ARTICULO INVESTIGACIÓN
Presencia de cerambícidos en plantaciones de Eucalyptus urograndis
Presence of Cerambycids in Plantations of Eucalyptus urograndis
 Claudio Fernández Herrera
Martha Fonseca Angulo
Claudio Fernández Herrera
Martha Fonseca Angulo
Presencia de cerambícidos en plantaciones de Eucalyptus urograndis
Colombia Forestal, vol. 28, no. 1, pp. 1-16, 2025
Proyecto Curricular de Ingeniería Forestal, Facultad del Medio Ambiente y Recursos Naturales, Universidad Distrital Francisco José de Caldas.
Received: 19 October 2024
Accepted: 06 November 2024
Resumen: Eucalyptus urograndis es un híbrido entre Eucalyptus urophylla y Eucalyptus grandis ampliamente plantado en el departamento de Córdoba, Colombia, hogar de muchas especies de cerambícidos. Por ello, en este estudio se realizaron muestreos en plantaciones y su vegetación circundante mediante búsqueda directa. La composición de cerambícidos en estas áreas agrupó 52 individuos de la subfamilia Lamiinae, que presentó la mayor abundancia al interior de las plantaciones, registrando 42 especímenes distribuidas entre Dorcasta dasycera (Erichson, 1848), Lepturges sp., Adetus tayronus Galileo y Martins, 2003, Oedopeza sp. y Steirastoma histrionica White, 1855. Entretanto, Cerambycinae registró 5 especies: Oxymerus aculeatus lebasi Dupont, 1838, Ceralocyna sp., Hexoplon albipenne Bates, 1872, Chrysoprasis chevrolati Lameere, 1884 y Trachyderes succinctus succinctus (Linnaeus, 1758). Se concluyó que, en relación con la diversidad de cerambícidos al interior de las plantaciones de E. urograndis, la subfamilia Lamiinae es favorecida por las precipitaciones.
Palabras clave: Comportamiento, caracterización, Eucalyptus, estudios forestales, longicornios.
Abstract: Eucalyptus urograndis is a hybrid between Eucalyptus urophylla and Eucalyptus grandis that is widely planted in the department of Córdoba, Colombia, which is home to many species of cerambycids. Therefore, this study conducted sampling activities using direct search in plantations and their surrounding vegetation. The composition of cerambycids in these areas grouped 52 individuals from the subfamily Lamiinae, which exhibited the greatest abundance within the plantations, recording 42 specimens distributed among Dorcasta dasycera (Erichson, 1848), Lepturges sp., Adetus tayronus Galileo and Martins, 2003, Oedopeza sp., and Steirastoma histrionica White, 1855. Meanwhile, the subfamily Cerambycinae recorded five species: Oxymerus aculeatus lebasi Dupont, 1838, Ceralocyna sp., Hexoplon albipenne Bates, 1872, Chrysoprasis chevrolati Lameere, 1884, and Trachyderes succinctus succinctus (Linnaeus, 1758). It was concluded that, concerning the diversity of cerambycids within E. urograndis plantations, the subfamily Lamiinae is favored by rainfall.
Keywords: Behavior, characterization, eucalyptus, forest studies, longhorns.
Highlights
-
En Córdoba, Colombia, Eucalyptus urograndis actualmente presenta un área de 11 000 ha.
-
Los cerambícidos utilizan las plantaciones de E. urograndis como nicho ecológico.
-
Las plantaciones de E. urograndis estudiadas asociaron a la subfamília Lamiinae y Cerambycinae.
-
Lamiinae registró la mayor abundancia al interior de las plantaciones.
-
Dorcasta dasycera (Erichson, 1848), Lepturges sp. y Adetus tayronus Galileo y Martins, 2003 fueron las especies más abundantes.
INTRODUCCIÓN
Eucalyptus (L'Héritier, 1789) es un género de árboles que, por su rendimiento y calidad, permiten múltiples usos (Singh & Dhillon, 2020) y de los cuales existen aproximadamente 57 mil hectáreas sembradas en Colombia (PROFOR, 2017). En las regiones tropicales del mundo, se destaca por su buena adaptación y producción la especie Eucalyptus urograndis, un híbrido entre Eucalyptus urophylla y Eucalyptus grandis, conocida por su rápido crecimiento (mayor a 45 m³.ha.año-1). Esta especie crece normalmente hasta los 25 m de altura, y en ocasiones alcanza los 50 m, con diámetros de 0.3 a 1.5 m (Carvalho, 2003; Marco de Lima et al., 2019; Salvador et al., 2019). Hace parte de un ecosistema forestal que es el hábitat de muchas especies de insectos, entre ellos los cerambícidos, que lo utilizan como su nicho ecológico. La familia Cerambycidae es conocida como escarabajos longicornios debido a la longitud de las antenas de sus individuos, que por lo general son más largas que el resto del cuerpo. Ecológicamente, este grupo de coleópteros contribuye a la circulación de los minerales hacia el suelo al momento de barrenar, permitiendo la invasión de hongos (Martínez, 2000). Su alimentación es variada e incluye troncos, ramas, frutos, semillas, raíces y tallos herbáceos (Morales-Morales et al., 2012; Noguera, 2014; Álvarez et al., 2015).
La presencia de algunas especies de cerambícidos puede estar determinada por diversos factores, como lo son la composición taxonómica de los árboles, la cubierta del dosel y los árboles en descomposición (Fahri et al., 2016). Algunos autores como Calderón-Cortés et al. (2011), Lemes et al. (2015) y Novais et al. (2017) los consideran como ingenieros ecológicos debido a sus interacciones directas e indirectas con los hábitats donde se asocian.
