Resumen: El objetivo fue analizar en animales con retardo prenatal de crecimiento (RPC) el efecto de la subnutrición proteico-calórica lactacional y postlactacional sobre la morfología craneofacial, particularizando en el crecimiento de los componentes funcionales neural y facial. Ratas Wistar fueron divididas en los grupos: Control, RPC (inducido por ligamiento parcial de ambas arterias uterinas el día 15 de gestación) y Sham-operado (con igual técnica quirúrgica que RPC aunque sin ligamiento de las arterias). A su vez, el grupo RPC se dividió en: (a) crías lactantes de madres con nutrición normal y a partir del destete alimentadas ad-libitum y (b) crías lactantes de madres con restricción alimentaria del 25% y a partir del destete alimentadas con el 50% de lo consumido por un animal control. Se tomaron radiografías a las edades 1, 21, 42, 63 y 84 y se midieron longitud, ancho y altura de los componentes neural y facial. Se calcularon los índices volumétricos neural y facial y morfométrico neurofacial. Se aplicaron ANOVA y pruebas post-hoc y se calcularon diferencias porcentuales entre medias. Los resultados permitieron concluir que el estrés primario ocurrido durante la vida intrauterina resulta crítico en lo inmediato y en la vida postnatal, ya que aun mediando normonutrición postnatal el retardo de crecimiento perdura. Además, cuando al estrés prenatal le continúa restricción nutricional postnatal los efectos adversos son aditivos provocando retardo del crecimiento aún mayor. Finalmente, mientras que el componente neural es más resistente a las deficiencias nutricionales, el facial presenta mayor plasticidad, hecho que se evidencia en cambios de forma.
Palabras clave:crecimiento craneofacialcrecimiento craneofacial, desnutrición pre y postnatal desnutrición pre y postnatal, craneometría funcional craneometría funcional .
Abstract: The aim of the study was to analyze the effect of protein-calorie malnutrition during lactation and post-lactation on craniofacial morphology in intrauterine growth-retarded (IUGR) animals, particularly in the neural and facial functional components. Wistar rats were divided into the following groups: Control, IUGR (induced by partial bending of both uterine vessels at day 15 of gestation), and sham-operated (with the same surgical technique as IUGR, but without vessel bending). The IUGR group was further divided into (a) nursing pups of mothers with normal nutrition and fed ad-libitum at weaning, and (b) nursing pups of mothers with 25% food restriction and fed with 50% of the food ingested by controls at weaning. Radiographs were taken at 1, 21, 42, 63, and 84 days. Neural and facial length, width and height were measured, and neural and facial volumetric and morphometric indices were calculated. ANOVA and post-hoc tests were applied, and percentage differences between means were determined. Results showed that intrauterine stress is critical during early and postnatal life, since even when postnatal nutrition is normal, growth retardation persists. Furthermore, when prenatal stress is followed by postnatal nutritional restriction, adverse effects are additive and cause even greater growth retardation. Finally, while the neural component is more resistant to nutritional deficiencies, the facial component has greater plasticity, as reflected in the shape changes observed.
Keywords: craniofacial growth, prenatal and postnatal undernutrition, functional craniometry.
EFECTO DEL RETARDO PRENATAL DE CRECIMIENTO Y LA SUBNUTRICIÓN POSTNATAL EN EL CRECIMIENTO CRANEOFACIAL
Asociación de Antropología Biológica Argentina

Esta obra está bajo una Licencia Creative Commons Atribución-NoComercial 4.0 Internacional.
Recepción: 09 Junio 2014
Aprobación: 21 Octubre 2014
Fuente: Universidad Nacional de La Plata
Nº de contrato: 11/ V203
El crecimiento y desarrollo es un proceso dinámico y continuo a través del cual los seres vivos al mismo tiempo que incrementan su tamaño corporal, maduran y adquieren progresivamente su plena capacidad funcional como resultado de la interacción entre factores genéticos y extragenéticos (Bogin, 2001). El crecimiento se manifiesta en una secuencia ordenada y similar en todos los seres humanos, sin embargo no es un proceso homogéneo, ya que los distintos tejidos y segmentos corporales presentan tasas de crecimiento diferentes que dan como resultado alometrías (Shea, 1985; 1992; Miller y German, 1999).
