COMUNICACIONES
EFECTO A CORTO PLAZO DE LA APLICACIÓN DE COMPOST SOBRE LA DIVERSIDAD DE HONGOS DE SUELOS DE UN OLIVAR EN EL VALLE DE AZAPA, CHILE
EFEITO A CURTO PRAZO DA APLICAÇÃO DE COMPOSTO NA DIVERSIDADE DE FUNGOS DE SOLOS DE OLIVEIRA NO VALE DE AZAPA, CHILE
SHORT-TERM EFFECT OF THE APPLICATION OF COMPOST ON THE DIVERSITY OF FUNGI OF OLIVE ORCHARD SOILS IN THE AZAPA VALLEY, CHILE
EFECTO A CORTO PLAZO DE LA APLICACIÓN DE COMPOST SOBRE LA DIVERSIDAD DE HONGOS DE SUELOS DE UN OLIVAR EN EL VALLE DE AZAPA, CHILE
Interciencia, vol. 44, núm. 7, pp. 394-399, 2019
Asociación Interciencia
Recepción: 10 Diciembre 2018
Corregido: 15 Junio 2019
Aprobación: 20 Junio 2019
Financiamiento
Fuente: Proyecto UTA–Mayor 9721–18
Nº de contrato: Proyecto UTA–Mayor 9721–18
Beneficiario: Germán Sepúlveda Chavera, Juan Carvallo Ibáñez , Oscar Martínez Viveros
Resumen: Los suelos del norte de Chile son pobres en materia orgánica (MO), lo que define un espectro de hongos filamentosos cultivables reducido. En su práctica tradicional, la olivicultura aplica MO para mejorar las condiciones del suelo, estimulando el desarrollo y expresión del bioma edáfico. En este trabajo se determinó la estructura y dinámica de comunidades fúngicas cultivables presentes en el suelo de huertos de olivo del valle de Azapa. Utilizando criterios de abundancia y diversidad de hongos filamentosos cultivables se comparó dos olivares, demostrando el efecto a corto plazo de la aplicación de compost maduro. El análisis de la población fúngica evaluó las unidades formadoras de colonia (UFC) después de cinco días de incubación en PDA más cloramfenicol a 25 ±1°C. Se registró cada colonia fúngica, agrupando aquellas con el mismo crecimiento y características. A partir de cultivos monospóricos, se identificó morfológicamente las taxas aisladas. Se generaron curvas de rarefacción y se agrupó los distintos suelos antes y después de la aplicación. Se comparó la composición de las comunidades en ambos suelos utilizando diagramas de Venn. Con el índice de diversidad de Shannon–Wiener se calculó el efecto de la aplicación de compost en la diversidad. La aplicación de compost no generó cambios determinantes en la abundáncia y diversidad, pero modificó la composición favoreciendo la abundancia de hongos descomponedores y biocontroladores, lo que es resultado del incremento de MO, mejorando la sanidad y actividad biológica del suelo.
Palabras clave: Biodiversidad , Desierto de Atacama , Hongos filamentosos .
Resumo: Os solos no norte do Chile são pobres em matéria orgânica (MO), que define um espectro de fungos filamentosos cultiváveis reduzidos. Em sua prática tradicional, a olivicultura aplica MO para melhorar as condições do solo, estimulando o desenvolvimento e a expressão do bioma do solo. Neste trabalho foram determinadas a estrutura e dinâmica de comunidades fúngicas cultiváveis presentes no solo de pomares de oliveiras no vale de Azapa. Utilizando critérios de abundância e diversidade de fungos filamentosos cultiváveis, compararam-se dois pomares, demonstrando o efeito a curto prazo da aplicação do composto maduro. A análise da população fúngica avaliou as unidades de formação de colônias (UFC) após cinco dias de incubação em PDA mais cloramfenicol a 25 ±1°C. Cada colônia fúngica foi registrada, agrupando aqueles com o mesmo crescimento e características. Das culturas monospóricas, os táxons isolados foram identificados morfologicamente. Curvas de rarefação foram geradas e os diferentes solos agruparam–se antes e após a aplicação. A composição das comunidades em ambos os solos foi comparada usando diagramas de Venn. Com o índice de diversidade de Shannon–Wiener, calculou-se o efeito da aplicação do composto na diversidade. A aplicação de composto não gerou mudanças significativas na abundância e diversidade, mas modificou a composição, favorecendo a abundância de fungos de descomponedores e biocontroladores, o que é resultado do aumento de MO, melhorando a saúde e a atividade biológica do solos.
