Resumen: La memoria espacial se considera un factor se- lectivo en la evolución de la inteligencia. Los estudios en animales permiten pensar que los procesos cognitivos des- criptos en humanos comparten características con los de otras especies. Se estudió el comportamiento espacial vin- culado a la búsqueda del alimento de los monos capuchinos y su capacidad de retener información sobre la ubicación y estado del mismo. Durante los seguimientos de tres grupos de Cebus nigritus silvestres se aplicó un diseño observacio- nal cualitativo que evaluó las cantidades de fruta madura e inmadura presentes en los árboles frutales consumidos por los monos en sus desplazamientos diarios. Las visitas a los árboles de frutos indicaron que los monos retienen infor- mación particular sobre la ubicación y condición de cada árbol. Los resultados obtenidos son fundamentales para la comprensión del funcionamiento de los mecanismos cogni- tivos no mediados por el lenguaje, lo cual permite ampliar los conocimientos de la base biológica del comportamien- to espacial no humano.
Palabras clave:Cebus=Sapajus nigritusCebus=Sapajus nigritus, árboles frutales árboles frutales, velocidad de acercamiento velocidad de acercamiento, visita visita.
Abstract: Spatial memory is considered a selective fac- tor in the evolution of intelligence. Animal studies lead us to believe that cognitive processes described in humans may have much in common with those of other species. We studied the foraging spatial behavior of capuchin monkeys and their ability to retain information about food location and quality. We analyzed whether individuals were able to integrate stored information about food sites and changes in the ripening of the resource. A qualitative observational design was used to monitor three groups of wild Cebus nigritus in their daily movements, evaluating the amounts of ripe and unripe fruit present in the fruit- ing trees consumed by monkeys. Visits to fruiting trees showed that monkeys remember specific information on the location and condition of each tree. These findings are essential to understand cognitive mechanisms not mediated by language, allowing a more comprehensive analysis of the biological basis of non-human spatial be- havior.
Keywords: Cebus=Sapajus nigritus, fruiting trees, speed of approach, visit.
MEMORIA ESPACIAL EN MONOS CAPUCHINOS DE ARGENTINA: UN ESTUDIO OBSERVACIONAL EN VIDA SILVESTRE
Asociación de Antropología Biológica Argentina

Esta obra está bajo una Licencia Creative Commons Atribución-NoComercial 4.0 Internacional.
Recepción: 01 Febrero 2015
Aprobación: 31 Marzo 2015
Fuente: Idea Wild. American Society of Mammalogists. Universidad Nacional de La Plata. Consejo Nacional de Investigaciones Científicas y Técnicas
La memoria espacial se considera un factor selectivo en la evolución de la inteligencia (Milton, 1981) asociado con múltiples procesos cognitivos (Fagan et al., 2013) y evolutivos (Milton, 1981). Numerosas investigaciones han determinado que los primates realizan movimientos directos hacia parches de alimento de una forma que sugiere un propósito y la utilización de un mapa espacial del ambiente (Garber y Dolins, 1996; Platt et al., 1996; Garber y Paciulli, 1997; Janson, 1998; Pochron, 2001; Cunningham y Janson, 2007; Janson y Byrne, 2007; Noser y Byrne, 2007).
En el caso de los primates que habitan selvas tropicales y subtropicales (entre los que se encuentran los primates sudamericanos), la diversidad de plantas que son fuente de alimento y la forma en que éstas se distribuyen en tiempo y espacio han sido señaladas como la mayor fuerza selectiva en el desarrollo de las complejidades cerebrales avanzadas (Milton, 1981), permitiendo el abordaje de la conducta de búsqueda del alimento como uno de los ámbitos más accesibles a un enfoque cognitivo comparado (Tujague, 2013). Si se asume que el hecho de minimizar las distancias recorridas hacia fuentes de recurso es un objetivo de la estrategia de forrajeo de primates, entendiendo al forrajeo como toda pauta de comportamiento que implique la búsqueda, localización y consumo de alimento, (Boyer et al., 2003), observar que se mueven al recurso disponible más cercano es consistente con la idea de que conocen dónde se ubican los recursos alternativos (Janson y Byrne, 2007). A su vez, los patrones de maduración de frutos de cada especie vegetal ofrecen una ventaja a aprovechar por los frugívoros si fueran capaces de memorizar estos esquemas (Janmaat et al., 2006; Anzelc, 2009). Observaciones en vida silvestre demostraron que distintas especies de primates viajaban de manera directa hacia los recursos sugiriendo que anticipaban dónde y cuánto alimento iban a encontrar: Cebus apella (Janson, 1998); Papio hamadryas cynocephalus (Pochron, 2001, 2005); Cercocebus atys atys, Lophocebus albigena johnstoni (Janmaat et al., 2006); Phitecia phitecia (Cunningham y Janson, 2007); Papio urcinus (Noser y Byrne, 2007); Ateles geoffroyi yucatanensis (Valero y Byrne, 2007), aun cuando las cantidades variaran en el tiempo como es el caso de los frutos (Janmaat et al., 2006; Janmaat et al., 2013). En este contexto, los investigadores han utilizado el aumento de la velocidad en el movimiento de los primates como evidencia de estas capacidades cognitivas de anticipación (Janson, 1998; Janmaat et al., 2006; Janson y Byrne, 2007).
