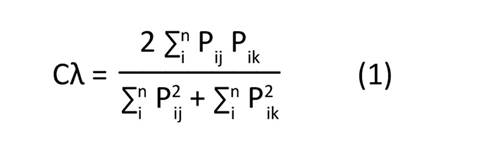Resumen:
Phytelephas macrocarpa es una palmera de relevancia ecológica y económica, especialmente por la exportación de su semilla para artesanías. Sin embargo, enfrenta amenazas por la sobreexplotación y la degradación de hábitat, afectando su sostenibilidad y el sustento de las comunidades locales. Este estudio evaluó su densidad, estructura poblacional y regeneración en el noroeste de la Amazonía peruana, describiendo además su ambiente donde crece. Se establecieron diez parcelas de 1000 m2, registrándose 3824 individuos en cuatro categorías: plántulas (2134), juveniles (436), subadultos (252) y adultos (1002). Las plántulas tuvieron una mayor densidad (2134 ind.10 000m-2), destacando a la localidad de Ahuashillo con el índice de regeneración más alto (4.200). La especie prospera en bosques secos, premontanos y húmedos, hasta los 663 m de altitud, y la mitad de sus poblaciones presentan una regeneración sostenible, mientras que las demás reflejan su manejo actual e histórico, así como cambios en la cobertura vegetal.
Palabras clave: Arecaceae, clases etarias, fragmentación de hábitat, marfil vegetal, yarina.
Abstract:
Phytelephas macrocarpa is a palm tree of ecological and economic importance, especially due to the export of its seeds for handicrafts. However, it suffers from overexploitation and habitat degradation, which affects its sustainability and the livelihoods of local communities. This study evaluated its density, population structure, and regeneration in the northwestern Peruvian Amazon while also describing the environment which it grows. Ten 1000 m2 plots were established, recording 3824 palms in four categories: seedlings (2134), saplings (436), subadults (252), and adults (1002). Seedlings showed the highest density (2134 ind.10 000m-2), with the locality of Ahuashillo standing out for the highest regeneration index (4.200). This species thrives in dry, premontane, and humid forests, up to 663 m in altitude, and half of its populations exhibit sustainable regeneration, whereas the others reflect their current and historical management, as well as changes in vegetation cover.
Keywords: Arecaceae, age classes, habitat fragmentation, vegetal ivory, yarina.
Carátula del artículo
Artículos de investigación
Densidad, estructura poblacional y regeneración de Phytelephas macrocarpa Ruiz & Pavón en el noroeste de la Amazonía peruana
Density, Population Structure and Regeneration of Phytelephas macrocarpa Ruiz & Pavón in the Northwest of the Peruvian Amazon
 Danixa Alessandra Raygada Rengifo
Danixa Alessandra Raygada Rengifo
Instituto de Investigaciones de la Amazonía Peruana, Perú
E. Hanz Rodríguez-Cabrera
Instituto de Investigaciones de la Amazonía Peruana, Perú
Flavia Alexa Urbina-Arévalo
Instituto de Investigaciones de la Amazonía Peruana, Perú
Danter Cachique Huansi
Instituto de Investigaciones de la Amazonía Peruana, Perú
Ángel Alejandro Salazar-Veja
Instituto de Investigaciones de la Amazonía Peruana, Perú
Ires Paula A. Miranda
Instituto Nacional de Pesquisas da Amazônia, Brasil
Kember Mejía
Instituto de Investigaciones de la Amazonía Peruana, Perú
Ángel Martín R. del-Castillo
Estación Experimental Agraria San Ramón, Instituto Nacional de Innovación Agraria. Yurimaguas, Perú. , Perú
Colombia Forestal, vol. 28, no. 2, e22662, 2025
Proyecto Curricular de Ingeniería Forestal, Facultad del Medio Ambiente y Recursos Naturales, Universidad Distrital Francisco José de Caldas.
Received: 08 September 2024
Accepted: 03 April 2025
INTRODUCCIÓN
Las palmeras (Arecaceae) son de gran importancia cultural y económica para las comunidades indígenas y rurales, pues cubren sus necesidades básicas de subsistencia, como lo son la alimentación y la construcción de viviendas (Macía et al., 2011; Martins et al., 2014; Avalos et al., 2022). Además, son un componente estructural importante en los ecosistemas forestales y desempeñan funciones ecológicas fundamentales al suministrar alimento y refugio para varias especies de animales (Kahn & de Granville, 1992; Galetti & Aleixo, 1998; Velazco et al., 2020).
Aun considerando el valor de estas plantas, las palmeras neotropicales se ven afectadas por perturbaciones naturales, incendios, defaunación de dispersores de semillas, herbivoría por parte de animales domésticos y, principalmente, la deforestación, que tiene un efecto de borde en los ecosistemas (Svenning, 1998; Galetti et al., 2006; Armenteras et al., 2013; Lozada & Moraes, 2013; van Lent et al., 2014). Estas circunstancias ralentizan el crecimiento lento de las plántulas e influyen en la mortalidad de individuos adultos y semillas (Wright & Duber, 2001; Montúfar et al., 2011; Soares et al., 2015). Asimismo, la explotación intensiva de las palmeras como productos forestales no maderables está relacionada con la fragmentación de hábitats y ha conllevado cambios significativos en la diversidad genética, la densidad y la estructura de sus poblaciones (Avalos et al., 2013).
Las especies del género Phytelephas se encuentran distribuidas desde Panamá hasta Bolivia, Brasil y Perú (Barfod, 1991; Dransfield et al., 2008; Escobar et al., 2022). Durante los siglos XIX y XX, la época dorada de la tagua, las poblaciones de algunas especies de Phytelephas fueron ampliamente aprovechadas por la industria del marfil vegetal (Barfod, 1989; Bernal, 1992). Las semillas se cosechaban a gran escala para ser exportadas a los mercados de Europa y América del Norte, principalmente para la producción de botones (Barfod et al., 1990; Pülschen, 2000). Sin embargo, después de casi un siglo de cosecha, las poblaciones de Phytelephas en Colombia se habían mantenido en buenas condiciones (Bernal & Galeano, 2013). Esto, debido a la longevidad de las palmeras, que amortiguaron los cambios en el número de plántulas y juveniles, permitiendo que las poblaciones recuperaran su estructura normal (Bernal, 1998).
En el caso de Perú, las semillas de Phytelephas macrocarpa Ruiz & Pavón fueron fuertemente cosechadas durante la época dorada de la tagua, lo que redujo severamente la regeneración de sus poblaciones (Pülschen, 2000; Saldaña & Montoya, 2007; Brokamp et al., 2011; Smith, 2015). Actualmente, sus poblaciones silvestres, situadas en el noroeste de la Amazonía, se encuentran en descenso debido a la pérdida de la cobertura vegetal y la fragmentación del hábitat por la deforestación (Ríos et al., 2018; Quinteros-Gómez et al., 2024) ocasionada por el incremento de los cultivos de palma aceitera y cacao (GIZ, 2016; IBC, 2022).
Esta palmera es considerada como un indicador de suelos fértiles y se desarrolla en zonas de afloramiento de piedra caliza, bosques húmedos de terrenos accidentados, bosques inundables y ecosistemas interandinos de bosques tropicales estacionalmente secos (BTES) (Barfod, 1991; Henderson et al., 1995; Mejía et al., 2014).
