El concepto de homología en Evo-Devo y ciencias cognitivas: un enfoque histórico-epistemológico
El concepto de homología en Evo-Devo y ciencias cognitivas: un enfoque histórico-epistemológico
Cuicuilco, vol. 23, núm. 65, pp. 133-164, 2016
Instituto Nacional de Antropología e Historia
Resumen: La extrapolación de la perspectiva Evo-Devo hacia las ciencias cognitivas es prometedora y puede entrañar interesantes problemas epistémicos. Éstos tienen que ver con la diferencia entre los tipos y conceptos de causalidad en los sistemas jerárquicos propios de Evo-Devo, vis à vis las explicaciones provenientes de sistemas jerárquicos característicos de las ciencias cognitivas. Frans de Waal y Pier F. Ferrari han establecido una hipótesis de abajo-hacia-arriba para explicar las conductas imitativas de los primates en contextos afectivos. Desde este punto de vista sugieren que el término de homología profunda de Evo-Devo puede aplicarse al análisis de los niveles jerárquicos inferiores que subyacen al funcionamiento de las neuronas espejo. De acuerdo con mi estudio, el modelo propuesto por estos autores no aclara satisfactoriamente si las neuronas espejo de las distintas especies de primates tienen un mismo conjunto de determinantes genético embriológicos, condición indispensable para un uso válido del concepto de homología profunda. Cabe la posibilidad de que redes genéticas regulatorias diferentes podrían dar lugar a grupos de neuronas espejo con propiedades funcionales similares, sería delicado suponer que las capaci- dades conductuales, que en sí mismas también son propiedades funcionales, dependan de una sola implementación anatomo-funcional en los primates. A partir de un análisis histórico-epistemo- lógico del concepto de homología se revisa la hipótesis anterior y se proponen aspectos relevantes para su evaluación. Finalizo con un comentario a este horizonte epistémico que se inserta en los confines de la antropología biológica.
Palabras clave: homología, homología profunda, neuronas espejo, antropología biológica, Evo-Devo.
Abstract: The extrpolation of the Evo-Devo perspective in the cognitive sciences realm is promising but at the same time it can carry some interesting epistemic problems. Such considerations are engaged with the different type of explanation or conceptualization of causality in the hierarchichal systems of Evo-Devo, vis-à-vis the typical explanations of characteristic hierarchichal systems of cognitive sciences. Frans de Waal and Pier F. Ferrari had established a bottom-up hypothesis for explaining the imitative behaviors of primates during affective interac- tions. From this point of view, they suggest that “deep homology” a concept from Evo-Devo can be used to analyze the most inferior hierarchichal levels that underlie the functioning of mirror neurons. In accordance with my analysis, the model proposed by the authors do not satisfactorily calrify if the mirror neurons of different primate species share a set of common genetic-embriologic determinants, a mandatorily condition for a valid use of the concept of deep homology. Due to the possibility that different regulatory networks could give place to sets of mirror neurons with similar functional properties, it would be frail to establish that behavioral capabilities, which are in fact functional properties, depend on a straightforward anatomical and functional implementation in primates. From a historical and epistemological analysis of the concept of homology I revise the previous hypothesis and propose some relevant aspects for its assessment. I close this paper with a commentary on the broad epistemic landscape in which this interesting hypothesis is framed, the bioanthropological discussions.
Keywords: homology, deep homology, mirror neurons, biological anthropology, Evo-Devo.
Introducción
En la actualidad, el enfoque de la biología evolutiva del desarrollo (conocida también como Evo-Devo) se ha introducido en el campo de las ciencias cognitivas como una alternativa científica válida, la cual permitiría responder a múltiples interrogantes vinculadas a la conducta y cognición humanas [Fitch 2012]. Dicho abordaje está inmerso en un contexto evolucionista que, además, busca incorporar información de especies biológicas que se encuentran justo en la vecindad filogenética —presente y pasada— de Homo sapiens, dando lugar a un campo que bien podría caracterizarse como “Evo- Devo” humano [Mitteroecker y Gunz 2012].
Es importante anotar que en la interacción entre Evo-Devo y los abor- dajes de investigación sobre la cognición existe un subconjunto de intereses caracterizados como “ciencias cognitivas” sensu lato: un campo donde se incluyen la neuropsicología, la etología primatológica, la antropología biológica, la filosofía de la mente y otras disciplinas relacionadas. Evo-Devo es un programa de investigación integrativo que sobresale en el horizonte actual de la biología, el cual ha inspirado a algunos especialistas en ciencias cognitivas para resolver problemas específicos de su quehacer. Aquí argumento que la extrapolación de la perspectiva Evo-Devo hacia las ciencias cognitivas, si bien promete ser productiva puede entrañar algunos problemas epistémicos, los cuales se relacionan con la diferencia entre las explicaciones y conceptos de causalidad que pueden establecerse o aplicarse en los
sistemas jerárquicos propios de Evo-Devo, vis-à-vis las explicaciones provenientes de sistemas jerárquicos de las ciencias cognitivas.
Tomando en cuenta estas problemáticas interdisciplinarias, de corte epistemológico comparativo, este trabajo lo inicio con un repaso histórico del concepto de homología en biología, así como de los contextos comparativos en etología y psicología. Sigo con una revisión y un diálogo directo con algunas investigaciones recientes de biólogos y filósofos de la biología que discuten los problemas contemporáneos del término de homología, especialmente atiendo los argumentos de Brigandt y Griffiths [2007], Balari y Lorenzo [2014] y Wagner [2014], así como la propuesta de Ereshefsky [2007], que considera que las categorías psicológicas y cognitivas son susceptibles de relacionarse entre sí qua homólogos. Luego recupero el trabajo de Ehab Abouheif quien —desde una teorización sustentada en trabajo empírico en Evo-Devo— muestra la naturaleza interactiva de los sistemas del desarrollo y propone criterios que permitirían establecer con certeza distintas relaciones jerárquicas de homología, los cuales corresponden a diferentes escenarios de evolución [Abouheif 1997a, 1997b, 1999]. Sobre esta base analizo la relación entre la naturaleza intrínsecamente jerárquica de los modelos provenientes de Evo Devo y los aspectos epistémicos de las explicaciones también jerarquizadas proporcionadas por los científicos cognitivos, a partir de un estudio de caso. Finalizo con una reflexión desde la perspectiva de la antropología biológica y la etología primatológica, en función de mi orientación disciplinar.
La homología profunda en cIencIas cognitivas: un avance
Para abordar el análisis epistémico de las nociones de homología en el ámbito de la jerarquía cognitive conductual, el caso específico que analizo obtenido de la literatura en ciencias cognitivas corresponde al uso del concepto de homología profunda que puede distinguirse en los esquemas jerárquicos de Abouheif (figura 1). Esta aplicación de la idea de homología profunda la presentaron los científicos cognitivos de orientación primatológica Frans de Waal y Pier Francesco Ferrari [2010], quienes vincularon a las “neuronas espejo” [Gallesse et al. 1996; Rizzolatti 1996; Cook et al. 2014] con los mecanismos conductuales de imitación en primates, en el contexto de un abordaje de abajo-hacia arriba (bottom-up approach en inglés) para examinar la cognición animal y humana.
Desde este punto de vista, De Waal y Ferrari sugieren que el concepto de homología profunda de Evo-Devo cuya definición obtienen del
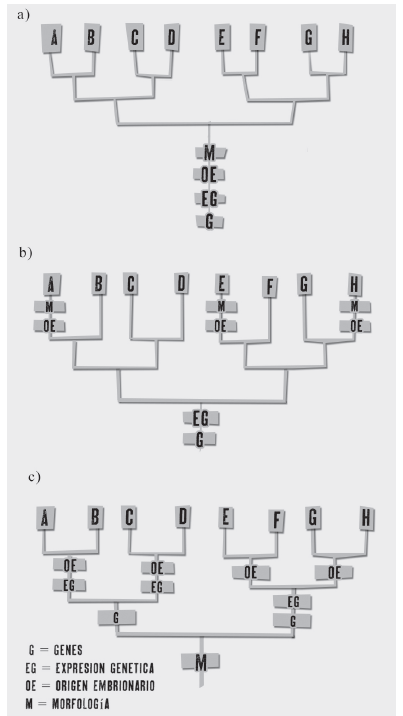
Figura 1
Las tres imágenes que componen la figura representan distintos tipos de relaciones evolutivas entre ocho diferentes taxa (AH). Los niveles de organización incorporados en las representaciones jerárquicas de homología de Abouheif [1997a] son cuatro: morfología (M), origen embrionario (OE), expresión génica (EG) y genes (G). Cada uno de estos correspon de a un nivel de organización propio de un carácter particular en las ocho especies. En la figura se distinguen tres escenarios evolutivos: a) homología verdadera: coincidencia com pleta entre niveles, b) homología profunda: coincidencia entre los dos niveles básicos (genes y su expresión), y c) convergencia de caracteres: conservación de los niveles superiores. Modificado de Abouheif [1997a]. conocido trabajo de Shubin, Tabin y Carroll [2009]— puede aplicarse al análisis de los niveles jerárquicos inferiores que subyacen al funcionamiento de las neuronas espejo. De acuerdo con mi análisis, el modelo propuesto por estos autores no aclara satisfactoriamente si las neuronas espejo de las distintas especies de primates tienen un mismo conjunto de determinantes genético-embriológicos, condición indispensable para un uso válido del término de homología profunda. Al final del trabajo argumento que los estudios recientes de Catmur [2007 y 2008] y Heyes [2010a, 2010b] indican que probablemente no todos los conjuntos de neuronas espejo se derivan de los mismos mecanismos en los niveles inferiores de la jerarquía de niveles de homología de Abouheif. Lo anterior, sugiere que en lugar de homología profunda, la relación jerárquica entre genes, neuronas espejo y conductas de imitación corresponde a otro tipo de homología.