Otro aspecto importante de los cerambícidos es su relación con factores ambientales, lo que permite catalogarlos como organismos bioindicadores (Noguera et al., 2012; Noguera, 2014). También se resalta su incidencia en los sistemas forestales, como lo indican Giraldo-Aristizábal et al. (2022), quienes registraron la presencia de larvas y un adulto de Cerambycidae causando daño y deterioro evidente en ramas de Quercus humboldtii Bonpl. 1805.
El género Phoracantha Pascoe (1866), que se ha adaptado a la región neotropical sin depredadores autóctonos, es una plaga que ataca a especies del género Eucalyptus (López-Pérez, 2007). Beéche et al. (2003) describen el daño causado por larvas de Phoracantha semipunctata (Fabricius, 1775) al alimentarse de tejidos conductores y de crecimiento, provocando la muerte por anillamiento. De igual manera, Berti Filho (1997) observó a Sphallenum tuberosum (Bates, 1870) perforando troncos de Eucalyptus citriodora y Eucalyptus tereticornis. Estos antecedentes merecen una adecuada atención y, debido a la estrecha relación que guardan, se planteó como hipótesis caracterizar el papel de los cerambícidos en la fenología de las plantaciones de E. urograndis. Para probar dicha hipótesis, se analizó la variación temporal de la comunidad de cerambícidos en cuatro localidades con presencia de plantaciones de E. urograndis.
MATERIALES Y MÉTODOS
Los muestreos se realizaron en los municipios de Valencia (Localidades Volcanes, 8.29515 N -76.20875E, y Guadual, 8.12577 N -76.33606 E), Montelíbano (Localidad San Francisco del Rayo, 8.003187 N -75.827850 E), Arbolete (Localidad Campanito, 8.526669N -76.444854E) y Urabá (San Pedro de Urabá, 8.275N -76.376944E). Para cada localidad, se registraron la temperatura, la precipitación diaria (mm3) mediante un pluviómetro y la humedad utilizando un higrómetro.
Los muestreos se realizaron dentro de las plantaciones de E. urograndis de cada localidad y en relictos de vegetación nativa como especies arbóreas de la familia Fabaceae, arbustos de la familia Euphorbiaceae, Malvaceae y Verbenaceae, y plantas de la familia Poaceae (gramíneas). De igual manera, se realizó una búsqueda directa, con una intensidad de 15 minutos por punto, en los hábitats que ocupan los cerambícidos, como la hojarasca y los árboles en descomposición. En cada localidad se delimitaron cuatro parcelas cuadradas de 50 x 3 m, separadas por 400 m. Para aplicar la técnica de fogging, se utilizaron equipos de protección y el insecticida ASTUTO 200 SC (Registro Nacional ICA No. 1540). Este insecticida, en forma de suspensión concentrada y con fipronil 200 g.l-1 como ingrediente activo, se aplicó sobre la cubierta de árboles de diferentes edades y características, a saber: a) árboles de 1 año y una altura aproximada de 8-10 m; b) árboles de 3 años y una altura aproximada de 14-15 m; c) árboles de 5 años y una altura aproximada de 20-22 m. Para esta técnica, se instaló una tela tull por encima del suelo y entre las plantas, con el fin de atrapar los insectos que caían tras la aplicación del insecticida. Los especímenes se recolectaron 48 horas post-aplicación. Además, se realizaron recorridos periódicos dentro y fuera de las plantaciones durante el periodo de lluvias, el cual abarcó los meses de abril a junio y de agosto a noviembre, normalmente con precipitaciones entre las 09:00 y las 15:00.
El material biológico se almacenó en bolsas de papel Kraft para extraer la humedad y posteriormente se depositó en bolsas plásticas Ziploc debidamente selladas y rotuladas. Acto seguido, los cerambícidos colectados fueron trasladados al laboratorio de entomología de la Universidad de Córdoba, donde se realizó un análisis de completitud que consideró las coberturas estandarizadas de las muestras (IC=95 %), la diversidad verdadera y el grado de recambio de especies reales. Esto, a través de los valores q = 0, 1 y 2, donde q = 0 representa la diversidad de orden cero e implica que no se consideran abundancias, por lo que equivale a la riqueza de especies; q = 1 es la diversidad de Shannon, que incluye todas las especies con un peso proporcional a su abundancia en el ensamblaje, siendo este el índice de diversidad; y q = 2 representa la diversidad de Simpson, que tiene en cuenta las especies dominantes y excluye las raras. Además, se realizó un análisis multivariado de correspondencia múltiple, y se identificaron las principales especies con la ayuda de un estereoscopio (Advanced Optical JSZ6S con cámara INFINITY 1 incorporada) y utilizando claves taxonómicas (Toledo, 1997; Verdugo, 2004; Toledo & Hovore, 2005; Maes et al., 2010a, 2010b, 2010c, 2010d, 2010e; Enríquez, 2012; Monné & Bezark, 2014).
RESULTADOS
Caracterización de cerambícidos en plantaciones de E. urograndis
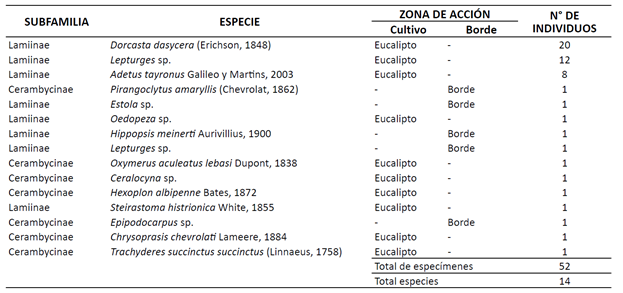
Se registró un total de 52 especímenes de cerambícidos en las plantaciones de E. urograndis analizadas, correspondientes a las subfamilias Lamiinae y Cerambycinae. Las especies más abundantes fueron Dorcasta dasycera (Erichson, 1848) con 20 individuos, Lepturges sp. con 13 individuos, Adetus tayronus Galileo y Martins, 2003 con 8 individuos y, por último, las especies Pirangoclytus amaryllis (Chevrolat, 1862), Estola sp., Ceralocyna sp., Chrysoprasis chevrolati Lameere, 1884, Epipodocarpus sp., Hexoplon albipenne Bates, 1872, Hippopsis meinerti Aurivillius, 1900, Oedopeza sp., Oxymerus aculeatus lebasi Dupont, 1838, Steirastoma histrionica White, 1855 y Trachyderes succinctus succinctus (Linnaeus, 1758), todas con un espécimen (Figura 1).