Las comunidades rurales conocen a P. macrocarpa con los nombres de yarina, llarina, polponta o poloponta (Kahn & Moussa, 1994; Paniagua-Zambrana et al., 2014; Smith, 2015), y son quienes utilizan las hojas para el techado de casas temporales. Las semillas maduras las emplean para la fabricación de adornos y artesanías, y se alimentan del mesocarpo de los frutos maduros, así como de las semillas inmaduras, que a su vez sirven como componentes medicinales (Albán et al., 2008; Paniagua-Zambrana et al., 2014; Argüello, 2016). Debido a su importancia, la palmera presenta estudios sobre su aprovechamiento y estado de conservación (Saldaña & Montoya, 2007; Pronaturaleza & Amazon Ivory, 2008; Acuña et al., 2022), pero aún se desconocen aspectos sobre la dinámica y la estructura de sus poblaciones (Gonzáles, 1974; Aponte, 2012; Machahua, 2012; Rimachi & Oliva, 2018; Pasapera, 2019; Freitas et al., 2020).
Los estudios sobre la densidad y estructura poblacional de las palmeras son herramientas que ayudan a mejorar el uso de los recursos y proporcionan información importante para el desarrollo de planes de manejo y conservación (Thompson et al., 2009; Moraes et al., 2016). Estos estudios demuestran la historia natural y el estado actual de una población cuando no se dispone de datos a largo plazo (de Souza et al., 2010; Giroldo & Scariot, 2015). También permiten documentar la competencia con otras especies vegetales en sus etapas de crecimiento, además de la competencia por mejores condiciones para la regeneración de sus poblaciones, explicando de esta manera indicadores de reclutamiento y mortalidad (Thompson et al., 2009; Blacutt & Moraes; 2011; Lozada & Moraes, 2013).
Considerando lo anterior, el objetivo de este estudio fue evaluar la densidad, la estructura poblacional y la regeneración de P. macrocarpa en el noroeste de la Amazonía peruana, así como describir las condiciones del ambiente donde se desarrolla la palmera, permitiendo generar información sobre el estado actual y la dinámica de regeneración de sus poblaciones naturales, información necesaria para desarrollar posteriores planes de aprovechamiento y conservación.
MATERIALES Y MÉTODOS
Área de estudio
Este estudio se realizó en localidades situadas en la cuenca central del río Huallaga, al noroeste de la Amazonía peruana, a saber: Ahuashillo (6°26’41” latitud sur - 76°22’13” longitud oeste), Bello Horizonte (6°32’12” latitud sur - 76°17’35” longitud oeste), Caynarachi (6°19’25” latitud sur - 76°17’19” longitud oeste), Churuzapa (6°28’20” latitud sur - 76°29’18” longitud oeste), Leoncio Prado (6°59’37” latitud sur - 76°13’57” longitud oeste), Pucacaca (6°51’29” latitud sur - 76°20’40” longitud oeste), Pucayaquillo (6°3531” latitud sur - 76°13’29” longitud oeste), San Antonio (6°59’23” latitud sur - 76°12’13” longitud oeste), Urcupata (6°23’45” latitud sur - 76°31’14” longitud oeste) y Winge (6°57’01” latitud sur - 76°20’12” longitud oeste) (Figura 1). Esta zona se caracteriza por una topografía irregular moldeada por la altitud, una precipitación media de 1400 mm anuales y una temperatura media anual de 24.4 °C (MIDAGRI, 1989; Rodríguez et al., 2008; Linares-Palomino et al., 2022), así como por presentar bosques húmedos tropicales, bosques estacionalmente secos, bosques transicionales premontanos y andinos, y fragmentos de bosques primarios producto de la deforestación (Ferreyra, 1996; Rodríguez et al., 2008; Mejía et al., 2014). Se seleccionaron estas localidades debido a la presencia de rodales remanentes de yarina, conocidos como yarinales (Coomes & Burt, 1997; Smith, 2015).
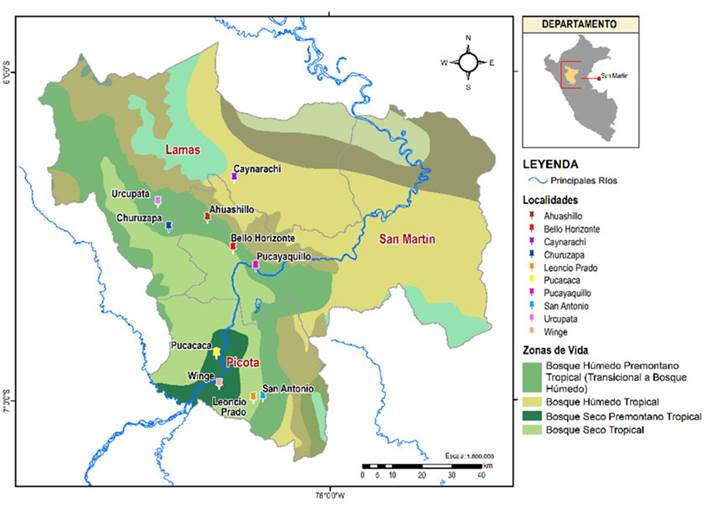 Figura 1
Ubicación de las poblaciones evaluadas de P.macrocarpa
Figura 1
Ubicación de las poblaciones evaluadas de P.macrocarpa
Muestreo de las poblaciones
Debido a la heterogeneidad de la cobertura vegetal en las localidades de muestreo, se emplearon transectos tipo Gentry (1998), que fueron modificados a 50 x 20 m o 0.1 ha (1000 m2) (Moraes et al., 2016). Se elaboró un instrumento de clasificación basado en las divisiones sugeridas por Balslev et al. (2010) y Moraes et al. (2016), donde se consideraron cuatro categorías de crecimiento: plántulas (individuos germinados con eófilos u hojas pinnadas de hasta 1 m de altura), juveniles (individuos acaulescentes de 1 a 2 m de altura, con hojas pinnadas), subadultos (individuos acaulescentes o caulescentes, con rasgos de una palmera adulta sin indicios de estructuras reproductivas) y adultos (individuos acaulescentes o caulescentes que han alcanzado la madurez, donde se evidencian las estructuras reproductivas o restos de ellas). El censo de los individuos se realizó siguiendo movimientos en forma de zigzag (Lozada & Moraes, 2013), con la finalidad evitar el conteo repetido.
Descripción del ambiente de la yarina
Para describir el ambiente donde se desarrolla P. macrocarpa, se utilizó la ficha de toma de datos de campo del OSINFOR (2013), en la que se consideraron, según la clasificación del ONERN (1976), las características del área, el tipo de vegetación de los alrededores (hierba, arbusto o árbol), el tipo de suelo, la altitud y la zona de vida.
Análisis de datos
Los datos se analizaron mediante estadística descriptiva. Se elaboraron gráficos lineales para visualizar la tendencia de la estructura poblacional por localidad, y se determinó el tipo de curva según la clasificación de Peters (1996). Para analizar el patrón de distribución de la estructura poblacional de P. macrocarpa y detectar diferencias significativas entre las categorías de crecimiento, se aplicó un modelo lineal generalizado mixto (GLMM). En dicho modelo, utilizando una distribución de Poisson, se consideró el número de individuos por categoría como variable de respuesta y la categoría de crecimiento como variable de predicción. Se empleó este modelo debido a que los datos no cumplieron el supuesto de normalidad.
