REFERENCIAS
Antonelli, A. (2020). Kew State of the World's Plants and Fungi. Kew Gardens.
Armenteras, D., Cadena-V., C., & Moreno, R. P. (2007). Evaluación del estado de los bosques de niebla y de la meta 2010 en Colombia. Instituto de Investigación de Recursos Biológicos Alexander von Humboldt.
Arévalo, R., & Betancur, J. (2004). Diversidad de epífitas vasculares en cuatro bosques del sector suroriental de la serranía de Chiribiquete, Guayana Colombiana. Caldasia, 26(2), 359-380.
Avellaneda-Cadena, C., & Betancur, J. (2007). Las plantas vasculares de los afloramientos rocosos de Suesca, Cordillera Oriental Colombiana. Actualidades Biológicas, 29(86), 25-39.
Baptiste, M. P., Benavides-Barrero, L., Betancur, J., Cárdenas, C., Castellanos-Castro, C., Castro, C., & Vallejo, M. I. (2019). Diversidad y estado de conservación de las orquídeas de Cundinamarca. En C. Castellanos-Castro & G. Torres-Morales (Eds.), Orquídeas de Cundinamarca: Conservación y aprovechamiento sostenible (pp. 86-91). Instituto de Investigación de Recursos Biológicos Alexander von Humboldt, Pontificia Universidad Javeriana, Jardín Botánico de Bogotá José Celestino Mutis, Corporación Colombiana de Investigación Agropecuaria Corpoica, Gobernación de Cundinamarca.
Benzing, D. H. (1990). Vascular epiphytes: General biology and related biota. Cambridge University Press.
Betancur, J., Sarmiento, H., Toro-González, L., & Valencia, J. (2015). Plan para el estudio y la conservación de las orquídeas en Colombia. Ministerio de Ambiente y Desarrollo Sostenible, Universidad Nacional de Colombia.
Bruijnzeel, L. A., Kappelle, M., Mulligan, M., & Scatena, F. S. (2010). Tropical montane cloud forests: State of knowledge and sustainability perspectives in a changing world. En L. A. Bruijnzeel, F. N. Scatena, & L. S. Hamilton (Eds.), Tropical Montane Cloud Forests: Science for Conservation and Management. Cambridge University Press
Calderón-Sáenz, E. (Ed.). (2007). Libro rojo de plantas de Colombia. Orquídeas, primera parte. Instituto de Investigación de Recursos Biológicos Alexander von Humboldt, Ministerio de Ambiente, Vivienda y Desarrollo Territorial.
Callaway, R. M., Reinhart, K. O., Moore, G. W., Moore, D. J., & Pennings, S. C. (2002). Epiphyte host preferences and host traits: Mechanisms for species-specific interactions. Oecologia, 132(2), 221-230.
Corporación Autónoma Regional de Cundinamarca (CAR) & Fundación Bachaqueros (2003). Plan de Manejo Ambiental de la Cuchilla de Peñas Blancas y del Subia. CAR-SIBAC.
Cascante-Marín, A., & Trejos Hernández, C. (2019). Diversidad y vulnerabilidad de la flora orquideológica de un bosque montano nuboso del Valle Central de Costa Rica. Lankesteriana, 19(1), 31-55. https://doi.org/10.15517/lank.v19i1.37031
Catling, P. M., & Lefkovitch, L. P. (1989). Association of vascular epiphytes in a Guatemalan cloud forest. Biotropica, 21, 35-40.
Caves, E. M., & Johnsen, S. (2018). AcuityView: an R package for portraying the effects of visual acuity on scenes observed by an animal. Methods in Ecology and Evolution, 9(3), 793-797. https://doi.org/10.1111/2041-210X.12911
CITES. (2022). Checklist of CITES species. Convention on International Trade in Endangered Species of Wild Fauna and Flora. https://checklist.cites.org
Cleef, A. M. (2008). Influencia humana en los páramos. En J. P. Castañeda R. (Ed.), Panorama y perspectivas sobre la gestión ambiental de los ecosistemas de páramo: memorias (Colección Asuntos Ambientales, no. 5, pp. 26-33). Procuraduría Delegada para Asuntos Ambientales y Agrarios.