Figura 1
Cerambícidos presentes en plantaciones de E. urograndis. Lamiinae: a. Lepturges sp.; b. Dorcasta dasycera; c. Hippopsis meinerti; d. Adetus tayronus; e. Estola sp. f. Oedopeza sp.; g. Steirastoma histrionica; Cerambycinae: h. Oxymerus aculeatus lebasi; i. Pirangoclytus amaryllis; j. Ceralocyna sp.; k. Hexoplon albipenne; l. Epipodocarpus sp.; m. Chrysoprasis chevrolati; n. Trachyderes succinctus succinctus.
Los cerambícidos resaltaron al interior de las plantaciones de eucalipto, siendo esta la zona de mayor abundancia relativa; la subfamilia Lamiinae registró 41 especímenes, destacando a D. dasycera, Lepturges sp. y A. tayronus, teniendo en cuenta que el resto de las especies fueron registradas como singletones. Por su parte, en los bordes de las plantaciones, todas las especies estuvieron representadas por una alta incidencia de singletones de la subfamilia Lamiinae, con Estola sp., H. meinerti y Lepturges sp.; y de la subfamilia Cerambycinae, con P. amaryllis y Epipodocarpus sp. (Tabla 1).
El análisis de cobertura (sample coverage, SC) permitió medir la completitud de la muestra en cuanto a la cantidad de especies con respecto a las potenciales. El valor obtenido para el cultivo de eucalipto fue del 82 %, y para borde fue del 11 % (Figura 2).
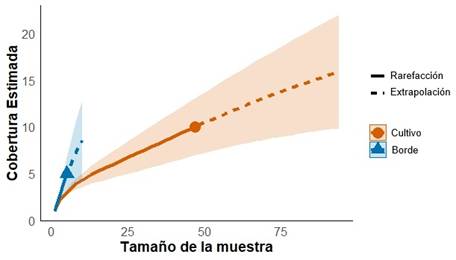
Figura 2
Análisis de completitud de la muestra al interior y em los bordes de las plantaciones de E. urograndis
Los índices de diversidad de Shannon presentaron un valor de 18.56 para los bordes, lo que sugiere que las especies en este hábitat presentan una distribución de abundancia relativamente equitativa. Entretanto, para el cultivo de eucalipto, este valor fue de 6.46, i.e., hay algunas especies con mayor abundancia. La riqueza de especies en los bordes fue de 5.0, con un estimado de 13.0 y un error estándar (SE) de 3.08. En cambio, para el cultivo de eucalipto, el valor fue de 10.0, con un estimado de 30.55 y un SE de 5.10 (Tabla 2).
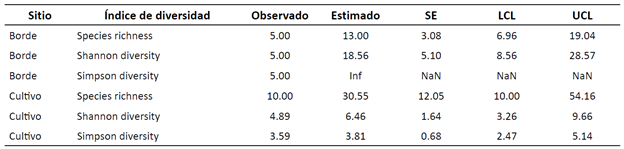
*(SE): error estándar
La diversidad de Simpson para los bordes se muestra como ‘Inf’ (infinito), lo que implica que ninguna especie domina el ecosistema. Por su parte, en el cultivo de eucalipto se registró 3.81, i.e., algunas especies dominan en este hábitat (Figura 3).
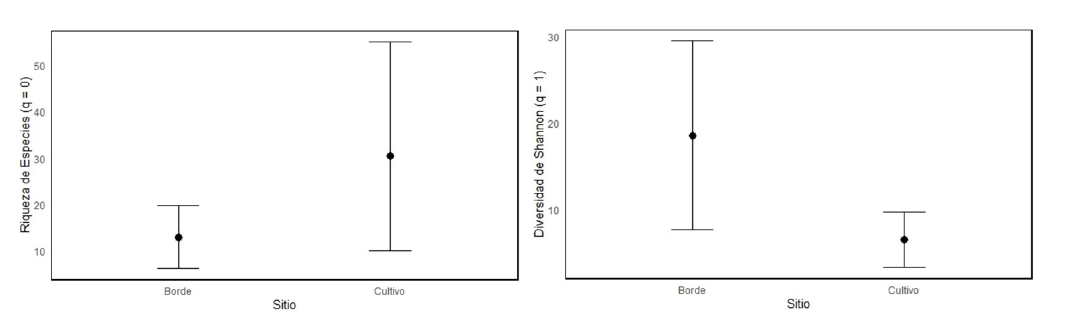
Figura 3
Riqueza de especies (q=0); índices de diversidad de Shannon (q=1) para los bordes y el cultivo de las plantaciones de eucalipto
Por su parte, el análisis de varianza para las subfamilias registradas muestra que la abundancia de especies de Lamiinae depende únicamente de la precipitación (valor p<0.05) y que es favorecida de manera positiva por el incremento de estas condiciones climáticas (Tabla 3). El modelo obtenido corresponde a: Log (y) = 21.49 - 1.44 * Temp. media + 0.12 * Temp. máxima + 0.02 * Precipitación + Humedad relativa * 0.06
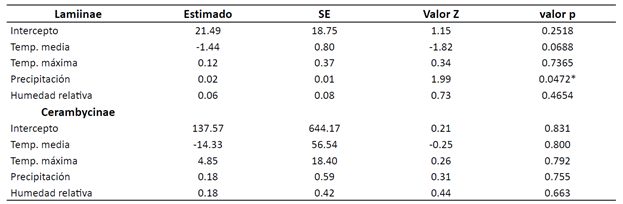
En cambio, la subfamilia Cerambycinae registro 5 especímenes pertenecientes a las especies O. aculeatus lebasi, Ceralocyna sp., H. albipenne, C. chevrolati y T. succinctus succinctus. El análisis de varianza muestra que la abundancia de especie no fue influenciada por ninguno de los factores ambiental (p-valor > 0.05).
