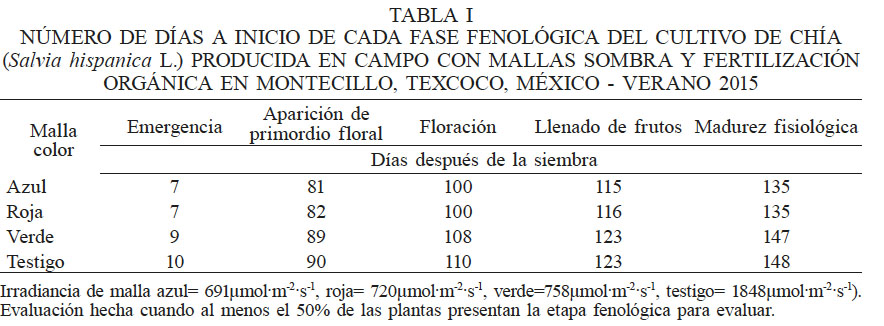
Irradiancia de malla azul= 691µmol·m-2·s-1, roja= 720µmol·m-2·s-1, verde=758µmol·m-2·s-1, testigo= 1848µmol·m-2·s-1). Evaluación hecha cuando al menos el 50% de las plantas presentan la etapa fenológica para evaluar.
La incorporación de mallas sombra de colores azul y rojo acelera el desarrollo de S. hispanica L. y acorta su ciclo de vida. Cambios en los días a ocurrencia de las etapas fenológicas en chía se han observado al evaluar el efecto de fechas de siembra (Arriagada, 2014;Tello, 2014). Se ha documentado que la chía es sensible al fotoperíodo; en Argentina, Busilacchi et al. (2013) reportaron un ciclo de cultivo de chía hasta de 170 días cuando se siembra a principios de enero, y está relacionado con la latitud de la región de siembra.
Razón de área foliar, razón de peso foliar y área foliar específica
El análisis estadístico (P≤0,05), mostró diferencias altamente significativas por efecto de factores principales (color de malla y fertilización) para RAF, RPF y AFE (Tabla II). En los tres índices hubo incremento conforme avanzó el ciclo de cultivo, a excepción del último muestreo, donde RAF y RPF presentaron reducción debido a la disminución del área foliar, ocasionada por senescencia del cultivo. El uso de mallas sombra incrementó el material asimilatorio por unidad de materia seca (mayor RAF) y la aplicación de fertilizantes orgánicos influyó en la RAF a los 100 y 130 dds, ya que las plantas donde se aplicó vermicompost al suelo y el té de vermicompost foliar registraron los valores más altos.
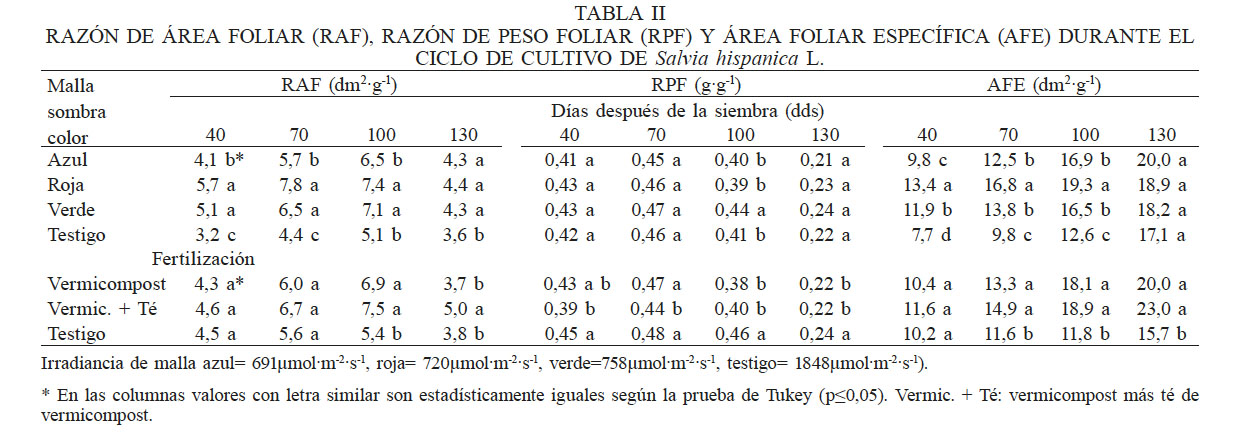
* Irradiancia de malla azul= 691µmol·m-2·s-1, roja= 720µmol·m-2·s-1, verde=758µmol·m-2·s-1, testigo= 1848µmol·m-2·s-1). * En las columnas valores con letra similar son estadísticamente iguales según la prueba de Tukey (p≤0,05). Vermic. + Té: vermicompost más té de vermicompost
El mayor gasto de materia seca para formar la maquinaria fotosintética (RPF) se obtuvo con la malla verde (758µmol·m-1·s-1) a los 100 dds (Tabla II), lo cual se refleja en una mayor producción de biomasa (Tabla IV), mientras que con el testigo (1848µmol·m-1·s-1) se observó mayor acumulación de materia seca en las hojas en relación a la total acumulada en el dosel (RPF). De acuerdo a lo reportado por Hunt et al. (2002), el uso de estos índices permite comparar la eficiencia del dosel vegetal entre especies, cultivares y tratamientos en función de fechas de siembra, nutrimentos y cultivares.