La afinidad en las estructuras de las poblaciones fue obtenida por medio de un análisis de conglomerados, a partir de una matriz de similitud y con base en la abundancia relativa de las categorías de crecimiento, mediante el índice de Morisita-Horn (Palacio et al., 2020):
donde:
-
Pij: proporción de la categoría i en muestra j
-
Pik: proporción de la categoría i en la muestra k
-
n: número total de categorías de crecimiento (i=1, 2, 3, …, n)
La densidad poblacional se calculó por medio de la siguiente Ecuación (Moraes et al., 2016):
Para obtener el índice de regeneración en relación con la población reproductiva total, se utilizó el cociente de la densidad de plántulas y de adultos (Michea, 1988), a través de la siguiente Ecuación:
Los análisis estadísticos y los gráficos fueron realizados en el software Rstudio, versión 2024.04.2+764 (RStudio Team, 2024).
RESULTADOS
Estructura poblacional
Se registró un total de 3824 individuos de P. macrocarpa: plántulas = 2134 (55.81 %); juveniles = 436 (11.40 %); subadultos = 252 (6.59 %); adultos = 1002 (26.20 %). Las cuatro categorías de crecimiento fueron significativamente diferentes (Tabla 1).
Tabla 1
Resultados del GLMM. Se presentan diferencias significativas en el promedio de individuos de P. macrocarpa entre categorías de crecimiento.
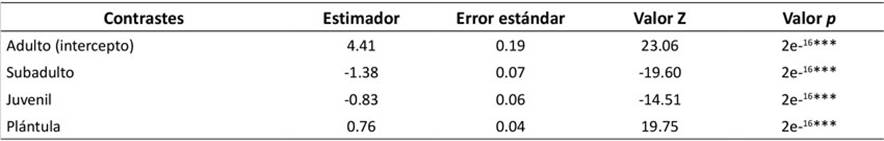 (***) = p < 0.001
(***) = p < 0.001
En las poblaciones de Bello Horizonte, Caynarachi, Leoncio Prado, Pucayaquillo y Urcupata se registró el mayor número de plántulas en comparación con los otros estadíos, observándose una curva de tipo I en forma de J invertida (Figura 2). Entre tanto, las poblaciones de Ahuashillo y San Antonio presentaron una discontinuidad entre los estadíos juvenil y subadulto, por lo que mostraron una curva de tipo II (Figura 2). En el caso de las poblaciones de Churuzapa y Winge, se registraron menores proporciones de juveniles y subadultos, formando una curva en forma de U (Figura 2). La población de Pucacaca fue la única que presentó una curva de tipo III, debido a que el número de palmeras adultas fue mayor que el de las demás categorías de crecimiento (Figura 2).
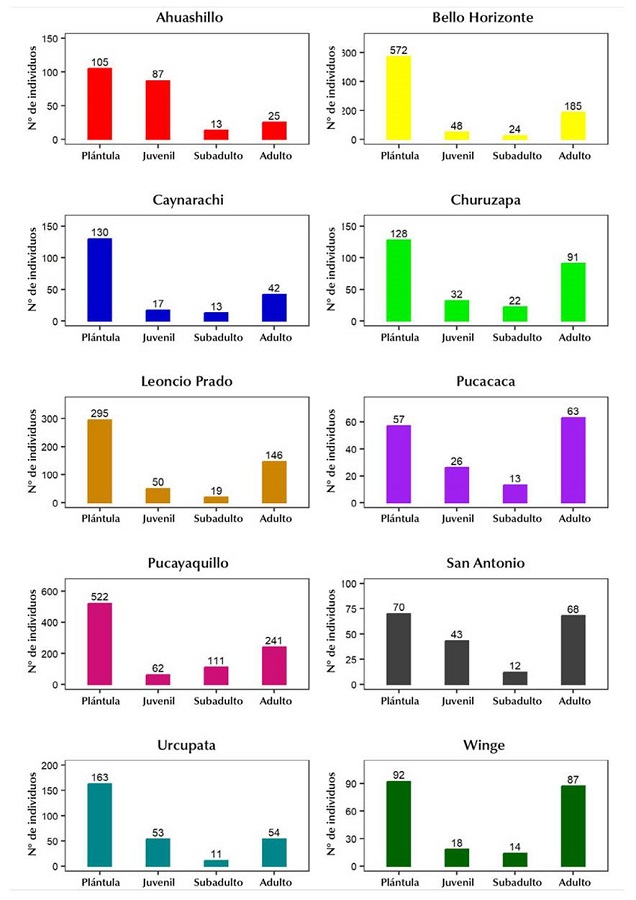 Figura 2
Estructura de las poblaciones evaluadas de P. macrocarpa en el noroeste de la Amazonía peruana
Figura 2
Estructura de las poblaciones evaluadas de P. macrocarpa en el noroeste de la Amazonía peruana
Conglomerado de las poblaciones de yarina
La abundancia relativa de las categorías de crecimiento permitió la formación de tres grupos, con un coeficiente de aglomeración de 0.872 (Figura 3). Se obtuvo la media y la desviación estándar de los tres grupos formados (Tabla 2). En el primer grupo (G1), la media de las plántulas fue más alta que la de las demás categorías. En el segundo (G2), la media de los juveniles fue mayor que la de los otros grupos, y la media de los adultos fue la más baja entre los grupos. El tercer grupo (G3) presentó una media más alta en los adultos, a diferencia de G1 y G2.
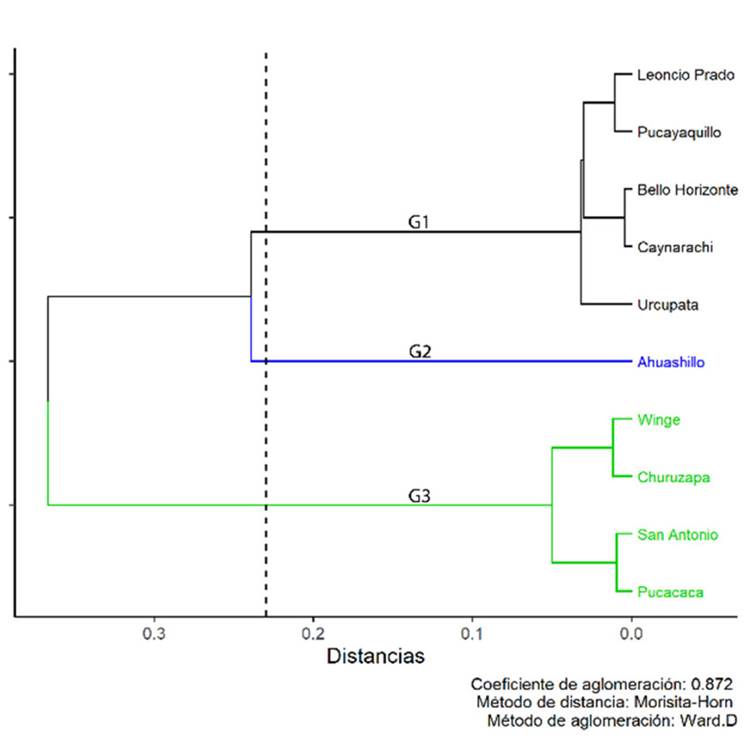 Figura 3
Dendrograma del análisis de conglomerados basado en la similitud de composición por categorías de crecimiento
Figura 3
Dendrograma del análisis de conglomerados basado en la similitud de composición por categorías de crecimiento
Tabla 2
Media y desviación estándar de las categorías de crecimiento según los grupos formados en el análisis de aglomeración 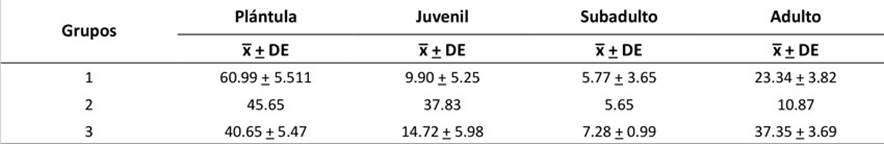

Densidad poblacional
La población de Pucayaquillo presentó la mayor densidad (936 ind.1000m-2), seguida de Bello Horizonte (829 ind.1000m-2) y Leoncio Prado (510 ind.1000m-2). En cambio, San Antonio (193 ind.1000m-2) y Pucacaca (159 ind.1000m-2) reportaron las densidades más bajas (Figura 4). Los individuos correspondientes a las categorías juvenil y subadulto fueron los de menor densidad, con 436 ind.10 000m-2 y 252 ind.10 000m-2 respectivamente, y la categoría plántula fue la de mayor densidad (2134 ind.10 000m-2). Estos valores aumentaron moderadamente al llegar a la etapa adulta (1002 ind.10 000m-2).