Cuando los niños se encuentran expuestos a factores ambientales desfavorables pueden presentar alteraciones en su patrón de crecimiento (Hernández Rodríguez, 2007; Kuzawa, 2008). Durante la infancia y la niñez la desnutrición crónica (talla baja para la edad), la desnutrición aguda severa y el retardo del crecimiento intrauterino, constituyen los principales factores de riesgo de mortalidad (Black et al., 2008). Entre las causas más frecuentes de retardo prenatal de crecimiento (RPC) figura la insuficiencia úteroplacentaria, condición que provoca disminución en la tasa de crecimiento fetal y otros efectos negativos sobre la salud, tanto a corto como a largo plazo (Clausson et al., 2001; Gomez Roig, 2002; Schwitzgebel et al., 2009). Así, estudios epidemiológicos han demostrado la relación entre la deficiencia nutricional ocurrida durante las primeras etapas de vida y las diferentes enfermedades que se manifiestan durante la adultez, principalmente aquellas relacionadas con el metabolismo de la glucosa (Godfrey y Barker, 2000; Flanagan et al., 2000), la hipertensión y el daño cardiovascular (Hales et al., 1997; Ozanne y Hales, 1999). A partir de estos estudios ha surgido la hipótesis de “programación de la vida fetal” la cual propone que la desnutrición intrauterina desencadena adaptaciones endócrinas que modifican de manera permanentemente la morfología y el metabolismo (Godfrey y Barker, 2000).
La multiplicidad de factores ambientales estresantes que provocan RPC ha dificultado alcanzar un conocimiento acabado del accionar de cada uno de ellos. Una forma de estudiar el RPC es a través de la aplicación del método experimental y el empleo de una amplia variedad de procedimientos. En algunos se han aplicado restricciones alimentarias a las madres gestantes entre las que se incluyen la reducción calórica media (15%), moderada (50%) o severa (70%) (Pucciarelli y Oyhenart, 1987a; Vuguin, 2007) y en otros se ha realizado el ligamiento de las arterias uterinas en ratas gestantes a fin de reducir el flujo sanguíneo materno-fetal simulando la insuficiencia útero-placentaria ocurrida en humanos (Wigglesworth, 1964; Oyhenart et al., 1998; Huizinga et al., 2004, Vuguin, 2007). Este último procedimiento involucra un conjunto de condiciones que provocan restricción del aporte de glucosa y aminoácidos y alteraciones fisiológicas que afectan la homeostasis fetal. Resultados obtenidos empleando este modelo informaron disminución del peso y tamaño corporal al nacimiento, retardo en el crecimiento craneano y postcraneano con efecto alométrico, modificaciones de la composición corporal e inhibición del dimorfismo sexual (Oyhenart et al., 2003; Quintero et al., 2005; Vuguin, 2007; Gonzalez et al., 2011a).
El período postnatal es una oportunidad para recuperar cualquier déficit de crecimiento. No obstante, es conocido que las posibilidades de recuperación dependen tanto de la intensidad con que actuó la restricción prenatal (Houdijk et al., 2000; Oyhenart et al., 2008) como de la variación en las condiciones ambientales en que los organismos crecen (Gonzalez et al., 2011b). Al respecto, si bien se han realizado estudios aplicando modelos de restricción alimentaria en los períodos lactacional y/o postlactacional (Pucciarelli y Oyhenart, 1987b; Dressino y Pucciarelli, 1999; Fernándes et al., 2008), aún no se ha profundizado en el análisis del crecimiento craneano cuando al estrés intrauterino le continúan deficiencias nutricionales postnatales.