Abstract: Soils in northern Chile are poor in organic matter (OM), which defines a reduced spectrum of cultivable filamentous fungi. In its traditional practice, olive growers apply OM to improve soil conditions, stimulating the development and expression of the soil biome. In this work the structure and dynamics of cultivable fungal communities present in the soil of olive orchards in the Azapa valley were determined. Using criteria of abundance and diversity of cultivable filamentous fungi, two olive groves were compared, demonstrating the short-term effect of the application of mature compost. The analysis of the fungal population evaluated the colony forming units (CFU) after five days of incubation in PDA plus chloramphenicol at 25 ±1°C. Each fungal colony was recorded, grouping those with the same growth rate and characteristics. From monosporic cultures, isolated taxa were identified morphologically. Rarefaction curves were generated, and the different soils were grouped before and after the application of OM. The composition of the communities in both soils was compared using Venn diagrams. With the Shannon-Wiener diversity index, the effect of compost application on diversity was calculated. The application of compost did not generate significant changes in the abundance and diversity, but modified the composition favoring the abundance of decomposing fungi and biocontrollers, which is a result of the increase of OM, improving the health and biological activity of the soil.
Introducción
El valle de Azapa, ubicado en la región de Arica y Parinacota, Chile, es reconocido por sus centenarios huertos de olivo y la producción de aceitunas (Sepúlveda y Leyton, 2017). Las características edafoclimáticas y el origen aluvial de sus suelos presentan en forma natural deficiencia de materia orgánica (Torres y Acevedo, 2008), lo que promueve la aplicación de enmiendas orgánicas, principalmente estiércol fresco, como una práctica tradicional de los olivicultores (Sepúlveda y Tapia, 2018). Si bien el estiércol fresco puede aportar nutrientes y favorecer la microbiota del suelo, el grado de maduración, la fuente de estiércol y otros factores pueden afectar los resultados, pudiendo ser una práctica no siempre recomendable (Edmeandales, 2003). Debido que el compost proviene de un proceso biológico, normalmente estandarizado, su uso como enmienda orgánica asegura resultados más estables y replicables que el uso de estiércol u otra enmienda fresca en suelos y/o determinados cultivos (Cooperband, 2000). La aplicación de compost maduro ha sido probada como una práctica rentable y ecológicamente sustentable (Leite et al., 2010), que impacta positivamente en las propiedades físicoquímicas y en la actividad microbiológica del suelo (Bulluck et al., 2002).
Hongos y bacterias constituyen un componente esencial del microbioma e influyen en la fertilidad del suelo y el crecimiento de las plantas, mediante la regulación de la disponibilidad e intercambio de nutrientes (Borken et al., 2002; Govaerts et al., 2008) y en la mitigación del efecto de las enfermedades (El-Sharouny, 2015). Así, es posible mejorar el rendimiento y sanidad de los cultivos utilizando prácticas sustentables, tal como la aplicación de compost, que actúa favoreciendo la actividad de los microorganismos. Se estima que 1,5 millones de especies de hongos están presentes en los ecosistemas del planeta, pero solo 5 a 10% se han descrito formalmente (Hawksworth, 2001).
La investigación sobre la diversidad y abundancia de los hongos cultivables puede proporcionar una base para proyectar el rol ecológico de estos microorganismos en los agroecosistemas (Drinkwater et al., 1995; Girvan et al., 2004). Sin embargo, la mayoría de las publicaciones sobre la dinámica poblacional de microorganismos en la rizosfera ha descrito cambios en las comunidades bacterianas, aun cuando los hongos saprófitos son importantes habitantes de la rizosfera y el rizoplano (Boer et al., 2005).
El Desierto de Atacama es un laboratorio natural para estudiar diversidad microbiológica (Gómez-Silva et al., 2008), especialmente microorganismos extremofilos y extremotolerantes, debido a los límites químicos y físicos que limitan la vida (Bull et al., 2016). Los hongos son un grupo de microorganismos diverso y ubicuo, presente en todos los ambientes del planeta (Blackwel, 2011). Sin embargo, la diversidad y riqueza de hongos varía de acuerdo a parámetros bióticos y abióticos, dependiendo principalmente de la disponibilidad de MO y agua. Este trabajo es un primer acercamiento al conocimiento de la estructura y dinámica de comunidades fúngicas cultivables presentes en el suelo de huertos de olivo del valle de Azapa, norte de Chile. Utilizando criterios de abundancia y diversidad de hongos filamentosos cultivables en dos olivares de este valle andino, mostramos el efecto, a corto plazo, de la aplicación de compost maduro.