El concepto de homología en biología y en filosofía de la biología: un breve recorrido histórico
En la revista Biology and Philosophy, editada por los filósofos de la biología Ingo Brigandt y Paul Griffiths [2007], se discutió cuál es la importancia del concepto de homología tanto para la biología como para la filosofía de la biología. El capítulo introductorio del presente número destaca que éste es uno de los términos más importantes en biología [De Beer 1971; Donoghue 1992; Wake 2003], el cual tiene una larga trayectoria histórica, con un origen que se puede ubicar en la época predarwiniana. De acuerdo con Williams [2005], la noción de homología fue consolidada en la primera mitad del siglo xix en el marco de la morfología y embriología comparativa. La definición original, acuñada por el paleontólogo inglés Richard Owen (1843), se refería a “el mismo órgano en diferentes animales bajo cualquier variedad de forma y función” [Owen 1843: 379]. Tanto en la Europa continental como en Inglaterra, el concepto de homología fue relevante en las discusiones transformistas. En el contexto de la constitución de la “teoría de la evolución por selección natural” marcó una pauta importante que llevaría a la postre a asimilar la noción de “ancestro común” [Desmond 1989; Richards 1993; Amundson 2007].1
En trabajos recientes de biología y filosofía de la biología se argumenta convincentemente sobre el valor de las nociones ligadas a homología en la biología molecular y del desarrollo en Evo-Devo. Los filósofos Brigandt y Griffiths [2007: 634] apuntan que “una vez que la morfología se interpretó como un reflejo de ancestría común demostró ser la herramienta epistémica central para los estudios filogenéticos, como los clásicos estudios desarrollados durante el siglo xix en la morfología evolutiva [Gegenbaur 1870; Lankester 1870; Nyhart 1995]. Ya para la segunda mitad del siglo xx, la sistemática filogenética moderna (la cladística) introdujo métodos más confiables para la distinción entre homologías y homoplasias,2 lo cual permitió la construcción de filogenias al estilo de Willi Hennig”.
Esta nueva forma de sistematizar la homología dio pie a la generación de un cuerpo teórico que trascendió el ámbito de la morfología dando lugar ya entrado el siglo XX— a la psicología y particularmente a la etología al establecer dicho marco teórico como punto de referencia. En ese sentido, el también filósofo de la biología Marc Ereshefsky [2007] hizo notar que los patrones de conducta y los rasgos cognitivos pueden ser reconocidos, cada vez más, como rasgos homólogos entre diferentes taxa.
Desde una mirada histórica enfocada a la etología y otros campos interesados en el comportamiento, la primera mitad del siglo xx se caracterizó por la disputa entre los conductistas y los etólogos clásicos. En relación con la segunda corriente (a la cual pondremos mayor atención), los trabajos de Konrad Lorenz y Niko Tinbergen marcaron el devenir de la etología, hicieron de este campo del conocimiento una disciplina fundamentada en la observación de la conducta animal y de las interacciones de los organismos con el ambiente. La etología demarcó sus límites con respecto a etapas pre- vias de estudio [Romanes 1883 y James 1890] al enfatizar la importancia de observar sus interpretaciones en un contexto evolucionista que le daba relevancia a las relaciones filogenéticas de las diferentes especies. Cabe destacar las críticas a esta tradición efectuadas por Lehrman [1953], donde señalaba el poco cuidado que había con los aspectos ontogenéticos de la conducta. Esta crítica es vigente para algunos abordajes actuales a la evolución de la cognición.
En las décadas siguientes, la etología fue influida por los programas adaptacionistas. Los estudios se centraron en explicar los diferentes componentes de las facultades psicológicas y mentales, como si éstas fueran adaptaciones. En esa época destacaron programas de investigación como la ecología de la conducta, la sociobiología y la psicología evolucionista [Bateson 2003, Laland y Brown 2011]. Se debe resaltar, como lo hace Ereshefsky [2007], que —aunque los programas funcionalistas y adaptacionistas otorgan un peso importante a los aspectos evolutivos— prestan poca atención a la interacción de los elementos ontogenéticos y filogenéticos de dichos rasgos psicológicos. En la actualidad, esta labor se realiza no sin dificultades desde Evo-Devo.
Para la presente investigación es relevante “la posibilidad de individualizar las clases o categorías de los fenómenos cognitivos y mentales en términos de homología (en lugar de su función evolutiva); la noción de homología tiene implicaciones para la filosofía de la mente y las ciencias cognitivas” [Brigandt y Griffiths 2007: 633]. Lo anterior permite establecer vecindades filogenéticas independientemente de su función adaptativa, y cumplir uno de los principales objetivos de la biología evolutiva.
En términos de las explicaciones causales, la anterior caracterización de homología nos ayuda a entender que “un carácter homólogo comparte muchas propiedades biológicas con diferentes organismos en los que se presenta y, por ello, existe una base causal para explicar la razón de que las compartan (ancestría común y mecanismos del desarrollo compartidos)” [Brigandt y Griffiths 2007: 634]. También desde la filosofía de Evo Devo, Love [2007] destaca la importancia de este punto, pues —en la época postoweniana darwinista— la única manera de establecer confianza en una relación de homología se fundamenta en la posibilidad de apelar a un origen común, y sólo secundariamente a la función del rasgo.
Desde una perspectiva metodológica, Griffiths [2007] retoma las ideas de Amundson y Lauder [1994] al respecto del estatus epistémico de la homología. De acuerdo con estos autores, concebimos la homología en términos de la función adaptativa del rasgo o como una inferencia que relaciona dos especies, dos órganos u otras entidades en función de que comparten un ancestro común. Con base en este argumento, Griffiths, Amundson y Lauder consideran que el establecimiento de homologías fundamentado en la función adaptativa del rasgo es más demandante epistémicamente hablando— de lo que resulta el método inferencial ya referido. Dicho de otro modo: contamos con diferentes métodos para establecer homologías pero, ¿cuál es la relación entre métodos analíticos y los tipos de homología?
Evo-devo y la discusión actual sobre los tipos de homología
En la literatura especializada de la biología evolutiva es posible encontrar diferentes tipos de homología. La homología táxica o taxonómica (también homología de caracteres morfológicos) se refiere a que dos especies dísimiles comparten un carácter por ancestría común, que Owen distinguió como una “homología especial”. Reconoció homologías en el interior de un propio organismo, “la homología serial”, por ejemplo, las vértebras de la espina dorsal son homologías seriales entre sí, mientras que los huesos correspondientes a las extremidades anteriores de distintas especies de tetrápodos son el resultado de un proceso de homología especial [Griffiths 2007].3
En términos generales, la homología táxica se reduce a la comparación de estructuras morfológicas y no considera a la conducta como un aspecto susceptible de ser caracterizado como un rasgo homólogo. Es importante apuntar que estos dos tipos de (conceptos) homología4 son el resultado de un análisis eminentemente descriptivo, en el cual los elementos causalesmecanísticos que favorecen la ocurrencia repetida de ciertos rasgos en el interior del mismo organismo, o que éstos se compartan entre especies, quedan totalmente indeterminados.
Dentro de una línea de investigación que comenzó en la década de los ochenta la cual recientemente se publicó en un libro, el biólogo y filósofo de la biología Gunter P. Wagner sugiere la posibilidad de definir el concepto de homología en términos de los mecanismos de desarrollo que generan correspondencias entre los progenitores y la descendencia. A esta noción Wagner ha denominado homología biológica [Wagner 1989, 2007, 2014]. Un primer elemento que es necesario destacar desde la perspectiva epistemológica es justamente la diferencia que se establece ante la homología táxica.
Como ya señalé, mientras que la homología serial y especial son recursos epistémicos que permiten describir la forma del rasgo de una especie en comparación con otra, la homología basada en factores ontogenéticos —específicamente el concepto de homología biológica sensu Wagner— intenta establecer los aspectos mecanísticos que subyacen al hecho de que dos especies distintas compartan o no elementos procedentes de un ancestro común.
En alusión a los conceptos wagnerianos, Griffiths [2007] destaca que “la única versión operativa del ‘concepto biológico de homología’, desde la perspectiva del desarrollo, define a la homología en función de la continuidad genética expresada en la ontogenia del rasgo; sin embargo, esto es inadecuado” [Griffiths 2007: 650]. A este respecto, Wagner alega que es necesario entender por qué las estructuras y procesos morfológicos retienen su identidad a través del tiempo evolutivo, de modo parcialmente independiente de la identidad específica de las secuencias del genoma que están involucradas en su ontogenia [Wagner 1989, 2007, 2014].
