Comunicaciones
DISTRIBUCIÓN DE Solanum chilense, S. peruvianum Y S. lycopersicoides EN LA CUENCA DEL RÍO LLUTA, CHILE, Y SU RELACIÓN CON FACTORES EDÁFICOS
DISTRIBUTION OF Solanum chilense, S. peruvianum AND S. lycopersicoides IN THE BASIN OF THE LLUTA RIVER, CHILE, AND ITS RELATION WITH EDAPHIC PARAMETERS
DISTRIBUIÇÃO DE Solanum chilense, S. peruvianum E S. lycopersicoides NA BACIA DO RIO LLUTA, CHILE, E SUA RELAÇÃO COM FATORES EDÁFICOS
DISTRIBUCIÓN DE Solanum chilense, S. peruvianum Y S. lycopersicoides EN LA CUENCA DEL RÍO LLUTA, CHILE, Y SU RELACIÓN CON FACTORES EDÁFICOS
Interciencia, vol. 44, núm. 10, pp. 593-599, 2019
Asociación Interciencia
Recepción: 02 Octubre 2018
Corregido: 13 Noviembre 2019
Aprobación: 14 Noviembre 2019
Financiamiento
Fuente: La presente investigación se realizó gracias al patrocinio del Centro de Investigaciones del Hombre en el Desierto (CIHDE)
Beneficiario: Vitelio Goykovic Cortés
Resumen: Se analizó el patrón de distribución de Solanum chilense, S. peruvianum y S. lycopersicoides en la cuenca del río Lluta, en relación a la conductividad eléctrica del suelo, pH, niveles de: boro, sodio, potasio, calcio, magnesio, cloruros, sulfatos, nitratos, bicarbonatos, y altitud de los sitios en que prosperan estas poblaciones. Se extrajeron y analizaron 96 muestras de suelo entre los 113 y 3580m de altitud, siempre asociadas al sitio de crecimiento de alguna planta de tomate silvestre. Los datos generados se analizaron mediante el método multivariado por análisis de componentes principales y gráficos biplot, además de estadística descriptiva uni y bivariada, y correlación de Spearman; también se espacializó la información de las variables que resultaron de la selección del análisis de componentes principales, CEs, K y pH, además de la covariable altura. El pH resultó ser indiferente al gradiente altitudinal, no así CEs y K. Respecto a la CEs en la zona baja del valle se apreciaron los mayores valores, con alta variabilidad, mientras que tuvo menor variabilidad y valores menores en los puntos muestreados en la parte alta. Para K se encontró que pocas muestras exhibieron valores altos, la mayoría resultó homogénea en ellos y con valores bajos. El crecimiento de S. peruvianum está asociado a suelos salino-bóricos por su estrecha asociación con las variables CEs, Na+, Ca2+, Mg2+, Cl-, SO4=, NO3- y B; mientras que las especies S. chilense y S. lycopersicoides están más asociadas a suelos con menos sales y boro, ubicados a mayores alturas.
Palabras clave: Boro, Distribución Geográfica, Salinidad, Tomates Silvestres.
Abstract: The distribution pattern of Solanum chilense, S. peruvianum and S. lycopersicoides in the basin of the Lluta River was analyzed in relation to the soil electric conductivity; pH; levels of boron, sodium, potassium, calcium, magnesium, chlorides, sulfite, nitrates, bicarbonates; and altitude of the sites where these populations thrive. Ninety sixsoil samples from sites between 113 and 3580m of altitude that were associated with the growing site of wild tomato plants were extracted and analyzed. The generated data was analyzed through the multivariate method by main components analysis and biplot graphics, as well as univariate and bi-variate descriptive statistics, and Spearman correlation. The information of the resulting variables from the selection analysis of the main components CEs, K and pH, and the altitude co-variate was spatialized. The pH resulted indifferent to the altitude gradient, unlike CEs and K. Regarding CEs, the highest values were found in the lower valley area, with significant variability, although it has a lower variability and values in the sample points at higher areas. For K, it was found that fewer samples exhibit higher values; most of them revealed to be homogeneous with low values. Growing of S. peruvianum is associated to boric-saline soils by its close associations to variables CEs, Na+, Ca2+, Mg2+, Cl-, SO4=, NO3- and B; while species of S. chilense and S. lycopersicoides are closely associated to soils with less salinity and boron, located at higher altitudes.
Resumo: Foi analisado o padrão de distribuição de Solanum chilense, S. peruvianum e S. lycopersicoides na bacia do rio Lluta, em relação à condutividade eléctrica do solo, pH, níveis de: boro, sódio, potássio, cálcio, magnésio, cloretos, sulfatos, nitratos, bicarbonatos, e altitude dos locais onde prosperam estas populações. Foram extraídas e analisadas 96 amostras de solo entre 113 e 3580m de altitude, sempre associadas ao local de crescimento de alguma planta de tomate silvestre. Os dados gerados foram analisados mediante o método multivariado por análise de componentes principais e gráficos biplot, além de estatística descritiva uni e bivariada, e correlação de Spearman; também foi espacializada a informação das variáveis que resultaram da seleção da análise de componentes principais, CEs, K e pH, além da covariável altura. O pH resultou ser indiferente ao gradiente altitudinal, diferente de CEs e K. Em relação a CEs na zona baixa do vale se apreciaram os maiores valores, com alta variabilidade, enquanto houve menor variabilidade e valores menores nos pontos de amostragem na parte alta. Para K foram observadas poucas amostras exibindo valores altos, a maioria resultou homogénea com valores baixos. O crescimento de S. peruvianum está associado a solos salino-bóricos por sua estreita associação com as variáveis CEs, Na+, Ca2+, Mg2+, Cl-, SO4=, NO3- e B; enquanto que as espécies S. chilense e S. lycopersicoides estão mais associadas a solos com menos sais e boro, encontrados em maiores alturas.