En este trabajo se describe el estudio llevado a cabo por una de las autoras (MPT) cuyo objetivo fue evaluar el comportamiento espacial vinculado a la búsqueda del alimento de Cebus nigritus, considerado actualmente por Lynch Alfaro y colaboradores (2011, 2012) como Sapajus nigritus, y su capacidad de retener información sobre la ubicación y estado del mismo (memoria espacial) en el Parque Nacional Iguazú, Argentina. El mono capuchino es una especie de primate diurna con una dieta basada predominantemente en frutos y artrópodos. En Iguazú, los capuchinos consumen pulpas de frutos de al menos 81 especies diferentes de árboles, enredaderas, arbustos y epífitas. Viven en grupos polígamos multimacho-multihembra, generalmente de 7-45 individuos independientes (Janson et al., 2012), con hembras filopátricas. Es una especie que presenta habilidades cognitivas sorprendentes desde el punto de vista adap tativo en relación a la localización y adquisición de recursos (Visalberghi y Fragazsy, 1995; Janson, 1998; Visalberghi et al., 2003; Potì et al., 2005), incluyendo el uso de herramientas (Moura y Lee, 2004; Garber et al., 2012). Estos monos presentan también un conjunto de convergencias morfológicas y comportamentales con hominoideos, incluyendo un cerebro alométrico en relación a su peso corporal (semejante al que existe en el género Pan sp.), un incremento en el desarrollo de la neocorteza y un pulgar altamente móvil (Costello y Fragaszy, 1988; Fragaszy y Mitchell, 1990 en Cleveland et al., 2003; Westergaard y Kuhn, 2001 en Cleveland et al., 2003). Dadas sus conocidas capacidades cognitivas y manipulativas como también su distancia filogenética con simios y humanos, los capuchinos representan una oportunidad única para analizar las presiones grupales y ecológicas que llevan a las convergencias existentes entre Cebus sp., Pan sp. y Homo sp. (Westergaard, 1998 en Cleveland et al., 2003).
Experimentos realizados en el mismo sitio de estudio que el presente trabajo por Janson (1998, 2007) demostraron que las decisiones que los capuchinos toman durante sus movimientos diarios se vinculan tanto a las cantidades de alimento como a las distancias entre los sitios donde el mismo está ubicado. En sus experimentos Janson instaló plataformas provisionadas con frutos: mandarinas (Janson, 1998) y bananas (Janson, 1997), con un esquema en el cual en los primeros experimentos la cantidad de fruta variaba dependiendo del diseño, mientras que en los experimentos posteriores las diferentes recompensas de alimento variaban con relación a distintas distancias entre los sitios de plataformas experimentales. El grupo estudiado se desplazó en línea recta y con mayor frecuencia hacia plataformas más cercanas siendo la cantidad de comida en las plataformas un predictor de la elección a realizar, pero también la distancia entre sitios en relación a las cantidades de comida: los monos visitaban sitios de baja recompensa en el camino hacia sitios más alejados pero con mayor recompensa sólo en los casos en los que la distancia hacia el sitio de baja recompensa no los obligaba a desviarse significativamente de la ruta más directa hacia el sitio de recompensa alta.
Hasta la actualidad son pocos los estudios en los que se ha encontrado evidencia de que los primates son exitosos diferenciando entre determinados árboles frutales de una misma y de varias especies en su estado de maduración además de en cantidad (Janmaat et al., 2006, 2012; Cunningham y Janson, 2007). Janmaat y colaboradores (2006) estudiaron los movimientos de dos grupos silvestres de Cercocebus atys atys y Lophocebus albigena johnstoni en sus recorridos diarios visitando árboles de fruta previamente seleccionados por los investigadores. Por medio de la estimación de las cantidades de fruta madura e inmadura de los árboles, y analizando los patrones de visita y la velocidad con la cual los grupos se acercaban a los mismos. Los investigadores concluyeron que los monos utilizaban información sobre las visitas previas para anticipar tanto la cantidad como el estado de los frutos en las visitas futuras. En otro estudio sobre los desplazamientos diarios de un grupo de Pithecia pithecia, Cunningham y Janson (2007) encontraron que los monos utilizaban una combinación de conocimientos obtenidos del uso repetido de su territorio junto con información sobre la ubicación y condición de los recursos, tomándose un mayor tiempo para regresar a árboles de frutos que estuvieran inmaduros. Por último, en un estudio reciente, Janmaat y colaboradores (2012) investigaron la capacidad de Lophocebus albigena johnstoni para utilizar una estrategia de inspección de los árboles basada en la fructificación sincronizada de los mismos. Analizando los patrones de inspección de los árboles de una especie en particular en función de las diferencias de sincronización en tiempo y espacio de la misma al momento de fructificar, encontraron que los monos inspeccionaban más árboles durante la temporada de fructificación, indicando que utilizaban los patrones de sincronización de los árboles como indicador de la presencia de frutos.
