Comunicaciones
Recepción: 25 Enero 2019
Corregido: 19 Enero 2021
Aprobación: 22 Enero 2021
Resumen:
El objetivo de la investigación fue estudiar la respuesta fisiológica de tres variedades de caña de azúcar y establecer diferencias en base a su capacidad fotosintética en respuesta a dos épocas del año y el efecto en el rendimiento y la calidad industrial. Se evaluaron tres variedades comerciales de caña de azúcar bajo condiciones de secano en dos épocas del año y diferentes horas del día, la tasa fotosintética y la respuesta varietal en variables como rendimiento y calidad del jugo (°Brix). La variedad Mex 79-431 tuvo la mayor tasa fotosintética (46,39μmolCO2·m-2·s-1) durante el verano y fue la variedad más eficiente en el aprovechamiento de la radiación fotosintéticamente activa (RFA), comparada con CP 72-2086 (45,51μmolCO2·m-2·s-1) y Mex 68 P-23 (37,34μmolCO2·m-2·s-1). Las variedades CP 72-2086 y Mex 79-431 se adaptaron mejor a las condiciones ambientales de la época del año ya que fueron superiores a Mex 68 P-23 en la tasa fotosintética tanto en verano como en invierno, con promedios de 13,5; 16,0 y 16,9μmolCO2·m-2·s-1 respectivamente. La variedad Mex 68 P-23 fue superior en rendimiento, pero con calidad del jugo similar en las tres variedades (23°Brix). Se concluye que el ambiente influye en la respuesta fotosintética de las plantas de caña de azúcar ya que en invierno se registraron reducciones de hasta 54% con respecto a verano.
PHOTOSYNTHESIS RATE OF SUGARCANE GENOTYPES UNDER RAINFED CONDITIONS DURING TWO SEASONS
Palabras clave: Calor, Estrés Hídrico, Fotosíntesis, Saccharum officinarum.
Abstract:
The goal of the research was to study the physiological response of three varieties of sugarcane and establish differences based on their photosynthetic capacity in response to two seasons and the effect in yield and industrial quality. Three commercial varieties of sugarcane were evaluated under rainfed conditions in two seasons of the year and at different times of the day, considering the photosynthetic rate and the varietal response in variables such as yield and juice quality (°Brix). The Mex 79-431 variety had the highest photosynthetic rate (46.39μmolCO2·m-2·s-1) during the summer and was the most efficient variety in the use of photosynthetically active radiation (RFA), compared to CP 72-2086 (45.51μmolCO.·m-2·s-1) and Mex 68 P-23 (37.34μmolCO2·m-2·s-1). The varieties CP 72-2086 and Mex 79-431 were better adapted to the environmental conditions of the time of the year since they were superior to Mex 68 P-23 in the photosynthetic rate in both summer and winter, with averages of 13.5, 16.0 and 16.9μmolCO2·m-2·s-1, respectively. The Mex 68 P-23 variety was superior in yield but with a similar juice quality in the three varieties (23°Brix). It is concluded that the environment influences the photosynthetic response of sugarcane plants since in winter there were reductions of up to 54% with respect to summer.
TAXA FOTOSINÉTICA DE GENÓTIPOS DE CANA-DE-AÇÚCAR EM CONDIÇÕES DE SECO EM DUAS EPOCAS DO ANO
Resumo: O objetivo da pesquisa foi estudar a resposta fisiológica de três genótipos de cana-de-açúcar e estabelecer diferenças a partir de sua capacidade fotossintética em resposta a duas estações do ano e sua resposta em produtividade e qualidade industrial. Foram avaliados três genótipos comerciais de cana-de-açúcar em condições de sequeiro em duas estações do ano e em diferentes momentos do dia, a taxa fotossintética e a resposta varietal em variáveis como rendimento e qualidade do suco (°Brix). O varietal Mex 79-431 apresentaram a maior taxa fotossintética (46,39μmolCO2·m-2·s-1) durante o verão e a utilização mais eficiente da variedade na radiação fotossinteticamente ativa (PAR) em comparação com CP 72-2086 (45,51μmolCO2·m-2·s-1) e Mex 68 P-23 (37,34μmolCO2·m-2·s-1). Os genótipos CP Mex 72-2086 e Mex 79-431 resultaram melhor adaptados para as condições ambientais da estação como eles foram superiores Mex 68 P-23 na taxa fotossintética no verão e no inverno médias de 13,5; 16,0 e 16,9μmolCO2·m-2·s-1, respectivamente. Mex 68 P-23 foi superior em rendimento, mas com qualidade de suco muito semelhante nos três genótipos (23°Brix). Conclui-se que o ambiente influencia a resposta fotossintética das plantas de cana-de-açúcar pois no inverno houve reduções de até 54% em relação ao verão.
Introducción
La caña de azúcar (Saccharum officinarum L.) es un cultivo importante para la producción de azúcar, y uno de los convertidores cuánticos más eficientes, dotado de gran capacidad de macollaje y rebrote, pero también cada vez alcanza más importancia como cultivo bioenergético debido a su fenomenal capacidad de producción de biomasa (Shrivastava et al., 2015).