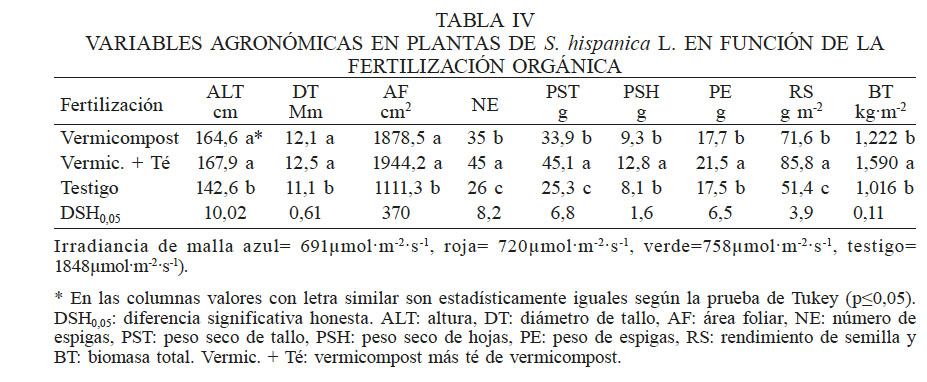
* Irradiancia de malla azul= 691µmol·m-2·s-1, roja= 720µmol·m-2·s-1, verde=758µmol·m-2·s-1, testigo= 1848µmol·m-2·s-1).
* En las columnas valores con letra similar son estadísticamente iguales según la prueba de Tukey (p≤0,05). DSH0,05: diferencia significativa honesta. ALT: altura, DT: diámetro de tallo, AF: área foliar, NE: número de espigas, PST: peso seco de tallo, PSH: peso seco de hojas, PE: peso de espigas, RS: rendimiento de semilla y BT: biomasa total. Vermic. + Té: vermicompost más té de vermicompost.
El espesor de la hoja, que se relaciona con el AFE, presentó diferencias estadísticas significativas (P≤0,05) por efecto de las mallas sombra a los 40, 70 y 100 dds. Con la malla roja se presentó la mayor producción de hojas y se refleja con el valor más alto de AFE (Tabla II). Este desarrollo foliar está relacionado con el efecto Emerson, ya que bajo los filtros fotoselectivos rojos, las longitudes de onda están en el rango de los 670 a los 700nm, y en esta condición hay un aumento en la tasa de fotosíntesis. Bajo la luz roja, que es un componente del espectro visible, es suficiente para que la planta crezca y realice su proceso fotosintético (Lu et al., 2012; Olle y Virsile, 2013), considerando que las auxinas son responsables de la distensión celular y que son capaces de migrar a los sitios menos iluminados de ambas caras de la lámina foliar y concentrarse, lo que provoca mayor crecimiento celular (Rodríguez et al., 2016).
El cociente entre el área y el peso de la hoja indica que la planta en sus primeras etapas de crecimiento invierte en la formación de hoja, por lo que esta expresión es útil para explicar en ocasiones hasta el 80% de la variación la tasa de crecimiento de la planta (Vega et al., 2012), no obstante que otros factores puedan contribuir sobre la tasa de crecimiento, como la proporción de biomasa distribuida en los diferentes órganos, las tasas de fotosíntesis y de respiración, o la composición química de la hoja (Villar et al., 2004). A lo largo del desarrollo del cultivo el AFE presentó diferencias estadísticas (P≤0,05) significativas entre las plantas fertilizadas (en suelo ó en suelo + foliar) y el testigo (sin fertilización) que presentó el AFE más bajo, valor que se asocia con hojas más duras o más densas, lo cual es una ventaja en un ambiente de baja fertilidad, las hojas persisten más y el periodo de retorno del carbono y nitrógeno invertidos se prolonga (Poorter et al., 2009; Brouillette y Donovan, 2011).