Considerando los antecedentes presentados, las hipótesis (H) y predicciones (P) asociadas del presente trabajo fueron las siguientes: (a) H1: Los capuchinos retienen la información espacial aprendida acerca de la presencia de árboles que periódicamente fructifiquen dentro del territorio del grupo, P1: el grupo focal visitará los árboles de manera periódica, P2: los patrones de visitas dependerán de la presencia de frutos en los mismos y (b) H2: Los acerca mientos a árboles frutales están determinados por la presencia de frutos en distintas cantidades y estados de maduración, P1: el grupo focal aumentará su velocidad a medida que se aproxima a un árbol visitado con anterioridad y que aun presenta frutos, P2: el grupo focal visitará con mayor frecuencia y a mayor velocidad árboles con mayor cantidad de frutos en estado maduro.
Las observaciones se desarrollaron en el Parque Nacional Iguazú, ubicado en la provincia de Misiones, Argentina (25º40’43”S, 54º26’57”O). El área comprende una porción de selva subtropical semidecidua caracterizada por clima subtropical húmedo, carente de estación seca predecible (Brown y Zunino, 1990). La producción de frutos es estacional, con el pico de baja producción entre junio y agosto y el de máxima entre octubre y noviembre (Janson y Di Bitetti, 1997) y con periodos de maduración cortos (Placci et al., 1994). Se realizaron seguimientos diarios continuos desde el amanecer al anochecer de tres grupos de Cebus nigritus en estado silvestre dentro del parque nacional durante los meses de octubre a diciembre y marzo a abril entre los años 2008 y 2011(Tabla 1).
Años de estudio de cada grupo, días de seguimiento, tamaño de los grupos y composición por sexo y edad
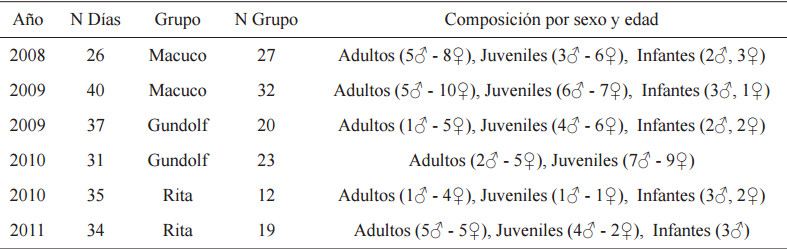
N Días total de días de seguimientoN grupo tamaño de los grupos
La metodología aplicada consistió en comparar datos de observación con las predicciones de un modelo cualitativo en el que el observador selecciona previamente árboles de determinadas especies consumidas por los monos y evalúa los mismos en función de la ausencia/presencia de frutos y su correspondiente estado madurativo. Se utilizó una modificación del método de árboles focales desarrollado por Janmaat et al. (2006) para analizar la capacidad de los grupos de estudio de visitar y re-visitar los frutales dentro de su territorio, en función de su ubicación espacial y cantidad de alimento disponible. Previo al inicio de las observaciones de comportamiento se seleccionaron 25 especies de árboles frutales y 2 especies de enredaderas cuyos frutos eran consumidos por los monos y no presentaban señales de su estado de maduración (olfativas ni visuales) a larga distancia (Tabla 2). Cuando los monos se encontraban en las cercanías de un árbol focal (AF), un observador se adelantaba al mismo y registraba la presencia de frutos en el mismo según las siguientes categorías: 0 (0%), 1 (1-25%), 2 (26-50%), 3 (51-75%), 4 (76-100%) (Janmaat et al., 2006). Esto se realizó dividiendo visualmente la copa en cuatro cuadrantes siguiendo como ejes a los puntos cardinales y realizando una estimación visual promedio del porcentaje de ramas que contenían fruta por cuadrante, desde la base al final de las mismas. Un árbol categoría 4 tendría entonces todas las ramas de todos los cuadrantes con frutos. Cuando un árbol no presentaba ramas en alguno de los cuatro cuadrantes se contó a ese cuadrante como con 0% de fruta. A su vez, siempre que fue posible distinguir la presencia de fruta madura de la inmadura se contaron las ramas por separado y se asignó un porcentaje promedio a las categorías finales. Dado que los árboles pueden presentar diferencias en los estados de maduración y cantidades de fruta dentro de un mismo individuo como causa de la diferente influencia del sol y la temperatura (Houle et al., 2007), esta metodología de estimación permite controlar esta variable.
Listado total de especies frutales consumidas (N=27) por los grupos de studio

ENR enredadera
e cada AF se tomaron los siguientes datos botánicos (Dallmeier, 1992): 1) DAP, diámetro a la altura del pecho en centímetros, 2) altura de la copa en metros y 3) diámetro de la copa en metros. Estas dos últimas medidas se utilizaron para calcular el volumen de la copa. De acuerdo con Janmaat et al. (2006) se incluyeron como AF aquellos árboles con DAP y diámetro de
De cada AF se tomaron los siguientes datos botánicos (Dallmeier, 1992): 1) DAP, diámetro a la altura del pecho en centímetros, 2) altura de la copa en metros y 3) diámetro de la copa en metros. Estas dos últimas medidas se utilizaron para calcular el volumen de la copa. De acuerdo con Janmaat et al. (2006) se incluyeron como AF aquellos árboles con DAP y diámetro de copa mayor que el más pequeño observado con frutos durante el período de muestreo. Se excluyeron del muestreo los árboles que llevaban lianas u otro tipo de fruto que no perteneciera al AF.