En el mismo tono Griffiths afirma que “la homología [entendida así]5 desde el desarrollo no desplazará a las explicaciones basadas en ancestría común, ya que ambas tienen elementos estrictamente complementarios para una explicación completa, al menos en el caso de la homología táxica. Además, parece posible que cuando los procesos que generan homologías queden absolutamente comprendidos, la homología morfológica podrá ser subdividida en diferentes categorías basadas en el proceso, de la misma manera que ha sucedido con la homología molecular” [Griffiths 2007: 651]. Considero que los argumentos vertidos por Wagner en su reciente libro [2014] —en el que sintetiza gran parte de su trabajo de varias décadas centrado en la importancia de homología en las ciencias comparativas— son una buena plataforma para responder a los cuestionamientos de Griffiths. El estudio señala que la homología puede ser el elemento unificador de las perspectivas funcionalista y estructuralista de la biología evolutiva contemporánea, es decir, los aspectos vinculados a las trayectorias de desarrollo que se comparten entre especies distintas parecen reflejar de mejor manera el origen común a lo que puede extraerse del análisis enfáticamente histórico que nos ofrece la homología en su versión clásica.
No obstante, el estudio que se realice desde una visión que enfatice aspectos funcionalistas o adaptacionistas no es soslayable. En todo caso, considero necesario ubicar en su dimensión a los diferentes mecanismos y procesos en la dinámica de los sistemas vivos que se encuentran sujetos tanto a los efectos de la selección natural como a las restricciones que cana- lizan el desarrollo de los individuos. La relación entre estos dos elementos fundamentales del proceso evolutivo implica reflexiones importantes en el debate actual del papel que Evo-Devo —y también las ciencias cognitivas— juegan dentro del conjunto total de las aproximaciones científicas evolucionistas.
El trabajo de Balari y Lorenzo [2014] es otra referencia reciente que merece incorporarse en el contexto de las actuales discusiones epistemológi- cas sobre el concepto de homología biológica en ciencias cognitivas —incluyendo, en este caso, temáticas de una filosofía naturalista de la mente. Estos autores se enfocan en la discusión entre la “teoría de la identidad entre mente y cerebro” y la “hipótesis de la realización múltiple”. Aunque el tema escapa a los intereses de la presente investigación, algunas de sus reflexiones pueden ser interesantes para apuntalar lo dicho hasta el momento.
De manera central, Balari y Lorenzo [2014] se interesan en mostrar las ventajas que caracterizan al abordaje del concepto biológico de homología ante el concepto histórico que se enfoca en el criterio principal de la ancestría común. En otras palabras, ellos argumentan que el conocimiento preciso de las trayectorias de desarrollo en especies que se comparan son un mejor indicador de procesos y redes de regulación genética compartida a lo que puede establecerse con el criterio historicista de homología.
Un ejemplo que puede dar luz a estas afirmaciones es el de las redes genéticas del desarrollo que generan como resultado los ojos de diversas especies de animales e insectos. Taxa tan alejadas filogenéticamente como los artrópodos y los mamíferos comparten los genes que regulan la morfo- génesis de los aparatos visuales de las especies correspondientes a estos grupos animales [Gehring e Ikeo 1999; Gehring 2005], contrariamente a lo que el mismo Darwin pensaba. Cuando se comparan superficialmente estas estructuras orgánicas parecieran no ser el resultado de la ancestría común. En cambio, una mirada más cuidadosa refleja que son la consecuencia de la retención y conservación prolongada de trayectorias ontogenéticas que modifican la relación entre los aspectos estructurales y funcionales de los individuos, lo cual genera fenotipos muy diversos procedentes de materiales genéticos compartidos. En resumen, Balari y Lorenzo [2014] vuelven a enfatizar la importancia que tiene el análisis empírico y conceptual de los tipos de homología en las discusiones actuales sobre la relación entre ontogenia y filogenia.
Homología profunda: un concepto fundamental en evo-devo y ciencias cognitivas
Uno de los aspectos conceptuales más sobresaliente a considerar en este trabajo es la comprensión y utilización que, en temas específicos dentro de las ciencias cognitivas, realizan diferentes grupos de investigación de los términos rectores de Evo-Devo. Existen algunas nociones que, al ser centrales para Evo-Devo, pueden tener una utilidad potencial al aplicarse en el dominio de las ciencias cognitivas.
Como se anticipó en la introducción, entre estos conceptos sobresale el de homología profunda (deep homology en inglés). La homología profunda es uno de los términos fundacionales de Evo-Devo como un campo disciplinario. Vale la pena destacar que —en contraste con los conceptos “clásicos” de homología— éste se ha derivado del sorprendente descubrimiento de la existencia de los genes del desarrollo, conservados a través de grupos taxonómicos animales con muy diversos planes corporales —en investigaciones que son precisamente la base del trabajo de Balari y Lorenzo, ya comentado líneas arriba.6
En un trabajo preparado para la celebración del 150 aniversario de la publicación del Origen de las especies de Darwin, los biólogos Evo-Devoístas Neil Shubin, Cliff Tabin y Sean Carroll [2009: 818] precisaron este concepto como “una comunidad de procesos causales genético-moleculares que no obstante fundamentan la divergencia embriológica y morfoanatómica característica de numerosos grupos taxonómicos”. Esta definición proviene de las reflexiones de estos autores en relación con la genética molecular del desarrollo, la paleontología y la morfología comparativa de las extremidades de los animales, que se remonta a un trabajo previo conjunto [Shubin, Tabin y Carroll 1997].
En cuanto a los estudios de evolución humana influenciados por Evo-Devo es importante destacar un trabajo reciente que discute la aplicabilidad del concepto de homología profunda al caso de la facultad lingüística. En su estudio Fitch [2012] indica la distinción entre su postura y la tradición derivada del trabajo de Noam Chomsky, la cual considera a esta conducta humana como carente de estructuras y funciones homólogas con otras especies. El trabajo de Fitch es una referencia relevante acerca de cómo algunos aspectos de la cognición y la conducta en Homo sapiens son abordados desde esta perspectiva.
Ontología intrínsecamente jerárquica en Evo-Devo y la homología conductual
En una tríada de publicaciones de trascendencia para la teoría en Evo-Devo, Ehab Abouheif [1997a, 1997b, 1999] estableció que los sistemas Evo-Devo pueden caracterizarse en términos de una dinámica interaccionista entre diferentes niveles de organización, en forma de esquemas jerárquicos. Los niveles de organización incorporados en las representaciones jerárquicas de homología de Abouheif son cuatro: morfología, origen embrionario, expresión génica y genes (del desarrollo, figura 1, a, b y c).
Al revisar las diferentes posibilidades de coincidencia —o conservación —de estados de carácter en cada nivel jerárquico, en un nodo determinado de un cladograma o árbol filogenético, Abouheif [1997a] distingue tres escenarios evolutivos: (i) el de coincidencia completa entre niveles;
-
el de coincidencia entre los dos niveles básicos (genes y su expresión), y
-
el de conservación de los niveles superiores (morfologías y orígenes ontogenéticos de la misma; figura 1, a, b y c). El primer caso —el menos problemático, o “ideal” (figura 1 a)— pertenece a la homología total o “verdadera” descrita por Bolker y Raff [1996];7 el segundo —denominado por Abouheif [1997a] como “de oportunidad ontogenética” (figura 1 b)— corresponde a la homología profunda [Shubin, Tabin y Carroll 2009].8 El tercer esquema —denominado originalmente por Abouheif [1997a] como “de manteni- miento de rasgos morfológicos por la selección natural, a pesar de la fija- ción de nuevas bases genético-embriológicas” (figura 1 c)— será de especial interés al analizar la manera como De Waal y Ferrari [2010] emplean el concepto de homología profunda.
Otra referencia destacada para este trabajo es la del filósofo de la biología Marc Ereshefsky [2007], quien ha defendido la importancia de conside- rar las categorías psicológicas como homologías. Al parecer, este autor hace eco de algunos biólogos de orientación etológica, quienes —no necesariamente en el contexto de la discusión sobre el carácter ontológico de las jerarquías planteadas por Evo-Devo— desecharon que la homología con- ductual deba identificarse con una estructura morfológica o un movimiento particular [e.g. Wenzel 1992; Lauder 1994]. Desde este punto de partida,
¿cómo debería concebirse a la conducta y la cognición sin abandonar la perspectiva conceptual de la homología ofrecida por Evo-Devo? ¿De qué manera debería incorporarse la visión de Abouheif al respecto?
La noción de que las homologías conductuales no requieren la presencia de un correlato morfológico refleja una visión jerárquica de la homología.
De acuerdo con Hall [1994], una visión de este tipo del concepto de homología señala la posibilidad de analizar diferentes niveles de organización de tal estructura homóloga; lo cual demuestra la clara naturaleza jerárquica del examen de los rasgos homólogos [Ereshefsky 2007]. Sin embargo, hay dos cosas que vale la pena recordar: “La primera es que las homologías conductuales no necesitan reducirse a subestructuras físicas, ya que una homología conductual puede ser el resultado de estructuras morfológicas y movimientos diferentes. La segunda, y que va de la mano, es que las instancias de una homología conductual son ampliamente variables. No obstante, la naturaleza jerárquica de la homología conductual ayuda a explicar esa variación” [Ereshefsky 2007].