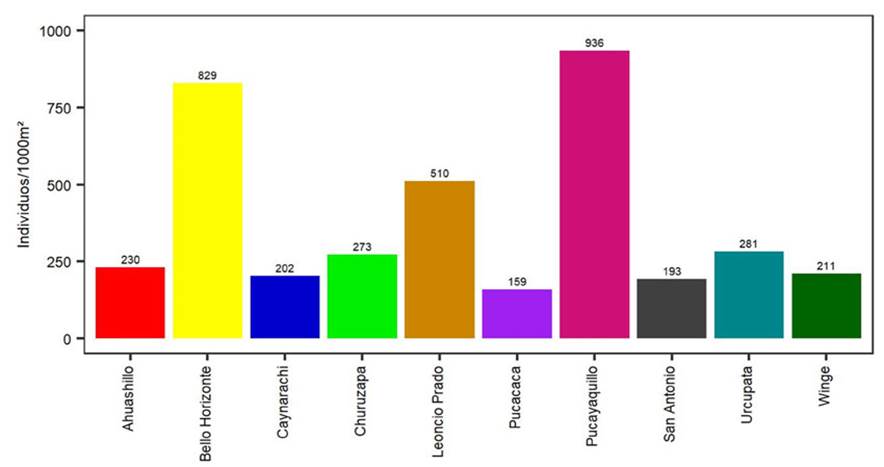 Figura 4
Densidad de las poblaciones evaluadas de P. macrocarpa en 1000 m2
Figura 4
Densidad de las poblaciones evaluadas de P. macrocarpa en 1000 m2
Índice de regeneración
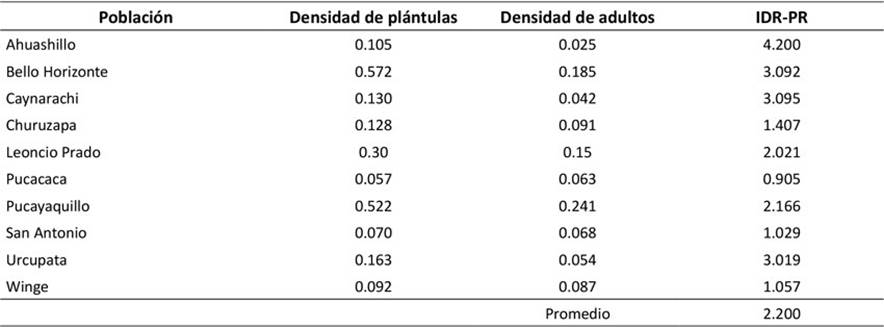
La localidad de Ahuashillo (4.200) presentó el mayor índice de regeneración en relación con la población adulta (IDR-PR), mientras que Pucacaca fue la de menor valor (0.905). El promedio del IDR-PR para las poblaciones de P. macrocarpa evaluadas fue 2.200 plántulas por adulto (Tabla 3). El IDR-PR general para la zona noroeste de la Amazonía peruana fue de 2.129 plántulas por adulto.
Tabla 3
Índice de regeneración de P. macrocarpa en relación con la población adulta
Ambiente de la yarina
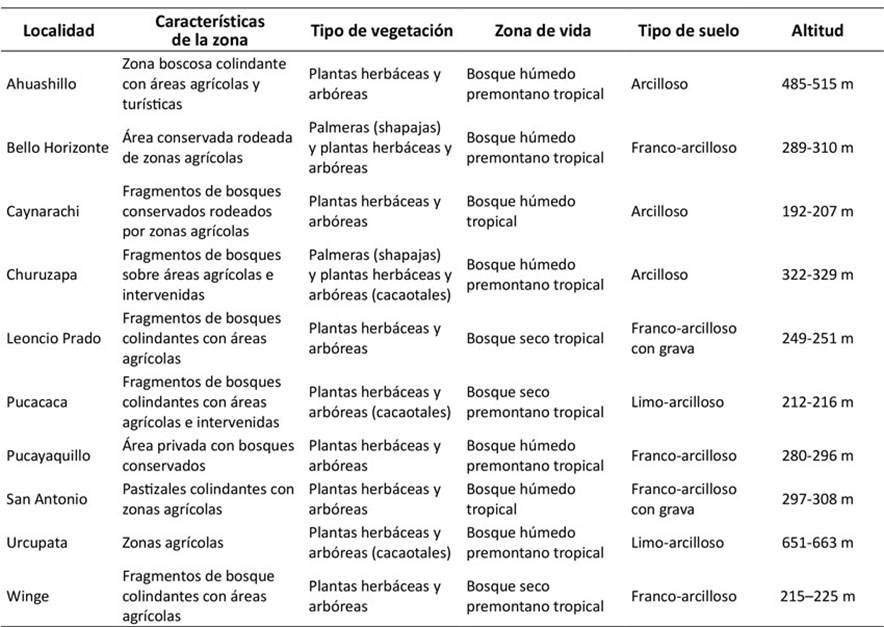
La yarina fue encontrada a altitudes de entre 192 y 663 m, sobre suelos mayormente arcillosos acompañados de una vegetación generalmente herbácea y arbórea. Se encontró que las poblaciones están siendo afectadas por actividades antropogénicas como la deforestación, impulsada por la agricultura, a excepción de algunas zonas como Pucayaquillo y Bello Horizonte, que presentaron bosques conservados. A continuación, en la Tabla 4 se detallan las características del entorno de las diferentes poblaciones de yarina evaluadas.
Tabla 4
Descripción de los ambientes donde fue encontrada la yarina
DISCUSIÓN
Estructura poblacional
La estructura poblacional de P. macrocarpa en las diez localidades se puede organizar en tres grupos según la abundancia de sus categorías de crecimiento. En este contexto, G1 se caracterizó por una mayor densidad de plántulas, mientras que G3 presentó una mayor densidad de adultos, además de la similitud en las condiciones ecológicas de las poblaciones. En estudios previos, la variación en las estructuras poblacionales observadas en Attalea phalerata Mart ex. Spreng y Euterpe precatoria Mart. ha sido atribuida tanto al uso que se le da a cada especie como a las características de sus hábitats antropizados (Giroldo et al., 2012; Avalos et al., 2013).