Se consideró una distancia mínima de separación entre árboles focales calculada como el doble de la dispersión longitudinal del grupo durante el forrajeo lento. La dispersión se estimó en función del número de individuos de cada grupo a partir de estudios previos (Janson y Di Bitetti, 1997; Janson, 1998). Se definió un radio crítico (RC) alrededor de cada árbol, como la mitad de la distancia mínima entre árboles focales. A esta distancia se consideró que el grupo era incapaz de detectar el árbol. Durante el seguimiento de los grupos de monos en sus visitas a los AF se registraron hora y ubicación del grupo en coordenadas UTM, antes de que los monos entraran al RC, en el momento en que ingresaban al mismo y a la mitad entre el RC y el AF. Estos datos de ubicación permitieron estimar las siguientes velocidades de movimiento (Noser y Byrne, 2007), medidas en metros por minuto (m/min): a) Velocidad pre radio crítico (V Pre RC) con la que el grupo se movía antes de entrar al radio crítico, b) Velocidad post radio crítico (V Post RC) con la que el grupo se movía inmediatamente luego de entrar al radio crítico y c) Velocidad media radio crítico (V Media RC) con la que el grupo se acercaba al AF, estimada a la distancia media entre el árbol focal y el límite del radio crítico.
Finalmente se definió a una visita como cualquier ingreso a un AF, con o sin consumo de fruta, mientras que un acercamiento o entrada a un RC se entendió como cualquier instancia en la que el grupo entrara a la distancia alrededor de un AF dentro de la cual no es posible aun visualizar el árbol. Luego del ingreso a un RC, el grupo pudo visitar o no el árbol.
Para el análisis de los patrones de visitas a árboles con y sin fruta se utilizaron el test de Chi cuadrado con corrección de continuidad de Yates (Yates, 1934) y el test de Phi para estimar el tamaño del efecto (Trusty et al., 2004), donde 0 indica ausencia de asociación y 1, asociación fuerte.
Para evaluar las diferencias entre las velocidades de acercamientos y visitas a los árboles se utilizaron test de Student para muestras pareadas y para muestras independientes y test de probabilidad combinada de Fisher para muestras con N bajo (Fisher, 1932). Para todos los test de Student se calculó el tamaño del efecto (effect size) mediante el método de Cohen (d de Cohen) (Nakawaga y Cuthill, 2007; Morales Vallejo, 2012) como análisis complementario al análisis de diferencias entre promedios. Cohen (1988 en Nakawaga y Cuthill, 2007) ha propuesto valores convencionales como puntos de referencia para interpretar el valor de d: bajos (d=0.2 ó menores), medios o moderados (d=0.5) y altos (d=0.8 ó mayores). Los datos de velocidad no se ajustaron a una distribución normal por lo que se les aplicó transformación logarítmica para poder utilizar test paramétricos.
Se compararon las velocidades Media RC de visita según los porcentajes de frutos separados en las siguientes categorías: árboles con más del 50% de fruta (madura e inmadura) presente y aquellos con el 50% o menos, árboles con más del 25% de fruta (madura e inmadura) presente y aquellos con el 25% o menos, árboles con más del 50% de fruta madura presente y aquellos con el 50% o menos, árboles con más del 25% de fruta madura presente y aquellos con el 25% o menos.
Finalmente se realizaron correlaciones de Spearman entre las velocidades y las medidas de los AF (volumen de la copa y DAP), para evaluar si los monos recordaban otras características vinculadas al tamaño del árbol además de las categorías de frutos, aumentando la velocidad de desplazamiento en función de las mismas. Se descartaron del análisis las correlaciones con un N menor a 20 (Packard y Boardman, 2008).
Todas las investigaciones realizadas en este manuscrito adhirieron a los principios éticos para el tratamiento de primates no humanos de la American Society of Primatologists, a los protocolos aprobados por el Animal Care and Use Committee y a los requerimientos legales de la Administración de Parques Nacionales de Argentina.
El promedio de árboles visitados por día y el número de especies varió según el grupo (Tabla 3).
Aquellas visitas a AF en las que se registró la presencia de otros frugívoros (pájaros y/o coatíes) en el árbol antes de la llegada del grupo fueron excluidos del análisis a fin de controlar la posibilidad de que el grupo se acercara al árbol atraído por la audición de otros individuos alimentándose y no por el registro de la presencia de frutos en el mismo.