El objetivo del presente estudio fue analizar en animales con retardo prenatal de crecimiento el efecto de la subnutrición proteico-calórica lactacional y postlactacional sobre la morfología craneofacial, particularizando en el crecimiento de los componentes funcionales neural y facial.
Se realizó un estudio longitudinal empleando ratas de la especie Rattus norvegicus albinus (cepa Wistar) provenientes del bioterio del Instituto de Genética Veterinaria (IGEVET) de la Facultad de Ciencias Veterinarias (UNLP-CCT-La Plata CONICET).
Machos y hembras fueron colocados en apareo diariamente (entre las 17 y 18 horas). En las primeras horas del día siguiente, se realizó un examen exfoliativo vaginal para determinar la presencia de espermatozoides que, en caso positivo, fue considerado como inicio de la gestación. Las hembras preñadas fueron separadas en cajas individuales y mantenidas durante la gestación con dieta stock y agua ad libitum.
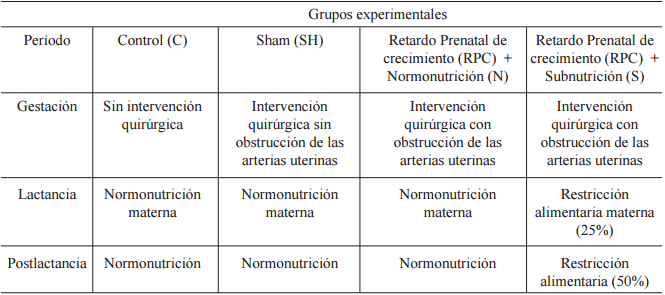
Se conformaron los siguientes grupos cuyo tamaño muestral (n) no varió en todo el experimento (Tabla 1):
Control (C) (n:16 machos, 15 hembras): crías nacidas de madres con nutrición normal y sin ningún tipo de tratamiento.
Retardo prenatal de crecimiento (RPC): crías nacidas de madres con nutrición normal a las que se les realizó una obstrucción parcial de las arterias uterinas el día 15 de gestación. Esta técnica introducida por Wigglesworth (1964) y posteriormente modificada por Oyhenart et al. (1998), se realizó mediante laparotomía abdominal bajo anestesia, con retracción de ambos cuernos uterinos y ligamiento simple con compresión parcial en el extremo inferior de cada arteria. Una vez ligados, los cuernos uterinos fueron vueltos a la cavidad abdominal y se procedió a suturar por planos. Al nacimiento este grupo fue subdividido en:
b1) RPC+N (n:15 machos, 15 hembras): crías lactantes de madres con nutricional normal y a partir del destete alimentadas ad-libitum.
b2) RPC+S (n:15 machos, 15 hembras): crías lactantes de madres con restricción alimentaria del 25% y a partir del destete alimentadas con el 50% de lo consumido por un animal de la misma edad y sexo.
Sham-operado (SH) (n:15 machos, 15 hembras): a las madres se les realizó el mismo procedimiento quirúrgico que al grupo RPC, pero en este caso las arterias uterinas no fueron obstruidas con el propósito de aislar el posible efecto de la técnica. La alimentación fue igual al grupo C.
Grupos experimentales de acuerdo al tratamiento aplicado
Los animales fueron radiografiados a los 1, 21, 42, 63 y 84 días de edad, en normas lateral y dorsal utilizando un equipo de rayos X Heliophos 4 240 MA, 125 KV. Sobre cada placa radiográfica se realizó el relevamiento métrico siguiendo los criterios de la Teoría Craneana Funcional que considera la estructura craneofacial como un complejo formado por distintos componentes (mayores y menores) los cuales tienen una función determinada (Moss y Young, 1960).