Cuellar, I. D. (2009). Estrategia ecoturística en el Distrito de Manejo Integrado Cuchilla de Peñas Blancas y el Subia, Municipio El Colegio, Cundinamarca (Trabajo de especialización, Universidad Nacional de Colombia).
Dueñas Gómez, H., & Fernández, A. J. (2009). Sinopsis de la subfamilia Spiranthoideae (Orchidaceae) en Colombia, Parte II. Revista de la Academia Colombiana de Ciencias, 33(127), 157-181.
Durán, C., Rivero, M., & Seemann, P. (2007). Identificación de endomicorrizas en la orquídea nativa Gavilea araucana (Phil.) Correa. Agro Sur, 35(2), 67-69.
Escobar, R., Múner, J. M., & Valdivieso, P. O. (1991). Orquídeas nativas de Colombia (vols. I-VI). Editorial Colina.
Estepa-Ruiz, M. T., & Campos, L. V. (2022). Diversidad de hepáticas epífitas (Marchantiophyta) del sector Suasie en el Parque Nacional Natural Chingaza. Acta Biológica Colombiana, 27(1), 52-60. https://doi.org/10.15446/abc.v27n1.87497
Flanagan, N. S., & Mosquera-Espinosa, A. T. (2016). Una estrategia integrada para la conservación y uso sostenible de las especies nativas de vainilla en Colombia. Lankesteriana: International Journal on Orchidology, 16(2), 201-218. https://doi.org/10.15517/lank.v16i2.26007
Flores-Palacios, A., & García-Franco, J. G. (2001). Sampling methods for vascular epiphytes: Their effectiveness in recording species richness and frequency. Selbyana, 22(2), 181-191.
Flores-Palacios, A., & Valencia-Diaz, S. (2007). Local illegal trade reveals unknown diversity and involves a high species richness of wild vascular epiphytes. Biological Conservation, 136, 372-387.
Fotosíntesis Colombia (2017). Epifitario, un jardín dentro del Bosque. Fotosíntesis Colombia.
Freiberg, M. (1996). Spatial distribution of vascular epiphytes on three emergent canopy trees in French Guiana. Biotropica, 28(3), 345-355.
Galindo, R., & Cadena, J. J. (2000). Caracterización y composición de la vegetación andina en el Santuario de Fauna y Flora Guanentá-alto Rio Fonce, Departamentos de Santander y Boyacá (Tesis de doctorado, Universidad Industrial de Santander).
García-Franco, J. G., & Toledo-Aceves, T. (2008). Epífitas vasculares: bromelias y orquídeas. En R. Manson, V. Hernández-Ortiz, S. Gallina & K. Mehltreter (Eds.), Agroecosistemas Cafetaleros de Veracruz. Biodiversidad, Manejo y Conservación (pp. 69-82). Instituto Nacional de Ecología.
García-González, A., Riverón-Giró, F. B., González-Ramírez, I. S., Escalona Domenech, R. Y., Hernández Montero, Y., & Palacio Verdecia, E. (2016). Ecología y estructura poblacional del endemismo cubano Tetramicra malpighiarum (Orchidaceae), en el Parque Nacional Desembarco del Granma, Cuba. Lankesteriana: International Journal on Orchidology, 16(1), 1-11. https://doi.org/10.15517/lank.v16i1.23379
Gerlach, G. (2020). Notas sobre el género Acineta en los Andes (Colombia, Venezuela, Ecuador y Perú) y una lista incluyendo sus especies. Orquideología, 37(1), 60-84.
Granados-Sánchez, D., López-Ríos, G. F., Hernández-García, M. Á., & Sánchez-González, A. (2003). Ecología de las plantas epífitas. Revista Chapingo. Serie Ciencias Forestales y del Ambiente, 9(2), 101-111.
Haber, W. A. (2000). Plants and vegetation. En N. M. Nadkarni & N. T. Wheelwright (Eds.), Monteverde: Ecology and Conservation of a Tropical Cloud Forest (pp. 39-69). Oxford University Press.
Hágsater, E., & Sánchez, L. (Eds.). (2006). Icones Orchidacearum: Fascicle 8 (Lamin. 801-900). Instituto Chinoin & Herbario AMO.