Tasa de crecimiento absoluto
La dinámica de la TCA de la chía en función de la irradiancia (M) y la fertilización orgánica (F) se ajustó a un modelo de regresión cuadrática (Figura 1a, b). Las plantas con malla verde presentaron una TCA más alta con un máximo de 23,3g·m-2·d-1 (95 dds), superior en 43, 70 y 72% a la TCA encontrada con la malla roja, testigo y malla azul. En la Figura 1a la ecuación con la malla verde, muestra a los 95 dds la TCA más alta (3,89g·m-2·d-1), seguido de la malla roja con 2,3g·m-2·d-1. Los valores más bajos corresponden al testigo y a la malla azul con 1,78 y 1,70g·m-2·d-1, respectivamente.
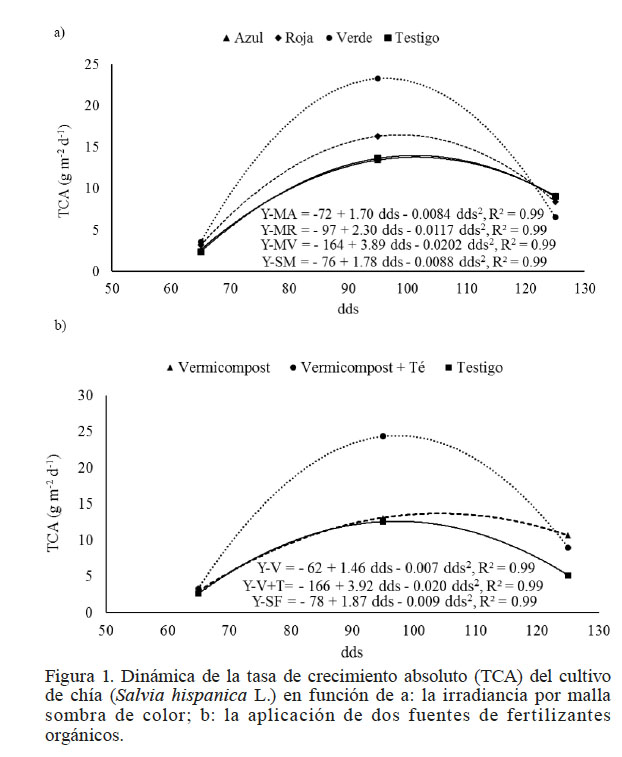
Figura 1
Dinámica de la tasa de crecimiento absoluto (TCA) del cultivo de chía (Salvia hispanica L.) en función de a: la irradiancia por malla sombra de color; b: la aplicación de dos fuentes de fertilizantes orgánicos.
La fertilización con vermicompost y aplicación foliar de té de vermicompost estimuló significativamente (P≤0,05) la TCA, con 24,3g·m-2·d-1. La TCA más baja se encontró en el testigo y cuando se aplicó únicamente vermicompost con 1,.6 y 13,1g·m-2·d-1, respectivamente. Esto indica que en ocasiones la aplicación de vermicompost no es suficiente para cubrir el requerimiento nutrimental del cultivo, y se presenta una respuesta favorable con la fertilización foliar.
La aplicación foliar del té de vermicompost, incrementó la producción de materia seca en cada fecha de muestreo (Figura 1b). El incremento en la TCA con FVT pueden atribuirse tanto al contenido mineral como a las sustancias promotoras del crecimiento presentes en estas fuentes orgánicas (Ingham, 2005;Pant et al., 2011), lo que provoca aumentos en el tamaño del dosel vegetal, medido a través del área foliar. Además, se retrasa la senescencia foliar y se prolonga su duración y actividad fotosintética (Fageria y Baligar, 2005). Al final del ciclo de cultivo la TCA se reduce porque los fotoasimilados se distribuyen a los nuevos sitios de demanda, en este caso a la formación de semilla.
El análisis de varianza mostró diferencias significativas (P≤0,05) por efecto de malla, fertilización y la interacción. Los cambios en irradiancia afectaron la altura de la planta, el diámetro de tallo, el área foliar, peso seco de tallo, hoja y espiga, el rendimiento de semilla y la biomasa total. La fertilización orgánica influyó en todas las variables en estudio, a excepción del número de nudos y brotes.