Se relevaron las siguientes mediciones lineales de los componentes craneanos funcionales mayores, empleando un calibre digital Tesa CAL IP 67 (0.01mm de precisión) (Fig. 1):
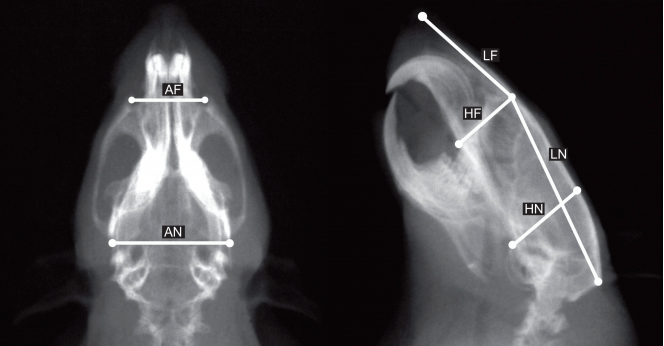
Fig. 1
Radiografía en vista dorsal (izquierda) y lateral (derecha) del cráneo de la rata mostrando las mediciones empleadas: LN:longitud neural, AN: ancho neural, HN:altura neural, LF: longitud facial, AF:ancho facial, HF:altura facial
Componente neural: Longitud: distancia Nasion-Opistocráneo (LN); Ancho: distancia Eurion-Eurion (AN); Altura: distancia entre la sincondrosis basioccipital y el Vértex (HN).
Componente facial: Longitud: distancia Prostion-Nasion (LF); Ancho: distancia máxima entre los puntos más anteriores de los arcos cigomáticos (AF); Altura: distancia entre el piso del paladar -por delante del primer molar superior- y el Nasion (HF).
Con estas variables se calcularon los índices craneanos volumétricos neural y facial (estimadores de tamaño) y morfométrico neurofacial (estimador del crecimiento relativo de ambos componentes mayores), empleando las siguientes fórmulas:
Índice Volumétrico Neural (IVN)=3 √LN x AN x HN Índice Volumétrico Facial (IVF)=3 √LF x AF x HF Índice Morfométrico Neurofacial (IMNF)= (IVN/IVF)
El proyecto de investigación fue evaluado y aprobado por la Secretaría de Ciencia y Técnica de la Universidad Nacional de La Plata. El trabajo experimental se realizó siguiendo las recomendaciones nacionales e internacionales contenidas en las normas de la Asociación Americana de Medicina Veterinaria, la Ley Nacional 14346 de la República Argentina y de los Derechos Internacionales de los Animales (ONUUNESCO).
Se calcularon parámetros de tendencia central (media) y dispersión (desvío estándar) de los índices. La normalidad de las distribuciones fue testea aplicando la prueba de Kolmogorov-Smirnov para una muestra. Para valorar el efecto de los factores edad, sexo y tratamiento se aplicó análisis multifactorial de la varianza (ANOVA). En los casos de diferencias significativas se procedió a realizar comparaciones entre grupos mediante el método de la diferencia mínima significativa (LSD). Se consideró como nivel de significación los valores de p<0.05.
A los efectos de estandarizar y graficar la variación morfológica entre tratamientos se calcularon diferencias porcentuales entre medias (DPM) empleando la siguiente fórmula:
DPM=((X1 -X2 )/X1 )x100, donde: X1 =valor promedio del grupo RPC y X2 =valor promedio del grupo C.
El análisis de la varianza mostró diferencias significativas para los factores edad, sexo y tratamiento en todos los índices (Tabla 2).
Análisis de la varianza (ANOVA) para índices craneanos
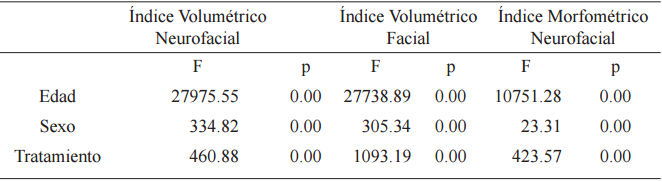
p<0.05
En las comparaciones post hoc hubo diferencias significativas entre C y SH, por lo tanto se procedió a emplear a este último grupo como control (Tabla 3).