Introducción
La cuenca del río Lluta, ubicada en la región de Arica y Parinacota, Chile, entre los 18º y 18º30'S, y los 70º20' y 69º22''O abarca una superficie de 3.378km2. Las precipitaciones se concentran en los sectores superiores de la cordillera (4000-5000msnm) y su promedio anual aumenta gradualmente desde 0 en la parte baja del valle de Lluta, hasta 350mm en la Cordillera de los Andes (DGA, 2010). La cuenca presenta un sistema hidrogeológico que se desarrolla en la parte baja del valle de Lluta, entre la localidad de Rosario y Lluta Panamericana, el cual está constituido por dos sistemas de acuíferos con un volumen total de almacenamiento estimado en 110×106 m3 (DGA, 2010). En sus suelos crecen las especies silvestres de tomate Solanum chilense, S. peruvianum y S. lycopersicoides (Chetelat y Rick, 2004). Sin embargo, estas especies en la cuenca del río Lluta presentan un patrón de distribución altitudinal diferente a la detectada en otras cuencas del extremo norte de Chile y peruanas, donde también son endémicas.
La cuenca del río Lluta se caracteriza porque su principal cauce contiene aguas de muy mala calidad por su alto contenido de boro y sales, las que restringen fuertemente el número de especies posibles de explotar y, a la vez, los rendimientos de algunas que los agricultores cultivan en el valle de Lluta (Albornoz et al., 2007; Doussoulin y Quezada, 2008;Torres y Acevedo, 2008;Copaja y Muñoz, 2018). Esto se debe a que algunos de sus tributarios, como el río Azufre, presentan un alto contenido de boro y aporta también arsénico, azufre, hierro, manganeso y zinc (DGA, 2008), mientras que otros, como el río Colpitas, presenta una alta conductividad eléctrica (DGA, 2004) y también contribuye con boro proveniente de afloramientos hidrotermales que desembocan en él (DGA, 2008). Los elementos que contribuyen a la salinización y borificación de las aguas se incorporan a éstas por solubilización de depósitos de evaporitas, solubilización de compuestos presentes en los suelos y arrastre de sólidos por escorrentía, entre otros mecanismos. (DGA, 2004).
Respecto al clima en que se desarrollan las especies bajo estudio, S. peruvianum se localizó creciendo en los distritos climáticos 15-1 (Arica), con una altura media de 176msnm; distrito 15-2 (El Tambo), con una altura media de 607msnm y distrito 15-2-2 (María Elena) con una altura media de 1090msnm, todos pertenecientes al tipo climático desierto con influencia marina y régimen de humedad xérico (BWnXe). En el distrito 15-1 la temperatura varía entre un máximo de 27,4°C en el mes de enero y un mínimo de 12,8°C en julio, no se presentan heladas y la precipitación media anual es de 1mm. En el distrito 15-2, ubicado más al oriente, la temperatura varía entre un máximo de 29,2°C en el mes de enero y un mínimo de 12,1°C en julio, tampoco se presentan heladas y la precipitación media anual es de 1mm. En el distrito 15-2-2, situado más al oriente y a mayor altura, la temperatura varía entre los 29,7°C en el mes de enero y un mínimo de 6,8°C en julio, y la precipitación media anual es de 0mm (Santibañez et al. 2017).
Más al este, en el distrito 15-3-3, con una altura media de1998msnm, perteneciente al tipo climático desierto interior y régimen de humedad xérico (BWxe) se localizó la especie S. chilense. Este clima se caracteriza por presentar una temperatura máxima de 26,5°C en enero y una mínima de 3,4°C en julio, y la precipi- tación media anual es de 4mm. En el año se registra un promedio de 25 heladas (Santibañez et al. 2017).
Siempre al oriente, en el distrito 15-3-4 con una altura media de 2869msnm, y un clima de desierto de altura y régimen de humedad xérico (BWHXe) se detectó creciendo S. lycopersicoides junto a S. chilense. En este clima la temperatura máxima de enero es 20°C y la mínima en julio -0,1°C. En el año se registra un promedio de 157heladas (Santibañez et al. 2017).
Tomando en consideración todas las variables expuestas anteriormente, el presente estudio tuvo como objetivo determinar si la distribución de Solanum chilense, S. peruvianum y S. lycopersicoides está relacionada con el contenido de sales y boro que presentan los suelos de la cuenca u obedece a un factor altidudinal relacionado con la temperatura.
Materiales y Métodos
Considerando que las poblaciones de S. peruvianum crecen en la parte baja del valle de Lluta (entre longitudes 0,70°15.114' y 0,69°53.878') y que tanto S. chilense como S. lycopersicoides se desarrollan principalmente entre las longitudes 0,62°52.114' y 0,69°3.605' se recolectaron muestras de suelo en estas dos áreas, procurando que la distancia entre una y otra muestra, la cual siempre estuvo asociada a alguna población de tomate silvestre, fuese de ~5km.