Diversidad de cerambícidos asociados en las diferentes localidades con plantaciones de E. urograndis
Las diferentes localidades que registraron especies de cerambícidos presentaron una temperatura promedio de 28 ºC, una humedad relativa del 83 % y altitudes desde los 100 hasta los 223 m. El total de especímenes registrados por localidad y zona de recolección (interior del cultivo y bordes), mostró que Volcanes (Valencia) posee la mayor diversidad, con el 78.8% de los muestreos, correspondientes a las especies D. dasycera, Lepturges sp., A. tayronus, P. amaryllis y Estola sp. La localidad de Campanito (Arboletes) presenta un 5.8 % de los muestreos, asociados a las especies Oedopeza sp., H. meinerti y Lepturges sp. Por su parte, Guadual (Valencia) asocia el 5.8 %, registrando a D. dasycera., O. aculeatus lebasi y Ceralocyna sp., y la localidad de San Francisco del Rayo (Montelíbano) registra un 5.8 % para las especies H. albipenne, S. histrionica y Epipodocarpus sp. Por último, la localidad de San Pedro de Urabá (Urabá) posee la menor diversidad, registrando el 3.8 % de los muestreos y asociando a las especies C. chevrolati y T. succinctus succinctus.
De acuerdo con el análisis de correspondencia múltiple, las especies D. dasycera, Adetus tayronus, y Oedopeza sp. se registraron como trozadoras al interior de las plantaciones de seis meses de E. urogarndis, comprendiendo las localidades de Volcanes y Campanito. De igual manera se comportaron las especies T. succinctus succinctus, C. chevrolati y Ceralocyna sp. en las localidades de Guadual y San Pedro de Urabá (SPU). En los bordes de las plantaciones de E. urogarndis, se registraron las especies H. meinerti, P. amaryllis, S. histrionica, H. albipenne y Epipodocarpus sp. actuando como barrenadoras en la localidad de San Francisco del Rayo (SFR) (Figura 4).
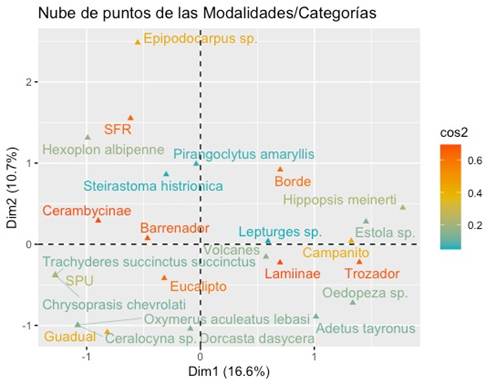
Figura 4
Análisis de correspondencia múltiple por localidades y rol trófico de las principales especies presentes en las plantaciones de E. urograndis
DISCUSIÓN
En cuanto a la composición de cerambícidos, los resultados muestran que las subfamilias Lamiinae y Cerambycinae son las más abundantes en las plantaciones de E. urograndis. Según esta asociación, la comunidad de cerambícidos de la subfamilia Lamiinae registró mayor abundancia en el interior que en el borde. Estos resultados concuerdan con los reportados por Goldsmith et al. (2007), Noguera et al. (2012) y Botero (2018), quienes afirman que muchas especies de cerambícidos están fuertemente asociadas a su planta hospedera. Según Santos y Pereira-Colavite (2017), además, la variación de la vegetación a través del tiempo tiene un papel fundamental. Esta incidencia se vio reflejada en la época de lluvias (abril-octubre), que reportó precipitaciones entre 110 mm (abril, el mes más seco) y 374 mm (septiembre, el mes más lluvioso).
Estos resultados guardan relación con los reportados por Gutiérrez et al. (2014) y Botero (2018), quienes afirman que Lamiinae es más diversa y abundante en bosques tropicales lluviosos. También concuerdan con los de Sugiarto y Mardji (2016) y Noguera et al. (2017): cuando llegan las lluvias, aumentan los recursos en el bosque y las actividades de alimentación y apareamiento de los adultos de muchas especies de cerambícidos. En contraste, la subfamilia Cerambycinae, al estar más ligada a bosques tropicales secos, reportó una menor abundancia, semejante a las reportadas por Toledo et al. (2002), Noguera et al. (2009, 2012) y Bernardi et al. (2011). Estos autores mencionan que muchos de estos cerambícidos muestran picos poblacionales en los meses de mayor temperatura.
En cuanto a la medición de la abundancia y los tipos de muestreos realizados, la colecta manual fue más eficiente, agrupando a la mayoría de las especies de la subfamilia Lamiinae, cuyo comportamiento está relacionado con sus hábitos alimenticios. Sin embargo, se esperaría que este tipo de muestreo favoreciera más a las especies de la subfamilia Cerambycinae (MacKay et al., 2014; Álvarez et al., 2015; Wang, 2017).