La interacción de las mallas de color y la fertilización modificó el diámetro de tallo, número de espigas, biomasa de tallo, hojas y total. En todas estas variables la diferencia fue altamente significativa (P≤0,05).
Efecto de la irradiancia por sombra de color en las variables agronómicas
La ALT y AF de las plantas bajo malla roja y azul fue superior al testigo en 5% y 26%, respectivamente (Tabla III). Esto reviste importancia porque la estructura del dosel influye en los procesos de fotosíntesis, transpiración, alargamiento de células, crecimiento y competencia entre especies, ya que influye en la temperatura, concentración de vapor y el régimen de radiación en el medio de la planta; (Daughtry et al., 1983; Montemayor et al., 2006). Los cambios morfológicos en chía, podrían deberse a la presencia de fitocromos y criptocromos, fotorreceptores fotomorfogénicos responsables de la transducción de señales en las plantas superiores, que absorben en las regiones rojo y rojo lejano del espectro en el caso de los fitocromos y los criptocromos con los espectros de acción en la región UV-A de la luz y luz azul, ya que estimulan eventos relacionados con los niveles hormonales como giberelinas, auxinas, etileno, ABA, etc., responsables de alteraciones en la morfología de la planta, su fisiología, desarrollo y metabolismo (Franklin y Whitelam, 2005; Möglich et al., 2010; Hu et al., 2013; Christie et al., 2015).

* Irradiancia de malla azul= 691µmol·m-2·s-1, roja= 720µmol·m-2·s-1, verde=758µmol·m-2·s-1, testigo= 1848µmol·m-2·s-1).
* En las columnas valores con letra similar son estadísticamente iguales según la prueba de Tukey (p≤0,05). DSH0,05: diferencia significativa honesta. ALT: altura, DT: diámetro de tallo, AF: área foliar, PST: peso seco de tallo, PSH: peso seco de hojas, PE: peso de espigas, RS: rendimiento de semilla y BT: biomasa total.
Con la malla verde se alcanzó mayor diámetro de tallo, peso seco de tallo y biomasa total (Tabla III). Algunos estudios mencionan que la luz azul mantiene el potencial eléctrico en las membranas de las células guarda y regula la apertura estomática, lo que provoca aumento de la concentración del CO2 dentro del mesófilo disponible para la fotosíntesis y hay mayor producción de biomasa (Kinoshita et al., 2003;Moni et al., 2014). En el presente estudio no se observó esta respuesta, ya que se redujo la biomasa con la malla azul. Resultados similares consignaron Monteiro et al. (2012) en orégano (Origanum vulgare L.) donde la biomasa de tallo y raíz se redujo bajo cobertura azul en relación a las mallas roja y negra. La producción de biomasa aumentó en plantas de brócoli expuestas a cobertura roja (Casierra y Rojas, 2009), mientras que en tomate cherry, lo hicieron con mallas de color azul y perla (Márquez et al., 2014). Estos resultados indican que los cultivos tienen respuesta diferencial al color de las mallas y que la misma está correlacionada directamente con el rendimiento (Kooman et al., 1996; Ayerza y Coates, 2009). En el presente estudio no hubo relación entre el uso de mallas de color y el incremento en el rendimiento de semilla de chía; por el contrario, las plantas bajo malla verde, aunque registraron mayor acumulación de materia seca, disminuyeron el rendimiento en 24%, lo que indica que el efecto es sobre el índice de cosecha. Por ello no se recomienda el uso de malla de color verde. El grado de sombra afecta a varios procesos fisiológicos (Casal, 2013), la menor irradiancia en la sombra limita la fotosíntesis, pero reduce también la posibilidad de daño por exceso de radiación fotosintéticamente activa (RFA) y luz ultravioleta (UVB).