Se recolectaron cuatro muestras del suelo de cada población de tomate; estas se extrajeron de los primeros 30cm del suelo y siempre debajo de la proyección de la copa de la planta. Se utilizó como envase para estas muestras, bolsas de polietileno provistas de etiquetas, en las cuales se registró el número de la muestra, la ubicación geográfica, nombre del sector y fecha del muestreo. El peso final de las muestras a analizar fue de ~2kg. Las coordenadas geográficas y altitud de cada población muestreada se registraron mediante un GPS de doce canales (GPS-Plus; Garmin).
Las muestras se analizaron en el laboratorio de suelos y agua de la Universidad de Tarapacá. Los parámetros químicos que se valoraron en cada muestra de suelo fueron: conductividad eléctrica del suelo (CEs), pH, boro, sodio, potasio, calcio, magnesio, cloruros, sulfatos, nitratos y bicarbonatos. Se procedió según metodología señalada por Sadzawka (2006).
Los procedimientos de análisis y unidades de medida fueron CEs (dS·m-1): extracto de saturación y determinación por conductivimetría; pH: suspensión y determinación potenciométrica; boro (mg·l-1): extracto de saturación y determinación colorimétrica con azometina-H; calcio, magnesio, potasio y sodio (meq·l-1): extracción con solución de acetato de amonio 1mol·l-1 a pH 7.0 y determinación por espectrofotometría de absorción y emisión atómica, con lantano; cloruros (meq·l-1): extracto de saturación y determinación por titulación potenciométrica con nitrato de plata; sulfatos (meq·l-1): extracción con solución de dihidrógeno fosfato de calcio 0,01mol·l-1 y determinación turbidimétrica; nitratos (meq·l-1): colorimetría con ácido nitrofenoldisulfónico; y bicarbonatos (meq·l-1): extracto de saturación y determinación por titulación potenciométrica con ácido.
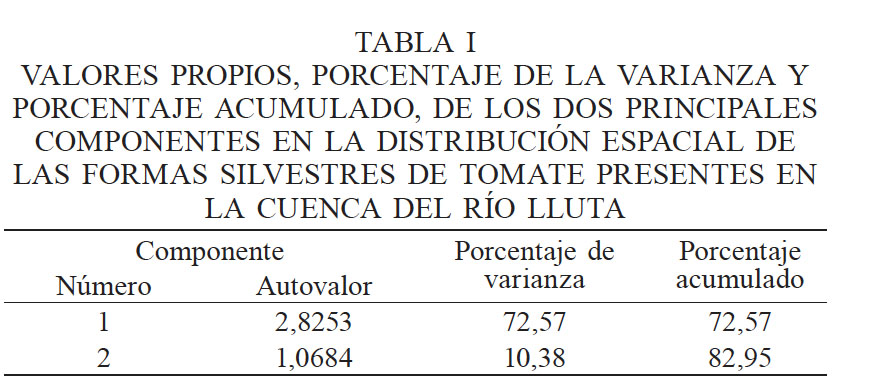
Los datos obtenidos se analizaron mediante enfoque descriptivo uni y bivariado mediante matriz de correlación con el método de Spearman, entendiendo la distribución con sesgo de la mayoría de las variables. Posteriormente se aplicó el método multivariado de análisis de componentes principales (ACP), el cual permitió reducir las variables a tres clusters: CEs e iones, K-HCO3 y pH. Las variables CEs, pH, potasio y altura se complementaron con espacialización de la información apoyándose en las coordenadas geográficas y la covariable altitud. Se excluyeron observaciones que mostraron valores extremos para algunas de las variables de interés, con el fin de suavizar y encontrar una tendencia útil para la ulterior interpretación. En todo momento se utilizó planilla de cálculo para crear máscara de ingreso y realizar ingeniería de caracteres, y posteriormente se procedió analíticamente mediante software estadístico de código abierto R y la parte espacial mediante su librería Spatial.
Resultados y Discusión
Con los valores de las 96 muestras de suelo asociadas a las especies S. chilense, S. peruvianum y S. lycopersicoides y la altitud en que se halló cada población en la cuenca del río Lluta se procedió a realizar ACP con datos estandarizados, el cual arrojó tres clusters diferenciados: CEs e iones, el binomio K-HCO3, y el pH con un comportamiento distinto al resto de las variables (Figura 1). Este enfoque multivariado convergió a que los dos primeros componentes explican el 82,95% de la variabilidad de los datos (Tabla I). El primer componente acumuló a: CEs, Na, Ca, Mg, Cl, SO4 y NO3, mientras que el segundo componente juntó a: K con HCO3 y pH, pero con signo negativo (Tabla II).