Si nos basamos en el trabajo de M. Ereshefsky [2007] es posible establecer dos conclusiones preliminares: i) diferentes factores ontogenéticos y sustratos morfológicos subyacentes dan como resultado la misma conducta porque ésa es una instancia de las mismas unidades filogenéticas (homólogas), y ii) la filogenia es esencial para la identificación de variantes conductuales del mismo comportamiento.
A partir de lo anterior surge la necesidad de estudiar y tomar en cuenta los procesos ontogenéticos de la morfología y la conducta. Esto permitiría mejorar nuestro entendimiento de las categorías psicológicas en sus aspectos filogenéticos y ontogenéticos; particularmente la relación de estos dos elementos constitutivos. Una conclusión preliminar es que el estudio de los rasgos psicológicos puede abordarse desde el marco de la biología evolutiva del desarrollo [Ploeger y Galis 2011].
Un elemento más que resulta de interés es lo que Ereshefsky [2007] considera importante recuperar de las reflexiones de los etólogos en relación con el estudio de las categorías psicológicas. En primer lugar, el estudio de la filogenia de los rasgos psicológicos nos da un mejor entendimiento de esos rasgos que abordarlos desde una perspectiva puramente adaptacionista o funcionalista. Es decir, no basta con los postulados de la “síntesis moderna” y los programas adaptacionistas para estudiar y explicar la cognición en humanos y animales. En segundo término poner atención al desarrollo de los rasgos psicológicos permite acceder a explicaciones causales mejor estructuradas que la perspectiva que ofrecen los enfoques puramente adaptacionistas. Por último, la etología muestra que esta forma de conceptualizar la homología nos faculta para analizar diferentes aspectos de la cognición. La implementación de métodos para el examen de los caracteres psicológicos o cognitivos con los diversos métodos comparativos brinda un compendio robusto para estudiar las categorías psicológicas como homologías [Ereshefsky 2007: 671].
Para finalizar este apartado es importante señalar que existen posturas contrarias a la idea aquí expresada. Es decir, para algunos investigadores [Balari y Lorenzo 2014] no hay forma de estudiar la conducta desde la perspectiva de la homología si no se toman en cuenta los aspectos ontogenéticos de la conducta. Esta consideración implica reflexionar en los sustratos subyacentes a la conducta; con lo cual despojar a las homologías psicológicas de sus correlatos físicos es problemático. No es posible decretar una homología conductual (en la visión evo-devoísta de la homología) si no se considera el sustrato genético. Esto no significa que los genes son el elemento decisivo para estipular la homología, sin embargo, sí es necesario conocer esa información para realizar el análisis. Además, en relación directa con las reflexiones de Ereshefsky no queda claro de qué manera debe incorporarse la dimensión ontogenética al análisis de los rasgos psico- lógicos homólogos. El señalamiento de que en primera instancia parece una idea interesante, no obstante, es apremiante más que señalar su relevancia, pues se requiere aterrizar esta información en una estructura física, con lo cual la propuesta de homología conductual sensu Ereshefsky [2007] (o etológica) es incompleta.
Extrapolaciones de la homología profunda desde evo-devo hacia las ciencias cognitivas: el caso de las neuronas espejo
El concepto de homología profunda, tal y como expuso Shubin y sus colaboradores [2009], lo ha empleado recientemente Frans B. M. de Waal y Pier Francesco Ferrari, como parte de su propuesta de un programa de investigación sobre homología de la conducta y sus bases neurobiológicas en primates.9 El sistema con el cual estos autores encontraron acomodo para la noción de homología profunda es una jerarquía de niveles de organización típica de la disciplina de las ciencias cognitivas. En dicha jerarquía un conjunto de conductas ligadas al aprendizaje por imitación en situaciones afectivas constituye el nivel superior, mientras que el inferior está representado por el célebre mecanismo celular-tisular denominado “neuronas espejo” [De Waal y Ferrari 2010: 203].
Antes de profundizar en la problemática epistémica de esta propuesta es necesario señalar que, para realizar el análisis que aquí se plantea, es necesario conceder a estos autores que la imitación en el orden taxonómico Primates constituye un comportamiento compartido por todo el grupo —es decir, la imitación dentro del orden es una homología conductual. Esta concesión tiene que ver con el debate que se sostiene desde hace dos décadas en relación a la imitación y la emulación. Algunos autores [Tomasello 1996] han sostenido que los primates no humanos, particularmente los chimpancés, no son capaces de imitar como lo hacemos los humanos. En contraste, lo que hacen los simios es emular el comportamiento de los demás. La diferencia entre estas dos conductas es sutil; por ejemplo, la imitación se refiere a la capacidad de los niños y los humanos para copiar la secuencia completa de pasos para cumplir un objetivo. En cambio, la emulación implica ciertas pistas ambientales que en última instancia se redu- cen al resultado del objetivo. Es decir, la emulación no copia la secuencia completa de pasos, sino simplemente la consecución del mismo resultado.
En un estudio reciente se discutió nuevamente este problema y se llegó a la conclusión de que esta dicotomía conductual entre primates humanos y no humanos no se ajusta a las observaciones realizadas [Whiten et al. 2009]. Existe evidencia de que los chimpancés y los primates no humanos cuentan con capacidades imitativas muy sofisticadas y son capaces de emular. Lo que resulta interesante es discutir si esa conducta compartida por las diversas especies de primates tiene un origen común o no. Para los propósitos de este trabajo, avalo la premisa de que las conductas imitativas de los primates son en realidad la misma conducta.
Una breve aproximación a los problemas epistémicos de la extrapolación entre evo-devo y ciencias cognitivas propuesta por de waal y ferrari
¿De qué manera emplean De Waal y Ferrari [2010] a la homología profunda en su trabajo? La respuesta es simple. El nivel inferior de organización identificado por estos autores está constituido por grupos específicos de neuronas espejo —evidentemente diferenciadas y funcionales—, los cuales presumiblemente encarnan “mecanismos homólogos profundos” del aprendizaje por imitación en todos los primates, desde su ancestro común, o antes incluso [De Waal y Ferrari 2010: 204] (figura 2 b).
Es importante destacar que de acuerdo con De Waal y Ferrari, no hay necesidad de postular un nivel jerárquico inferior a las neuronas espejo. Sin embargo, como hemos descrito antes, el concepto de homología pro- funda propio de Evo-Devo, expuesto por Shubin y sus colaboradores [2009], estipula que los mecanismos que se encuentran en el nivel más inferior son
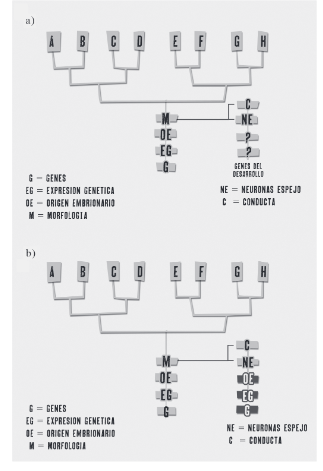
Figura 2
La imagen A es un esquema donde se compara la propuesta teórica de Abouheif y la interpretación que yo he hecho de la hipótesis de De Waal y Ferrari [2010]. De acuerdo con esta interpretación, los genes del desarrollo asociados a las neuronas espejo no están presentes en la propuesta, además de que desconocemos la expresión genética y el origen embrionario de las neuronas espejo. El inciso B muestra (en rojo) los datos faltantes para establecer la hipótesis postulada por De Waal y Ferrari [2010]. Si se tuviera esa información y coincidiese su contenido en los distintos niveles jerárquicos podríamos establecer con confianza la presencia de una homología profunda de las neuronas espejo entre los primates. Pero se carece de tal información.
obligatoriamente de naturaleza genético molecular. Siendo así, el esquema “Cogno-Evo-Devo” de De Waal y Ferrari [2010] no es equivalente al de los biólogos evo-devoístas, pese a su alusión directa al uso del término de homología profunda en dicha publicación de la literatura Evo-Devo.
Dado que el estatuto de homología profunda asignado por De Waal y Ferrari [2010] al nivel de organización jerárquica correspondiente a las neuronas espejo no hace referencia a genes, expresión génica, u origen embrionario de las estructuras, está “desacoplado” de la jerarquía usual en un sistema Evo-Devo, que comienza con un nivel genético-molecular y pasa por niveles de complejidad superior (celular, tisular, organográfico, etcétera) hasta terminar en el nivel morfológico (adulto) [Abouheif 1997] e, incluso, en el conductual (figura 2 a y b).
El “desacoplamiento” constituye un problema epistémico importante a identificar en el trabajo de De Waal y Ferrari, aunque no es el único. Otro elemento interesante es la posibilidad de evolución convergente de las capacidades imitativas de los primates, como ya se apuntó. En ese sentido, en las jerarquías tradicionalmente estudiadas por Evo-Devo —y analizadas de manera exhaustiva por Abouheif, de manera muy conveniente para el estudio de sus aspectos epistemológicos la homología puede establecerse únicamente si descartamos la posibilidad de la evolución convergente de las similitudes observadas en los niveles de organización relevantes y suponemos causalmente relacionados.