En México la caña de azúcar se cultiva en casi 829.000ha, produciendo 56,7x106t con una productividad promedio de 74,4Mg·ha-1 (SIAP, 2015); sin embargo, casi el 58% se cultiva bajo condiciones de secano, en las que la precipitación es insuficiente. A pesar de esta superficie plantada en esas condiciones, ha habido poco énfasis en la investigación y el desarrollo sobre la tolerancia de su cultivo al estrés hídrico y al calor. En regiones como el Noreste de México (Tamaulipas), donde la producción de caña de azúcar es en gran medida dependiente de la precipitación, las pérdidas de rendimiento debido a la sequía y/o al calor son comunes.
El verano, particularmente en el periodo llamado ‘canícula’, suele estar acompañado de estrés por agua y/o calor, con temperaturas que exceden el rango óptimo para el crecimiento y las funciones fisiológicas de muchas plantas y en algunos casos alcanzando niveles dañinos. Esta variabilidad en el clima, junto con la aparición de episodios cortos de altas temperaturas, será una de las características importantes de los climas futuros (Djanaguiraman et al., 2011), ya que los modelos climáticos predicen que las temperaturas de la superficie terrestre seguirán aumentando (IPCC, 2007).
El aumento de la temperatura ambiente es un factor crítico para el crecimiento y desarrollo de las plantas (Wahid et al., 2007). La temperatura tiene efectos significativos sobre la fotosíntesis; en plantas que tienen una temperatura óptima, por encima y por debajo de esta, la fotosíntesis puede llegar a ser inhibida casi en su totalidad (Taiz y Zeiger, 2002) antes de que otras funciones celulares se vean afectadas (Djanaguiraman et al., 2011). En las plantas cultivadas, la fotosíntesis frecuentemente es limitada por la luz en condiciones de campo; tal limitación es el resultado de la variación de la intensidad de la luz durante la época del año y el día (Marchiori et al., 2014) y del estrés abiótico por la disponibilidad del agua (Zhao et al., 2013) y/o el estrés por alta temperatura (Wahid et al., 2007).
El efecto de la temperatura alta sobre el metabolismo de la planta depende de la intensidad y la duración de las temperaturas supra-óptimas en combinación con la tasa de incremento de la temperatura (Wahid et al., 2007).
El objetivo de esta investigación fue estudiar la respuesta fisiológica de tres variedades de caña de azúcar y establecer diferencias varietales en base a su capacidad fotosintética en respuesta a dos condiciones ambientales durante el año y su respuesta en el rendimiento y la calidad industrial asumiendo estrés por calor y/o hídrico.
Materiales y Métodos
Se realizó un experimento de campo en el sur del estado de Tamaulipas, México, en el municipio de Cd. Ocampo (22º51'N, 99º20'O, 325m de altitud) durante las zafras 2012-2013 y 2013-2014. Este lugar tiene un clima de (A)Ca(m)(w)(e)gw'' (clima semi-cálido subtropical), según el Sistema de Köppen (García, 1988). El ciclo de la caña de azúcar en esta región comienza con la siembra a principios del otoño (septiembre), el crecimiento vegetativo tiene lugar durante los meses siguientes hasta el final de la primavera (junio), y el inicio de la cosecha se produce generalmente en noviembre-diciembre.
Se estudiaron tres variedades comerciales de caña: Mex 68 P-23, CP72-2086 y Mex 79-431. Estas variedades se seleccionaron por su alto rendimiento en sacarosa, pero ellas difieren en relación al período de maduración, amplio rango de características de crecimiento fenotípico y debido a la importancia en superficie sembrada en la región sur de Tamaulipas, bajo condiciones de secano. CP 72-2086 es una variedad de madurez temprana y Mex 79-431 es una variedad de madurez tardía. CP 72-2086 y Mex 68 P-23 tienen un buen amacollamiento (Flores, 2001). La fertilización se realizó a la siembra siguiendo las recomendaciones locales basadas en las pruebas de suelo realizadas por el ingenio azucarero. Las malezas se controlaron con herbicidas (ametrina + 2,4-D en dosis de 4lxha-1) durante el experimento con el fin de minimizar el estrés biótico por la competencia de malezas. Las enfermedades y los insectos no fueron controlados porque las poblaciones estaban por debajo de los umbrales económicos. Todas las otras prácticas de manejo del cultivo fueron similares a los predios comerciales en la zona. La siembra se realizó utilizando semilla ciclo planta de cada cultivar en surcos a 1,30m de separación el 29/11/2012.