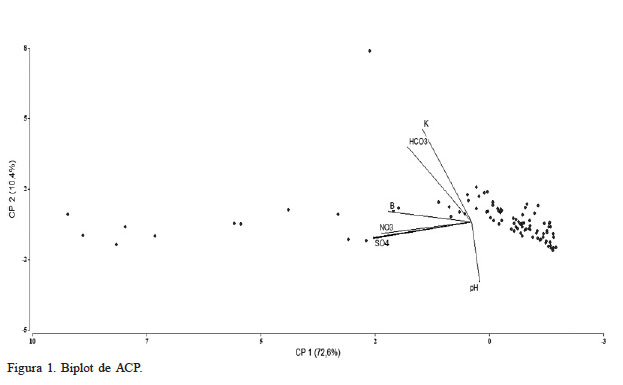
Figura 1
Biplot de ACP
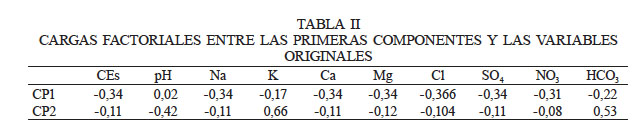
La Tabla III muestra la matriz de correlación bivariada por método de Spearman entre las variables originales, destacando altos valores absolutos en numerosos binomios, lo cual es indicio de alta correlación multivariada; es decir, hay redundancia de información. Destaca la CEs fuertemente correlacionada con todas las variables excepto el pH.
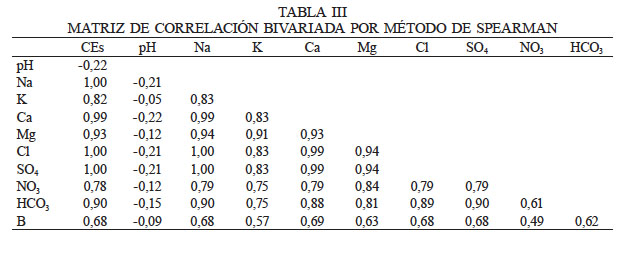
En cuanto a la alta correlación bivariada (Tabla I) entre CEs y iones, es de esperar este resultado, dado que la CEs es un indicador de la salinidad, de manera que se encuentra influenciada por la concentración y composición de las sales disueltas. Estas sales incluyen los aniones cloruro (Cl-), sulfatos (SO42-), carbonato (CO32-), bicarbonato (HCO3-) y algunas veces nitrato (NO3-), y los cationes sodio (Na+), calcio (Ca2+), magnesio (Mg2+) y en ocasiones potasio (K+) (Meenakshi 2013).
La CEs de los suelos muestreados fue variable. En el caso de las plantas de la población de S. peruvianum el 65% de las plantas se encontró creciendo en suelos con CEs entre 1,0 y 24,45dS·m-1 y elevadas concentraciones de Na+, siendo la máxima registrada de 127,14meq·l-1. La CEs promedio y desviación estándar de los suelos muestreados fue de x= 4,16 y σ= ±5,53 respectivamente. Niveles de CEs de este orden quedarían clasificados según SSSA (2008) como salinos al presentar una CE >4,0dS·m-1. Si se considera que existieron plantas prosperando en suelos con CEs= 0,22 el rango de salinidad (R) en que crece esta especie en la cuenca del río Lluta es elevado (R= 24,24), de manera que S. peruvianum presenta una gran capacidad de adaptación a ambientes con un alto o bajo contenido de sales. Teniendo en cuenta que a CEs >16dS·m-1 solo unas pocas plantas, muy tolerantes, crecen satisfactoriamente (Kotuby-Amacher et al. 2000), S. peruvianum sería una de estas. Al respecto, estudios de Chetelat et al. (2009) en tomates silvestres en el norte de Chile mostraron que muchas de las muestras de suelo donde crecían estas especies presentaban CEs >4dS·m-1 y elevados niveles de Na+, Cl- y otros iones. Cabe señalar que para el tomate cultivado, emparentado con esta especies silvestres, umbrales de salinidad del suelo >2-3dS·m-1 comienzan a afectar el rendimiento (McFarland et al. 2014).
En la especie S. chilense se detectó que solo el 12,5% de la población prospera en suelos con CEs >1,0dS·m-1; (x = 0,48; σ= ±0,45; R= 2,11). La CEs más alta registrada y asociada al crecimiento de S. chilense fue de 2,15dS·m-1 y a la vez presentaba una concentración de Na+ de 11,18meq·l-1. Al respecto Chetelat et al. (2009), en relación a esta especie estudiada en el norte del país, informaron que su adaptación a la extrema aridez y salinidad es evidente al florecer y fructificar en profusión en estas áreas. Al comparar los niveles de Na+ entre S. peruvianum y S. chilense esta última especie estaría creciendo en suelos con un 91,2% menos de este ión. Estos resultados sugieren que un factor limitante para la distribución de esta especie en los sectores más bajos de la cuenca del río Lluta sea el alto contenido de sales en estos suelos.
En S. lycopersicoides, al igual que S. chilense, no se detectó ninguna planta creciendo en los sectores más bajos de la cuenca, caracterizados por una mayor salinidad. La CEs más alta detectada y asociada a alguna planta de esta especie fue de 0,89dS·m-1; el promedio y la desviación estándar fueron x = 0,35 y σ= ±0,34 respectivamente, mientras que el rango fue bastante bajo, no alcanzando la unidad (R= 0,87). Teniendo presente estos resultados y el experimento realizado, de cultivar esta especie, al igual que S. chilense en el valle de Azapa, donde fueron capaces de crecer sin problemas al regarse con agua con una CE de 1,32dS·m-1, se puede sugerir que una posible limitante para su distribución en las partes bajas de la cuenca del río Lluta esté vinculada a la mayor salinidad de los suelos en estas áreas.