Materiales y Métodos
Estudio de comunidades fúngicas en suelos olivicolas de Azapa
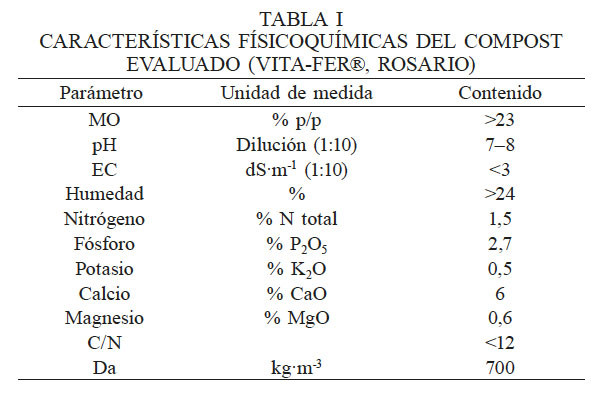
Para estudiar las comunidades fúngicas presentes en suelos de olivares del valle de Azapa y el efecto de la aplicación de compost sobre éstas, se recolectaron muestras de suelo antes (01/08/2009) y tres meses después (01/11/2009) de la aplicación de compost, en dos huertos de olivos. Los dos huertos se encuentran a lo largo de la carretera central del valle A27, ubicados en los kilómetros 11½ (VC) y 4½ (CB). Para la toma de muestras se escogieron al azar cinco olivos que se rotularon e identificaron. Del suelo de cada olivo elegido se tomaron cuatro submuestras correspondientes a cada punto cardinal de la taza de riego de cada árbol, obteniéndose un total de 20 submuestras compuestas. El procedimiento se repitió en los dos huertos. Para la extracción de la submuestra, se utilizó un barreno y se perforó la capa superficial del suelo en el borde interior de la taza de riego, a 25cm de profundidad, hasta obtener 200-250g de suelo. Tres días después del primer muestreo de suelo, se aplicó 150kg de compost (VITA–FERT®, Rosario Inc), cuyas características fisicoquímicas se muestran en la Tabla I. Esta enmienda se incorporó en ambas unidades.
En el segundo período, después de tres meses de la aplicación de compost, se procedió a una segunda colecta de muestras. Para ello, se repitió la misma operación en cada unidad muestral, utilizando los mismos puntos y procedimientos de muestreo ya descritos.
Análisis de riqueza y diversidad
El estudio de la composición y diversidad de las comunidades fúngicas se llevó a cabo en el laboratorio de Patología Vegetal de la Universidad de Tarapacá, ubicada en el Km 12 del Valle de Azapa a 250msnm, en la XV Región (Arica-Parinacota), Chile. Para el análisis de la población fúngica, se siguió el método de dilución en serie (Johnson y Curl, 1972), utilizando medio PDA complementado con cloramfenicol (Fluka Biochemicals). Se inocularon placas de Petri en una sala de cultivo estéril a 25 ±1°C. Se determinó el número de unidades formadoras de colonia (UFC) después de cinco días de incubación. Cada colonia fúngica se registró como un individuo. Se generó agrupamientos de colonias con crecimiento y características similares y se tomaron como individuos pertenecientes a un mismo agrupamiento específico. Desde éstas se generaron cultivos puros, monospóricos, y se identificó las taxas aisladas de acuerdo a caracteres morfológicos, a nivel de género y especie, estableciendo índices de abundancia de cada taxa por gramo de suelo. Se generaron curvas de rarefacción y se agrupó los distintos suelos antes y después de la aplicación de MO utilizando el programa estadístico PAST. Se comparó la composición de las comunidades en ambos suelos, antes y después de la aplicación de compost, utilizando diagramas de Venn generados on line con el programa de acceso libre depositado en http://bioinformatics.psb.ugent.be/software. Para comparar el efecto de la aplicación de compost en la diversidad, se calculó el índice H de diversidad de Shannon-Wiener (1963) según la formula H’= -1∑ [(ni/N)Ln(ni/N)], donde ni: número de colonias de una especie determinada obtenidas por gramo de suelo, y N: número total de colonias de dicha muestra. Para establecer la relación entre las variables asociadas al suelo y la estructura de las comunidades fúngicas se llevó a cabo un análisis de componentes principales (PCA) utilizando R (Torriente y Torres, 2010).