Sobre el comportamiento de las especies, se puede afirmar que D. dasycera fue la más abundante en este estudio, seguida de Lepturges sp. y A. tayronus. Por consiguiente, al ser la especie dominante, D. dasycera puede adaptarse a modificaciones en su entorno relacionadas con la expansión del cultivo, lo que favorecería su aumento poblacional y podría causar daños económicos. Estos resultados pueden atribuirse, según Sataral et al. (2015) y Noerdjito (2012), al hecho de que muchos cerambícidos de menor tamaño tienden a ser más ricos y abundantes en ecosistemas tropicales, pues se encuentran asociados a ramas pequeñas, donde sus larvas pueden vivir y desarrollarse.
Por su parte, la baja abundancia de las especies P. amaryllis, Estola sp., Oedopeza sp., H. meinerti, Lepturges sp., O. aculeatus, Ceralocyna sp., H. colombianum, S. histriónica, Epipodocarpus sp., C. chevrolati y T. succinctus puede atribuirse a su actividad y a las técnicas de muestreo utilizadas -muchas de estas especies son nocturnas. Esto guarda relación con lo expresado por Basset et al. (2012), quienes atribuyeron este comportamiento a una característica de los grupos megadiversos en ambientes tropicales, donde se registra una alta riqueza de especies con bajas abundancias.
En cuanto a la subfamilia Cerambycinae, la alta incidencia de singletones para las especies P. amaryllis, O. aculeatus lebasi, Ceralocyna sp., H. albipenne, Epipodocarpus sp., C. chevrolati y T. succinctus succinctus en las localidades de Volcanes, Guadual, San Francisco de Rayo y San Pedro de Urabá se debe a la alteración de la flora y a las condiciones ambientales de los muestreos. Nuestros resultados se asemejan a los reportados por Wang (2002, 2017) y Álvarez et al. (2015), según los cuales esto guarda relación con los cortos períodos de vida y alimentación de los adultos -las especies asociadas dependen de la temporada de floración de la planta hospedera.
También se pudo observar que las especies P. amaryllis, O. aculeatus lebasi, Ceralocyna sp., H. albipenne, Epipodocarpus sp., C. chevrolati y T. succinctus succinctus son más comunes en el bosque seco, lo que concuerda con los resultados de Luna-León et al. (2015), Toledo et al. (2002), Noguera et al. (2012) y Morales-Morales et al. (2012). Estos autores argumentan que las especies de la subfamilia Cerambycinae se establece más en bosque tropical caducifolio de Guerrero, México. Nuestros resultados también guardan relación con los reportados por Álvarez-Ramon et al. (2022) en las selvas de México y a los de Gerónimo-Torres et al. (2024) en manglares. Por su parte, Noerdjito (2012) afirma que la heterogeneidad y el recambio de la vegetación podrían explicar la baja similitud de especies de cerambícidos, si bien Marini et al. (2013), Stadelmann et al. (2013) y Gerónimo-Torres et al. (2021, 2024) establecen que la fluctuación de estas especies está ligada a las precipitaciones.
Al comparar la diversidad de especies asociadas por localidades, Campanito, Guadual, San Francisco del Rayo y San Pedro de Urabá presentaron una diversidad similar en las comunidades de cerambícidos, lo que obedece al tipo de muestreo utilizado (captura manual y fogging). Estos resultados se asemejan a los de Sataral et al. (2015), quienes mencionan que este comportamiento está ligado a la repartición del espacio de los cerambícidos; al colocar las trampas a diferentes alturas, se pueden capturar especies asociadas a diferentes estratos vegetales. Nuestros hallazgos también concuerdan con los de Noguera et al. (2012) y Noguera (2014). Según estos estudios, la relación entre los factores ambientales y los cerambícidos permite catalogarlos como organismos bioindicadores, y su riqueza, abundancia y distribución pueden verse alteradas por cambios en el uso del suelo y la tala y extracción de madera, antecedentes que se presentan en las localidades donde realizamos nuestros muestreos.
Estos antecedentes, junto con la presencia de las especies registradas de Cerambycinae y Lamiinae en las diferentes localidades, pueden traducirse en problemas con estas plagas. Esta afirmación es consistente con lo reportado por Zanuncio et al. (1994), quienes afirman que un incremento del área reforestada con esencias exóticas puede aumentar los problemas relacionados con plagas de insectos. Esto, debido a cambios ambientales en los monocultivos.
CONCLUSIONES
La diversidad de cerambícidos estuvo marcada por las especies asociadas al interior del cultivo y representadas por las subfamilias Cerambycinae y Lamiinae. Esta última expresa un comportamiento diferenciado y es altamente favorecida por las condiciones alimenticias y de precipitación en las zonas de estudio. En vista de lo anterior, es necesario implementar otros sistemas de muestreo para estudios posteriores que permitan establecer la diversidad de este grupo de insectos en plantaciones de E. uraograndis en época seca.12
AGRADECIMIENTOS
A la Universidad de Córdoba, a la Reforestadora del Sinú, al laboratorio de Entomología de la Universidad de Córdoba y al personal que apoyó la toma de muestras en la zona donde se desarrolló la investigación.