Hágsater, E. (2013). The genus Epidendrum, Part 10 (Icones Orchidacearum, 14: Plates 1401-1500). En E. Hágsater & L. Sánchez-Saldaña (Eds.), Icones Orchidacearum (Fascículo 14(2)). Instituto Chinoin & Herbario AMO.
Hágsater, E. (2016). The genus Epidendrum, Part 11 (Icones Orchidacearum, fasc. 15(2), láminas 1569-1600). Instituto Chinoin & Herbario AMO.
Hágsater, E., & Ordóñez-Blanco, J. C. (2022). Epidendrum viridialpicola Hágsater et Ordóñez-Blanco, sp. nov. Icones Orchidacearum, 19(2), 35-36.
Hernández-Rosas, J. I. (2001). Ocupación de los portadores por epífitas vasculares en un Bosque Húmedo Tropical del Alto Orinoco, Estado Amazonas, Venezuela. Acta Científica Venezolana, 52(4), 292-303.
Hernández, M., Rosales, N., & Cortés, S. P. (2011). Riqueza y diversidad florística de un bosque de niebla subandino en la Reserva Forestal Laguna De Pedro Palo (Tena-Cundinamarca, Colombia). Revista Facultad de Ciencias Básicas, 7(1), 32-47.
Instituto de Hidrología, Meteorología y Estudios Ambientales (IDEAM). (2010). Leyenda Nacional de Coberturas de la Tierra. Metodología CORINE Land Cover adaptada para Colombia Escala 1:100.000. IDEAM.
Ingram, S., Ferrell-Ingram, K., & Nadkarni, N. M. (1996). Floristic composition of vascular epiphytes in a Neotropical cloud forest, Monteverde, Costa Rica. Selbyana, 17(1), 88-103.
INSAT (2006). Mapa de Cobertura Vegetal y Cuerpos de Agua Del Departamento de Cundinamarca Escala 1:100.000. INSAT Tecnologías en Sistemas de Información Geográfica y Satelital.
Izuddin, M., & Webb, E. L. (2015). The influence of tree architecture, forest remnants, and dispersal syndrome on roadside epiphyte diversity in a highly urbanized tropical environment. Biodiversity and Conservation, 24(8), 2063-2077. https://doi.org/10.1007/s10531-015-0932-6
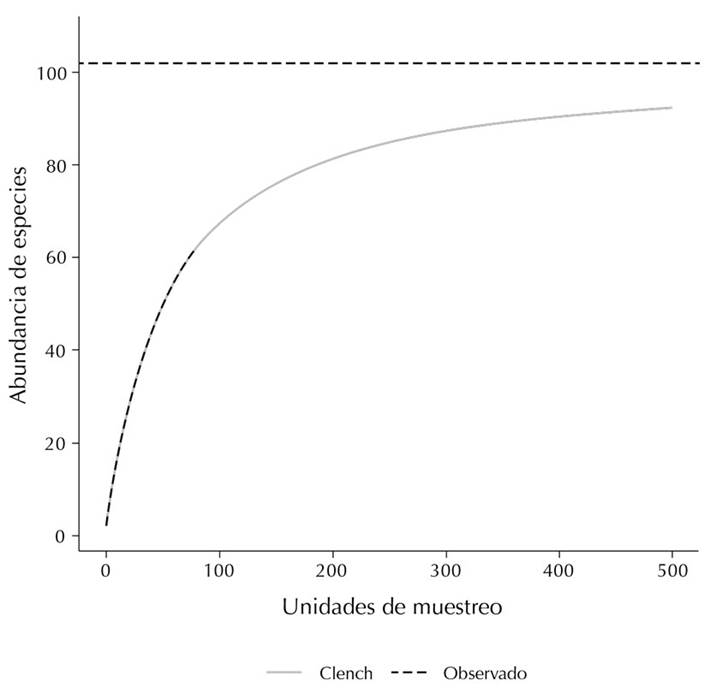
Jiménez-Valverde, A., & Hortal, J. (2003). Las curvas de acumulación de especies y la necesidad de evaluar la calidad de los inventarios biológicos. Revista Ibérica de Aracnología, 8, 151-161.