Resultados
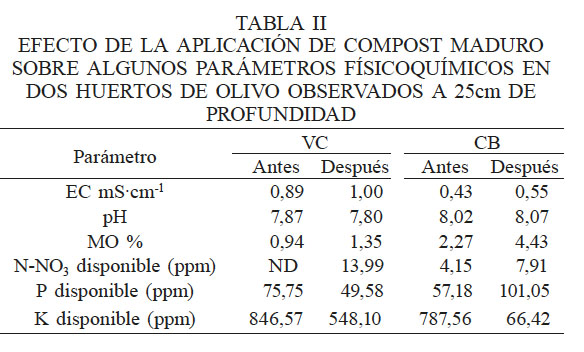
En las distintas muestras de suelo se encontró un total 40 especies de hongos filamentosos cultivables presentes en forma diferencial y pertenecientes a 11 géneros (Figura 1). La mayor abundancia de hongos se obtuvo en el suelo CB, antes y después de la aplicación de compost, lo que puede estar relacionado con una mayor cantidad de materia orgánica (MO) en este suelo. En este sentido, se puede afirmar que la aplicación de compost aumentó el porcentaje de MO y los niveles de nitratos. Los fosfatos no mostraron cambios que puedan ser asociados a la aplicación del compost; en VC tras la aplicación de compost disminuyó el fosfato y en la muestra CB aumentó. Por otro lado, el potasio disminuyó tras la aplicación de compost (Tabla II).
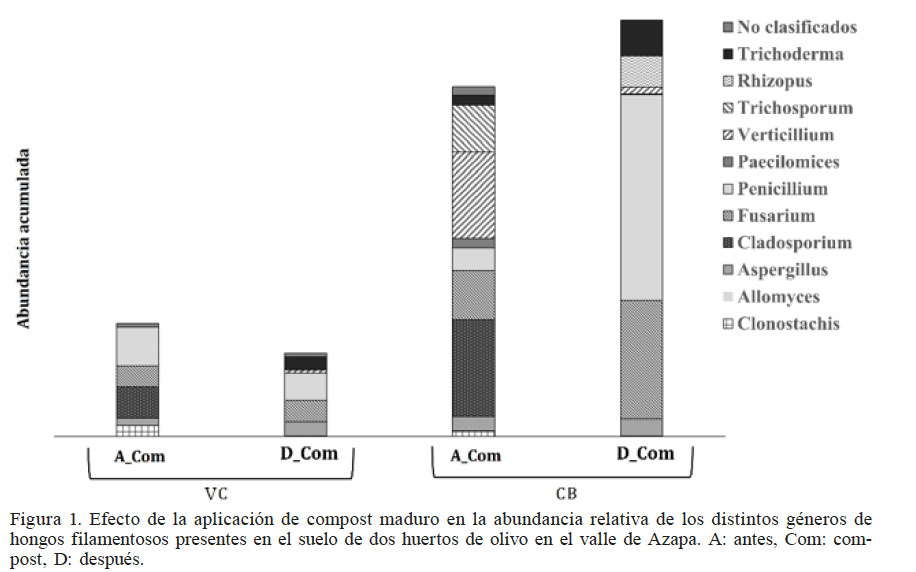
Figura 1
Efecto de la aplicación de compost maduro en la abundancia relativa de los distintos géneros de hongos filamentosos presentes en el suelo de dos huertos de olivo en el valle de Azapa. A: antes, Com: compost, D: después.
Si bien no se observaron cambios importantes en la abundancia acumulada tras la aplicación del compost, algunos géneros específicos se vieron favorecidos y otros fueron desplazados, destacando Trichoderma, que aumentó su abundancia en los dos suelos tras la aplicación de compost, mientras que por el contrario Cladosporium fue desplazado (Figura 1).
Con base en el diagrama de Venn (Figura 2) se puede afirmar que la aplicación de compost produjo importantes cambios en la composición de las comunidades en ambos suelos. En el suelo VC se desplazaron ocho especies que fueron reemplazadas por otras 11, entre las que destacan distintas especies de Aspergillus y Fusarium, además de T. harzianum y V. lecanii (Figuras 1 y 2A), aunque este último presentó baja abundancia. Lo mismo ocurrió en CB, donde ocho especies presentes antes de la aplicación de compost, fueron desplazadas por nueve especies nuevas (Figura 2B), destacando por su abundancia distintas especies de Penicillium, Aspergillus y Fusarium (Figura 1). En este caso, T. harzianum no se detectó como una especie nueva resultante de la aplicación de compost, pero su abundancia se incrementó con este tratamiento. Se puede inferir la presencia de un núcleo (core) de hongos filamentosos compuesto de siete especies (Aspergillus ochraceus, Clonostachys rosea, Fusarium oxysporum, F. solani, F. tricinctum, Penicillium aurantiogriseum y P. glabrum; (Figura 3C), destacando por su abundancia y ubicuidad Penicillium y Fusarium. Además, la aplicación de compost tuvo un efecto transformador en la composición de especies en el suelo de VC, donde se registró menor contenido de materia orgánica y nitrógeno disponible (Figuras 2A y C).