Se midió el intercambio de gases con un analizador de gas infrarrojo (LI-6400, Li-Cor, Lincoln, NE, EEUU), con fuente de luz Red Blue (6400-02B, Li-Cor). En cada parcela se midió la tasa de asimilación neta de CO2 (A), la conductancia estomática (g.), la transpiración (Tr) y la temperatura de la hoja (Th). Las mediciones de intercambio de gases se llevaron a cabo a lo largo del período de luz diurna a intervalos de dos horas entre las 7:00 y las 19:00, en la segunda hoja madura más joven de cada planta por parcela. El intercambio de gases en la hoja se midió en dos épocas contrastantes del año: verano (16/08/2013) e invierno (01/02/2014). Al medir la tasa fotosintética, la radiación fotosintéticamente activa en la cámara de la hoja se estableció como un día soleado normal al momento de la lectura; la humedad relativa se ajustó a un nivel cercano al ambiente (50%); la concentración de CO2 de la cámara se fijó en 400μml·l-1; el flujo de aire se fijó en 400mol·s-1 y la temperatura en la cámara no fue controlada, ya que se consideró debía ser similar a la temperatura del aire. Además de las variables de intercambio de gases a la cosecha se midieron variables tales como calidad de la caña basada en determinaciones de °Brix con un refractómetro digital (Model 300051, Thomas Scientific) y el rendimiento de campo (Mg·ha-1), para lo cual los tallos de la parcela útil se pesaron en una balanza de reloj de capacidad de 100kg y se transformaron a Mg·ha-1.
Se registraron precipitaciones diarias (mm), temperaturas máximas (Tmax) y mínimas (Tmin) durante el experimento con la estación meteorológica Hobo cada hora. El diseño experimental fue de bloques completos al azar con cuatro repeticiones. Cada repetición estuvo formado por cuatro surcos de 10m de longitud y la parcela útil de dos surcos centrales en una dirección N-S. En todas las variables se aplicó un análisis de varianza (SAS, 2003); la prueba de separación de medias de Tukey (P=0,05) se aplicó para establecer diferencias estadísticas.
Resultados y Discusión
Se encontraron diferencias contrastantes entre las épocas del año en las que se realizaron las lecturas en términos de temperatura del aire y precipitación (Figura 1). Se observan períodos prolongados de poca disponibilidad de agua fuera del período de lluvia (julio a septiembre) y temperaturas máximas por arriba de 35°C durante ocho meses. La Tmax fue de 36,5 y 24,4°C para verano e invierno respectivamente, es decir una diferencia de 12°C.