Las poblaciones de Caynarachi, Pucayaquillo, Bello Horizonte, Urcupata y Leoncio Prado presentaron una curva de tipo I, donde la frecuencia de los individuos disminuía a medida que estos aumentaban de tamaño, formando una curva en forma de J invertida, según el modelo propuesto por Peters (1996). Estas poblaciones fueron encontradas en entornos que iban desde bosques conservados hasta fragmentos de bosque rodeados por zonas agrícolas, con un comportamiento poblacional similar a los registrados para P. tenuicaulis (Barfod) A.J. Hend. en Tumupasa (Bolivia) y para Euterpe precatoria en San Martín de Amacayacu (Colombia), siendo la curva tipo I muy característica en especies con capacidad de regeneración natural en áreas conservadas (Isaza et al., 2014; Moraes et al., 2016). Adicionalmente, se encontró que Acrocomia aculeata (Jacq.) Lodd. ex Mart. presenta el mismo comportamiento autoregenerativo (curva tipo I), a pesar de encontrarse sobre áreas perturbadas de bosques fragmentados, donde la regeneración de su población es atribuida a la dispersión de semillas por parte de animales domésticos (Lozada & Moraes, 2013).
En Churuzapa y Winge se observó una curva en forma de U, caracterizada por una baja presencia de las categorías juvenil y subadulto. Este comportamiento poblacional es afín con lo reportado por Iriartea deltoidea Ruiz & Pavón (Moraes et al., 2016) y Oenocarpus bataua Mart. (Miranda et al., 2009). La baja presencia de juveniles y subadultos sugiere una regeneración continua, pero influenciada por diferentes factores, ecológicos y antropogénicos que afectan temporalmente el crecimiento y la supervivencia de las plantas jóvenes (Escalante, 2000).
Ahuashillo y San Antonio, caracterizadas por niveles discontinuos en la incorporación de nuevos individuos, evidenciaron una curva tipo II. Este tipo de estructura está asociada a perturbaciones físicas o biológicas del ambiente que afectan la estabilidad general de la población (Pinard & Putz, 1992; Peters, 1996) -como en el caso de este estudio- debido a su proximidad con zonas agrícolas y turísticas. Andrade-Erazo y Galeno (2016) documentaron un comportamiento similar en Sabal mauritiiformis (H. Karst.) Griseb. & H. Wendl., donde las perturbaciones producto del cambio en el uso del suelo alteraron la estructura poblacional de la especie.
En Pucacaca se encontró una curva tipo III; hubo mayor predominancia de palmeras adultas por sobre las plántulas debido a la perturbación de los fragmentos de bosque y su cercanía a zonas agrícolas, lo que demuestra la limitada incorporación de nuevos individuos y la baja supervivencia de las etapas iniciales. Con A. phalerata se obtuvieron resultados similares; esta especie también mostró una baja incorporación de nuevos individuos y una limitada presencia de categorías reproductivas en áreas ecológicamente alteradas (Paniagua-Zambrana & Moraes, 2009). En este sentido, la densidad-dependencia podría ser un factor relevante para estos resultados, pues diferentes alteraciones ambientales pueden interrumpir los procesos de crecimiento (Peters, 1996). Sin embargo, Cano-Calderón et al. (2024) evidenciaron que la población de Mauritia flexuosa L.f. tendió hacia niveles bajos de regeneración en un área conservada, considerándola como una población de transición sujeta a eventos reproductivos intermitentes y a la variación de las condiciones ambientales de la zona, las cuales influyeron en el bajo reclutamiento de nuevos individuos.
Densidad poblacional
La densidad poblacional más baja se registró en la localidad de Pucacaca, lo cual puede atribuirse a sus condiciones y perturbaciones ambientales, evidenciadas parcialmente por la presencia de fragmentos de bosque. En este contexto, la fragmentación del hábitat desempeña un papel clave al reducir la conectividad entre parches de vegetación, lo que limita la dispersión de semillas y la adición de individuos jóvenes a la dinámica poblacional (Kahn & de Granville, 1992; Correa-Gómez & Vargas-Ríos, 2009). En este estudio se describieron algunos aspectos ambientales (características del área, interacción con la vegetación, zona de vida, tipo de suelo y altitud) que complementan los resultados, los cuales son congruentes con los hallazgos de Portela y Santos (2014), quienes indicaron que Astrocaryum aculeatissimum (Schott) Burret fue la especie más afectada debido al tamaño de los fragmentos del bosque en su área de estudio.
Si bien en Pucacaca no se observó una ausencia total de individuos juveniles y subadultos, la baja representación de las categorías intermedias se debe a posibles limitaciones en el reclutamiento, especialmente en poblaciones pequeñas, tal y como fue observado en otro grupo de plantas como Araucaria (Souza, 2007).
Al analizar la densidad de los individuos de P. macrocarpa por categoría de crecimiento, se observó un menor registro en las categorías juvenil y subadulto. De acuerdo con Smith y Smith (2000), este comportamiento guarda relación con la curva tipo I, donde se evidencia un mayor número de plántulas que disminuye a medida los individuos van creciendo y madurando. Sin embargo, se podría asumir también que las categorías juvenil y subadulto compiten por recursos como luz, agua y nutrientes con otras plantas o individuos adultos de la misma especie; y también que las palmeras tienen ciclos de vida largos, por lo que pueden tardar en alcanzar su madurez (Uriarte et al., 2005; Kunstler et al., 2016).
Índice de regeneración
Mostacedo y Frederiksen (2001) indican que valores de regeneración inferiores a 0.2 plántulas por adulto demuestran una baja capacidad regenerativa, y que esto puede variar entre especies. En nuestro caso, el bajo índice de regeneración encontrado en Pucacaca (0.905) es mayor que el mencionado. Por otra parte, el índice de regeneración de Geonoma deversa (Poit.) Kunth registró un valor más alto (4.64-5.5 plántulas.adulto-1) (Moraes et al., 2016), superior a la regeneración que observamos en Ahuashillo (cuatro plántulas por adulto), localidad que presentó el mayor valor en este estudio, a pesar de su baja densidad. Otras revisiones de la literatura presentaron valores de regeneración mayores que lo registrado en esta investigación, como es el caso del estudio de Ceroxylon parvifrons (Engel) H. Wendl realizado por Rimachi y Oliva (2018), con 85 plántulas.adulto-1. Sin embargo, los autores sostienen que sus resultados no son definitivos, ya que estos pueden variar de acuerdo con las condiciones del entorno, que podrían transformar una población autoregenerativa en una regresiva.
Miguez et al. (2020) indicaron que la baja regeneración de Ceroxylon pityrophyllum (Mart.) Mart. ex H. Wendl se debe a la fuerte presión de cosecha y a la presencia de ganado y herbivoría. Las perturbaciones ambientales y la explotación de P. macrocarpa para diferentes usos afectan negativamente su regeneración poblacional (Saldaña & Montoya, 2007; Smith 2015). Los resultados de este estudio corroboran estas afirmaciones, pues la baja regeneración de la mitad de las poblaciones de la yarina en la Amazonía peruana está relacionada con la extracción intensiva y otros factores como la fragmentación del hábitat y el estrés hídrico.
Ambiente
Moraes (2014) indicó que P. macrocarpa crece sobre los estratos más bajos de los bosques bolivianos, mientras que Miranda et al. (2001) la encontraron en bosques húmedos de baja altitud y ocasionalmente en áreas inundables de Brasil. En el caso del Perú, esta especie ha sido reportada en estribaciones de los Andes a lo largo de los ríos Huallaga y Ucayali, en los bosques de tahuampa de tierras bajas adyacentes (Barfod, 1991), en bosques de transición en la periferia amazónica con estación seca (Pintaud et al., 2008), en bosques tropicales estacionalmente secos y subhúmedos (Mejía et al., 2014) y en los bosques de las llanuras aluviales (Smith, 2015). Los lugares donde se desarrolla P. macrocarpa en la zona noroeste de la Amazonía peruana estuvieron conformados por bosques secos premontanos tropicales y bosques húmedos premontanos tropicales (transicionales a bosque húmedo), coincidiendo con las descripciones de Barfod (1991), Pintaud et al. (2008), Mejía et al. (2014) y Moraes (2014).