En este contexto, al hablar de “evolución convergente” hago referencia especialmente a la que podría tener lugar en el nivel inferior de la jerarquía en el ejemplo descrito por De Waal y Ferrari —i.e. el nivel de las neuronas espejo. En este punto considero crucial destacar que De Waal y Ferrari [2010: 202] son conscientes de que “sólo si conocemos los sustratos genéticos y/o neurales subyacentes, podemos clasificar a las capacidades (cognitivas) con confianza, dentro de las categorías de ‘análogas’ u ‘homólogas’”.
Es necesario insistir en que aún no se ha publicado información definitiva acerca de las similitudes o diferencias entre los complejos circuitos regulatorios que dan lugar a los núcleos de neuronas espejo en las distintas especies de primates con capacidades imitativas. Esta información es indispensable para establecer con confianza que la relación jerárquica de homología que involucra a dichos elementos es una homología profunda (figura 2 b). Como cabe la posibilidad de que redes genéticas regulatorias disímiles generen grupos de neuronas espejo con propiedades funcionales similares, es delicado suponer —más allá del análisis del trabajo de De Waal y Ferrari [2010] que las capacidades conductuales, que en sí mismas son propiedades funcionales, dependan de una sola implementa- ción anatomo-funcional (molecular-celular-tisular) en los primates.
Reflexiones relativas a la propuesta que considera a las neuronas espejo como una homología profunda
La biología evolutiva del desarrollo (Evo-Devo) ha tomado un camino alternativo y complementario a la “síntesis moderna” para estudiar la evolución de los organismos. Principalmente es una propuesta de integración de los procesos de desarrollo ontogenético como factores cruciales en la evolución, tan importantes como el mecanismo de la selección natural, la deriva génica y los demás elementos que forman parte del arsenal explicativo de la teoría evolutiva convencional. De acuerdo con algunos autores, la configuración teórica producto de la unión entre Evo-Devo y las perspectivas tradi- cionales de la evolución podría desembocar en una “síntesis evolutiva extendida” [e.g. Pigliucci y Müller 2010].
Desde una perspectiva teórica —predominantemente epistemológica pero bien informada acerca de las discusiones empíricas en la intersección entre Evo-Devo y las ciencias cognitivas que abordan los problemas primatológicos—, en este trabajo me he enfocado en uno de los conceptos rectores de la biología evolutiva: la homología. Presenté un resumen histórico que apuntala que a partir del estudio clásico de Owen [1843], el término de homología se ha reformulado, dando pie a nociones más sofisticadas que se relacionan con consideraciones ontogenéticas —e.g. la homología biológica sensu Wagner [2014] y referencias del mismo autor ahí citadas— en las que se encuentra una de las ideas más distintivas de Evo-Devo como aproxima- ción teórica en evolución: la homología profunda [Shubin et al. 2009].
He destacado que —en el contexto de varias discusiones actuales sobre el uso del concepto de homología respecto de la conducta y la cognición en los animales— la extrapolación de teorías y modelos de Evo-Devo hacia las ciencias cognitivas resulta interesante. Sin embargo, este tipo de extrapolaciones debe hacerse con sumo cuidado para evitar problemas epistémicos. Es el caso para el trabajo de De Waal y Ferrari [2010], quienes afirman que las neuronas espejo son el locus anatomo-fisiológico de una homología profunda en los primates, subyacente a sus conductas imitativas en situaciones afectivas.
La propuesta de De Waal y Ferrari [2010] debe revisarse desde el marco epistémico proporcionado por Evo-Devo, en particular con el apoyo de la visión jerárquica de la homología articulada por Ehab Abouheif en varios de sus trabajos de finales de la década de los noventa. En su esquema, De Waal y Ferrari destacan a las neuronas espejo como el nivel más básico del sistema espejo que subyacen a la conducta imitativa. Si concedemos que efectivamente las neuronas espejo representan el nivel inferior en una jerarquía de las conductas imitativas de los primates se demuestra que es posible esquematizar los procesos conductuales desde una perspectiva jerárquica. Sin embargo, De Waal y Ferrari [2010] han ido más allá al interpretar esta esquematización como la fundamentación de que las neuronas espejo, en los primates, representan el elemento causal común de su comportamiento imitativo de los primates. En ese sentido, dicha conducta tendría un origen filogenético común, cuya base biológica —justamente, la función de las neuronas espejo— sería también compartida.
La interpretación anterior es el asunto central que se ha revisado y criticado en este trabajo. Como he argumentado, el desacoplamiento en el esquema de De Waal y Ferrari [2010] respecto a la relación jerárquica que en la teoría Evo-Devo recibe el nombre de homología profunda (figura 2 a, b), implica una comprensión equivocada de lo que esta última significa originalmente (figura 1 c). Una homología profunda supone la comunidad de información en los niveles más inferiores o “básicos” de una jerarquía biológica —niveles que corresponden a la expresión de información de naturaleza genético molecular.
El elemento característico de la prerrogativa denominada “homología profunda” sensu Abouheif [1997] no está presente en el esquema propuesto por De Waal y Ferrari, aunque las neuronas espejo representan el nivel más básico del esquema sugerido por estos investigadores, ese rango no está ocupado por ningún elemento que haga referencia a genes o a expresión génica (figura 2a). Todo lo contrario, las neuronas espejo, en el modelo de De Waal y Ferrari [2010], hacen referencia a células ya diferenciadas funcionalmente que pueden o no tener un origen genético distinto. Éste es el problema epistemológico fundamental de la propuesta de De Waal y Ferrari [2010] respecto de homología profunda, el cual sólo podría subsanarse si se contara con información desde la genética molecular, que demostrara que la diferenciación celular de las neuronas espejo efectivamente depende de la expresión de las mismas redes genéticas regulatorias.
La disyuntiva epistemológica implícita en el caso analizado se relaciona con el problema de la homología tanto en el sentido de Evo-Devo como en el sentido clásico. De Waal y Ferrari [2010] no aclaran si las neuronas espejo —que en última instancia conforman el sistema homólogo en las diferentes especies de monos y en los humanos— tienen o no un origen genético distinto entre sí y sólo comparten los niveles más altos de la jerarquía, representados por la conducta en sí misma: la imitación. Desde la perspectiva clásica de la homología, esta última configuración se consideraría como una homología, incluso, desde la concepción conductual podría aceptarse como una homología conductual.
Recordemos que en la visión etológica la homología fue despojada de cualquier elemento físico o morfológico; entendida así, la homología conductual puede establecerse incluso en aquellos comportamientos que no se fundamentan en las estructuras físicas de dos o más especies. En cambio, en el caso de la homología profunda no existe alternativa que coincida con la información genética de las dos especies en cuestión para descartar con certeza la presencia de una convergencia evolutiva. Lo que se observa en la propuesta de De Waal y Ferarri [2010] es la carencia de información para fijar con confianza absoluta que las conductas imitativas de las dife- rentes especies de primates no humanos y los humanos descansan en los mismos genes y se expresan de la misma manera.
Como anoté previamente, podría suceder que diferentes genes, con expresión genética distinta, fueran reclutados eventualmente —a lo largo de la filogenia de las especies y de la ontogenia de los individuos— para participar en la diferenciación de (las redes de) neuronas espejo. Estas redes sustentarían comportamientos similares, pero éstos no estarían fundamen- tados en la expresión de información compartida —es decir, homóloga, en el sentido clásico— en los niveles más básicos.
Las neuronas espejo y los estudios experimentales sobre conducta
¿Qué perspectivas ofrece la literatura más reciente acerca de la evolución de los mecanismos genético moleculares que subyacen a las neuronas espejo? Catmur et al. [2007 y 2008] y Heyes [2010a y 2010b] mostraron que la respuesta de este tipo de neuronas puede modificarse con la experiencia sensorio motriz. En mi opinión, esto sugiere la posibilidad de que esas células tengan orígenes genéticos distintos y sólo durante su desarrollo —a partir de procesos epigenéticos, probablemente— den lugar a conductas homólogas que descansan en estructuras genéticas no homólogas. El debate de la evolución del “sistema de neuronas espejo” implica que se trata de una adaptación que facilita la comprensión de la acción. De manera alternativa, las neuronas espejo pueden ser un subproducto del aprendizaje asociativo.
Es interesante preguntarse como lo ha hecho Heyes, ¿de dónde vienen las neuronas espejo? ¿Cómo podemos explicar su capacidad para hacer coincidir las acciones observadas con las ejecutadas? Una posibilidad —como se apuntó anteriormente— es que las neuronas espejo sean una adaptación o una característica que evolucionó para cumplir una función particular, probablemente para comprender la acción propia y la de los demás [Rizzolatti et al. 2001]. De acuerdo con esta perspectiva, las neuronas espejo fueron favorecidas por la selección natural porque les permitió a los individuos comprender lo que otros estaban haciendo; se conoce como “hipótesis adaptacionista”.