REFERENCIAS
Álvarez, G., Ammagarahalli, B., Hall, D. R., Pajares, A., & Gemeno, C. (2015). Smoke, pheromone and kairomone olfactory receptor neurons in males and females of the pine sawyer Monochamus galloprovincialis (Olivier) (Coleoptera: Cerambycidae). Journal of Insect Physiology, 82, 46-55. https://doi.org/10.1016/j.jinsphys.2015.08.004
Álvarez-Ramón, Ó., Pérez-de la Cruz, M., Magaña-Alejandro, M., Oporto-Peregrino, S., & Gerónimo-Torres, J. (2022). Diversidad y fluctuación anual de cerambícidos (Coleoptera: Cerambycidae) en una selva tropical del sureste de México. Acta Biológica Colombiana, 27(1), 79-87. https://doi.org/10.15446/abc.v27n1.89421
Basset, Y., Cizek, L., Cuénoud, P., Didham, R., Guilhaumon, F., Missa, O., Novotny, V., Ødegaard, F., Roslin, T., Schmidl, J., Tishechkin, A., Winchester, N., Roubik, D., Aberlenc, H., Bail, J., Barrios, H., Bridle, J., Castaño, G., … Leponce, M. (2012). Arthropod diversity in a tropical forest. Science, 338(6113), 1481-1484. https://doi.org/10.1126/science.1226727
Beéche, M., Coycoolea, C., Rothman, S., Sandoval, A., Ide, M., & Espinoza, S. (2003). Detección y control biológico de los taladradores de eucalipto en Chile. Phoracantha semipunctata y Phoracantha recurva Newman (Coleoptera: Cerambycidae). Ministerio de Agricultura, Gobierno de Chile. https://doi.org/10.46908/tayacaja.v6i1.202
Bernardi, O., Silveira-Garcia, M., Ely e , Silva , E. , Fialho-Zazycki , L. , Bernardi , D. , Miorelli & D. , Finkenauer É., (2011). Besouros cerambycidae associados a Eucalyptus spp. no município de Pinheiro Machado, RS. Ciência Florestal, 21(1), 23-30. http://dx.doi.org/10.5902/198050982744
Berti Filho, E. (1997). Impacto de Coleoptera: Cerambycidae en florestas de Eucalyptus no Brasil. Scientia Forestalis, 52(1), 53-56. https://www.ipef.br/PUBLICACOES/SCIENTIA/nr52/cap6.pdf
Botero, J. P. (2018). La familia Cerambycidae (Coleoptera: Chrysomeloidea) en Colombia. En C. Deloya & H. Gasca (Eds.), Escarabajos del neotrópico (Insecta: Coleoptera) (pp. 153-170). S & G Editores. https://www.researchgate.net/publication/330656517_La_familia_Cerambycidae_Coleoptera_Chrysomeloidea_en_Colombia
Calderón-Cortés, N., Quesada, M., & Escalera-Vázquez, L. (2011). Insects as stem engineers: Interactions mediated by the twig-girdler Oncideres albomarginata chamela enhance arthropod diversity. PLoS ONE, 6(4), e19083. https://doi.org/10.1371/journal.pone.0019083
Carvalho, G., Carvalho, M., Ferreira, D., Faria, J., & Broz R., . (2001) Diterpenos, triterpenos e esteróides das flores de Wedelia paludosa. Quimica Nova, 24(1), 24. http://dx.doi.org/10.1590/S0100-40422001000100006
Enríquez, A. (2012). Caracterización de la comunidad Cerambycidae (Coleoptera) en Igapó y Moretal de la Amazonía Ecuatoriana, período 2008 - 2010. (Tesis de maestría, Universidad Central del Ecuador). http://www.dspace.uce.edu.ec/handle/25000/590
Fahri, A., Atmowidi, T., & Noerdjito, W. (2016). Diversity and abundance of cerambycid beetles in the four major land-use types found in Jambi Province, Indonesia. HAYATI Journal of Biosciences, 23(2), 56-61. https://doi.org/10.1016/j.hjb.2016.06.001
Gerónimo-Torres, J., Pérez-de la Cruz, M., de la Cruz-Pérez, A., Arias-Rodríguez, L., & Burelo-Ramos, C. (2021). Diversidad y distribución vertical de escarabajos barrenadores (Coleópteros: Bostrichidae, Curculionidae: Scolytinae, Platypodinae) en un manglar de Tabasco, México. Caldasia, 43(1), 172-185. https://doi.org/10.15446/caldasia.v43n1.84499
Gerónimo-Torres, J., Rios-Rodas, L., Alvarez-Ramon, O., Sanchez-Gutiérrez, F., & Pérez-de la Cruz, M. (2024). Diversidad de escarabajos longicornios (Coleoptera: Cerambycidae) asociados a un manglar de Tabasco, México. Revista de Biología Tropical, 72, e55604. https://doi.org/10.15517/rev.biol.trop.v72i1.55604
Giraldo-Aristizábal, I., Mera-Velasco, A., & Rivas-Zúñiga, C., et al. (2022). Evaluación fitosanitaria de Juglans neotropica y Quercus humboldtii en arbolado urbano de Popayán-Cauca, Colombia. Revista de la Academia Colombiana de Ciencias Exactas, Físicas y Naturales, 46(178), 169-181. https://doi.org/10.18257/raccefyn.1542
Goldsmith, S., Gillespie, H., & Weatherby, C. (2007). Restoration of Hawaiian montane wet forest: endemic longhorned beetles (Cerambycidae: Plagithmysus) in koa (Fabaceae: Acacia koa) plantations and in intact forest. The Southwestern Naturalist, 52(3), 356-363. http://www.jstor.org/stable/20424840
Gutiérrez, N., Márquez, J., & Noguera, F. (2014). Coleoptera: Cerambycidae (Insecta: Coleoptera) de una localidad con bosque mesófilo de montaña en Hidalgo, México. Dugesiana, 21(2), 143-150. https://doi.org/10.32870/dugesiana.v21i2.4153