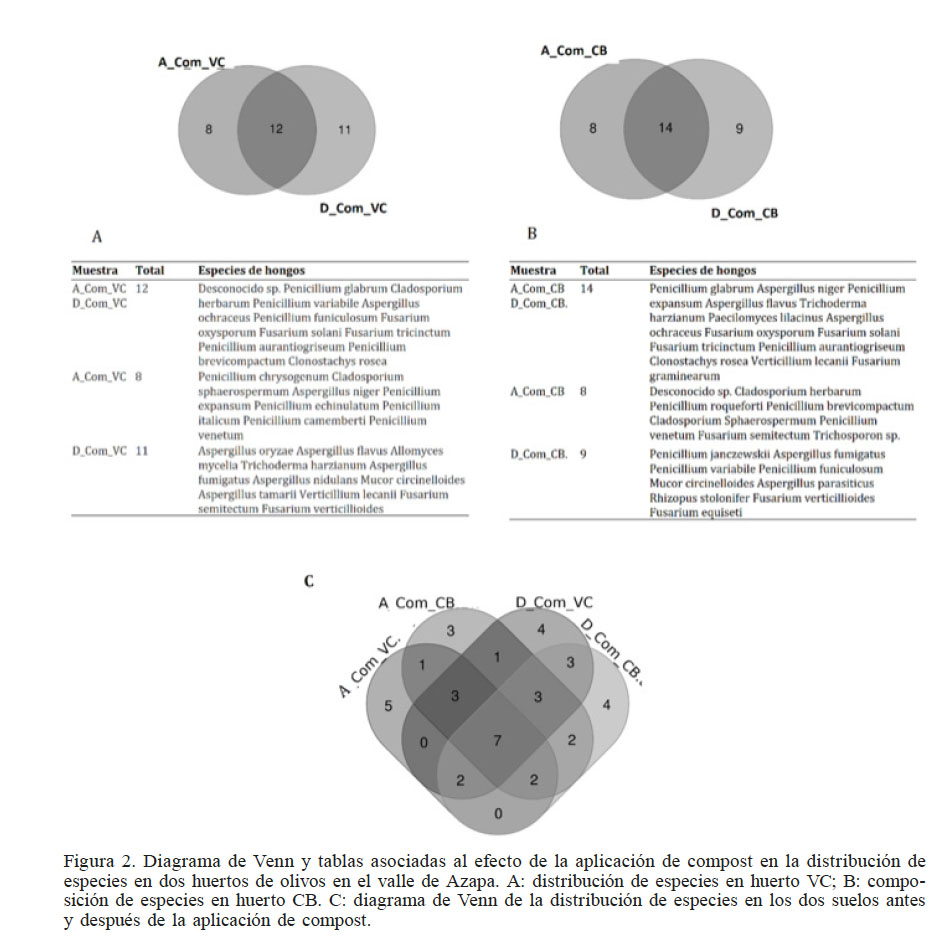
Figura 2
Diagrama de Venn y tablas asociadas al efecto de la aplicación de compost en la distribución de especies en dos huertos de olivos en el valle de Azapa. A: distribución de especies en huerto VC; B: composición de especies en huerto CB. C: diagrama de Venn de la distribución de especies en los dos suelos antes y después de la aplicación de compost
El análisis de la riqueza en los suelos antes y después de la aplicación del compost, generó curvas de rarefaccion (Figura 3A), donde se determinó una asíntota en 1500 individuos, lo que pone de manifiesto que se logró captar la diversidad en ambas condiciones de MO. La aplicación de compost en los dos suelos, aumentó la riqueza de especies, pasando en VC de 20 antes de la aplicación del compost a 23 especies tras la aplicación. Así mismo, en CB inicialmente se registró 22 especies y después de la aplicación de compost se reportaron 23. El suelo CB presentó inicialmente mayor riqueza que el VC y posterior a la aplicación de compost el suelo VC alcanzó una riqueza equivalente al suelo CB antes de la aplicación. Todo esto permite afirmar que la aplicación de compost aumenta la riqueza de especies. Sin embargo, no existe un aumento en la diversidad, como lo muestra el indice de Shanon–Weber (Figura 3B).