Otra característica importante de esta hipótesis es que, de manera innata, las neuronas espejo de los primates están diferenciadas funcionalmente. En consecuencia, considera que la experiencia juega un rol menor en su desarrollo. Sin embargo, Catmur et al. [2007 y 2008] y Heyes [2010a] apuntan que la experiencia sensorial puede desencadenar o facilitar el desarrollo de las neuronas espejo; aun cuando señalan que su capacidad para hacer coincidir acciones observadas con acciones ejecutadas está genéticamente heredada.
La perspectiva alternativa en esta discusión es la que estipula que las neuronas espejo son un producto del aprendizaje asociativo. La “hipótesis asociativa” sugiere que las neuronas espejo se forman en el curso del desarrollo individual a través de los mismos procesos de aprendizaje que se producen con el condicionamiento pavloviano [Shultz y Dickinson 2000, en Heyes 2010]. Esta “secuencia asociativa de aprendizaje” o “hipótesis asociativa” suscita que cada neurona espejo se forja a través de la experiencia sensorio motriz, por medio de la correlación de observar y ejecutar la misma acción.
Pero, ¿cuál es la dinámica que generan este tipo de células neurales diferenciadas con la propiedad especular, la de hacer coincidir acciones observadas con las ejecutadas? Se ha propuesto que los individuos comienzan su vida con neuronas visuales que responden a la observación de una acción y un conjunto distinto de neuronas se activan durante la ejecución de una acción. Algunas de las neuronas motoras se convierten en neuronas espejo si los individuos obtienen experiencia en la que la observación y ejecución de acciones similares están correlacionadas, cuando ocurren en un tiempo relativamente corto de tiempo, y la una predice a la otra [Heyes 2010]. Esta experiencia, que fortalece los vínculos entre las neuronas visuales y motoras que codifican acciones similares, es común cuando los infantes humanos se observan a sí mismos actuar directamente o con la utilización de un espejo; cuando son imitados por otros o se involucran en actividades sincrónicas como los deportes y la danza [Heyes 2005; Ray y Heyes 2011].
Estas nociones son consistentes con estudios recientes donde se muestra que la actividad de las neuronas espejo de músicos y bailarines es diferente que la de otras personas [D’ Ausilio et al. 2006; Haslinger et al. 2005] y de que los monos adquieren neuronas que “responden a la utilización de herramientas” por medio de la experiencia [Ferrari et al., 2005]. Si la hipótesis asociativa es correcta, el mecanismo que produce las neuronas espejo —el aprendizaje asociativo— debió evolucionar, pero como está presente en diversas especies y opera en un rango muy amplio de entradas de información, no existe ninguna razón para creer que el aprendizaje asociativo es una adaptación para la producción de neuronas espejo [Heyes 2010].
Existe una dimensión que se ha abordado de manera marginal, la ontogenia de las células que conforman el sistema espejo. Lo planteo nuevamente como una interrogante ¿la función asociada con las neuronas espejo está presente en los primates desde el nacimiento? La evidencia de la actividad del sistema de neuronas espejo en monos y humanos recién nacidos apoyarían bastante la hipótesis adaptativa sobre la asociativa; sin embargo, no existe certeza sobre ello. Los datos sobre imitación en recién nacidos no son suficientes para mostrar que las neuronas espejo se encuentran presentes desde el nacimiento [Heyes 2010].
Además —y en contra de la hipótesis adaptativa— hay aspectos vinculados a la experiencia. Por ejemplo, estudios con músicos y bailarines también indican que la experiencia modula la actividad del sistema de neuronas espejo de los humanos [D’Ausilio et al. 2006; Cross et al. 2006; Margulis et al. 2009]. Hay mayor activación especular en los pianistas que en los no pianistas durante la observación de movimientos de los dedos mientras tocan el piano [Haslinger et al. 2005], y en bailarines de ballet clásico que en bailarines de capoeira durante la observación de movimientos de ballet [Calvo- Merino et al. 2005].
Apelo a las evidencias que favorecen a la hipótesis asociativa por sobre la adaptativa que provienen de experimentos que muestran específicamente que la experiencia sensoriomotriz puede mejorar [Press et al. 2007], abolir [Heyes et al. 2005] e incluso revertir [Catmur et al. 2007, 2008] la activación especular en sujetos humanos. La hipótesis que dio lugar a la realización de estas pruebas es que si el desarrollo del sistema espejo depende del aprendizaje sensoriomotor, entonces se podría utilizar el entrenamiento sensoriomotor para cambiar el funcionamiento de un sistema espejo maduro e incluso dotarlo de propiedades “contraespeculares” [Catmur et al. 2007]. Los resultados de estos experimentos indican que las propiedades especulares del sistema espejo no son totalmente innatas ni están fijas una vez adquiridas, por el contrario, se despliegan a partir del aprendizaje sensoriomotor y son susceptibles de revertir su respuesta especular.
El experimento consistió en presentar el movimiento de un dedo de la mano, que estaba posicionada frente al sujeto, para que de manera especular lo moviera de acuerdo con el movimiento del experimentador. Otro grupo de sujetos debía mover un dedo diferente del que meneaba el experimentador, en esta condición no había una acción especular. Tal como predijo la hipótesis del aprendizaje sensoriomotor, el entrenamiento incompatible (el segundo grupo de sujetos) causó la reversión de la reacción músculo específica durante la observación de la acción, mientras que el entrenamiento compatible mantuvo el patrón sin cambios. Es decir, el aprendizaje sensoriomotor reconfiguró la respuesta del sistema espejo.
Si la hipótesis asociativa es precisa, las neuronas espejo no sólo apoyan sino que están sustentadas en la socialidad humana. No son producto directo de la evolución (selección natural), sino de la experiencia sensorio-motriz, gran parte de ésta se obtiene a partir de la interacción con los demás individuos [Catmur et al. 2007 y 2008]. Las propiedades especulares del sistema espejo son genuinas pero no intrínsecas, dependen de la contingencia de la experiencia en lugar de la similitud objetiva entre estímulo y respuesta. Las respuestas del sistema espejo se desarrollan como resultado de procesos generales del aprendizaje sensoriomotor asociativo, lo cual implica que la naturaleza del estímulo que puede permear estas asociaciones no tiene restricciones [Catmur et al. 2007].
En el nivel conductual una posible conclusión preliminar es que el sistema espejo, desde la perspectiva de la teoría sensoriomotora, es un producto y un proceso de la interacción social, contribuye en la adquisición del lenguaje, en la teoría de la mente y en nuestra capacidad de interacción social compleja [Catmur et al. 2007 y 2008; Heyes 2010a]. Es decir, la presencia de un mecanismo donde las representaciones sensoriales y motoras de una acción, experimentadas de manera correlacionada durante el desarro- llo como resultado de la interacción social, configuran al sistema espejo humano para que coincidan acciones ejecutadas con las observadas.
Finalmente, la información genética que se necesita para establecer el tipo de homología, que sería el de las neuronas espejo de los primates, todavía está ausente. Se requiere de más trabajo para determinar con confianza si las neuronas espejo de las distintas especies de primates comparten la misma información genética, que a la postre se expresa en las conductas imitativas de los primates. Si fuese así, habría que discutir si lo que tenemos ante nosotros es una homología profunda o una homología verdadera.
Un apunte historiográfico final
La crítica efectuada en el presente trabajo se ha expresado desde una perspectiva enfáticamente epistemológica. Sin embargo, el trabajo de De Waal y Ferrari [2010] se enmarca en los límites de la etología primatológica; una de las corrientes que conforman la visión de las ciencias cognitivas sensu lato. En ese sentido, es importante reflexionar sobre la relevancia que tiene la antropología biológica en estos menesteres. El enfoque actual de este horizonte epistémico tiene su raíz en las propuestas clásicas de Washburn [1951, 1973]. El primero de los estudios mencionados es una referencia crucial para los antropólogos físicos, pues se hace un llamado al establecimiento de métodos y técnicas particulares para el pensamiento antropofísico, es un parteaguas en los postulados teóricos de la disciplina.
El estudio que he referido es The New Physical Anthropology [1951] y aborda una nueva manera de estudiar el fenómeno humano con especial énfasis en la relación entre los elementos biológicos constitutivos del ser humano, así como de sus características históricas, sociales y culturales. De manera sintética, el trabajo de Washburn es un exhorto a trascender el profundo compromiso con la tarea descriptiva de esta disciplina en relación con las singularidades físicas de las diversas poblaciones humanas. Es decir, no es suficiente considerar los aspectos fenotípicos de las personas y compararlos entre sí, sino que se busca explicar de manera amplia la correspondencia entre las particularidades físicas de los individuos y su relación con el ambiente.
El segundo trabajo citado que considero relevante en este contexto es el de “la promesa de la primatología” (The Promise of Primatology). En 1973 Washburn señaló cuáles eran las bondades de utilizar la primatología como marco de referencia para el estudio de la evolución humana. No únicamente en los aspectos anatómicos la primatología tiene algo que decir en relación con la evolución humana, incluso en aquéllos tan complejos como la evolución de la cognición y la conducta es un requisito indispensable del análisis.