Lemes, P., Cordeiro, G., Jorge, I., Dos Anjos, N., & Zanuncio, J. (2015). Cerambycidae and other Coleoptera associated with branches girdled by Oncideres saga Dalman (Coleoptera: Cerambycidae: Lamiinae: Onciderini). The Coleopterists Bulletin, 69(1), 159-166. https://doi.org/10.1649/0010-065X-69.1.159
López-Pérez, J. (2007). Corología de los Phoracanthini (Coleoptera: Cerambycidae) en la provincia de Huelva (Andalucía, suroeste de España). Boletín de la Sociedad Entomológica Aragonesa, 41, 441-443. https://typeset.io/pdf/corologia-de-los-phoracanthini-coleoptera-cerambycidae-en-la-30oaqkolbl.pdf
Luna-León, C., Domínguez-Márquez, V., & Ordoñez-Reséndiz, M., Mundo-Bahena, A. (2015). Diversidad de la familia Cerambycidae (Coleoptera) de un bosque tropical caducifolio de la comunidad de Taxco el Viejo, Guerrero, México. Entomología Mexicana, 2, 835-839. https://www.acaentmex.org/entomologia/revista/2015/SM/PAG%20%20835-839.pdf
Mackay, C., Sweeney, D., & Hillier, N. (2014). Morphology of antennal sensilla of the brown spruce longhorn beetle, Tetropium fuscum (Fabr.) (Coleoptera: Cerambycidae). Arthropod Structure & Development, 43(5), 469-475. https://doi.org/10.1016/j.asd.2014.04.005
Maes, J., Berghe, E., Dauber, D., Audureau, A., Nearns, E., Skillman, F., Heffern, D., & Monné, M. (2010a). Catálogo de los Cerambycidae de Nicaragua: Parte I, Parandrinae Prioninae-Aseminae. Revista Nicaragüense de Entomología, 70(1), 102. https://www.zin.ru/animalia/coleoptera/pdf/nicaragua_cerambycidae_2010-s2-cerambycinae.pdf
Maes, J., Berghe, E., Dauber, D., Audureau, A., Nearns, E., Skillman, F., Heffern, D., & Monné, M. (2010b). Catálogo de los Cerambycidae de Nicaragua: Parte II, Cerambycinae. Revista Nicaragüense de Entomología, 70(1), 640. https://www.zin.ru/animalia/coleoptera/pdf/nicaragua_cerambycidae_2010-s2-cerambycinae.pdf
Maes, J., Berghe, E., Dauber, D., Audureau, A., Nearns, E., Skillman, F., Heffern, D., & Monné, M. (2010c). Catálogo de los Cerambycidae de Nicaragua: Parte III, Lepturinae. Revista Nicaragüense de Entomología, 70(1), 31. http://www.bio-nica.info/RevNicaEntomo/70-2010-S1-Prioninae.pdf
Maes, J., Berghe, E., Dauber, D., Audureau, A., Nearns, E., Skillman, F., Heffern, D., & Monné, M. (2010d). Catálogo de los Cerambycidae de Nicaragua: Parte IV, Lamiinae-Disteniinae. Revista Nicaragüense de Entomología, 70(1), 879. https://www.researchgate.net/publication/331640858_Catalogua_illustrado_de_los_Cerambycidae_Coleoptera_de_Nicaragua_-_Parte_III_-Lepturinae
Maes, J., Berghe, E., Dauber, D., Audureau, A., Nearns, E., Skillman, F., Heffern, D., & Monné, M. (2010e). Catálogo de los Cerambycidae de Nicaragua: Parte V, especies exóticas presentes en la colección del museo entomológico de León. Revista Nicaragüense de Entomología, 70(1), 422. https://www.researchgate.net/publication/331640940_Cataloguo_illustrado_de_los_Cerambycidae_Coleoptera_de_Nicaragua_Part_I_-_Parandrinae_-_Prioninae_-_Aseminae
Marco de Lima, B., Cappa, E., Silva, O., Garcia, C., Mansfield, S., & Grattapaglia, D. (2019). Quantitative genetic parameters for growth and wood properties in Eucalyptus "urograndis" hybrid using near-infrared phenotyping and genome-wide SNP-based relationships. PLoS ONE, 14(6), e0218747. https://doi.org/10.1371/journal.pone.0218747
Marini, L., Lindelöw, A., Jönsson, M., Wulff, S., & Schroeder, M. (2013). Population dynamics of the spruce bark beetle: a long-term study. Oikos, 122(12), 1768-1776. https://doi.org/10.1111/j.1600-0706.2013.00431.x
Martínez, C. (2000). Escarabajos longicornios (Coleoptera: Cerambycidae) de Colombia. Biota Colombiana, 1(1), 76-105. https://www.redalyc.org/articulo.oa?id=49110104
Monné, M., & Bezark, L. (2014). Cerambycoidea de México: Cerambycidae, Disteniidae, Vesperidae. Coleoptera Neotropical. http://www.coleopteraneotropical.org/paginas/2_PAISES/Mexico/Cerambycidae/cerambycidae-mex.html
Morales-Morales, C., Aguilar-Astudillo, E., Rosales-Esquinca, M., Quiroga-Madrigal, R., Alonso-Bran, R., & Gutiérrez-Hernández, R. (2012). Cerambícidos (Coleoptera: Cerambycidae) asociados al piñón (Jatropha curcas L), en cinco municipios de la Depresión Central de Chiapas, México. Biota Colombiana, 13(1), 35-46. https://doi.org/10.15517/rev.biol.trop..v72i1.55604
Noerdjito, W. (2012). Dampak kegiatan manusia terhadap keragaman dan pola distribusi kumbang sungut panjang (Coleoptera: Cerambycidae) di Gunung Salak, Jawa Barat. Journal Biologi Indonesia, 8(1), 57-69. https://doi.org/10.14203/jbi.v8i1.3066
Noguera, F., Ortega-Huerta, M., Zaragoza-Caballero, S., González-Soriano, E., & Ramírez-García, E. (2009). A faunal study of Cerambycidae (Coleoptera) from one region with tropical dry forest in México: Sierra de San Javier, Sonora. The Pan-Pacific Entomologist, 85(2), 70-90. https://doi.org/10.3956/2009-04.1