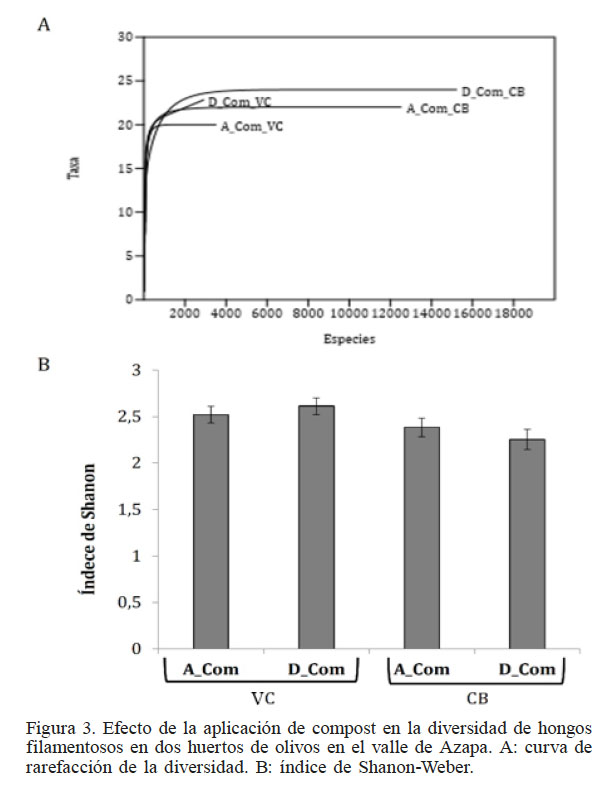
Figura 3
Efecto de la aplicación de compost en la diversidad de hongos filamentosos en dos huertos de olivos en el valle de Azapa. A: curva de rarefacción de la diversidad. B: índice de Shanon-Weber.
El análisis de componentes principales permitió agrupar y sintetizar el efecto de las distintas variables, donde los dos primeros componentes explicaron más del 90% de la varianza. Además, presenta estrecha relacion ente el pH, fosfato y MO, y estas variables tienen efecto positivo sobre el suelo CB antes y después del compost (Figura 4). Por otro lado, el suelo VC es afectado por la conductividad eléctrica (Ec) en mayor magnitud. Se puede inferir también que la abundancia (Ab) y la diversidad (Div) están negativamente correlacionadas. Así, el suelo con mayor abundancia de hongos (CB) tuvo menor diversidad. Es importante destacar que existe un marcado efecto del suelo que se mantiene también después de la aplicación del compost, lo que permite separar a los suelos en dos grupos (cluster) como revela el agrupamiento UPGMA (Figura 4B).
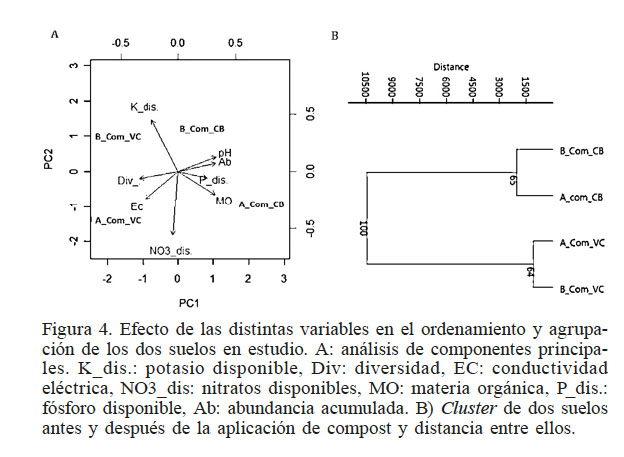
Figura 4
Efecto de las distintas variables en el ordenamiento y agrupación de los dos suelos en estudio. A: análisis de componentes principales. K_dis.: potasio disponible, Div: diversidad, EC: conductividad eléctrica, NO3_dis: nitratos disponibles, MO: materia orgánica, P_dis.: fósforo disponible, Ab: abundancia acumulada. B) Cluster de dos suelos antes y después de la aplicación de compost y distancia entre ellos.
Discusión
Utilizando técnicas basadas en la capacidad de crecimiento de las taxa en medio de cultivo, se llevó a cabo una descripción de las comunidades de hongos filamentosos presentes en olivares del valle de Azapa. En concordancia con los resultados obtenidos, Al-Sadi et al. (2015), utilizando técnicas basadas en medios de cultivo, reportaron que Eurotiomycetes y Sordariomicetes son dominantes en los suelos analizados. Cabe destacar que estudiamos hongos filamentosos cultivables que pueden representar solo una pequeña porción (1-10%) del total de hongos en la rizosfera. Sin embargo, estudios basados en técnicas de cultivo permiten una buena aproximación a la diversidad de hongos fitopatógenos, biocontroladores o descomponedores de materia orgánica (MO) presentes en mayor abundancia en la mayoría de los suelos agrícolas, tales como Ascomycetes (Eurotiomycetes: Penicillium y Aspergillus; Sordariomycetes: Fusarium (Hipocrea) (Al-Sadi et al., 2015; Kazeeroni y Al-Sadi, 2016; Yee et al., 2016; Kazerooni et al., 2017), que por su abundancia y ubicuidad son determinantes en los procesos biológicos del suelo (Nannipieri et al., 2003).