Se midieron variables de intercambio de gases en las variedades de interés. De acuerdo con el análisis de varianza (Tabla I), la época del año (ambiente) afectó de manera significativa (p˂0,001) el comportamiento de los genotipos involucrados, así como la hora del día y sus interacciones en todas las variables estudiadas, excepto para Genotipo x Ambiente en las variables temperatura de la hoja (Th) y transpiración (Tr). Esto indica que para la tasa fotosintética (A) las variedades estudiadas responden de manera diferente a las condiciones del ambiente y durante la hora del día, lo cual concuerda con lo señalado por Rea y de Sousa-Vieira (2001).
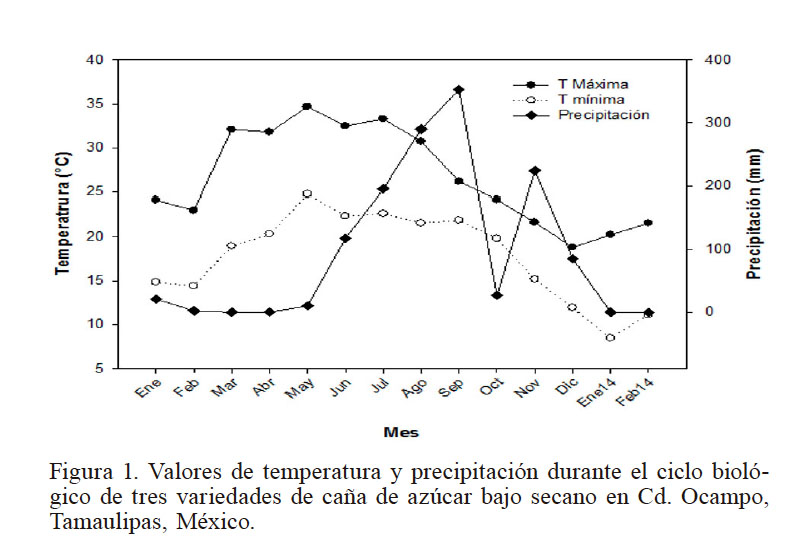
Figura 1
Valores de temperatura y precipitación durante el ciclo biológico de tres variedades de caña de azúcar bajo secano en Cd. Ocampo, Tamaulipas, México
CUADRADOS MEDIOS PARA LAS VARIABLES ESTUDIADAS DE TRES VARIEDADES COMERCIALES DE CAÑA DE AZÚCAR MEDIDAS EN DOS AMBIENTES EN CD. OCAMPO, TAMAULIPAS, MÉXICO
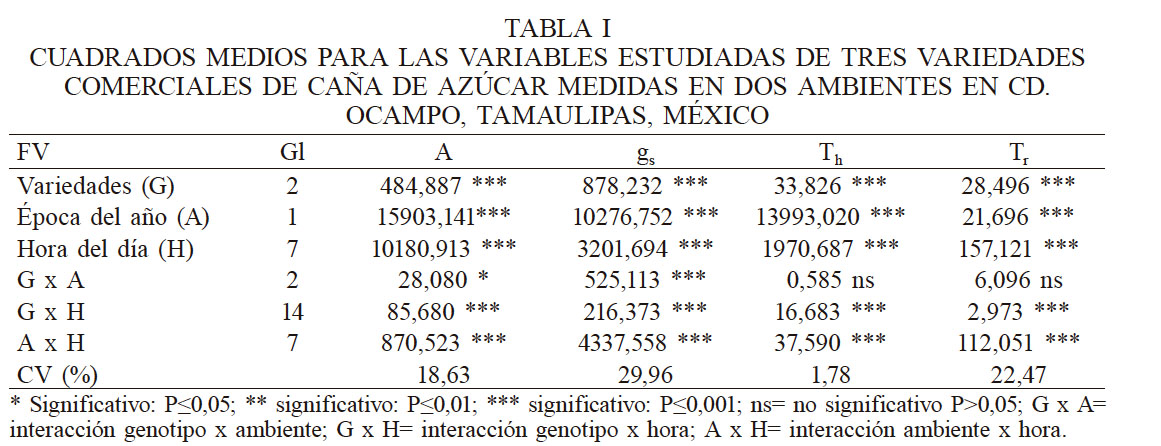
* * Significativo: P≤0,05; ** significativo: P≤0,01; *** significativo: P≤0,001; ns= no significativo P>0,05; G x A= interacción genotipo x ambiente; G x H= interacción genotipo x hora; A x H= interacción ambiente x hora.
El ambiente influyó en el comportamiento de la caña de azúcar; las variables de intercambio de gases cambian de un ambiente a otro durante el día y el año y depende de la variedad. En la Tabla II se observa que para el invierno la A se redujo significativamente (54%) en relación al verano, debido principalmente a la cantidad de radiación fotosintéticamente activa (RFA) interceptada por las hojas (Monteith y Unsworth, 1990), lo cual confirma que el primer efecto de una baja disponibilidad de la radiación visible es una reducción de la fotosíntesis y el aporte de fotoasimilados (Hashemi-Dezfouli y Herbert, 1992) y una reducción para las variables gs, Th y Tr (170, 33 y 14% respectivamente). Estos resultados permiten deducir que el ambiente está directamente relacionado con la respuesta de las variedades estudiadas, principalmente por el efecto de la temperatura, como lo demostraron Kubien et al. (2003).
PROMEDIOS PARA AMBIENTES EN LAS VARIABLES ESTUDIADAS BAJO TEMPORAL EN CD. OCAMPO, TAMAULIPAS, MÉXICO
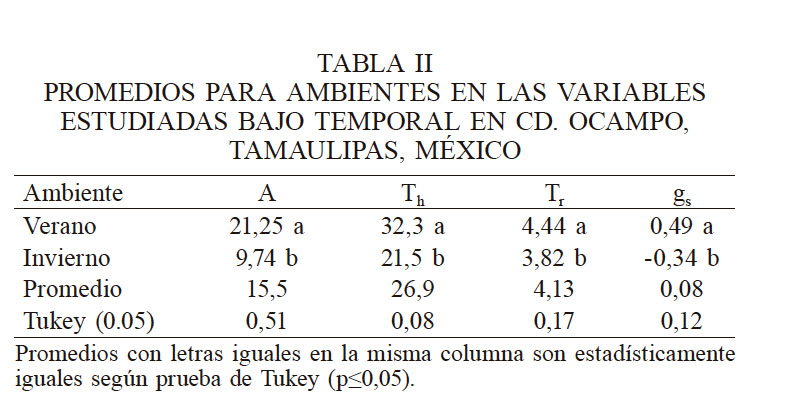
Promedios con letras iguales en la misma columna son estadísticamente iguales según prueba de Tukey (p≤0,05). Promedios con letras iguales en la misma columna son estadísticamente iguales según prueba de Tukey (p≤0,05)
Por otra parte, la temperatura durante el día es un factor del ambiente muy cambiante en función de la época del año (Pearcy et al. (2004). Bajo estas circunstancias, en este estudio las variedades tuvieron un comportamiento estadísticamente diferente (P<0,05) durante el día. En la Figura 2a se observa que entre las 11:00 y las 15:00 se tuvieron los valores más altos de la A (30,4 y 278μmolCO2·m-2·s-1 respectivamente). Esta misma respuesta se vio reflejada en las variables Th y gs.