Pruebas LSD para diferencias entres medias (DM) de los índices craneanos
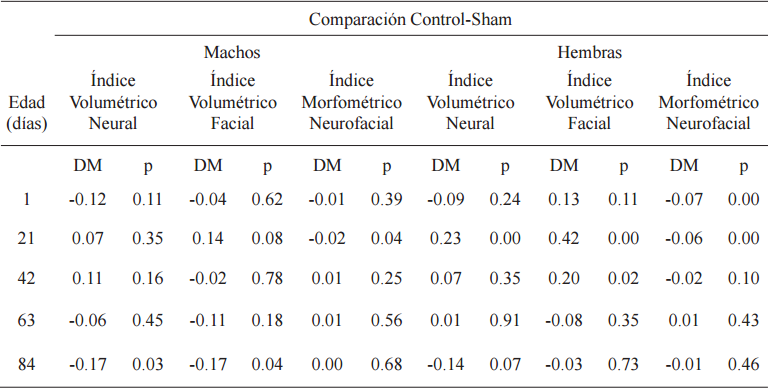
p< 0.05
Al nacimiento, en la comparación entre RPC+N y SH se observaron diferencias significativas en IVN, IVF e IMNF, con valores DPM negativos para los dos primeros y positivos para el tercero (Tabla 4, Fig. 2). En la lactancia RPC+N respecto de SH, indicó en machos diferencias significativas, con valores DPM negativos, para IVN e IVF y no significativas para IMNF. Las hembras no mostraron diferencias en ninguno de los índices analizados (Tabla 4, Fig. 3). Durante la postlactancia en IVN hubo diferencias significativas a los 63 y 84 días en machos y a los 42, 63 y 84 días en hembras, con valores DPM negativos en todos los casos.
Pruebas LSD para diferencias entres medias (DM) de los índices craneanos
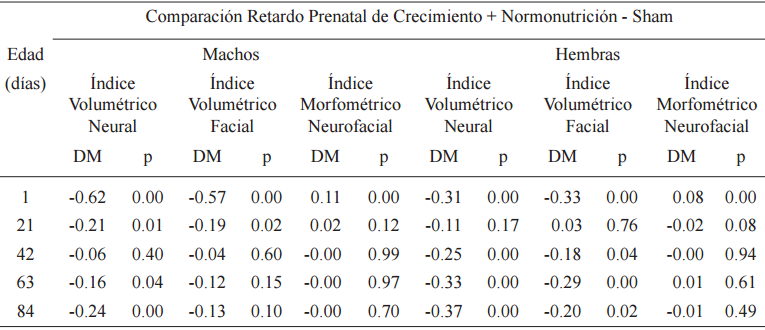
p<0.05
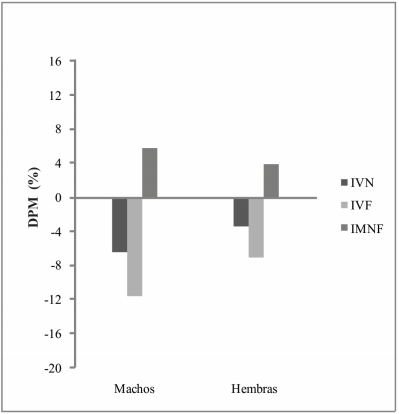
Fig. 2
Diferencias porcentuales entre medias (DPM) para la comparación entre RPC y SH al nacimiento. IVN:Índice Volumétrico Neural, IVF:Índice Volumétrico Facial; IMNF:Índice Morfométrico Neurofacial
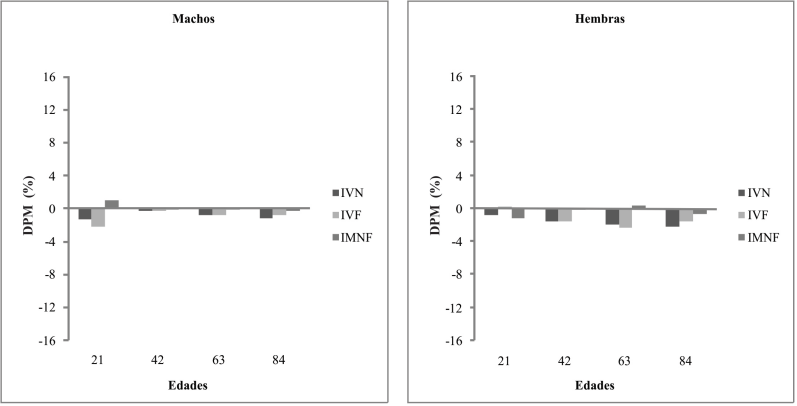
Fig. 3
Diferencias porcentuales entre medias (DPM) para la comparación entre RPC+N y SH. IVN:Índice Volumétrico Neural, IVF:Índice Volumétrico Facial; IMNF:Índice Morfométrico Neurofacial
En IVF en cambio, mientras que en los machos se presentaron diferencias no significativas, en hembras fueron significativas, con DPM negativos, a los 42, 63 y 84 días. En IMNF las diferencias fueron no significativas, en ambos sexos (Tabla 4, Fig. 3).
La comparación entre RPC+S y SH indicó a los 21, 42, 63 y 84 días de edad diferencias significativas en todos los índices, con excepción de IVN a los 21 días en hembras. Los valores DPM fueron negativos para IVN e IVF y positivos para IMNF (Tabla 5, Fig. 4).
Prueba de rango múltiple para índices volumétricos y mofométrico, neuro y esplacnocraneanos
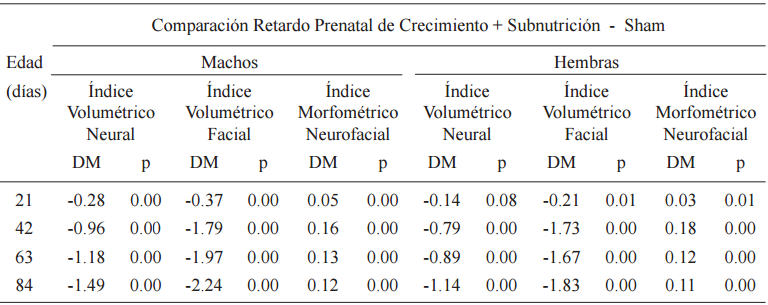