Es evidente que la primatología ha tenido un papel importante en los estudios de evolución humana desde mediados del siglo pasado, sin embargo, el enfoque que ha tenido mayor vinculación con la evolución humana es el de la antropología física o biológica. Siguiendo a Washburn [1973], la primatología es más que simplemente el estudio de nuestros parientes más próximos, después de todo, el ser humano es un primate [Washburn 1973]. Es un hecho que las ideas de este autor le dieron la razón, pues como señaló “el estudio de los primates no humanos es útil para dar luz sobre la naturaleza humana” [Washburn 1973].
Siguiendo el razonamiento anterior pondremos como ejemplo un investigador que realiza reflexiones que intersectan algunas de las dimensiones postuladas por Washburn [1951]. Agustín Fuentes es un antropólogo físico estadounidense que ha recuperado de manera muy clara la visión que tenía Washburn respecto del quehacer de la antropología física. Fuentes integra sus inquietudes epistémicas con su trabajo empírico —como en su momento hiciera Washburn— para generar cavilaciones que cuestionan el estatus actual de nuestra materia, así como las relaciones disciplinares que pueden establecerse con la biología evolutiva y la primatología en el contexto de la evolución humana.
El llamado de Fuentes [2010] —emulando a Washburn— es hacia el establecimiento de una “nueva antropología biológica” (The New Biological Anthropology). De la misma manera que Washburn dotara al pensamiento antropofísico de la perspectiva evolucionista —particularmente de los pos- tulados darwinistas y neodarwinistas—, hoy Fuentes llama a la incorporación de perspectivas actuales y alternativas que son discutidas en el ámbito de los estudios evolucionistas. De lo que se trata es de reevaluar los alcances de la selección natural. Asimismo se pretende incorporar en el discurso y, sobre todo en el análisis, teorías y conceptos que expliquen mejor los aspectos vinculados a la evolución humana.
Dichas explicaciones no se reducen a los elementos biológicos, sino que consideran la herencia y reproducción de conductas y tradiciones culturales, que no pueden explicarse desde el paradigma simplista de la adaptación de los individuos al ambiente. Por ejemplo, Fuentes [2010] precisa tres elementos teóricos concretos que deben incorporarse a esta nueva visión de la antropología biológica: i) los estudios sobre “teoría de sistemas en desarrollo” [Oyama 1999: 2001] es una vertiente de la psicología del desarrollo que señala la importancia de los procesos ontogenéticos en la evolución de los organismos y en el desarrollo de los individuos; ii) la evolución en cuatro dimensiones [Jablonka y Lamb 2005] consiste en un enfoque que establece la existencia de al menos cuatro niveles de organización distintos a partir de los cuales se puede estudiar la evolución humana: el genético, el epigenético, el conductual y el simbólico; que representan las diferentes formas como se heredan desde los genomas hasta las conductas, y iii) la “teoría de construcción de nicho” [Odling-Smee y Laland 1996, 2003; Laland y Odling-Smee 1996, 2001a y 2010b; Laland y O’Brien 2011], propuesta que difiere de los postulados neodarwinistas que consideran la relación organismo-ambiente como una interacción constructivista mutua y bidireccional.
Se mantiene la idea de que los organismos están sujetos a las presiones del ambiente en busca de su adaptación pero, además, se considera que de la misma manera los organismos representan una presión de selección para el ambiente. La influencia del medio en el individuo no es la única interacción presente en esta relación, también el ambiente responde a la presencia y a los efectos de los organismos. El individuo deja de ser un ente pasivo en esta vinculación organismo-ambiente.
Estas breves reflexiones tienen como propósito mostrar que la discusión epistemológica en términos teóricos y metodológicos en la antropología física es un debate abierto. El presente trabajo es un ejemplo del tipo de análisis que se puede realizar desde nuestra trinchera considerando elementos históricos, empíricos y filosóficos. Si bien la propuesta de De Waal y Ferrari no se inscribe necesariamente en el ámbito de los estudios antropofísicos es importante señalar que una lectura desde este enfoque tiene cosas que decir respecto de los detalles sin una clara y válida aplicación conceptual. No todos los estudios primatológicos son antropofísicos, pero cualquier investigación vinculada con primates no humanos puede tener una relación con los objetivos de investigación de la antropología biológica.
Agradecimientos
El presente trabajo corresponde a la primera etapa de mi proyecto de investigación doctoral en el Instituto de Investigaciones Filosóficas de la unam. Juan Manuel Argüelles y Francisco Vergara Silva hicieron comentarios puntuales a versiones previas del texto, las cuales forma parte de un estudio acerca de los usos de los conceptos de Evo-Devo en el ám- bito de la antropología biológica. Una versión más extensa a nuestra crítica sobre el trabajo de De Waal y Ferrari está en preparación. Agradezco tam- bién a Tom Froese, Fernando Zambrana y a Lluís Barceló-Coblijn, por sus interesantes aportaciones, así como los útiles comentarios y sugerencias de dos revisores anónimos y al Centro de Estudios Filosóficos, Políticos y Sociales Vicente Lombardo Toledano por proporcionarme las facilidades para realizar esta investigación.
Referencias
Abouheif, Ehab 1997a Developmental Genetics and Homology: a Hierarchical Approach, en Trends in Evolutionary Ecology, vol. 12: 405-408.
Abouheif, Ehab et al. 1997b Homology and Developmental Genes, en Trends in Genetics, vol. 13: 432-433.
Abouheif, Ehab 1999 Establishing Homology Criteria for Regulatory Gene Networks: Prospects and Challenges. Genetics, vol. 13: 432-433.
Amundson, Ronald 2005 The Changing Role of the Embryo in Evolutionary Thought: Roots of Evo-Devo. Cambridge Studies in Philosophy and Biology.
Amundson, Ronald 2007 Richard Owen and Animal Form, en On the Nature of Limbs: A Discourse, Amundson, R. (ed.). The University of Chicago Press.
Amundson, Ronald y George V. Lauder 1994 Function without Purpose: the Uses of Causal Role Function in Evolutionary Biology. Biology and Phylosophy, vol. 9: 443-469.
Balari, Sergio y Guillermo Lorenzo 2014 Ahistorical Homology and Multiple Relizability. Psychological Phylosophy. .
Bateson, Patrick 2003 The Promise of Behavioral Biology. Animal Behaviour, vol. 65: 11-17.
Bolker, Jessica A. y Rudolph A. Raff 1996 Developmental Genetics and Traditional Homology. BioEssays, vol. 18: 489-494.
Brigandt, Ingo 2003 Homology in Comparative, Molecular, and Evolutionary Developmental Biology: the Radiation of a Concept. Journal of Experimental Zoology, vol. 299: 9-17.
Brigandt, Ingo y Paul Griffiths 2007 The Importance of Homology for Biology and Philosophy. Biology and Philosophy, vol. 22: 633-641.
Calvo-Merino, Beatriz, et al. 2005 Action Observation Acquired Motor Skills: an MRI Study With Expert Dancers. Cerebral Cortex, vol. 15: 1243-1249.
Catmur, Caroline, Vincent Walsh y Cecilia Heyes 2007 Sensoriomotor Learning Configures the Human Mirror System. Current Biology, vol. 17: 1527-1531.
Catmur, Caroline, et al. 2008 Through the Looking Glass: Counter-Mirror Activation Following In- compatible Sensoriomotor Learning. European Journal of Neuroscience, vol. 28: 1208-1215.
Cook, Richard, et al. 2014 Mirror Neurons: from Origin to Function. Behavioral and Brain Sciences, vol. 37: 177-241.
Cross, Emily S., Antonia, F. de C. Hamilton y Scott T. Grafton 2006 Building a Motor Simulation the Novo: Observation of Dance by Dancers. Neuroimage, vol. 31: 1257-1267.
D’Ausilio, Alessandro, et al. 2006 Cross-Modal Plasticity of the Motor Cortex while Listening to a Re- hearsed Musical Piece. European Journal of Neuroscience, vol. 24: 955-958.
Desmond, Adrian 1989 The Politics of Evolution. Science and its Conceptual Foundations series.
De Beer, Gavin 1971 Homology, an Unsolved Problem. Oxford University Press.
De Waal, Frans B.M. y Pierre F. Ferrari 2010 Towards a Bottom-up Perspective on Animal and Human Cognition. Trends in Cognitive Sciences, vol. 14: 201-207.
Donoghue, Michael J. 1992 Homology, en Keyword in Evolutionary Biology, E.F. Keller y E.A. Lloyd (eds.). Harvard University Press. Cambridge: 170-179.
Ereshefsky, Marc 2007 Psychological Categories as Homologies: Lessons from Ethology. Biology and Philosophy, vol. 22: 659-674.
Ferrari, Pier Francesco, et al. 2005 Mirror Neurons Responding to the Observation of Actions Made with Tools in Monkey Ventral Premotor Cortex. Journal of Cognitive Neuroscince, vol. 17: 212-226.
Fitch, W. Tecumseh 2012 Evolutionary Developmental Biology and Human Language Evolution: Constraints on Adaptation. Evolutionary Biology, vol. 39: 613-637.