Promedio de árboles parches de fruta visitados por día ± desvío estándar, radio crítico y distancia mínima entre árboles focales en metros, número de especies consumidas (N sp.) para cada grupo en cada período de muestreo y sus áreas de acción en hectáreas
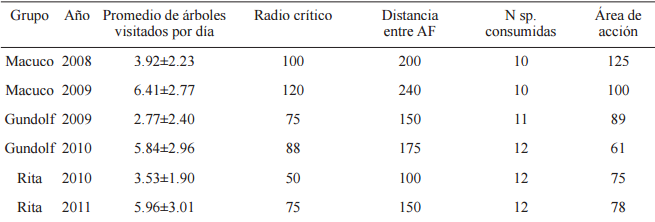
Se evaluó si los monos, habiendo entrado al RC de un árbol, seguían o no desplazándose hacia el árbol según si tenía o no frutos, hallándose para 4 de las 6 muestras (Macuco 2009; Gundolf 2009-2010 y Rita 2010), una mayor probabilidad de visitar un árbol cuando tenía frutos que cuando no tenía frutos (Macuco 2009: χ²=1846.76, p≤0.00, Phi=0.12, p=0.03; Gundolf 2009: χ²=10.55, p≤0.00, Phi=0.32, p≤0.00; Gundolf 2010: χ²=26.41, p≤0.00, Phi=0.38, p≤0.00; Rita 2010: χ²=59.72, p≤0.00, Phi=0.58, p≤ 0.00). Puede observarse que los tamaños del efecto fueron débiles pero no nulos, excepto en el caso de Rita 2010 donde la asociación fue elevada. Para el caso de Macuco 2008 la asociación no fue significativa pero el valor de Phi reveló una asociación fuerte entre las variables (χ²=2.13, p=0.14; Phi=0.68, p≤0.00). El grupo Rita 2011 no fue incluido en el análisis dado que de un total de 204 entradas a RC, sólo 3 fueron a árboles sin frutos.
En la muestra correspondiente a Gundolf 2009 existió una mayor cantidad de entradas a RC de árboles sin frutos que con frutos y la misma cantidad de visitas en ambas categorías (N visitas sin fruta=32; N visitas con fruta=33), sugiriendo que los monos podrían estar monitoreando en los árboles sin frutos la presencia de larvas de Homoptera presentes durante ese período de muestreo.
Para evaluar si los monos, una vez entrado el RC (y aun sin poder visualizar el árbol) tenían una expectativa de lo que iban a encontrar más adelante, se compararon los promedios de las velocidades Post RC sin visita y con visita a los AF, observando diferencias significativas (χ²F =21.42, p=0.04, gl=12) y mostrando que los monos, una vez entrado al RC, lo hacían a mayor velocidad cuando tal entrada terminaba en una visita.
Para analizar si los grupos incrementaban su velocidad en dirección hacia el AF cuando éste aún se encontraba fuera de su vista se compararon las velocidades Pre, Post y Media RC entre sí. El promedio de las V Pre RC fue menor que las Post RC, y a su vez el promedio de la V Post RC fue menor que el promedio de las V Media RC en todos los casos (Tabla 4).
Promedio (M) de las velocidades en metros/minuto, desvíos estándar (DS), intervalos de confianza (IC 95%) y número (N) de visitas Pre-RC, Post-RC y Medio-RC de cada grupo por período de muestreo
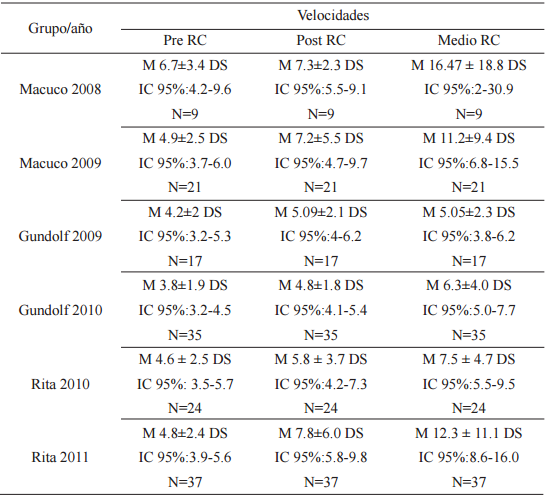
Se encontraron diferencias entre las velocidades Pre RC y Post RC de las visitas (χ²F =85.95, p≤0.00, gl=12) y entre las velocidades Post RC y Media RC (χ²F =78.61, p≤0.00, gl=12).
Se procedió a evaluar si los monos poseían algún tipo de registro sobre la cantidad y estado de maduración de los frutos en el árbol por visitar. Para esto se compararon las V Media RC de visita a los árboles según diferentes porcentajes de fruta presente (madura e inmadura juntas) y de fruta madura únicamente.
No se encontraron diferencias entre las V Media RC de acercamiento a árboles con menos o más del 50% de fruta, madura e inmadura, presente (test de probabilidad combinada de Fisher para muestras independientes, χ²F =7.07, p=0.71, gl=10). Ninguno de los grupos presentó diferencias significativas en los test de Student independientes sobre los que se basó el análisis y los cálculos de tamaño del efecto demostraron que no existió una diferencia considerable, excepto para las muestras de Macuco 2008 y Rita 2010, donde el tamaño del efecto fue moderado, pero los promedios mayores correspondieron a los árboles con 50% o menos de fruta presente (Tabla 5).