p<0.05
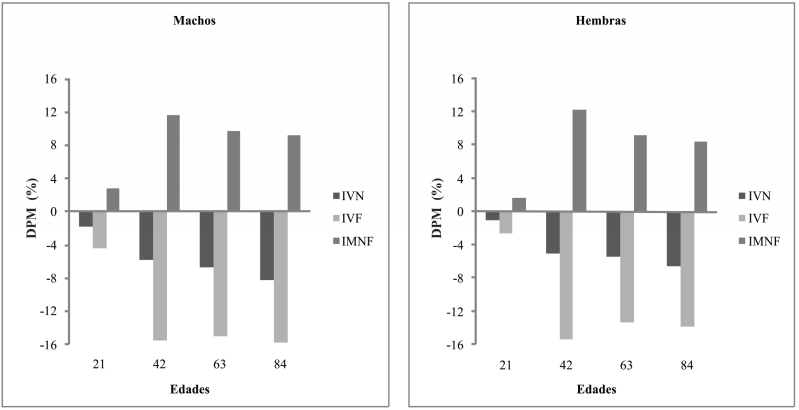
Fig. 4
Diferencias porcentuales entre medias (DPM) para la comparación entre RPC+S y SH. IVN:Índice Volumétrico Neural, IVF:Índice Volumétrico Facial; IMNF:Índice Morfométrico Neurofacial
Los resultados obtenidos, en coincidencia con estudios realizados previamente por Oyhenart et al. (1998; 2003) indicaron, que la disminución del flujo sanguíneo útero-placentario provocó al naci
Los resultados obtenidos, en coincidencia con estudios realizados previamente por Oyhenart et al. (1998; 2003) indicaron, que la disminución del flujo sanguíneo útero-placentario provocó al nacimiento retraso del crecimiento craneano, con cambios en la proporción entre ambos componentes mayores. En este sentido, el componente facial presentó mayor disminución de tamaño -12% en machos y 7% en hembras que el neural -7% en machos y 3% en hembras- (Fig. 2). La mayor estabilidad del neurocráneo puede ser explicada en relación al crecimiento en tamaño del cerebro que es el principal determinante de su morfología (Young, 1959; Ramírez Rozzi et al., 2005). Bajo estrés nutricional, el cerebro se modifica en menor proporción que otros órganos debido a su importancia funcional (Zamenhof y van Marthens, 1974; Dickerson y Pao, 1975; Sara et al., 1976). Al respecto, se considera que el feto en respuesta a la reducción del suministro de nutrientes desarrolla durante su crecimiento adaptaciones metabólicas que mejoran la capacidad de absorción y utilización de nutrientes (Baschat, 2004; Thorn et al., 2011). Ejemplo de ello es la adaptación hemodinámica por la cual el flujo sanguíneo es preferencialmente perfundido al cerebro, proceso conocido como brain sparing effect (Wladimiroff et al., 1987; van den Broek, et al., 2010).