La Figura 2 da cuenta de que los mayores valores de CEs (geográfico conductividad eléctrica) se encuentran asociados a las menores alturas (geográfico altura) donde se encuentra presente la especie S. peruvianum, la cual se halló creciendo desde los 112msnm hasta una altura máxima de 1082metros, y también resultó ser la especie que se desarrolló en el suelo más salino, con CEs= 24,45dS·m-1. En cambio S. lycopersicoides se encontró desarrollándose en suelos con la menor CEs (0,02dS·m-1) y también fue la especie de tomate silvestre que se ubicó a la mayor altura, 3581m. El quiebre en la continuidad de los puntos refleja el sector en el que no se encontró creciendo ninguna especie de tomate silvestre, y correspondió al transecto entre los 1082 y 1756m.
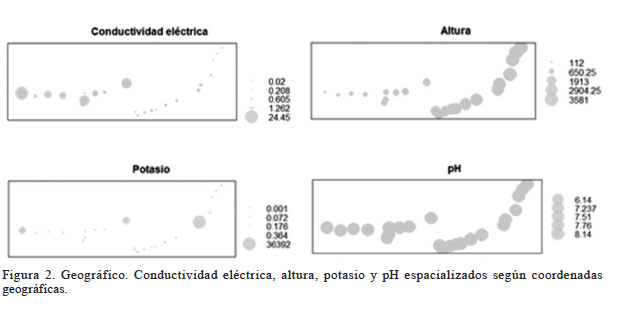
Figura 2
Geográfico. Conductividad eléctrica, altura, potasio y pH espacializados según coordenadas geográficas.
Junto con la salinidad, otro factor de estrés a considerar en la cuenca del río Lluta es el boro. Este micronutriente esencial para las plantas vasculares, cuando se encuentra en altas concentraciones en el suelo o agua, llega a ser tóxico para éstas (Polsia et al. 2016; Degryse 2017). Un 30% de la población de la especie S. peruvianum se encuentra creciendo en suelos con un alto contenido de boro, entre 12,35 y 24,75mg·l-1. El promedio de la concentración de este metaloide fue de 9,4mg·l-1 con σ= ±7,82 mientras que el rango fue amplio (R = 23,19); la concentración mínima en el suelo en que se encontró creciendo S. peruvianum fue de 1,56mg·l-1. Los resultados dan cuenta de la gran capacidad que presenta esta especie para adaptarse a ambientes tan diversos respecto al contenido de boro en el suelo. En este caso S. peruvianum no se ajustaría a lo señalado para este micronutriente, en que un rango estrecho de concentración puede significar la diferencia entre la deficiencia de la planta y la toxicidad. (Gupta et al. 2014).
No se detectó planta alguna de la especie S. chilense creciendo en suelos con concentraciones de B= 10,1mg·l-1. La concentración mínima de B en el suelo en que se observó prosperando a esta especie fue de 0,51mg·l-1, de modo que el rango fue de 9,59. El promedio alcanzó a los 5,7mg·l-1 con σ= ±3,85. Al comparar las concentraciones promedio de B en el suelo de esta especie con las de S. peruvianum, S. chilense se encuentra creciendo en suelos con una concentración 39,4% menor, lo que sugiere que la especie presenta más restricciones para habitar suelos con altas concentraciones del metaloide, y su distribución en las partes altas de la cuenca del río Lluta también puede estar condicionada por este factor.
Tampoco se registró muestra poblacional alguna de S. lycopersicoides creciendo en suelos con concentraciones de boro >4,07mg·l-1 y el valor mínimo de concentración de boro registrado fue de 0,73mg·l-1 mientras que el promedio alcanzó los 1,9mg·l-1 con σ= ±1,05 y el rango fue de 3,34. Al comparar las concentraciones de B en el suelo con S. peruvianum, S. lycopersicoides se encuentra creciendo en suelos con una concentración 79,8% menor, de forma que, al igual que S. chilense, su presencia en los pisos altitudinales de la cuenca del río Lluta se encuentra relacionada a un menor contenido de B de los suelos, situación que se puede explicar entre otros factores por el mayor nivel de precipitaciones que ocurre en la parte alta de la cuenca. De acuerdo a la DGA (2010) en Putre las precipitaciones son del orden de 197mm anuales, mientras que la parte baja del valle de Lluta (Arica) son de 1,0mm.
También destaca la correlación bivariada (Tabla I) entre el binomio HCO3 y K evidenciado en el gráfico Biplot de ACP (Figura 1), y en el plot de relación lineal (Figura 3) donde se obtuvo un coeficiente de correlación moderado de 0,63. Sin embargo, desde el punto de vista químico no existen interacciones de este carácter entre estos dos elementos. La concentración de K y HCO3 en los suelos muestreados fueron variables. De acuerdo con Crouse (2018) estas variaciones están relacionadas con los factores que intervienen en la formación de los suelos, como son el material parental, la actividad biológica, el clima, la topografía y el tiempo. El K fluctúo entre 0,01 y 1,03meq·l-1. Desde el punto de vista ecológico a este elemento se la han asociado algunos procesos fisiológicos como la resistencia al frío y la salinidad (Sardans y Peñuelas, 2015), situaciones a que están sujetas las especies analizadas. Otro estudio (Samet et al., 2015) concluyó que el suministro de K a plantas de Capsicum annuum L. atenúa parcialmente el efecto inhibitorio del exceso de B en su crecimiento. Respecto al HCO3, este varió entre 0,05 y 2,58meq·l-1. Este anión tiene un efecto fisiológico en las raíces, reduciendo la absorción de nutrientes (Morgan, 2014). A un mayor contenido de HCO3 el pH será más alto, lo que reduce la absorción del Mn, Fe, B Cu y Zn, entre otros (Pennisi y Thomas, 2015). Desde un punto de vista ecológico, especialmente en aquellas regiones áridas y semiáridas como la estudiada, el agua de riego contiene bicarbonatos disueltos los que pueden ser una fuente de emisiones de CO2 y por tanto contribuyen al aumento de los gases de efecto invernadero (Hannam et al. 2019).