Fuentes, Agustín 2010 The New Biological Anthropology: Bringing Washburn’s New Physical Anthropology into 2010 and Beyond –the 2008 AAPA Luncheon Lecture, en Yearbook of Physical Anthropology: 532-612.
Gallesse, Vittorio, et al. 1996 Action Recognition in the Premotor Cortex. Brain, vol. 119: 593-609.
Gegenbaur, Carl 1870 Grundzüge der Vergleichenden Anatomie (2a edición), Wilhelm Engelmann. Leipzig.
Gehring, William J. 2005 New Perspectives on Eye Development and the Evolution of Eyes and Photoreceptors. Journal of Heredity, vol. 96: 171-184.
Gehring, William J. y Kazuho Ikeo 1999 Pax 6: Mastering Eye Morphogenesis and Eye Evolution. Trends in Gene- tics, vol. 15: 371-377.
Griffiths, Paul 2007 The Phenomenon of Homology. Biology and Philosophy, vol. 22: 643-658.
Hall, Brian K. 1994 Homology: the Hierarchichal Basis of Comparative Biology. Academic Press. San Diego.
Haslinger, B., et al. 2005 Transmodal Sensoriomotor Networks During Action Observation in Professional Pianists. Journal of Cognitive Neuroscience, vol. 17: 282-293.
Heyes, Cecilia 2005 Imitation by Association, en Perspectives on Imitation: From Neuroscience to Social Science, vol. 1: Mechanisms of Imitation and Imitation in Animals, S. Hurley y N Chater (eds.). MIT Press. Cambridge: 157-176.
Heyes, Cecilia 2010a Where do Mirror Neurons Come From? Neuroscience and Behavioral Re- views, vol. 34: 575-583.
Heyes, Cecilia 2010b Mesmerizing Mirror Neurons. Neuroimage, vol. 51: 789-791.
Heyes, Cecilia, et al. 2005 Experience Modulates Automatic Imitation. Brain Research Cognitive Brain Research, vol. 22: 233-240.
Jablonka, Eva y Marion Lamb 2005 Evolution in Four Dimensions: Genetic, Epigenetic, Behavioral, and Symbolic Variation in the History of lLife. MIT Press. Cambridge.
James, William 1890/1983 The Principles of Psychology. Harvard University Press. Cambridge.
Laland, Kevin, John Odling-Smee y Marcus W. Feldman 1996 The Evolutionary Consequences of Niche Construction: a Theoretical In- vestigation Using Two-Locus Theory. Journal of Evolutionary Biology, vol. 9: 293- 233.
Laland, Kevin, John Odling-Smee y Marcus W. Feldman 2001a Cultural Niche Construction and Human Evolution. Journal of Evolutionary Biology, vol. 14: 22-33.
Laland, Kevin, John Odling-Smee y Marcus W. Feldman 2001b Niche Construction, Ecological Inheritance, and Cycles of Contingency in Evolution, en Cycles of Contigency: Developmental Systems and Evolution, Oyama, Susan, Paul Griffiths y Rusell Gray (eds.). MIT Press.
Laland, Kevin y Gillian Brown 2011 Sense and Non-Sense in Evolutionary Perspectives on Human Behaviour. Oxford University Press.
Laland, Kevin y Michael O’Brien 2011 Cultural Niche Construction: An Introduction. Biological Theory, vol. 6: 191-202.
Lankester, Ernst Ray 1870 On the Use of the Term Homology in Modern Zoology, and the Distinction between Homogenetic and Homoplastic Agreements. Annual Magazine of Natural History, Zoology Botany and Geology, vol. 6: 34-43.
Lauder, George V. 1994 Homology, form, and Function, en Homology: the Hierarchichal Basis of Comparative Biology, B. Hall (ed.). Academic Press. San Diego: 151-196.
Lehrman, Daniel S. 1953 A Critique of Konrad Lorenz’s Theory of Instinctive Behavior. The Quarterly Review of Biology, vol. 28 (4): 337-363.
Le Guyader, Hervé 2004 Etienne Geoffroy Saint-Hilare, 1772-1844: a Visionary Naturalist. University of Chicago Press. Chicago.
Love, Alan 2007 Functional Homology and Homology of Function: Biological Concepts and Philosophical Consequences. Biology and Philosophy, vol. 22: 691-708.
Margulis, Elizabeth Hellmuth, et al. 2009 Selective Neurophysiological Responses to Music in Instrumentalists with Different Listening Biographies. Human Brain Mapping, vol. 30: 267-275.
Mitteroecker Phillip y Phillip Gunz 2012 Human Evo Devo. Evolutionary Biology, vol. 39: 443-446.
Nyhart, Lynn K. 1995 Biology Takes Form: Animal Morphology and the German Universities, 1800- 1900. University Chicago Press. Chicago.
Odling-Smee, John y Kevin Laland 1996 Niche Construction. The American Naturalist, vol. 147 (4): 641-648.
Odling-Smee, John y Kevin Laland 2003 Niche Construction: the Neglected Process in Evolution. Monographs in Population Biology. Princeton University.
Owen, Richard 1843 Lectures on the Comparative Anatomy and Physiology of the Invertebrate Animals. Longman, Brown, Green and Longmans. Londres.
Oyama, Susan 1999 Locating Development: Locating Developmental Systems, en Conceptual Development: Piaget’s Legacy, Scholnick, Ellin K. et al. (eds.). Psychology Press.
Oyama, Susan, Paul Griffiths y Rusell Gray 2001 Introduction: what is Developmental Systems Theory, en Cycles of Contigency: Developmental Systems and Evolution, Oyama, Susan, Paul Griffiths y Rusell Gray (eds.). MIT Press.
Pigliucci, Massimo y Gerd Müller 2010 Evolution. The Extended Synthesis. MIT Press. Cambridge.
Ploeger, Annemie y Frietson Galis 2011 Evo Devo and Cognitive Science. Wiley Interdisciplinary Reviews: Cognitive Science, vol. 2 (4): 429-440.
Press, Clare, Helge Gillmeister y Cecilia Heyes 2007 Sensoriomotor Experience Enhances Automatic Imitation of Robotic Action. Proceedings of the Royal Society B, vol. 274: 2509-2514.
Ray, Elizabeth y Cecilia Heyes 2011 Imitation in Infancy: the Wealth of the Stimulus. Developmental Science, vol. 14: 92-105.
Richards, Robert 1993 The Meaning of Evolution. Science and its Conceptual Foundations Series.
Rizzolatti, Giacomo et al. 1996 Premotor Cortex and Recognition of Motor Actions. Cognitive Brain Re- search, vol. 3: 131-141.
Rizzolatti, Giacomo, et al. 2001 Neurophysiological Mechanisms Underlying the Understanding and Imitation of Action. Nature Reviews Neuroscience, vol. 2: 661-670.
Romanes, George 1883/1977 Animal Intelligence. Appleton. Nueva York.
Shubin, Neil, Cliff Tabin y Sean Carroll 1997 Fossils, Genes and the Evolution and the Evolution of Animal Limbs. Nature, vol. 388: 639-648.
Shubin, Neil, Cliff Tabin y Sean Carroll 2009 Deep Homology and the Origins of Evolutionary Novelty. Nature vol. 457: 818-823.
Shultz, W. y A. Dickinson 2000 Neuronal Coding of Prediction Errors. Annual Reviews of Neuroscience, vol. 23: 473-500.
Tomasello, Michael 1998 Emulation Learning and Cultural Learning. Behavioral and Brain Sciences, vol. 21: 703-704.
Wagner, Gunther, P. 1989 The Biological Homology Concept. Annual Reviews of Ecology and Syste- matics, vol. 20: 51-69.
Wagner, Gunther, P. 2001 Characters Units and Natural Kinds: an Introduction, en The Character Concept in Evolutionary Biology, G.P. Wagner (ed.). Academic Press. San Diego: 1-10.
Wagner, Gunther, P.2007 The Developmental Genetics of Homology. Natural Reviews of Genetics, vol. 8: 473-479.
Wagner, Gunther, P.2014 Homology, Genes, and Evolutionary Innovation. Princeton University Press.
Wake, David B. 2003 Homology and Homoplasy, en Keywords & Concepts in Evolutionary Developmental Biology, Hall, B. y W. Olson (eds.). President and Fellows of Harvard College.
Washburn, Sherwood Lancester 1951 The New Physical Anthropology. Transactions of the New York Academy of Sciences, vol. 13: 298-304.
Washburn, Sherwood Lancester 1973 The Promise of Primatology, en The New Physical Anthropology, Strum, S. et al. (eds.). Advances in Human Evolution Series: 43-48.
Wenzel, John W. 1992 Behavioral Homology and Phylogeny. Annual Review of Ecology and Systematics, vol. 23: 361-381.
Whiten, Andrew, et al.2009 Emulation, Imitation, Over-Imitation and the Scope of Culture for Child and Chimpanzee. Philosophical Transactions of the Royal Society B, vol. 364: 2417-2428.
Williams, David M. 2005 Homologues and Homology, Phenetics and Cladistics: 150 Years of Progres, en Milestones in Systematics, Williams, D. y P. Forey (eds.). CRC Press.
Notas