De acuerdo a lo expresado por Alippi et al. (2002) la desnutrición durante la vida fetal o neonatal incide en la posibilidad de recuperación del crecimiento posterior, que en caso de ocurrir puede ser parcial o total. Este crecimiento compensatorio es atribuido al mecanismo auto-estabilizante denominado “canalización” que se manifiesta como la capacidad de alcanzar la curva de crecimiento original después de una deprivación nutricional (Tanner, 1962). Al respecto, los resultados obtenidos indicaron que en los animales con RPC la posibilidad de canalización del crecimiento durante la etapa postnatal fue dependiente del sexo. Aún cuando las madres y las crías recibieron alimentación normal, los machos continuaron con retardo de crecimiento neural y facial hasta los 21 días de edad postnatal y a partir de esa edad, solo tuvieron crecimiento compensatorio en la región facial. Las hembras en cambio, durante la lactancia mostraron tendencia a recuperar el crecimiento en el tamaño craneofacial. Sin embargo, durante la postlactancia el retardo continuó en ambos componentes (Fig. 3). Probablemente la disminución en la tasa de división celular de tejidos y órganos, la cual conduce a alteración en la programación estructural y funcional, constituya una respuesta adaptativa frente al aporte inadecuado de nutrientes (Lucas, 1998).
Por otra parte, cuando al RPC le continuó una subnutrición proteico-calórica durante la lactancia, el patrón de crecimiento también resultó modificado. Nuevamente y tal como fuera observado al nacimiento, el retardo de crecimiento craneano se manifestó en ambos componentes 4% y 3% para el componente facial y 2% y 1% para el componente neural, en machos y hembras respectivamente (Fig. 4). Sin embargo, el impacto fue menor que el ocurrido en la etapa prenatal probablemente por el efecto amortiguador materno, aún cuando la madre fuera desnutrida (Oyhenart et al., 1999). Por último, cuando la subnutrición continuó en la postlactancia la disminución del crecimiento craneofacial fue mayor, en promedio cercana al 16% en machos y 14% en hembras en el componente facial y al 7% en machos y 6% en hembras en el componente neural (Fig. 4).