CONCLUSIONES
Este estudio demostró que el 50 % de las poblaciones de P. macrocarpa en el noroeste de la Amazonía peruana presentan una curva tipo I (en forma de J invertida), lo que indica que se mantienen en buen estado de autoregeneración, con patrones estables y sostenibles, y que podrían ser aprovechadas mediante planes adecuados de manejo. Entretanto, las poblaciones con curvas tipo II, III y en forma de U reflejan el manejo actual e histórico, y son producto del cambio de cobertura vegetal causado por la deforestación de estas localidades, lo que las ha transformado en terrenos dedicados a la agricultura y ha puesto en riesgo la dinámica poblacional de la yarina. Por ello, es necesario realizar evaluaciones permanentes encaminadas a recuperar la densidad de las categorías plántula y juvenil. Asimismo, las poblaciones de P. macrocarpa permitieron conformar tres grupos en función de la abundancia relativa y la densidad de plántulas y adultos, las cuales fueron influenciadas por las diferentes condiciones ecológicas de su respectiva localidad y el nivel de fragmentación de su hábitat. Esta palmera se ha adaptado a diferentes ambientes, e.g., bosques húmedos tropicales, bosques húmedos y secos premontanos tropicales, y bosques secos tropicales, a altitudes de hasta 663 m. Por último, se recomienda continuar estudiando la estructura poblacional de P. macrocarpa en la Amazonía peruana para entender mejor su regeneración a largo plazo y la dinámica de sus poblaciones y evitar la disminución de las mismas por la sobreexplotación y la fragmentación de su hábitat.
Agradecimientos
Los autores agradecen a Henrry Chávez, Caleb Gonzales, Nevel Julón, Sandro Ramírez, Karlene Yupe, Julio Raygada, Alan Martín Coral y Geny Rengifo por su apoyo durante las evaluaciones de campo; así como al señor Pascual Guerra por brindarnos los permisos para acceder a los lugares evaluados en el desarrollo de esta investigación.
Referencias
Acuña, M. A., Wipio, G. A., Díaz, E. A., & Guzmán, W. (2022). Recuperación del proceso constructivo para cubiertas ligeras de viviendas en comunidades awajún Condorcanqui, Amazonas-Perú. Revista Científica Dékamu Agropec, 3(2), 33-43. https://doi.org/10.55996/dekamuagropec.v3i2.97
Albán, J., Millán, B., & Kahn, F. (2008). Situación actual de la investigación etnobotánica sobre palmeras de Perú. Revista peruana de biología, 15(3), 133-142. https://doi.org/10.15381/rpb.v15i3.3344
Andrade-Erazo, V., & Galeano, G. (2016). La palma amarga (Sabal mauritiiformis, Arecaceae) en sistemas productivos del Caribe: estudio de caso en Piojó, Atlántico. Acta Biológica Colombiana, 21(1), 141-150. https://doi.org/10.15446/abc.v21n1.47280
Aponte, H. (2012). Efectos de las actividades agropecuarias sobre la estructura poblacional de Astrocaryum perangustatum F. Kahn & B. Millán (ARECACEAE) en Pozuzo (Pasco-Perú). Científica, 9(1), 17-32.
Armenteras, D., González, T. M., & Retana, J. (2013). Forest fragmentation and edge influence on fire occurrence and intensity under different management types in Amazon forests. Biological Conservation, 159, 73-79. https://doi.org/10.1016/j.biocon.2012.10.026
Avalos, G., Emilio, T., Andersen, K. M., & Álvarez-Clare, S. (2022) Editorial: Functional ecology and conservation of palms. Frontiers in Forests and Global Change, 5, 1021784. https://doi.org/10.3389/ffgc.2022.1021784
Avalos, G., Fernández, M., & Engeln, J. T. (2013). Successional stage, fragmentation and exposure to extraction influence the population structure of Euterpe precatoria (Arecaceae). Revista de Biología Tropical, 61(3), 1415-1424. https://doi.org/10.15517/rbt.v61i3.11968
Balslev, H., Eiserhardt, W., Kristiansen, T., Pedersen, D., & Grandez, C. (2010). Palms and palm communities in the upper Ucayali river valley - A little known region in the Amazon basin. PALMS, 54(2), 57-72.
Barfod, A. S. (1989). The rise and fall of vegetable ivory. Principes, 33(4), 181-190.
Barfod, A. S. (1991). A monographic study of the subfamily Phytelephantoideae (Arecaceae). Opera Botanica, 105, 1-73.
Barfod, A. S., Bergmann, B., & Pedersen, H. B. (1990). The vegetable ivory industry: Surviving and doing well in Ecuador. Economic Botany, 44(3), 293-300. https://doi.org/10.1007/BF03183910
Bernal, R. (1992). Colombian palm products. En M. Plotkin y L. Famolare (Eds.), Sustainable Harvest and Marketing of Rainforest Products (pp. 158-172). Island Press.
Bernal, R., & Galeano, G. (2013). Cosechar sin destruir - aprovechamiento sostenible de palmas colombianas. Facultad de Ciencias, Instituto de Ciencias Naturales, Universidad Nacional de Colombia.
Blacutt, E., & Moraes, R. M. (2011). Densidad, estructura y regeneración de la palmera endémica Syagrus yungasensis en Yanamayo - La Asunta (La Paz, Bolivia). Revista de la Sociedad Boliviana de Botánica, 5(1), 5-14.
Brokamp, G., Valderrama, N., Mittelbach, M., Grández, C., Barfod, A. S., & Weigend, M. (2011). Trade in palm products in north-western South America. The Botanical Review, 77, 571-606. https://doi.org/10.1007/s12229-011-9087-7
Cano-Calderón, Y. M., Rodríguez-Hurtado, J. D., Vásquez-Ramos, J. M., Quiñones Méndez, L. M., Castillo-Monroy, E. F., Serrano-Gómez, M., & Torres-Mora, M. A. (2024). Estructura poblacional de Mauritia flexuosa L. f. en la altillanura colombiana, Puerto Gaitán, Meta. Colombia Forestal, 27(1), e20126. https://doi.org/10.14483/2256201X.20126
Coomes, O. T., & Burt, G. J. (1997). Indigenous market-oriented agroforestry: Dissecting local diversity in western Amazonia. Agroforestry Systems, 37, 27-44 https://doi.org/10.1023/A:1005834816188
Correa-Gómez, D. F., & Vargas-Ríos, O. (2009). Regeneración de palmas en bosques nativos y plantaciones del santuario de fauna y flora Otún - Quimbaya (Risaralda, Colombia). Caldasia, 31(2), 195-212.
de Souza, I., Souza, A., Pizo, M., & Ganade, G. (2010). Using tree population size structures to assess the impacts of cattle grazing and eucalypts plantations in subtropical South America. Biodivsersity and Conservation, 19, 1683-1698. https://doi.org/10.1007/s10531-010-9796-y
Dransfield, J., Uhl, N. W., Asmussen, C. B., Baker, W. J., Harley, M. M., & Lewis, C. E. (2008). Genera Palmarum. The Evolution and Classification of Palms. Royal Botanical Garden, Kew.