Varios estudios han demostrado que los hongos cultivables son más sensibles a los fertilizantes orgánicos que a los químicos (He et al., 2008) y potencialmente podrían ser utilizados como bioindicadores. De nuestros resultados y otros estudios (Kazerooni et al., 2017) se desprende que mayor cantidad de MO es capaz de sostener mayor abundancia de hongos. Sin embargo, encontramos cambios más importantes en la composición que en la abundancia. Sofo et al. (2014) reportaron un aumento en la población fúngica en olivares producto de la aplicación de enmiendas orgánicas. Lo anterior fue consecuencia de la aplicación de compost u otra enmienda orgánica que resulta en la incorporación de carbono orgánico al suelo (Brady y Weil, 2008). Sin embargo, en el presente estudio no se apreció un aumento importante en la diversidad, pero si en la composición de especies, en concordancia con otros estudios que reportaron el efecto de las enmiendas orgánicas en la diversidad de microorganismos (Fraç et al., 2012). Un patrón que observamos en los dos suelos es que con la aplicación de compost aumentó la presencia de Trichoderma, siendo interesante destacarlo ya que T. harzianum es biocontrolador de Verticillium dalhiae, que produce verticilosis en olivo (Carrero-Carrón et al., 2018). En este sentido, en el suelo CB se evidencia que tras la aplicación de compost aumenta la abundancia de Trichodermay se reduce significativamente la proporción de Verticillium (Figura 1). Este hecho sugiere que la sanidad en olivo puede ser mejorada tras la aplicación de compost, dada la capacidad biocontroladora de Trichoderma (Carrero-Carrón et al., 2018).
Las propiedades fisicoquímicas del suelo determinan la diversidad y abundancia de hongos en el suelo y los efectos que puede tener la aplicación de enmiendas orgánicas permite concluir que la aplicación de compost en olivares del norte de Chile, a corto plazo, no genera importantes cambios en la abundancia y diversidad pero modifica la composición favoreciendo la abundancia de hongos descomponedores y biocontroladores, tales como Trichoderma, lo que al parecer es resultado de un aumento en la MO. Esto conlleva a una mejora en la sanidad y actividad biológica del suelo. Futuros estudios deberán considerar los efectos a largo plazo de la aplicación de compost, integrando el uso de herramientas moleculares de detección y la actividad enzimática del suelo. Es probable que una combinación de enfoques dependientes e independientes de cultivo revele mayor información sobre la composición de las comunidades fúngicas y microbianas del suelo.
Agradecimientos
Los autores agradecen al Proyecto UTA–Mayor 9721–18 por el apoyo en el desarrollo de este trabajo
EFECTO A CORTO PLAZO DE LA APLICACIÓN DE COMPOST SOBRE LA DIVERSIDAD DE HONGOS DE SUELOS DE UN OLIVAR EN EL VALLE DE AZAPA, CHILE
Al-Sadi AM, Al-Mazroui SS, Phillips AJL (2015) Evaluation of culture based techniques and 454 pyrosequencing for the analysis of fungal diversity in potting media and organic fertilizers. J. Appl. Microbiol. 119: 500–509.
Blackwel M (2011) The fungi: 1, 2, 3 ... 5.1 million species? Am. J. Bot. 98:426–438
Brady NC, Weil RR (2008) Elements of the Nature and Properties of Soils. 14a ed. Prentice Hall. Upper Saddle River, NJ, EEUU. 975 pp.
Boer W, Folman L, Summerbell R, Boddy L (2005) Living in a fungal world: impact of fungi on soil bacterial niche development. FEMS Microbiol. 29: 795-811.
Borken W, Muhs A, Beese F (2002) Changes in microbial and soil properties following compost treatment of degraded temperate forest soils. Soil Biol. Biochem. 34: 403-412.
Bull AT, Asenjo JA, Goodfellow M, Gómez-Silva B (2016) The Atacama Desert: Technical resources and the growing importance of novel microbial diversity. Annu. Rev. Microbiol. 70: 215-234.
Bulluck LRIII, Brosius M, Evanylo GK, Ristaino JB (2002) Organic and synthetic fertility amendments influence soil microbial, physical and chemical properties on organic and conventional farms. Appl. Soil Ecol. 19: 147-160.
Carrero-Carrón I, Rubio M, Niño-Sánchez J, Navas-Cortés J, Jiménez-Díaz R, Hermosa R (2018) Interactions between Trichoderma harzianum and defoliating Verticillium dahliae in resistant and susceptible wild olive clones. Plant Pathol. 67: 1758-1767.