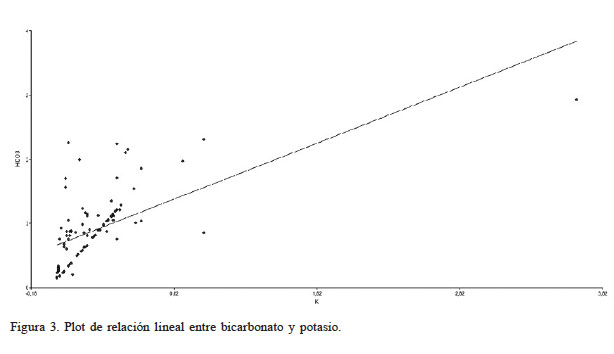
Figura 3.
Plot de relación lineal entre bicarbonato y potasio
El pH tuvo un comportamiento diferente a todas las demás variables (Figura 1); fue baja y negativa en la mayoría de los casos. En los suelos analizados varió moderadamente, entre 6,14 y 8,14 (Figura 2 - geográfico pH), valores que los tipifican como ligeramente ácidos, neutros y moderadamente alcalinos, y que corresponde a un rango donde la mayoría de los cultivos crecen bien (Horneck et al. 2011). En el transecto donde se ubicó S. peruvianum (112 y 1084msnm) el pH fue semejante al transecto donde se desarrollan S. chilense y S. lycopersicoides (1756 y 3581msnm) pues variaron entre 6,6-8,14 y 6,14-8,03 respectivamente. De acuerdo con Mc Cauley et al. (2017) valores de esta naturaleza se ajustan a la realidad de la cuenca del río Lluta, donde las precipitaciones son reducidas de manera que restringe la lixiviación de cationes formadores de bases.
Referente a las relaciones entre altura y poblaciones, las poblaciones de S. peruvianum en la cuenca del río Lluta se detectaron a alturas entre el nivel cercano al mar y los 1100msnm, resultados que se asemejan a los obtenidos por Suzuki et al. (1995), que en prospecciones efectuadas en el extremo norte de Chile, encontraron a S. peruvianum creciendo bajo los 1000msnm en áreas costeras cercanas a Arica, Paposo y Taltal, como también a colectas efectuadas por Chetelat et al. (2009) que registraron la accesión LA4318 creciendo a una elevación de 1154m. Sin embargo, otras colectas realizadas en Perú (TGRC, 2013) lo han registrado entre los 10m (accesión LA 0453) y los 2000m (accesiones LA 0453 y LA 1692). Estos resultados contribuyen a apoyar la tesis de Warnock (1991), de que existirían razas en esta especie que prosperan a bajas alturas o costeras y otras que crecen a mayores alturas o razas de montaña. De acuerdo con Cubillos (1996) en Chile probablemente solo existe la raza costera.
S. chilense se encontró en la cuenca del río Lluta creciendo a altitudes comprendidas entre los 1.000 y 3.800msnm. De acuerdo con Gonzáles (2013)S. chilense se distribuye desde el sur del Perú al norte de Chile, entre el nivel del mar y los 3000m. En prospecciones efectuadas por Susuki et al. (1995) fue detectado entre los 1000 a 3000msnm en la precordillera de Putre (Región de Arica y Parinacota) y en Calama (Región de Antofagasta). En cambio, Chetelat et al. (2009) coleccionaron esta especie en la Quebrada los Zanjones, cerca de Taltal (Región de Antofagasta), a una altura de 514m. Pliscoff et al. (2017) registraron su presencia en Alto Patache (Región de Tarapacá) entre los 800 y 850msnm. Estos resultados sugieren que la altura y por ende la temperatura no constituirían los factores determinantes en la distribución de S. chilense en la cuenca del río Lluta. Como tampoco se señala en la literatura la existencia de razas costeras y de montañas, es posible sugerir que sea la salinidad y el contenido de boro presentes en los suelos de los diferentes pisos altitudinales de la cuenca, los responsables de la distribución espacial de esta especie.