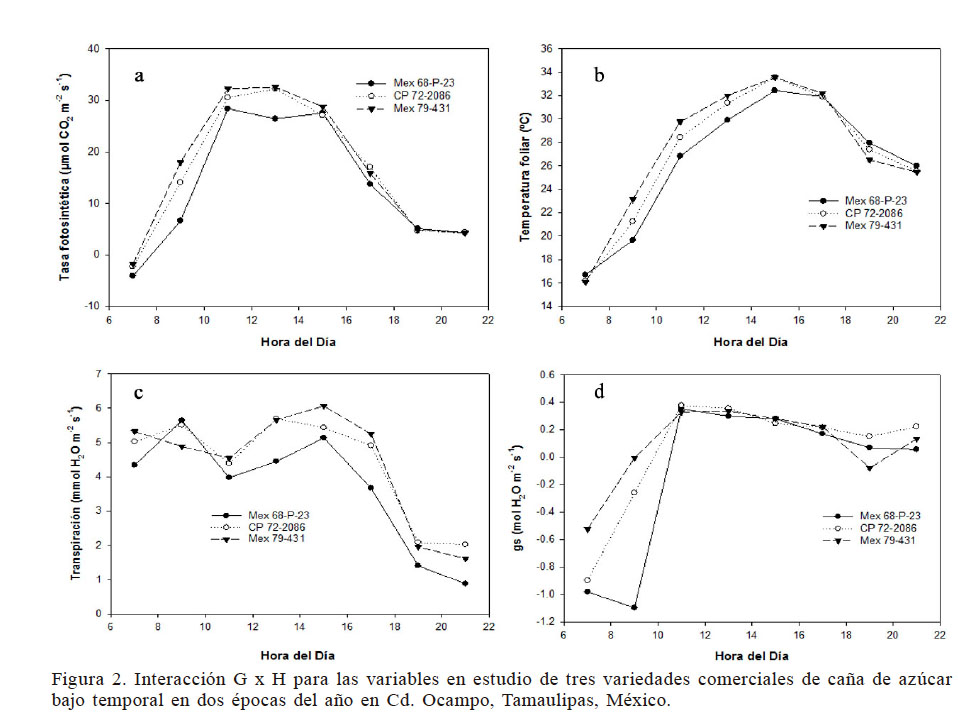
Figura 2
Interacción G x H para las variables en estudio de tres variedades comerciales de caña de azúcar bajo temporal en dos épocas del año en Cd. Ocampo, Tamaulipas, México
Además, en la Figura 2a se muestra la A de las variedades estudiadas durante el día, se observa una inestabilidad entre las 13:00 y 15:00, principalmente de la variedad Mex 68 P-23. Las variedades Mex 79-431 y CP 72-2086 mantuvieron un comportamiento muy similar entre sí durante todo el día y la A más alta y estable hacia el mediodía con promedios de 32.22 y 32,57µmolCO2·m-2·s-1 respectivamente. En las Figuras 2b, c y d se muestran las variables Th, Tr y gs respectivamente; en cada una de estas variables se observa un comportamiento de las variedades con la misma tendencia que con la A durante el día, pero con cierto nivel de estabilidad como resultado de la influencia de la temperatura, aunque respondiendo de manera diferente, dependiendo esto de la hora del día y de la variedad. De manera general las variedades Mex 79-431 y CP 72-2086 responden de manera estable durante el día, pero no así la variedad Mex 68 P-23; esta respuesta se asocia principalmente a que la RFA capturada por las plantas no es la misma en el transcurso de las horas del día (Marchiori et al., 2014), respondiendo las plantas con los valores más altos de fotosíntesis al medio día ya que la radiación es mayor que en el resto del día.
Estos resultados confirman la necesidad de identificar variedades de caña sobresalientes para este tipo de condiciones con altos riesgos de estrés abiótico (hídrico y/o calor). Una variedad adecuada para estas condiciones es un genotipo estable; es decir, aquel que tenga un rendimiento alto y se comporte consistentemente en diferentes ambientes (Rea y de Sousa-Vieira, 2001).
En general, para la A (Figura 3a) se observó que las tres variedades en estudio tuvieron en el ambiente de verano un comportamiento superior (P<0,05) al de invierno; dicha superioridad dependió en gran medida de la variedad, ya que existieron diferencias de hasta 55,8%. La variedad Mex 79-431 tuvo en promedio la mayor tasa fotosintética durante el verano (23,036µmolCO2·m-2·s-1); sin embargo, durante el invierno tuvo un comportamiento similar al de la variedad CP 72-2086, pero diferente a Mex 68 P-23. La disminución de la tasa fotosintética en invierno es atribuible principalmente a la reducción de la temperatura del aire (Figura 1), ya que es severamente reducida en condiciones en las que la temperatura es más baja que el óptimo (Inman-Bamber et al., 2010). En la Figura 3d se observa que para la g. en el ambiente de verano las variedades Mex 79-431 y CP 72-2086 tuvieron un comportamiento similar y superior a Mex 68 P-23; para invierno las variedades tuvieron un comportamiento similar al verano, en promedio la g. fue similar para las variedades Mex 79-431 y CP 72-2086 aunque con valores negativos, siendo la más afectada la variedad Mex 68 P-23. Ello indica que los estomas estuvieron cerrados, principalmente durante las primeras horas del día, lo cual se vio reflejado en una A muy baja, comportamiento encontrado también por Brugnoli y Lauteri (1991). Para la T. (Figura 3b) se observa que las variedades CP 72-2086 y Mex 79-431 tuvieron un comportamiento similar en las dos épocas del año, con una T. más alta durante el verano; en ambos ambientes la variedad Mex 68 P-23 tuvo la menor Th. Esto demuestra que las variedades CP 72-2086 y Mex 79-431 se adaptan mejor a las condiciones ambientales de la zona ya que tuvieron un mejor comportamiento comparado con Mex 68 P-23 en la A tanto en verano como en invierno. Esta respuesta entre genotipos se atribuye principalmente a las características genéticas propias de cada variedad, las que responden de manera diferente al ambiente.