Noguera, F., Zaragoza, S., Rodríguez, A., González-Soriano, E., Ramírez, E., Ayala, R., & Ortega-Huerta, M. (2012). Cerambícidos (Coleoptera: Cerambycidae) del bosque tropical caducifolio en Santiago Dominguillo, Oaxaca, México. Revista Mexicana de Biodiversidad, 83(3), 611-622. https://doi.org/10.7550/rmb.25088
Noguera, F. (2014). Biodiversidad de Cerambycidae (Coleoptera) en México. Revista Mexicana de Biodiversidad, 85(Suppl. 1), 290-297. https://doi.org/10.7550/rmb.32966
Noguera, F., Ortega-Huerta, M., Zaragoza-Caballero, S., González-Soriano, E. & Ramírez-García, E. (2017). Species richness and abundance of Cerambycidae (Coleoptera) in Huatulco, Oaxaca, Mexico; Relationships with phenological changes in the tropical dry forest. Neotropical Entomology, 47, 457-469. http://dx.doi.org/10.1007/s13744-017-0534-y
Novais, S., DaRocha, W., Calderón-Cortes, N., & Quesada, M. (2017). Wood-boring beetles promote ant nest cavities: Extended effects of a twig-girdler ecosystem engineer. Basic and Applied Ecology, 24, 53-59. https://doi.org/10.1016/j.baae.2017.09.001
Programa para los Bosques (PROFOR) (2017). Situación actual y potenciales de fomento de plantaciones forestales con fines comerciales en Colombia. Banco Mundial. https://www.profor.info/sites/profor.info/files/Informe%20Final%20-%20Plantaciones%20Comerciales%20en%20Colombia_1.pdf
Salvador, S., Schumacher, M., Sthal, J., Ludvichak, A., Momolli, D., & Consensa, C. (2019). Effects of soil type in nutrient amount in Eucalyptus urograndis: Macronutrients. Journal of Experimental Agriculture International, 35(3), 1-9. https://doi.org/10.9734/JEAI/2019/v35i330204
Santos, W. & Pereira-Colavite, A. (2017). Ocorrência de Acrocinus longimanus (Linnaeus) (Coleoptera: Cerambycidae) em área urbana, Nordeste do Brasil. EntomoBrasilis, 10(1), 57-59. https://doi.org/10.12741/ebrasilis.v10i1.664
Sataral, M., Atmowidi, T., & Noerdjito, W. (2015). Diversity and abundance of longhorn beetles (Coleoptera: Cerambycidae) in Gunung Walat Educational Forest, West Java, Indonesia. Journal of Insect Biodiversity, 3(17), 1-12. https://doi.org/10.12976/jib/2015.3.17
Singh, A., & Dhillon, G. (2020). Evaluation of Eucalyptus clones under seasonal waterlogging conditions in South-Western Punjab. Current Agriculture Research Journal, 8(2), 98-103. https://doi.org/10.12944/CARJ.8.2.04
Stadelmann, G., Bugmann, H., Wermelinger, B., Meier, F., & Bigler, C. (2013). A predictive framework to assess spatio-temporal variability of infestations by the European spruce bark beetle. Ecography, 36(11), 1208-1217. https://doi.org/10.1111/j.1600-0587.2013.00177.x
Sugiarto, B. & Mardji, D. (2016). Species diversity of cerambycid beetles at reclamation area of coal mining in Berau District, east Kalimantan, Indonesia. Biodiversitas17, 200-207. https://doi.org/10.13057/biodiv/d170129
Toledo, V. (1997). Revisión taxonómica del género Lagocheirus Dejean para México y Centroamérica (Coleoptera: Cerambycidae). Folia Entomológica Mexicana, 101, 1-58. https://doi.org/10.51643/tns.v101.618
Toledo, V., Noguera, F., Chemsak, J., Hovore, F., & Giesbert, E. (2002). The cerambycid fauna of the tropical dry forest of “El Aguacero,” Chiapas, México (Coleoptera: Cerambycidae). The Coleopterists Bulletin, 56(4), 515-532. https://doi.org/10.1649/0010065X
Toledo, V., & Hovore, F. (2005). Notes on the genus Lagocheirus Dejean: Records and descriptions (Coleoptera: Cerambycidae: Lamiinae: Acanthocinini). Zootaxa, 1021(1), 29-36. https://doi.org/10.11646/zootaxa.1021.1.3
Verdugo, P. (2004). Los cerambícidos de Andalucía (Coleoptera: Cerambycidae). Boletín de la Sociedad Andaluza de Entomología, 1, 1-148. https://www.researchgate.net/publication/28201528_Los_cerambicidos_de_Andalucia_Coleoptera_Cerambycidae
Wang, Q. (2002). Sexual selection of Zorion guttigerum Westwood (Coleoptera: Cerambycidae: Cerambycinae) in relation to body size and color. Journal of Insect Behavior, 15(5), 675-687. https://doi.org/10.1023/A:1020747907490
Wang, Q. (2017). Cerambycidae of the world: Biology and pest management (p. 643). CRC Press. https://www.researchgate.net/publication/326892957_Cerambycidae_of_the_World_Biology_and_Pest_Management
Zanuncio, J., Do Nascimento, E., García, J. & Zanuncio, T. (1994). Major lepidopterous defoliators of eucalypt in southeast Brazil. Forest Ecology and Management, 65, 53-63. https://doi.org/10.1016/0378-1127(94)90257-7
Contribución por autor:
Citation:
Author notes
* Autor de correspondencia: Karol Perez García, correo electrónico: kperez@correo.unicordoba.edu.co
Conflict of interest declaration