El efecto diferencial sobre los componentes facial y neural condujo a modificación de la forma craneana durante toda la etapa postnatal. Una posible explicación a ello podría radicar en el patrón de crecimiento propio de cada región. De acuerdo a lo informado por Clark y Smith (1993), al nacimiento y bajo condiciones normales, el neurocráneo ha completado la mayor parte de su crecimiento. En promedio, el cráneo neural de la rata alcanza el 93% del tamaño adulto a los 30 días de edad, mientras que el facial a la misma edad alcanza sólo el 75% (Lightfoot y German, 1998; Moore, 1966). El crecimiento y desarrollo de los vertebrados consiste en un proceso altamente ordenado que puede ser fragmentado en procesos relativamente disociados. Esta fragmentación, conocida como modularidad, posibilita la adaptación independiente de manera que cada módulo funcional puede ser modificado diferencialmente por influencias ambientales y presentar un potencial de recuperación propio sin pérdida de funcionalidad (Bonner, 1988; Polly et al, 2001). Al respecto Miller y German (1999) informaron que debido a que las demandas funcionales del esplacnocráneo son mayores después del nacimiento, la probabilidad que tiene de ser modificado por factores epigenéticos también es mayor.
Los resultados obtenidos permiten concluir que la disminución del flujo sanguíneo maternofetal modifica, al nacimiento, el tamaño y la forma craneana en ambos sexos.
Las consecuencias del estrés primario ocurrido durante la vida intrauterina resultan críticas no sólo en lo inmediato, sino también en la vida postnatal. Prueba de ello es que, aún mediando normonutrición, el retardo de crecimiento en las hembras perdura hasta la adultez, en tanto que en los machos se presenta crecimiento compensatorio incompleto.
Por otra parte, si luego del estrés prenatal continúa una restricción nutricional, los efectos adversos son aditivos, por cuanto el retraso del crecimiento craneano resulta aún mayor, particularmente en la región facial, persistiendo hasta edad adulta en ambos sexos.
Finalmente, mientras que el componente neural es más resistente frente a las deficiencias nutricionales, el componente facial presenta mayor plasticidad, hecho que queda evidenciado en cambios en la forma craneana.
Los resultados obtenidos aunque no directamente extrapolables al hombre permiten avanzar en el conocimiento de los procesos adaptativos y evolutivos en relación al estrés nutricional.
En memoria de la Med. Vet. Miriam Villanueva por el compromiso y profesionalismo puesto en el desarrollo del trabajo.
Los autores agradecen a Adriana Di Maggio por la revisión idiomática.
Dra. María Eugenia Luna. Instituto de Genética Veterinaria, Facultad de Ciencias Veterinarias, UNLP-CONICET. Calle 60 y 118. B1900ASV La Plata. Buenos Aires. Argentina
Grupos experimentales de acuerdo al tratamiento aplicado
Fig. 1
Radiografía en vista dorsal (izquierda) y lateral (derecha) del cráneo de la rata mostrando las mediciones empleadas: LN:longitud neural, AN: ancho neural, HN:altura neural, LF: longitud facial, AF:ancho facial, HF:altura facial
Análisis de la varianza (ANOVA) para índices craneanos
p<0.05
Pruebas LSD para diferencias entres medias (DM) de los índices craneanos
p< 0.05
Pruebas LSD para diferencias entres medias (DM) de los índices craneanos
p<0.05
Fig. 2
Diferencias porcentuales entre medias (DPM) para la comparación entre RPC y SH al nacimiento. IVN:Índice Volumétrico Neural, IVF:Índice Volumétrico Facial; IMNF:Índice Morfométrico Neurofacial
Fig. 3
Diferencias porcentuales entre medias (DPM) para la comparación entre RPC+N y SH. IVN:Índice Volumétrico Neural, IVF:Índice Volumétrico Facial; IMNF:Índice Morfométrico Neurofacial
Prueba de rango múltiple para índices volumétricos y mofométrico, neuro y esplacnocraneanos
p<0.05
Fig. 4
Diferencias porcentuales entre medias (DPM) para la comparación entre RPC+S y SH. IVN:Índice Volumétrico Neural, IVF:Índice Volumétrico Facial; IMNF:Índice Morfométrico Neurofacial