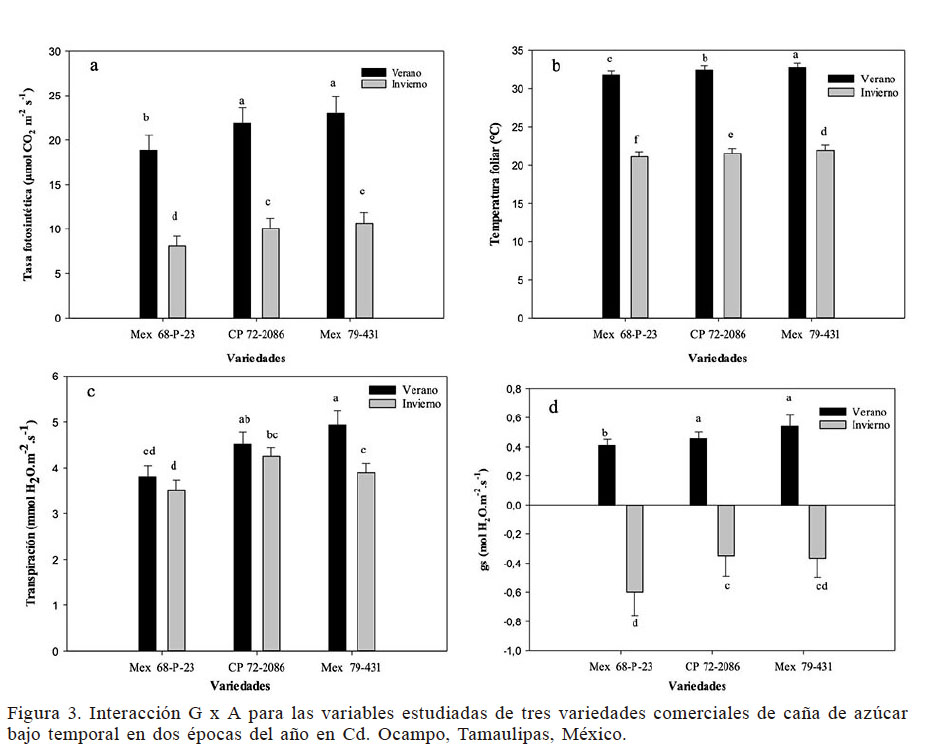
Figura 3
Interacción G x A para las variables estudiadas de tres variedades comerciales de caña de azúcar bajo temporal en dos épocas del año en Cd. Ocampo, Tamaulipas, México
La variedad Mex 79-431 tuvo en promedio la A más alta (16,9µmolCO2·m-2s·-1) con respecto a las otras variedades (Tabla III). Superó en 5,5% a CP 72-2086 (16,0µmolCO2·m-2·s-1) y en 25% a Mex 68 P-23 (13,5µmolCO2.m-2·s-1). Esta respuesta fue similar para la variable Tr (27,3°C); sin embargo, para las variables Tr y gsla variedad Mex 79-431 tuvo una respuesta similar (P>0,05) a la variedad CP 72-2086 pero diferente (P<0,05) a la variedad Mex 68 P-23. Los resultados permiten asumir que la variedad Mex 79-431, con mayor A, tendría la capacidad de producir más biomasa y por lo tanto mayor rendimiento económico, como lo señalan Mabrouk et al. (1990); sin embargo, esto no fue así ya que la tendencia indica que la variedad de mejor rendimiento económico fue Mex 68 P-23, aunque estadísticamente similar a CP 72-2086. Esto podría ser atribuido a que se tuvo la misma densidad de población y el mismo manejo del cultivo. Al respecto Williams et al. (1965) mencionan que el rendimiento máximo de un cultivo es una función directa de su IAFo (cantidad de follaje requerido para interceptar 95% de la radiación incidente), el cual está determinado por la densidad de población.