Escalante, S. (2000). Estudio demográfico y genético de Desmoncus quasillarius (Arecaceae). Informe final SNIB-CONABIO proyecto No. M066. Centro de Investigación Científica de Yucatán, AC.
Escobar, S., Helmstetter, A. J., Montúfar, R., Couvreur, T. L. P., & Balslev, H. (2022). Phylogenomic relationships and historical biogeography in the South American vegetable ivory palms (Phytelepheae). Molecular Phylogenetics and Evolution, 166, 107314. https://doi.org/10.1016/j.ympev.2021.107314
Freitas, L., Dávila, V., Pérez, P., Pezo, R., & Mejía, K. (2020). Estructura poblacional de Mauritia flexuosa y Oenocarpus bataua en tres comunidades de la cuenca alta del Putumayo, frontera Perú - Colombia. Ciencia Amazónica (Iquitos), 8(2), 151-166. https://doi.org/10.22386/ca.v8i2.294
Galetti, M., Donatti, C. I., Pires, A. S., Guimarães, P. R., & Jordano, P. (2006). Seed survival and dispersal of an endemic Atlantic Forest palm: The combined effects of defaunation and forest fragmentation. Botanical Journal of the Linnean Society, 151(1), 141-149. https://doi.org/10.1111/j.1095-8339.2006.00529.x
Gentry, A. H. (1988). Changes in plant community diversity and floristic composition on environmental and geographical gradients. Annals of the Missouri Botanical Garden, 75(1), 1-34.
Giroldo, A. B., Nascimento, A. R. T., Silva, P. P. F., & Pinho Júnior, G. V. (2012). Population structure and density of Attalea phalerata Mart. ex Spreng. (Arecaceae) in a semideciduous forest. Revista Árvore, 36, 637-645. https://doi.org/10.1590/S0100-67622012000400006
Giroldo, A. B., & Scariot, A. (2015). Land use and management affects the demography and conservation of an intensively harvested Cerrado fruit tree species. Biological Conservation, 191, 150-158. https://doi.org/10.1016/j.biocon.2015.06.020
Gonzáles, M. (1974). Estudio sobre la densidad de poblaciones de Aguaje (Mauritia sp.) en Tingo María-Perú. Revista Forestal del Perú, 5, 1-2.
Kahn, F., & de Granville, J.-J. (1992). Palms in forest ecosystems of Amazonia. En F. Kahn & J.-J. de Granville (Eds.), Palms and Forest Management in Amazonia (pp. 155-167). Springer. https://doi.org/10.1007/978-3-642-76852-1_8
Kahn, F., & Moussa, F. (1994). Las palmeras de Perú. Instituto Francés de Estudios Andinos.
Kunstler, G., Falster, D., Coomes, D. A., Hui, F., Kooyman, R. M., Laughlin, D. C., Poorter, L., Vanderwel, M., Vieilledent, G., Wright, S. J., Aiba, M., Baraloto, C., Caspersen, J., Cornelissen, J. H. C., Gourlet-Fleury, S., Hanewinkel, M., Herault, B., Kattge, J., Kurokawa, H., … Westoby, M. (2016). Plant functional traits have globally consistent effects on competition. Nature, 529(7585), 204-207. https://doi.org/10.1038/nature16476
Linares-Palomino R., Huamantupa-Chuquimaco I., Padrón E., La Torre-Cuadros M. A., Roncal-Rabanal M., Choquecota N., Collazos L., Elejalde R., Vergara N., & Marcelo-Peña J. L. (2022). Los bosques estacionalmente secos del Perú: un re-análisis de sus patrones de diversidad y relaciones florísticas. Revista Peruana de Biología, 29(4), e21613. http://dx.doi.org/10.15381/rpb.v29i4.21613
Lozada, S., & Moraes, R. M. (2013). Estructura poblacional del totaí (Acrocomia aculeata, Arecaceae) según presencia de ganado en localidades de Beni y Santa Cruz (Bolivia). Ecología en Bolivia, 48(2), 72-86.
Macía, M. J., Armesilla, P. J., Cámara-Leret, R., Paniagua-Zambrana, N., Villalba, S., Balslev, H., & Pardo-de-Santayana, M. (2011). Palm uses in northwestern South America: A quantitative review. The Botanical Review, 77, 462-570. https://doi.org/10.1007/s12229-011-9086-8
Martins R. C., Filgueiras, T. D. S., & Albuquerque, U. P. (2014). Use and diversity of palm (Arecaceae) resources in central western Brazil. The Scientific World Journal, 1, 1-14 https://doi.org/10.1155/2014/942043
Mejía, K., Pintaud, J.-C., del-Castillo, Á. M. R., Santa Cruz, L., Rojas-Fox, J., Jiménez, V., & Ramírez, R. (2014). Del bosque húmedo al bosque seco: adaptabilidad de las palmeras al cambio climático. En S. Gonzáles, J. J. Vacher & A. Grégoire (eds.). El Perú frente al cambio climático: resultados de investigaciones franco-peruanas (pp. 101-111). Institut de Recherche pour le Développement.
Michea, G. (1988). Estudio poblacional de la palma chilena (Jubaea chilensis) en el sector Ocoa, Parque Nacional La Campana. Medio Ambiente, 9(1), 124-130.
Miguez, S., Paniagua-Zambrana, N., & Moraes, M. (2020). Estado poblacional, aprovechamiento y cadena de comercialización de la palma de ramo (Ceroxylon pityrophyllum, Arecaceae). Ecología en Bolivia, 55(2), 127-139.
Miranda, I. P. de A., Rabelo, A., Bueno, C. R., Barbosa, E. M., & Ribeiro, M. N. S. (2001). Frutos de palmeiras da Amazônia. Ministério da Ciência e Tecnologia, Instituto Nacional de Pesquisas da Amazônia.
Miranda, J., Moraes R., M., & Müller, R. (2009). Estructura poblacional, producción de frutos y uso tradicional de la palmera “majo” (Oenocarpus bataua Mart.) en bosque montano (La Paz, Bolivia). La Revista GAB, 4, 1-10.
Montúfar, R., Anthelme, F., Pintaud, J.-C., & Balslev, H. (2011). Disturbance and resilience in tropical American palm populations and communities. The Botanical Review, 77, 426-461. https://doi.org/10.1007/s12229-011-9085-9
Mostacedo, B., & Fredericksen, T. (Eds.). (2001). Regeneración y silvicultura de bosques tropicales en Bolivia. Proyecto de Manejo Forestal Sostenible. El País.
Moraes, R. M. (2014). Palmeras útiles de Bolivia: las especies mayormente aprovechadas para diferentes fines y aplicaciones. Herbario Nacional de Bolivia, Universidad Mayor de San Andrés, Plural Editores.
Moraes R., M., Vargas, V., Miguez, S., Choque, V., & Sardán, A. (2016). Estructura poblacional de cinco especies de Arecaceae de Tumupasa (La Paz, Bolivia). Revista de la Sociedad Boliviana de Botánica, 9(1), 39-56. http://resbbo.sbb.org.bo/wp-content/uploads/2020/08/4.-Vol.-9-pp-39-56.pdf
Palacio, F. X., Apocada, M. J., & Crisci, J. V. (2020). Análisis multivariado para datos biológicos: teoría y su aplicación utilizando el lenguaje R. Fundación de Historia Natural Félix de Azara.