Cooperband L (2000) Composting: Art and science of organic waste conversion to a valuable soil resource. Lab. Med. 31: 283-290.
Drinkwater L, Letourneau D, Workneh F, Van Bruggen A, Shennan C (1995) Fundamental differences between conventional and organic tomato agroecosystems in California. Appl. Ecol. 5: 1098-1112.
Edmeandales D (2003) The long-term effects of manures and fertilizers on soil productivity and quality: a review. Nutr. Cycling Agroecosyst. 66: 165-180.
El-Sharouny E (2015) Effect of different soil amendments on the microbial count correlated with resistance of apple plants towards pathogenic Rhizoctonia solani AG-5. Biotechnol. Biotechnol. Equip. 29: 463-469.
Fraç M, Oszust K, Lipiec J (2012) Community level physiological profiles (CLPP), characterization and microbial activity of soil amended with dairy sewage sludge. Sensors 12: 3253-3268.
Girvan M, Bullimore J, Ball A, Pretty J, Osborn AM (2004) Responses of active bacterial and fungal communities in soils under winter wheat to different fertilizer and pesticide regimens. Appl. Environ.l Microbiol. 70: 2692-2701.
Gómez-Silva B, Rainey FA, Warren-Rhodes KA, McKay CP, Navarro-González R (2008) Atacama Desert soil microbiology. En Dion P, Nautiyal CS (Eds.) Microbiology of Extreme Soils. Springer. Berlin, Alemania. pp. 117-132.
Govaerts B, Mezzalama M, Sayre KD, Crossa J, Lichter K, Troch V, Vanherck K, De Corte P, Deckers J (2008) Long-term consequences of tillage, residue management, and crop rotation on selected soil micro-flora groups in the subtropical highlands. Appl. Soil Ecol. 38: 197-210.
Hawksworth D (2001) The magnitude of fungal diversity: the 1.5 million species estimate revisited. Mycol. Res. 105: 1422-1432.
He JZ, Zheng Y, Chen CR, He YQ, Zhang LM (2008) Microbial composition and diversity of an upland red soil under long-term fertilization treatments as revealed by culture-dependent and culture-independent approaches. J. Soils Sedim. 8: 349-358.
Johnson L, Curl A (1972) Method for the Research on Ecology of Soil Borne Plant Pathogens. Burgess. St. Paul, MN, EEUU. 247 pp.
Kazeeroni EA, Al-Sadi AM (2016) 454-pyrosequencing reveals variable fungal diversity across farming systems. Front. Plant Sci. 7: 314.
Kazerooni E, Maharachchikumbura S, Rethinasamy V, Al-Mahrouqi H, Al-Sadi M (2017) Fungal diversity in tomato rhizosphere soil under conventional and desert farming systems. Front. Microbiol. 8: 1462.
Nannipieri P, Ascher J, Ceccherini MT, Landi L, Pietramellara G, Renella G (2003) Microbial diversity and soil functions. Eur. J. Soil Sci. 54: 655-670.
Leite L, Oliveira FC, Araujo A, Galvão S, Lemos J, Silva E (2010) Soil organic carbon and biological indicators in an Acrisol under tillage systems and organic management in north–eastern Brazil. Aust. Soil Res. 48: 258-265.
Sepúlveda Chavera G, Leyton Carvajal J (2017) Olives and protected crops: spatial analysis of occupation Azapa Valley, Chile (2003–2013). Interciencia 42: 761-766.
Sepúlveda Chavera G, Tapia Contreras F (2018) Aceituna de Azapa. Ediciones Universidad de Tarapacá. Chile. 95 pp.
Shannon E, Wiener N (1963) The Mathematical Theory of Communication. University of Illinois Press. Urbana, IL, EEUU. 117 pp.
Sofo A, Ciarfaglia A, Scopa A (2014) Soil microbial diversity and activity in a Mediterranean olive orchard managed by a set of sustainable agricultural practices. Soil Use Manag. 30: 160-167.
Torres A, Acevedo E (2008) El problema de salinidad en los recursos suelo y agua que afectan el riego y cultivos en los valles de Lluta y Azapa en el Norte de Chile. Idesia 26(3): 31-44.
Torriente D, Torres V (2010) El análisis de componentes principales en la interpretación de sistemas agroecológicos para el manejo de rizobacterias promotoras del crecimiento vegetal para el cultivo de la caña de azúcar. Idesia 28(1): 23-32.
Yee TL, Tajuddin R, Nor NMIM, Mohd MH, Zakaria L (2016) Filamentous ascomycete and basidiomycete fungi from beach sand. Rendiconti Lincei 27: 603-607.
Notas de autor
gsepulve@uta.cl