En cuanto a S. lycopersicoides, esta especie se registró creciendo entre los 2300 y 3800m de altura, de modo que existen áreas en las cuales comparte el mismo espacio que S. chilense. Sin embargo, su densidad es muy inferior, dado que en el transecto analizado su presencia fue de solo un 40% con relación a S. chilense. Prospecciones realizadas por Susuki et al. (1995) informan de su presencia en sectores aledaños al camino internacional CH-11 Arica-Tambo Quemado (Bolivia) y en la localidad de Camiña, a alturas entre los 2000 y 3700msnm. En Perú (TGRC, 2013) su presencia se evidencia entre los 2800m (accesiones LA 1966 y LA 2387) y los 3250m (accesión LA1964). También respecto a S. lycopersicoides, es posible sugerir que la salinidad y el contenido de boro en los suelos sean los responsables de su presencia solo en zonas altas de la cuenca del río Lluta, dado que en estas áreas, las precipitaciones, al ser superiores a la parte baja del valle contribuirían a lixiviar las sales y el boro del suelo. También en un pequeño ensayo de jardín de variedades, efectuado en el km 12 del valle de Azapa a 250msnm, en el cual se plantaron algunos ejemplares de estas dos especies, en un suelo con CEs: 2,34dS·m-1 y una concentración de B= 1,91mg·l-1, las cuales fueron regadas con aguas del canal Azapa (0,98mg·l-1 de B y CEw de 0,75dS·m-1) se observó que las plantas crecieron y florecieron sin ningún inconveniente.
Si se considera que S. chilense y S. lycopersicoides son capaces de prosperar a elevadas altitudes en la cuenca del río Lluta, es posible sugerir que estas poblaciones pueden constituir una fuente de germoplasma para tolerancia al frío.
En resumen, existen diferencias en el patrón de distribución de las especies silvestres de tomate. Por ejemplo, la especie S. peruvianum se encuentra habitando los sectores bajos de la cuenca del río Lluta, inferiores a los 1.100m, lo que corresponde al valle de Lluta, el cual se caracteriza por presentar concentraciones de sales y boro superiores a la que registran los suelos ubicados a mayores gradientes topográficos, en que se detectaron las especies S. chilense y S. lycopersicoides. Este hecho denota que las poblaciones de S. peruvianum han desarrollado en su proceso evolutivo la capacidad de tolerar las sales y el boro, de modo que es posible sugerir que esta especie constituye un material genético de importancia para el mejoramiento del tomate cultivado en relación con estos elementos. En cambio, el hábitat de S. chilense en la cuenca del río Lluta se extiende entre los 1748 y los 3455msnm siendo por tanto la especie de mayor distribución altitudinal. Por esta distribución también sería la especie de tomate silvestre que presenta una tolerancia intermedia a la salinidad y boro. Al no detectarse en el valle de Lluta ninguna planta de S. lycopersicoides queda de manifiesto su menor tolerancia a salinidad y boro, y también su menor grado de evolución en relación a la adaptación a condiciones salino-bóricas.
El hecho que también S. chilense y S. lycopersicoides se desarrollen sin ningún inconveniente en el valle de Azapa, caracterizado por presentar temperaturas superiores a las de los sectores altos de la cuenca y suelos con menores contenido de sales y boro que el valle de Lluta, deja en evidencia que son estos dos últimos factores los limitantes para que S. chilense y S. lycopersicoides no se encuentren ocupando los pisos altitudinales más bajos de la cuenca. Los resultados contribuyen a confirmar la hipótesis de que existen diferencias en la distribución de las especies S. chilense, S. peruvianum y S. lycopersicoides en la cuenca del río Lluta, siendo la salinidad y el boro los factores limitantes en su distribución.
Agradecimientos
La presente investigación se realizó gracias al patrocinio del Centro de Investigaciones del Hombre en el Desierto (CIHDE).
REFERENCIAS
Albornoz F, Torres A, Tapia MI, Acevedo E (2007) Cultivo de tomate (Lycopersicon esculentum mill.) hidropónico con agua desalinizada y desborificada en el valle de Lluta. Idesia 25(2): 73-80.
Copaja SV, Muñoz FJ (2018) Heavy metals concentration in sediment of Lluta river basin. J. Chil. Chem. Soc. 63: 3878-3883.
Crouse DA (2018) Soils and plant nutrients. En Moore KA, Bradley LK (Eds) North Carolina Extension Gardener Handbook. Chapter 1. NC State Extension. Raleigh, NC, EEUU. https://content.ces.ncsu.edu/extension-gardener-handbook/1-soils-and-plant-nutrients
Cubillos AP (1996) Principios para la Conservación In Situ de Parientes Silvestres de Plantas Cultivadas: El Caso de las Especies de Lycopersicon en Chile. Seminario taller: Conservación in situ de especies silvestres del género Lycopersicon. Instituto de Investigaciones Agropecuarias. Centro Regional de Investigación la Platina. Ministerio de Agricultura. Serie la Platina N° 68.
Chetelat RT, Rick CM (2004) Tomato Genetics Resource Center Department of Vegetable Crops. University of California. http://tgrc.ucdavis.edu
Chetelat RT, Pertuzé RA, Faúndez L, Graham EB, Jones CM (2009) Distribution, ecology and reproductive biology of wild tomatoes and related nightshades from the Atacama Desert region of northern Chile. Euphytica 167: 77-93.
Degryse F (2017) Boron fertilizers: use, challenges and the benefit of slow-release sources - A review. Boron 2(3): 111-122.
DGA (2004) Diagnóstico y Clasificación de los Cursos y Cuerpos de Agua según Objetivos de Calidad. Cuenca del Río Lluta. Dirección General de Aguas. Ministerio de Obras Públicas. Chile. 105 pp.
DGA (2008) Propuesta de Calidad Objetivo y Análisis General de Impacto Económico y Social-Cuenca Río Lluta. Bases Conceptuales y Cuantitativas. Dirección General de Aguas. Ministerio de Obras Públicas. Chile. 213 pp.