Test de Student para muestras independientes para las velocidades Media RC sin transformar y log transformadas (metros/minuto) de las visitas a árboles focales con más del 50% versus 50% o menos de fruta (madura + inmadura) presente, y a árboles focales más del 25% versus 25 % o menos de fruta (madura + inmadura) presente (*) Promedios, desvíos estándar, y valores del d de Cohen
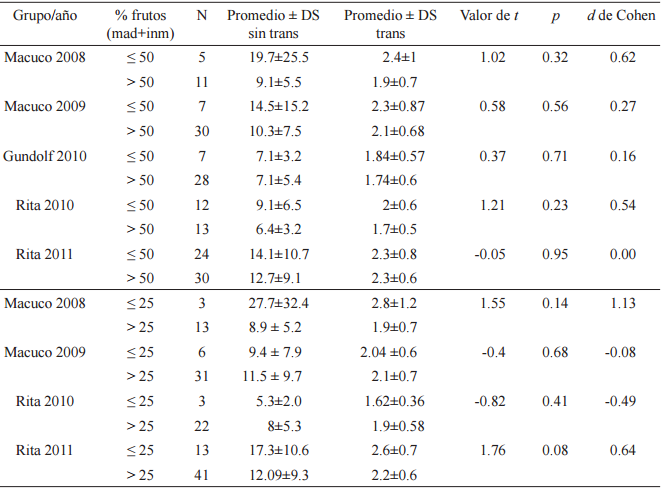
* Las muestras correspondientes al grupo Gundolf en los períodos 2009 y 2010 fueron excluidos del análisis por datos insuficientes para la categoría del 25%, y el período 2009 para la categoría del 50%.
Tampoco se encontraron diferencias entre las V Media RC para árboles con más o menos del 25% de fruta presente (test de probabilidad combinada de Fisher para muestras independientes, χ²F =11.39, p=0.18, gl=8). Ninguno de los grupos presentó diferencias significativas en los test de Student independientes sobre los que se basó el análisis y los cálculos de tamaño del efecto demostraron que existió una diferencia baja a moderada (Rita 2010 y 2011) y sólo alta en el caso de Macuco 2008 donde el N del análisis fue bajo (Tabla 5). Puede observarse que en algunos casos el valor del d de Cohen es negativo, indicando que el valor del segundo promedio de velocidad utilizado en el análisis (en este caso árboles con más del 25% de fruta presente) fue mayor. Nuevamente se registraron velocidades mayores para los árboles con 25% o menos de fruta presente, pero es importante notar que algunas de estas muestras tuvieron N bajos.
En cuanto al análisis de la presencia exclusiva de fruta madura, no se encontraron diferencias entre las V Media RC para árboles con más o menos del 50% de fruta madura presente (test de probabilidad combinada de Fisher para muestras independientes, χ²F =4.79, p=0.57, gl=6). Ninguno de los grupos presentó diferencias significativas en los test de Student independientes sobre los que se basó el análisis, y los cálculos de tamaño del efecto demostraron que existió una diferencia baja a moderada (Tabla 6). Puede observarse que en algunos casos el valor del d de Cohen es negativo indicando nuevamente que el promedio dos (en este caso árboles con más del 50% de fruta madura presente), fue mayor. A pesar de esto, en el caso en que se observó una magnitud del efecto más elevada (muestra correspondiente a Rita 2011) el mayor promedio se observó para las visitas a árboles con 50% o menos de fruta madura presente.
Test de Student para muestras independientes para las velocidades Media RC sin transformar y log transformadas (metros/minuto) de las visitas a árboles focales con más del 50% versus 50% o menos de fruta madura presente, y a árboles focales más del 25% versus 25 % o menos de fruta madura presente. Promedios, desvíos estándar, y valores del d de Cohen(*)
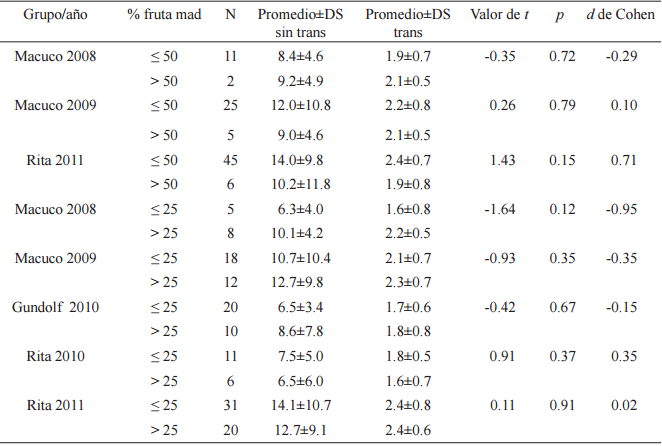
* Los grupos Gundolf (2009 y 2010) y Rita (2010) fueron excluidos del análisis por datos insuficientes.