Efecto de la fertilización orgánica en las variables agronómicas
La aplicación de vermicompost o la combinación de vermicompost y aplicación foliar de té, favoreció la ALT y DT, así como para la formación del dosel vegetal cuantificado mediante el AF, quedando muy por abajo las plantas testigo. En las variables de RS, PST, PSH, PE y BT, con la fertilización combinada (al suelo y vía foliar) se obtuvieron los mejores resultados (Tabla IV). Esto se atribuye a que además de los nutrimentos contenidos en el vermicompost, la planta absorbe lo aplicado al follaje, responde favorablemente a la aplicación del té (Albert et al., 2012) y se promueve el crecimiento vegetal (Tabla IV). En Argentina (Jamboonsri et al., 2012) la chía tiene bajo requerimiento de fertilizantes y riegos, y se fertiliza con un rango de 15-45kg·ha-1 de N y 37kg·ha-1 de P, mientras que en México se aplican 68kg·ha-1 de N (Ayerza y Coates, 2009), por lo que hacen falta más estudios sobre los requerimientos de fertilización del cultivo. Sin embargo, independientemente de los niveles óptimos de nutrientes, la fertilización orgánica resulta efectiva dada la importancia de producir en armonía con el ambiente, y ecológicamente sustentables.
Efecto de la interacción de la irradiancia bajo malla y la fertilización
Se observaron diferencias en DT, NE, PST, PSH y BT con respecto a la interacción de las mallas sombra de color y la fertilización en el cultivo de chía (Tabla V). En la variable DT los valores más bajos se registraron sin fertilización bajo malla roja y sin malla, mientras que los otros tratamientos fueron estadísticamente similares (P≤0,05). La fertilización, ya sea al suelo o combinada, afectó el crecimiento de la planta bajo las mallas roja y sin malla, a diferencia de las que crecieron bajo la malla verde, pues con o sin fertilización se obtuvieron resultados similares. El mayor NE se observó en las plantas con malla verde y aplicación de vermicompost más té de vermicompost y el menor resultó ser el testigo absoluto (Tabla V), lo cual pudo influir en el bajo rendimiento de semilla que se obtuvo bajo este color de malla. La combinación de la malla verde y la aplicación de vermicompost y té registró mayor PST en contraste con los tratamientos sin fertilización en todas las condiciones de malla sombra y testigo (Tabla V). Los valores más altos de PSH se obtuvieron con malla verde y la aplicación de vermicompost y té, mientras que los más bajos se observaron en las plantas cultivadas con malla roja sin fertilización (Tabla V).
La combinación de malla verde y fertilización con vermicompost y té produjo los valores más altos de BT y los más bajos en las plantas a las cuales no se les hizo aplicación de ninguna fuente de nutrimentos (Figura 2). Algunos estudios mencionan que la luz verde es esencial para la germinación, crecimiento de hojas y cotiledones, crecimiento de frutos, transpiración y conductancia estomatal, acumulación de pigmentos fotosintéticos, metabolismo de carbohidratos y lípidos (Golovatskaya y Karnachuk, 2015; Sato et al., 2015), lo cual pudo influir en las respuestas obtenidas en este estudio. La Figura 2 muestra que el color de la malla no es un factor que influye en la producción de biomasa, pero sí lo hace la fuente de nutrimentos. También se ha sugerido al fitocromo como uno de los receptores de luz verde, que está en un estado fotoestacionario. La intensidad umbral de la luz verde para la excitación del fitocromo es 2,5 veces mayor que la de luz roja, por lo que la capacidad para absorber luz verde permite a las plantas evaluar las condiciones de iluminación y de responder adecuadamente a los cambios durante el día o la temporada (Sato et al., 2015). Independientemente del color de la malla, la BT más baja se presentó en las plantas no fertilizadas, por lo cual se sugiere la utilización tanto del vermicompost como el té de vermicompost como fuentes de nutrición orgánica para el cultivo de chía.
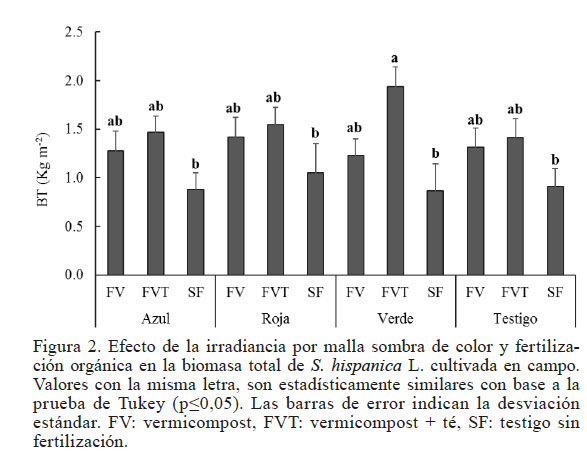
Figura 2
Efecto de la irradiancia por malla sombra de color y fertilización orgánica en la biomasa total de S. hispanica L. cultivada en campo. Valores con la misma letra, son estadísticamente similares con base a la prueba de Tukey (p≤0,05). Las barras de error indican la desviación estándar. FV: vermicompost, FVT: vermicompost + té, SF: testigo sin fertilización.