DGA (2010) Plan de Acción Estratégico para el Desarrollo Hídrico de la Región de Arica y Parinacota. Dirección General de Aguas. Ministerio de Obras Públicas. Chile. 89 pp.
Doussoulin EE, Quezada CL (2008) Introducción al problema de los suelos de las zonas áridas de Chile. En Quezada CL, Sandoval ME, Zagal EV (Eds.) Manejo de Suelos en Zonas Áridas. Chile. pp. 22-30
Gonzáles PA (2013) Distribución geográfica de los “tomates silvestres” (Solanum L. sect. Lycopersicon (Mill.) Wettst.: Solanaceae). Arnaldoa 20: 301-314.
Gupta U, Verma P, Solanki H (2014) Impact of high boron concentration on plants. Biolife 2: 610-614
Hannam KD, Midwood AJ, Neilsen D, Forge TA, Jones MD (2019) Bicarbonates dissolved in irrigation water contribute to soil CO2 efflux. Geoderma 337: 1097-1104
Horneck DA, Sullivan DM, Owen JS, Hart JM (2011) Soil Test Interpretation Guide. Oregon State University. EEUU. 12 pp.
Kotuby-Amacher J, Koenig R, Kitchen B (2000) Salinity and Plant Tolerance. Extension Bulletin N° HG460. Utah State University. EEUU. 8 pp.
McCauley A, Jones C, Olson-Rutz K (2017) Soil pH and Organic Matter. Nutrient management module N° 8. Montana State University. EEUU. 16 pp.
McFarland ML, Provin TL, Redmon LA, Boellstorff DE, McDonald AK, Stein LA, Wherley BG (2014) Soil and Water Salinity and Boron Thresholds for Selected Trees, Shrubs, Turfgrasses, Ornamental Ground Covers, Fruit and Nut Crops, Vegetables, Flowers, Herbaceous Crops, Field Grasses, Forages, and Field Crops. Texas A&M AgriLife Extension Service. College Station, TX, EEUU. 45 pp.
Meenakshi N, Rajni H (2013) Salt tolerance and physiological response of plants to salinity: A Review. Int. J. Sci. Eng. Res. 4(10): 44-67.
Morgan KT (2014) Understanding the potential problem with high bicarbonates in irrigation water. Southwest Florida research and education center immokalee. IFAS. University of Florida. EEUU. 18 pp. http://citrusagents.ifas.ufl.edu/events/GrowersInstitute2014/PDF/ Understanding%20the%20Potential%20Problem%20with%20High%20Bicarbonates-%20Morgan.pdf
Pennisi BV, Thomas PA (2015) Essential pH Management in Greenhouse Crops pH and Plant Nutrition. Extension Bulletin 1256. University of Georgia. EEUU. 10 pp.
Pliscoff P, Zanetta N, Hepp J, Machuca J (2017) Efectos sobre la flora y vegetación del evento de precipitación extremo de agosto 2015 en Alto Patache, Desierto de Atacama, Chile. Rev. Geogr. Norte Gde. 68: 91-103.
Polsia MP, Lupine A, Aranite F, Longo C, Mauceri A, Sunseri F, Abenavoli MR (2016) Boron Toxicity and Tolerance in Plants: Recent Advances and Future Perspectives. Dipartimento Agraria, Universita Mediterranea di Reggio Calabria. Salita Melissari, Italia. pp 115-147.
Sadzawka AR, Carrasco MA, Grez RZ, Mora MDLL, Flores HP, Neaman A (2006) Métodos Recomendados para los Suelos de Chile. Rev. 2006. Serie Actas INIA N° 34. Instituto de Investigaciones Agropecuarias. Ministerio de Agricultura. Chile. 164 pp.
Samet H, Cikili Y, Dursun S (2015) The role of potassium in alleviating boron toxicity and combined effects on nutrient contents in pepper (Capsicum annuum L.) Bulgar. J. Agric. Sci. 21: 64-70.
Santibañez FQ, Santibañez PV, Caroca CT, González PM (2017) Atlas Agroclimático de Chile. Estado Actual y Tendencias del Clima. Tomo I: Regiones de Arica y Parinacota, Tarapacá y Antofagasta. Universidad de Chile. Fundación para Innovación Agraria. 148 pp.
Sardans J, Peñuelas J (2015) Potassium: a neglected nutrient in global change. Global Ecol. Biogeogr. 24: 261-274.
SSSA (2008) Glossary of Soil Science Terms. Soil Science Society of America. EEUU. 88 pp. https://www.soils.org/publications/soils-glossary
Suzuki S, Okawara Y, Sato T, Rojas L, Matus I, Pezoa A (1995) Informe de Colecta. Especies Género Lycopersicon. Proyecto de recursos genéticos INIA-JICA. INIA Intihuasi. Chile. 18 pp.
TGRC (2013) Geographics Data on Wild Species Accessions. Tomato Genetics Resource Center. University of California, Davis. EEUU. https://tgrc.ucdavis.edu/Data/Acc/Wildspecies.aspx
Torres AH, Acevedo EH (2008) El problema de salinidad en los recursos suelo y agua que afectan el riego y cultivos en los valles de Lluta y Azapa en el norte de Chile. Idesia 26(3): 31-44.
Warnock SJ (1991) Natural habitats of Lycopersicon species. HortScience 26: 466-471.
Notas de autor
vgoykovi@uta.cl