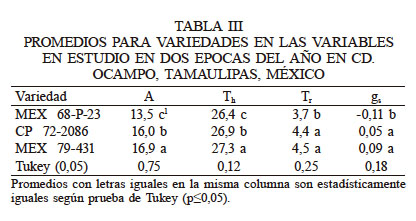
De manera general, la variedad Mex 79-431, con la mayor A, fue más eficiente en el aprovechamiento de la radiación fotosintéticamente activa (RFA) comparada con CP 72-2086 y Mex 68 P-23 así como una mayor Tr, aunque esto no se vio reflejado en una Th menor y un mayor rendimiento. Con los resultados obtenidos se asume que el genotipo estable fue Mex 79-43, aunque esto no se vio reflejado en un mayor rendimiento (Figura 4a) probablemente debido a que la densidad de siembra no es la adecuada para esta variedad. Es decir, no se ha alcanzado el índice de área foliar óptimo (IAF.) como lo señalan Valentinuz y Tollenaar (2006) para un mejor aprovechamiento fotosintético y aumentar el rendimiento del cultivo por unidad de superficie, además de aprovechar el mayor tamaño de hojas y mayor área foliar como señalan Castro et al. (2016)
Rendimiento de campo (A) y °Brix (B) de tres variedades comerciales de caña de azúcar evaluadas en condiciones de secano en Ocampo, Tamaulipas, México
En relación con la calidad del jugo medida en °Brix (Figura 4b), no se tuvieron diferencias estadísticas P>0,05 entre las variedades. Al momento del corte todas tuvieron una igual concentración de azucares, lo que significa que tienen la misma calidad del jugo. Al respecto, Aguilar y Debernardi (2004) reportaron resultados similares en concentración de azucares de la variedad Mex 72-2086, con 224% °Brix.
La interacción A x H fue altamente significativa (P<0,001) para todas las variables estudiadas. En la Figura 5a se observa que la A en invierno se redujo significativamente durante todo el día. Como era de esperarse, la A más alta se alcanzó al mediodía (42,9µmolCO2·m-2·s-1) durante el verano, como resultado de una mayor radiación. A partir de las 13:00 y hasta las 15:00, la A fue 40% menor que la obtenida en verano. La variedad Mex 79-431 tuvo la A más alta (46,39µmolCO2·m-2·s-1) a esa misma hora del día (13:00). En la Figura 5b se aprecia una reducción de la Th durante todo el día hasta de 12,8°C mientras que en verano fue de solo 5°C. En la Figura 5c se observa que la transpiración en invierno fue inestable en las primeras horas del día; a partir de las 11:00 y hasta las 15:00 la transpiración fue 54% menor que la transpiración obtenida en verano. En la Figura 5d se indica que en invierno, a partir de las 7:00 y hasta las 11:00 los estomas de la variedades estudiadas estuvieron cerrados; sin embargo, el resto del día estuvieron abiertos para el intercambio de gases, aunque en menor proporción que durante el verano, ya que en esta última época durante todo el día los estomas estuvieron abiertos, como resultado del nivel de humedad en el suelo y la temperatura del aire. De manera general el efecto de las condiciones ambientales en invierno se vio reflejado en una reducción del intercambio de gases como se muestra en la Figura 5, atribuido principalmente a la reducción de la temperatura del aire (Du et al., 1999).
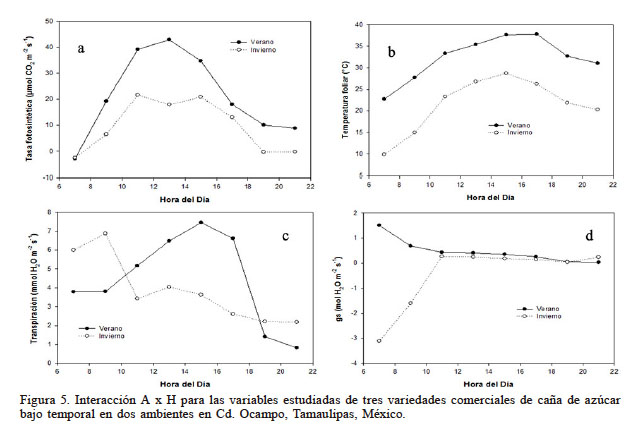
Figura 5
Interacción A x H para las variables estudiadas de tres variedades comerciales de caña de azúcar bajo temporal en dos ambientes en Cd. Ocampo, Tamaulipas, México
Conclusión
Como resultado del cambio de la época del año, las variables tasa fotosintética, conductancia estomática, temperatura de la hoja y transpiración fueron reducidas en 54, 171, 34 y 53% respectivamente. La tasa fotosintética más alta fue alcanzada en verano por la variedad Mex 79-431 con un promedio de 46,39μmolCO2·m-2·s-1, aunque fue estadísticamente similar a CP 72-2086. La variedad Mex 68 P-23 tuvo un rendimiento de 103t·ha-1 y fue superior a Mex 79-431 aunque ésta última podría tener un potencial de rendimiento superior dándole un manejo diferente, principalmente buscando el índice de área foliar óptimo. No existen diferencias entre variedades para la calidad del jugo medida en °Brix.