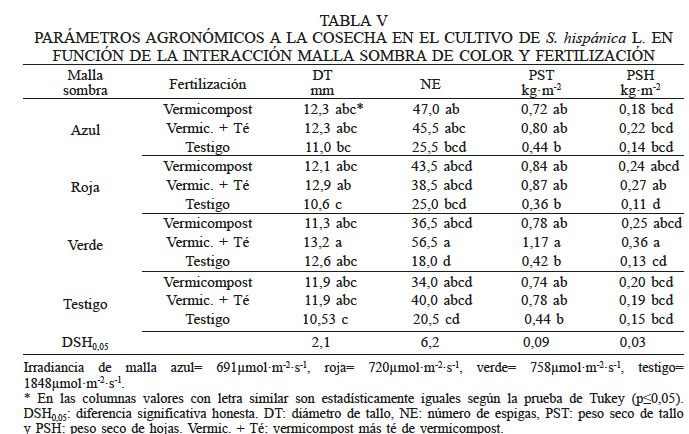
* Irradiancia de malla azul= 691µmol·m-2·s-1, roja= 720µmol·m-2·s-1, verde= 758µmol·m-2·s-1, testigo= 1848µmol·m-2·s-1. * En las columnas valores con letra similar son estadísticamente iguales según la prueba de Tukey (p≤0,05). DSH0,05: diferencia significativa honesta. DT: diámetro de tallo, NE: número de espigas, PST: peso seco de tallo y PSH: peso seco de hojas. Vermic. + Té: vermicompost más té de vermicompost.
Conclusiones
Los cambios en la irradiancia por el uso de malla de diferentes colores afectan el crecimiento de las plantas de chía. Con malla azul y roja (691 y 720µmol·m-2·s-1, respectivamente) se acortó el ciclo del cultivo, pero no se incrementó el rendimiento de semilla. La malla verde (758µmol·m-2·s-1) favoreció la tasa de crecimiento absoluto, más no la producción de semilla.
El vermicompost y aplicación foliar de té de vermicompost incrementan la biomasa y el rendimiento de semilla, por lo que es una alternativa de manejo orgánico para el cultivo de chía.
CRECIMIENTO Y PRODUCCIÓN DE CHÍA (Salvia hispanica L.) EN FUNCIÓN DE LA IRRADIANCIA Y FERTILIZACIÓN ORGÁNICA
Adhikary S (2012) Vermicompost, the story of organic gold: A review. Agric. Sci. 3: 905-917.
Albert N, Nazaire K, Hartmut K (2012) The relative effects of compost and non-aerated compost tea in reducing disease symptoms and improving tuberization of Solanum tuberosum in the field. Int. J. Agri. Res. Rev. 2: 504-512.
Arriagada GCD (2014) Efecto de la Fecha de Siembra en el Rendimiento en Grano en Chía (Salvia hispanica L.) y su Relación con el Crecimiento y Desarrollo. Tesis. Universidad de Chile. 29 pp.
Ayerza R, Coates W (2009) Influence of environment on growing period and yield, protein, oil and a-linolenic content of three chia (Salvia hispanica) selections. Ind. Crop. Prod. 30: 321-324.
Ben-Yakir, D, Antignus Y, Offir Y, Shahak Y (2012) Colored shading nets impede insect invasion and decrease the incidences of insect-transmitted viral diseases in vegetable crops. Entomol. Exp. Appl. 144: 249-257.
Brouillette LC, Donovan LA (2011) Relative growth rate and functional traits of a hybrid species reflect adaptation to a low-fertility habitat. Int. J. Plant Sci. 172: 509-520.
Busilacchi H, Quiroga M, Bueno, Di Sapio O, Flores V, Severin C (2013) Evaluación de Salvia hispanica L. cultivada en el sur de Santa Fe (República Argentina). Cult. Trop. 34: 55-59.
Casal JJ (2013) Photoreceptor signaling networks in plant responses to shade. Annu. Rev. Plant Biol. 64: 403-427.
Casierra-Posada F, Rojas JFB (2009) Efecto de la exposición del semillero a coberturas de colores sobre el desarrollo y productividad del brócoli (Brassica oleracea var. italica). Agron. Colomb. 27: 49-55.