Por último, tampoco se encontraron diferencias entre las V Media RC para árboles con más o menos del 25% de fruta madura presente (test de probabilidad combinada de Fisher para muestras independientes, χ²F =12.74, p=0.23, gl=10). Ninguno de los grupos presentó diferencias significativas en los test de Student independientes sobre los que se basó el análisis y los cálculos de tamaño del efecto demostraron que existió una diferencia baja a moderada (Tabla 6), excepto en el caso de Macuco 2008 donde el tamaño del efecto fue elevado, siendo mayores las velocidades a árboles con más del 25% de fruta, aunque nuevamente el N de esta comparación fue bajo.
A diferencia de los análisis anteriores, en el caso de las V Media RC para árboles con más o menos del 25% de fruta madura presente los promedios de velocidades fueron mayores a árboles con más del 25% de fruta madura presente en la mayoría de los casos (excepto Rita 2010 y 2011).
El análisis de las velocidades en relación al volumen de la copa reveló una correlación significativa de la V Pre RC para la muestra correspondiente a Rita 2011, y de la V Post RC para Macuco 2008. En cuanto al DAP, presentó una correlación significativa con la V Post RC para una sola de las 6 muestras analizadas, Rita 2011. El resto de las velocidades no se correlacionaron con el volumen de la copa o el DAP (Tabla 7).
Correlaciones de Spearman entre las variables Velocidad Pre Radio Crítico (V Pre RC),Post Radio Crítico (V Post RC) y Media Radio Crítico (V Media RC) en metros/minuto con el volumen de copa en metros cúbicos (VOL Copa) y diámetro a la altura del pecho en centímetros (DAP)(*)
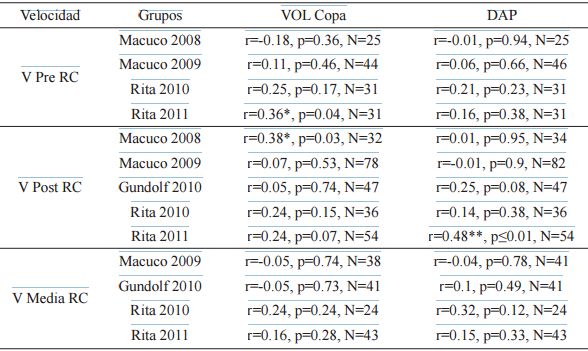
(*) Las muestras correspondientes a Macuco 2008 y Gundolf 2010 para las V Pre RC y Gundolf 2009 para todas las velocidades, fueron excluidas del análisis por presentar N menores a 20 ** diferencias significativas p<0.05 y 0.01 respectivamente><0.05 y 0.01 respectivamente
Los grupos estudiados visitaron los árboles focales de manera reiterada mientras los mismos tuvieron frutos, dirigiéndose a mayor velocidad cuando su objetivo final era una visita que cuando no lo era y aumentando gradualmente la velocidad a medida que se acercaban al árbol (como puede observarse en la tabla 4). Esto indica que existió una expectativa del recurso a encontrar y por lo tanto, un registro previo basado en las cantidades y condiciones de los frutos y no en otras características de los árboles tales como el volumen de la copa o el diámetro del tronco. Dados los recaudos tomados en cuanto a distancias de detección, puede considerarse que los monos no utilizaron señales visuales ni olfativas durante este proceso. Asimismo, vale considerar que los primates no humanos y humanos presentan el mismo rango de performance olfatoria que les impide detectar el olor de la fruta a más de 20cm (Janmaat et al., 2006).
Puede reafirmarse entonces que la velocidad es una buena medida de la expectativa del grupo, tal como se había señalado en estudios previos en esta especie (Janson y Di Bitetti, 1997) y en otras: Papio ursinus (Noser y Byrne, 2007), Papio hamadryas cynocephalus (Pochron, 2001, 2005).
En cuanto a la velocidad como medida para estimar la capacidad de los monos de diferenciar entre árboles con fruta madura e inmadura, la predicción de que los monos viajaban más rápido a árboles con una mayor cantidad de fruta madura presente no tiene una base teórica conocida, por lo cual la ausencia de este patrón en los datos presentados no necesariamente indica que los mismos no conocen el estado de fructificación del árbol. Podría haber ocurrido que una vez dentro del RC y no percibir señales auditivas de otros frugívoros compitiendo por el recurso en el sitio (coatíes, pájaros), viajar a mayor velocidad hacia el recurso que tuviera más frutos maduros podría no ser necesario (a pesar del ya mencionado incremento en la velocidad que existió a medida que se acercan al árbol). A su vez, si bien los test estadísticos no encontraron diferencias significativas, existieron tamaños del efecto elevados y velocidades mayores en dirección a árboles con mayor cantidad de fruta madura para algunas muestras. Resultados preliminares del análisis de los intervalos de re-visita a árboles según las cantidades de fruta en diferentes estados de maduración sugirieron que los monos capuchinos en Iguazú usan información sobre la cantidad pero también sobre el estado de maduración de los frutos al decidir si visitan un árbol y con qué frecuencia (Tujague y Janson, 2015). El tiempo transcurrido hasta la próxima visita decreció independientemente con relación al aumento de las cantidades de fruta madura e inmadura presente en la copa, indicando que los monos eran capaces de anticipar la maduración de los frutos presentes en la última visita. Entonces, o bien la velocidad no es un buen indicador de que los monos recuerdan el estado de maduración de los frutos en el árbol, o tal vez el ambiente ofrecía a los individuos suficientes nutrientes como para no necesitar visitar los árboles de manera más rápida, teniendo en cuenta que los objetivos nutricionales son determinantes de la selección de la dieta por parte de los primates (para una revisión sobre ecología nutricional ver Felton et al., 2009). La diferencia nutricional entre los frutos no fue una variable considerada en este trabajo, siendo interesante su incorporación para estudios futuros.