TASA FOTOSINTÉTICA DE GENOTIPOS DE CAÑA DE AZÚCAR BAJO CONDICIONES DE SECANO DURANTE DOS ÉPOCAS DEL AÑO
Aguilar RN, Debernardi ML (2004) Efecto de la floración en la calidad agroindustrial de la variedad de caña de azúcar CP 72-2086 en México. Caña de Azúcar 22(2): 19-37.
Brugnoli E, Lauteri M (1991) Effects of salinity on stomatal conductance, photosynthetic capacity and carbon isotope discrimination of salt tolerant (Gossypium hirsutum L.) and salt sensitive (Phaseolus vulgaris L.) C. Non-halophytes. Plant Physiol. 95: 628-635.
Castro-Nava S, Huerta AJ, Placido-de la Cruz JM, Mireles-Rodríguez E (2016) Leaf growth and canopy development of three sugarcane genotypes under high temperature rainfed conditions in northeastern Mexico. Int. J Agron. 2016: 2561026. doi:10.1155/2016/2561026.
Djanaguiraman M, Prasad PVV, Boyle DL, Schapaugh WT (2011) High-temperature stress and soybean leaves: Leaf anatomy and photosynthesis. Crop Sci. 51: 2125-2131.
Du YC, Nose A, Wasano K (1999) Effects of chilling temperature on photosynthetic rates, photosynthetic enzyme activities and metabolite levels in leaves of three sugarcane species. Plant Cell Environ. 22: 317-324.
Flores CS (2001) Las Variedades de Caña de Azúcar en México. 1ª ed. Asociación de técnicos Azucareros de México. 228 pp.
García E (1988) Modificaciones al Sistema de Clasificación Climática de Köppen. 4ª ed. Offset. Larios. México. 217 pp.
Hashemi-Dezfouli A, Herbert SJ (1992) Intensifying plant density response of corn with artificial shade. Agron. J. 84: 547-555.
Inman-Bamber NG, Bonnett D, Spillman MF, Hewitt MH, Glassop D (2010) Sucrose accumulation in sugarcane is influenced by temperature and genotype through the carbon source-sink balance. Crop Pasture Sci. 61: 111-121.
IPCC (2007) Cambio climático 2007. En Pachauri RK, Reisingger A (Eds) Contribución de los Grupos de Trabajo I, II y III al Cuarto Informe de evaluación del IPCC. Intergovernmental Panel on Climate Change. Ginebra, Suiza. 104 pp.
Kubien DS, Sage RF (2003) C4 grasses in boreal fens: their occurrence in relation to microsite characteristics. Oecologia137: 330-337.
Mabrouk GM, Helmy IM, Thampy KG, Wakil SJ (1990) Acute hormonal control of acety-CoA carboxylase. The roles of insulin, glucagon and epinephrine. J. Biol. Chem. 265: 6330-6338.
Marchiori PER, Machado EC, Ribeiro RV (2014) Photosynthetic limitations imposed by self-shading in fild-grown sugarcane varieties. Field Crops Res. 155: 30-37.
Monteith JL, Unsworth MH (1990) Principles of Environmental Physics. 2ª ed. Arnold. New York, EEUU. pp. 53-54.
Pearcy RW, Valladares F, Wright SJ, Lasso de PE (2004) A functional analysis of the crown architecture of tropical forest Psychotria species: do species vary in light capture efficiency and consequently in carbon gain and growth? Oecologia 139: 163-177.
Rea R, De Sousa-Vieira O (2001) Interacción genotipo x ambiente y análisis de estabilidad en ensayos regionales de caña de azúcar en Venezuela. Caña de Azúcar 19: 3-15.
SAS (2003) SAS Users Guide: Statistics. Versión 9.1 (TS1M3). SAS Institute, Inc. Cary, NC, USA.
Shrivastava AK, Solomon S, Rai RK, Singh P, Chandra A, Jain R, Shukla SP (2015) Physiological interventions for enhancing sugarcane and sugar productivity. Sugar Tech 17: 215-216.
SIAP (2015) Sistema Producto. Servicio de Información y Estadística Agroalimentaria y Pesquera. http://www.siap.gob.mx/.
Taiz L, Zeiger E (2002) Plant Physiology. 3ª ed. Sinauer. Sunderland, MA, EEUU. 690 pp.
Valentinuz OR, Tollenaar M (2006) Effect of genotype, nitrogen, plant density, and row spacing on the area-per-leaf profile in maize. Agron. J. 98: 94-99.
Wahid A, Gelani S, Ashraf M, Foolad MR (2007) Heat tolerance in plants: an overview. Environ. Exp. Bot. 61: 199-223.
Williams WA, Loomis RS, Lepley CR (1965) Vegetative growth of corn as affected by population density. I. Productivity in relation to interception of solar radiation. Crop Sci. 5: 211-215.
Zhao D, Glaz B, Comstock JC (2013) Sugarcane leaf photosynthesis and growth characters during development of water-deficit stress. Crop Sci. 53: 1066-1075.
Notas de autor
scastro02@yahoo.com.mx.