Casierra-Posada F, Peña-Olmos JE (2015) Modificaciones fotomorfogénicas inducidas por la calidad de la luz en plantas cultivadas. Rev. Acad. Colomb. Cienc. Ex. Fis. Nat. 39: 84-92.
Christie JM, Blackwood L, Petersen J, Sullivan S (2015) Plant flavoprotein photoreceptors. Plant. Cell. Physiol. 56: 401-413.
Di Sapio O, Bueno M, Busilacchi H, Quiroga M, Severin C (2012) Caracterización morfoanatómica de hoja, tallo, fruto y semilla de Salvia hispanica L. (Lamiaceae). Bol. Latinoam. Carib. Plant. Med. Aromat. 11: 249-268.
Daughtry CS, Gallo TKP, Bauer ME (1983) Spectral estimates of solar radiation intercepted by corn canopies. Agron. J. 75: 527-531.
Escalante EJAS, Kohashi SJ (1993) El Rendimiento y Crecimiento del Frijol. Manual para la Toma de Datos. Colegio de Postgraduados. Montecillo, México. 84 p.
Fageria NK, Baligar VC (2005) Enhancing nitrogen use efficiency in crop plants. Adv. Agron. 88: 97-185.
Franklin KA, Whitelam GC (2005) Phytochromes and shade-avoidance responses in plants. Ann. Bot. 96: 169-175.
García E (2004) Modificación al Sistema de Clasificación Climática de Koppen para Adaptarlo a las Condiciones de la República Mexicana. UNAM. México. 217 pp.
Gardner FP, Pearce RB, Mitchell RL (1990) Physiology of Crop Plants. 2a ed. Iowa State University Press. Ames, IO, EEUU. 327 pp.
Golovatskaya IF, Karnachuk RA (2015) Role of green light in physiological activity of plants. Rus. J. Plant Physiol. 62: 727-740.
González-Solano KD, Rodríguez-Mendoza MN, Sánchez-Escudero J, Trejo-Téllez LI, García-Cué JL (2013) Uso de té de vermicompost en la producción de hortalizas de hoja. Agroproductividad 7: 17-22.
Hu W, Franklin KA, Sharrock RA, Jones HA, Harmer SL, Lagarias JC (2013) Unanticipated regulatory roles for Arabidopsis phytochromes revealed by null mutant analysis. Proc. Natl. Acad. Sci. 110: 1542-1547.
Hunt R (1990) Basic Growth Analysis. Unwin Hyman. Londres, RU. 132 pp.
Hunt R, Causton DR, Shipley B, Askew AP (2002) A modern tool for classical growth analysis. Ann. Bot. 90: 485-488.
Ingham E (2005) The Compost Tea Brewing Manual. 5a ed. Soil Food Web Inc. Corvallis, OR, EEUU. 79 pp.
Jamboonsri W, Phillips TD, Geneve RL, Cahill JP, Hildebrand DF (2012) Extending the range of an ancient crop, Salvia hispanica L. - a new ω3 source. Genet. Resour. Crop Evol. 59: 171-178.
Kenyangi A, Blok W (2012) Vermicompost as a component in potting mixes for growth promotion in ornamental plants. Rwanda J. 28: 53-63.
Kinoshita T, Emi T, Tominaga M, Sakamoto K, Shigenaga A, Doi M, Shimazaki K (2003) Blue light and phosphorylation dependent binding of a 14-3-3 protein to phototropins in stomatal guards cells of broad bean. Plant Physiol. 133: 1453-1463.
Kooman PL, Fahem M, Tegera P, Haverkort AJ (1996) Effects of climate on different potato genotypes. 1. Radiation interception, total and tuber dry matter production. Eur. J. Agron. 5: 193-205.
Lu N, Maruo T, Johkan M, Hohjo M, Tsukagoshi S, Ito Y, Ichimura T, Shinohara Y (2012) Effects of supplemental lighting with light-emitting diodes (LEDs) on tomato yield and quality of single-truss tomato plants grown at high planting density. Environ. Control Biol. 50: 63-74.
Márquez-Quiroz C, Robledo-Torres V, Benavides-Mendoza A, Vázquez-Badillo ME, De la Cruz-Lázaro E, Estrada-Botello MA, López-Espinosa. ST (2014) Uso de mallas sombra: una alternativa para la producción de tomate cherry. Ecosist. Rec. Agropec. 1: 175-180.
Milenković L, Ilić ZS, Durovka M, Kapoulas N, Mirecki N, Fallik E (2012) Yield and pepper quality as affected by light intensity using colour shade nets. Agric. Forest. 58: 19-33.