Paniagua-Zambrana, N. Y., Bussmann, R. W., Vega, C., Téllez, C., & Macía, M. J. (2014). Kampanak se usa para el techo, pero ya no hay: uso y conservación de palmeras entre los Awajún, Amazonas, Perú. Ethnobotany Research and Applications, 14(3), 1-100. https://ethnobotanyjournal.org/index.php/era/article/view/13-004
Paniagua-Zambrana, N. Y., & Moraes R., M. (2009). Hacia el manejo del motacú (Attalea phalerata, Arecaceae) bajo diferente tipo de cosecha (Riberalta, depto. Beni, NE Bolivia): estructura y densidad poblacional. La Revista GAB, 4, 17-23.
Peters, C. (1996). The ecology and management of non-timber forest resources. World Bank Technical Paper.
Pinard, M. A., & Putz, F. E. (1992). Population matrix models and palm resource management. Bulletin de l’Institut Français d’Études Andines, 21(2), 637-649. https://doi.org/10.3406/bifea.1992.1078
Pintaud, J.-C., Galeano, G., Balslev, H., Bernal, R., Borchsenius, F., Ferreira, E., de Granville, J. J., Mejía, K., Millán, B., Moraes, M., Noblick, L., Stauffer, F. W., & Kahn, F. (2008). Las palmeras de América del Sur: Diversidad, distribución e historia Evolutiva. Revista Peruana de Biología, 15(3), 5-28. https://doi.org/10.15381/rpb.v15i3.2662
Portela, R. C. Q., & Santos, F. A. M. (2014). Impact of forest fragment size on the population structure of three palm species (Arecaceae) in the Brazilian Atlantic Rainforest. Revista de Biología Tropical, 62(2), 433-442. https://doi.org/10.15517/rbt.v62i2.9826
Pronaturaleza & Amazon Ivory EIRL. (2008). Plan de manejo de Phytelephas macrocarpa “yarina” en el área de influencia de la comunidad de Veinte de Enero, cuenca Yanayacu Pucate, Reserva Nacional Pacaya Samiria. Instituto de Investigaciones de la Amazonía peruana, Proyecto Focal Bosques. http://www.iiap.org.pe/archivos/publicaciones/Publicacion_2054.pdf
Quinteros-Gómez, Y., Cabrera, D. M., Macedo-Bedoya, J., Santos-Linares, V., & Salinas-Inga, A. (2024). Propagación vegetativa de Vanilla pompona subsp. grandiflora (Orchidaceae) en territorios inundables del Valle del Alto Mayo, Perú. Acta Botánica Mexicana, 131, e2243. https://doi.org/10.21829/abm131.2024.2309
Ríos, S., Liza, R., Smith, C., & Montes, M. R. (2018). Deforestación en el noroeste de la Amazonía peruana. En A. Chirif (Ed.), Deforestación en tiempos de cambio climático (pp. 93-104). IWGIA.
Rimachi, Y., & Oliva, M. (2018). Evaluación de la regeneración natural de palmeras Ceroxylon parvifrons en el bosque andino amazónico de Molinopampa, Amazonas. Revista de Investigación de Agroproducción Sustentable, 2(1), 42-49. https://doi.org/10.25127/aps.20181.383
Rodríguez, F., Limachi, L., Reátegui, F., Escobedo, R., Ramírez, J., Encarnación, F., Maco, J., Guzmán, W., Castro, W., Fachín, L., & Martínez, P. (2008). Las potencialidades y limitaciones del departamento de San Martín: Propuesta de zonificación ecológica y económica como base para el ordenamiento territorial. Gobierno Regional de San Martín, Instituto de Investigaciones de la Amazonía Peruana, Deutsche Gesellschaft für Technische Zusammenarbeit, LuzAzul Gráfica. http://terra.iiap.gob.pe/assets/files/macro/zee-san-martin/Propuesta_ZEE_SanMartin_2009.pdf
Saldaña, J. S., & Montoya, A. E. (2007, mayo 20-25). Potencial de cosecha de Phytelephas macrocarpa (Arecaceae) en tres comunidades de la cuenca Yanayacu- Pucate, Reserva Nacional Pacaya Samiria, Loreto-Perú (Artículo de conferencia). XVII Congreso Venezolano de Botánica, Maracaibo, Venezuela.
Smith, L., & Smith, T. M. (2000). Ecología. Addison Wesley.
Soares, L. A. S. S., Faria, D., Vélez-Garcia, F., Vieira, E. M., Talora, D. C., & Cazetta, E. (2015) Implications of habitat loss on seed predation and early recruitment of a keystone palm in anthropogenic landscapes in the Brazilian Atlantic rainforest. PLoS ONE, 10(7), e0133540. https://doi.org/10.1371/journal.pone.0133540
Svenning, J.-C. (1998). The effect of land-use on the local distribution of palm species in an Andean rain forest fragment in northwestern Ecuador. Biodiversity and Conservation, 7, 1529-1537. https://doi.org/10.1023/A:1008831600795
Thompson, B. N., Moraes, M., & Baudoin, W. M. (2009). Estructura poblacional de la palmera endémica Parajubaea torallyi (Mart.) Burret en zonas aprovechadas del Área Natural de Manejo Integrado El Palmar (Chuquisaca, Bolivia). Ecología en Bolivia, 44(1), 17-35.
Uriarte, M., Canham, C. D., Thompson, J., Zimmerman, J. K., & Brokaw, N. (2005). Seedling recruitment in a hurricane-driven tropical forest: light limitation, density-dependence and the spatial distribution of parent trees. Journal of Ecology, 93(2), 291-304. https://doi.org/10.1111/j.0022-0477.2005.00984.x
van Lent, J., Hernández-Barrios, J. C., Anten, N. P. R., & Martínez-Ramos, M. (2014). Defoliation effects on seed dispersal and seedling recruitment in a tropical rain forest understorey palm. Journal of Ecology, 102(3), 709-720. https://doi.org/10.1111/1365-2745.12216
Velazco, S. J. E., Svenning, J.-C., Ribeiro, B., & Laureto, L. M. O. (2020). On opportunities and threats to conserve the phylogenetic diversity of Neotropical palms. Diversity and Distributions, 27(3), 512-523. https://doi.org/10.1111/ddi.13215
Wright, S. J., & Duber, H. C. (2001). Poachers and forest fragmentation alter seed dispersal, seed survival, and seedling recruitment in the palm Attalea butyracea, with implications for tropical tree diversity. Biotropica, 33(4), 583-595. https://doi.org/10.1111/j.1744-7429.2001.tb00217.x
Notes
Notes
Citación: Raygada Rengifo, D. A., Rodríguez-Cabrera, E. H., Urbina-Arévalo, F. A., Cachique Huansi, D., Salazar-Vega, Á. A., Miranda, I. P. A., Mejía, K., & del-Castillo, Á. M. R. (2025). Densidad, estructura poblacional y regeneración de
Phytelephas macrocarpa Ruiz & Pavón en el noroeste de la Amazonía peruana.
Colombia Forestal, 28(2), e22662.
https://doi.org/10.14483/2256201X.22662
Conflict of interest declaration
Conflicto de intereses Los autores declaran que no existen conflictos de intereses.
Author notes
Contribución por autor D.A.R.R, E.H.R-C, A.M.R.d-C y F.A.U-A. conceptualizaron y definieron la metodología de la investigación. D.A.R.R, E.H.R-C y A.M.R.d-C. realizaron el registro de datos de campo. A.M.R.d-C. y E.H.R-C. se encargaron de la revisión y el tratamiento de los datos. D.A.R.R y E.H.R-C. realizaron el análisis de los datos. Todos los autores participaron en la redacción, edición, discusión y revisión del manuscrito