En el presente trabajo se pusieron de relieve novedosas cuestiones del comportamiento espacial de los sujetos estudiados, como es la capacidad de recordar características vinculadas a la cantidad y el estado de maduración del alimento distribuido en el ambiente, lo cual constituye un ejemplo de flexibilidad cognitiva y plasticidad. A pesar del carácter puramente observacional del diseño, las implicancias de considerar las variables “cantidad” y “estado de maduración” de los recursos es importante, porque permite analizar cómo la variación espacio-temporal del comportamiento de los primates se asocia al recuerdo de frutos disponibles para ser consumidos en los árboles. Esa información espacio-temporal aprendida en determinado momento tendrá valor en un tiempo posterior, sugiriendo que la posibilidad de conseguir alimento dependerá de su propio comportamiento pasado (Janson, 2000).
Estos aportes permiten conocer con mayor detalle las características de base de una especie ampliamente estudiada en trabajos de cognición comparada. Asimismo, los hallazgos descriptos son fundamentales para la comprensión del funcionamiento de los mecanismos cognitivos no mediados por el lenguaje, aportando elementos valiosos para el estudio de la base biológica del comportamiento espacial de los Primates y apoyando la idea de que en el estudio de primates no humanos pueden encontrarse las características que nos asemejan más allá de nuestra diversidad genómica y cultural.
A la Administración de Parques Nacionales (APN) por los permisos para llevar adelante el trabajo de campo en Iguazú. El mismo fue posible gracias a la asistencia de Alex Alcocer Aldunate, Ana Alvarez, María Celia Baldovino, María Verónica Cifre, Rocío Fernández Quintero, Emanuel Galetto, Elizabeth Gonzáles Valentín, Daniel Lambruschi, Luis Molina, Juan Pablo Peretti, Noelia Rivas, Taylor Rubin, Clara Scarry y Fermino Silva.
Dra. María Paula Tujague. Instituto de Biología Subtropical - UNaM, CONICET. Bertoni 85. 3370 Puerto Iguazú. Misiones. Argentina
Años de estudio de cada grupo, días de seguimiento, tamaño de los grupos y composición por sexo y edad
N Días total de días de seguimientoN grupo tamaño de los grupos
Listado total de especies frutales consumidas (N=27) por los grupos de studio
ENR enredadera
Promedio de árboles parches de fruta visitados por día ± desvío estándar, radio crítico y distancia mínima entre árboles focales en metros, número de especies consumidas (N sp.) para cada grupo en cada período de muestreo y sus áreas de acción en hectáreas
Promedio (M) de las velocidades en metros/minuto, desvíos estándar (DS), intervalos de confianza (IC 95%) y número (N) de visitas Pre-RC, Post-RC y Medio-RC de cada grupo por período de muestreo
Test de Student para muestras independientes para las velocidades Media RC sin transformar y log transformadas (metros/minuto) de las visitas a árboles focales con más del 50% versus 50% o menos de fruta (madura + inmadura) presente, y a árboles focales más del 25% versus 25 % o menos de fruta (madura + inmadura) presente (*) Promedios, desvíos estándar, y valores del d de Cohen
* Las muestras correspondientes al grupo Gundolf en los períodos 2009 y 2010 fueron excluidos del análisis por datos insuficientes para la categoría del 25%, y el período 2009 para la categoría del 50%.
Test de Student para muestras independientes para las velocidades Media RC sin transformar y log transformadas (metros/minuto) de las visitas a árboles focales con más del 50% versus 50% o menos de fruta madura presente, y a árboles focales más del 25% versus 25 % o menos de fruta madura presente. Promedios, desvíos estándar, y valores del d de Cohen(*)
* Los grupos Gundolf (2009 y 2010) y Rita (2010) fueron excluidos del análisis por datos insuficientes.
Correlaciones de Spearman entre las variables Velocidad Pre Radio Crítico (V Pre RC),Post Radio Crítico (V Post RC) y Media Radio Crítico (V Media RC) en metros/minuto con el volumen de copa en metros cúbicos (VOL Copa) y diámetro a la altura del pecho en centímetros (DAP)(*)
(*) Las muestras correspondientes a Macuco 2008 y Gundolf 2010 para las V Pre RC y Gundolf 2009 para todas las velocidades, fueron excluidas del análisis por presentar N menores a 20 ** diferencias significativas p<0.05 y 0.01 respectivamente><0.05 y 0.01 respectivamente