Möglich A, Yang X, Ayers RA, Moffat K (2010) Structure and function of plant photoreceptors. Annu. Rev. Plant Biol. 61: 21-47.
Mohd AN, Keong SY, Yong WH, Kee BB, Wei ST, Guan ST (2012) The promising future of chía, Salvia hispanica L. J. Biomed. Biotechnol. 2012: 1-9.
Moni A, Lee AY, Brigss WR, Han IS (2014) The blue light receptor Phototropin 1 suppresses lateral root growth by controlling cell elongation. Plant Biol. 17: 34-40.
Monteiro CR, Brasil JEP, Soares ER, Moreir CM (2012) Crescimento de plantas, teor e qualidade de óleo essencial de folhas de orégano sob malhas coloridas. Glob. Sci. Technol. 5: 11-22.
Montemayor TJA, Zermeño AG, Olague JR, Aldaco RN, Fortis MH, Salazar ES, Rodríguez JCR, Vázquez-Vázquez C (2006) Efecto de la densidad y estructura del dosel de maíz en la penetración de la radiación solar. Int. J. Exp. Bot. 75: 47-53.
Nassi DiN, Roncucci NN, Triana F, Tozzini C, Bonari E (2011) Productivity of giant reed (Arundo donax L.) and miscanthus (Miscanthus x giganteus Greef et Deuter) as energy crops: growth analysis. Ital. J. Agron. 6: 141-147.
Olle M, Virsile A (2013) The effect of light-emitting diode lighting on greenhouse plant growth and quality. Agr. Food Sci. 22: 223-234.
Ozdamar UH, Unlu H, Karakurt Y, Padem H (2011) Changes in fruit yield and quality in response to foliar and soil humic acid application in cucumber. Sci. Res. Ess. 6: 2800-2803.
Pant AT, Radovich JK, Hue NV, Arancon NQ (2011) Effects of vermicompost tea (aqueous extract) on pak choi yield, quality, and on soil biological properties. Compost Sci. Util. 19: 279-292.
Poorter H, Niinemets U, Poorter L, Wright IJ, Villar R (2009) Causes and consequences of variation in leaf mass per area (LMA): a meta-analysis. New Phytol. 182: 565-588.
Rodríguez LLA, Guevara FH, Gómez HC, Fonseca MF, Gómez JCC, Pinto RR (2016) Anatomía foliar relacionada con la ruta fotosintética en árboles de café (Coffea arabica L., var. Caturra Rojo) expuestos a diferentes niveles de radiación solar en la Sierra Maestra, Granma, Cuba. Acta Agron. 65: 248-254.
Romero FJC, Rodríguez MMN, Escalante EJAS, Gutiérrez CMC, Peña VCB, Cueto WJA, Burguete EH (2017) Dinamica de crecimiento de Stevia cultivada en sustratos orgánicos en invernadero. Rev. Fitotec. Mex. 40: 341-350.
SAS (2015) SAS/STAT User´s Guide. Release 9.4. SAS Institute, Inc. Cary, NC, USA. 440 pp.
Sato M, Nishiuchi T, Sakamoto T (2015) Responses to intermittent exposure to green light during the dark period in Arabidopsis thaliana and possible involvement of cryptochrome 2. Plant Biotechnol. 32: 263-266.
Sharma S, Pradhan K, Satya S, Vasudevan P (2005) Potentiality of earthworms for waste management and in other uses - A review. J. Am. Sci. 1: 4-16.
Tello HN (2014) Efecto de la Fecha de Siembra sobre el Crecimiento y Rendimiento de Chía Blanca (Salvia hispanica L.) Establecida en la Localidad de las Cruces, Provincia de San Antonio. Tesis. Universidad de Chile. 35 pp.
Vega R, Méndez TCS, Rodríguez WM (2012) Análisis del crecimiento de cinco híbridos de zanahoria (Daucus carota L.) mediante la metodología del análisis funcional. Agron. Costarric. 36: 29-46.
Villar RA, Ruiz RJ, Quero JL, Poorter H, Valladares F, Marañón T (2004) Tasas de crecimiento en especies leñosas: aspectos funcionales e implicaciones ecológicas. En Valladares F. (Ed) Ecología del Bosque Mediterráneo en un Mundo Cambiante. Ministerio de Medio Ambiente. Egraf. Madrid, España. pp: 191-227.
Notas de autor
marinie@colpos.mx