Johansson, D. R. (1974). Ecology of vascular epiphytes in West African rain forests. Acta Phytogeographica Suecia, 59, 1-136.
Krömer, T., Kessler, M., & Gradstein, S. R. (2007). Vertical stratification of vascularepiphytes in submontane and montane forest of the Bolivian Andes: The importance of the understory. Plant Ecology, 189(2), 261-278.
Krömer, T., García-Franco, J. G., & Toledo-Aceves, T. (2014). Epífitas vasculares como bioindicadores de la calidad forestal: impacto antrópico sobre su diversidad y composición. En J. L. Martínez-Hernández, A. T. Rojas, & M. A. Ramos-Prado (Eds.), Bioindicadores: guardianes de nuestro futuro ambiental (pp. 605-623). Instituto de Ecología A.C., Secretaría de Medio Ambiente y Recursos Naturales.
Lacher, T. E., Boitani, L., & da Fonseca, G. A. B. (2012). The IUCN global assessments: Partnerships, collaboration and data sharing for biodiversity science and policy. Conservation Letters, 5(5), 327-333. https://doi.org/10.1111/j.1755-263X.2012.00249
Leal, P., & León, H. (2014). Inventario taxonómico de especies de la familia Orchidaceae en un área de bosque andino del predio La Sierra, Santuario de Fauna y Flora Guanentá-Alto Río Fonce (Informe técnico). Instituto de Investigación de Recursos Biológicos Alexander von Humboldt.
Le Provost, G., Schenk, N. V., Penone, C., Thiele, J., Westphal, C., Allan, E., Ayasse, M., Blüthgen, N., Boeddinghaus, R. S., Boesing, A. L., Bolliger, R., Busch, V., Fischer, M., Gossner, M. M., Hölzel, N., Jung, K., Kandeler, E., Klaus, V. H., Kleinebecker, T., … Manning, P. (2023). The supply of multiple ecosystem services requires biodiversity across spatial scales. Nature Ecology & Evolution, 7(2), 236-249. https://doi.org/10.1038/s41559-022-01918-5
Lozano-Zambrano, F. H. (Ed.). (2009). Herramientas de manejo para la conservación de biodiversidad en paisajes rurales. Instituto de Investigación de Recursos Biológicos Alexander von Humboldt y Corporación Autónoma Regional de Cundinamarca.
Luer, C. A. (1986a). Icones Pleurothallidinarum II. Systematics of Masdevallia (Orchidaceae). Monographs in Systematic Botany from the Missouri Botanical Garden, 16, 1-63. https://doi.org/10.5962/bhl.title.149317
Luer, C. A. (1986b). Icones Pleurothallidinarum III: Systematics of Pleurothallis (Orchidaceae). Monographs in Systematic Botany from the Missouri Botanical Garden, 20, 1-109. https://doi.org/10.5962/bhl.title.149317
Luer, C. A. (1993). Icones Pleurothallidinarum X. Systematics of Dracula. Monographs in Systematic Botany from the Missouri Botanical Garden, 46, 1-244.
Luer, C. A. (1998). Icones Pleurothallidinarum XVII. Systematics of Pleurothallis subgenus Dracontia (Orchidaceae). Monographs in Systematic Botany from the Missouri Botanical Garden, 72, 65-85.
Mites, M., García-Mozo, H., Galán, C., & Oña, E. (2022). Análisis de la diversidad de orquídeas en la Reserva Pululahua, Ecuador: oportunidades y limitaciones en relación con la conservación de la biodiversidad del bosque nuboso de montaña. Plantas, 11(5), 698. https://doi.org/10.3390/plants11050698
Ministerio de Ambiente y Desarrollo Sostenible. (2024). Resolución 0126 de 2024, por la cual se establece el listado oficial de las especies silvestres amenazadas de la diversidad biológica colombiana continental y marino-costera, se actualiza el comité coordinador de categorización de las especies silvestres amenazadas en el territorio nacional y se dictan otras disposiciones. MinAmbiente.
Mulligan, M., & Burke, S. M. (2005). Global cloud forests and environmental change in a hydrological context: Final report (DFID FRP Project ZF0216). Department for International Development (DFID), Forestry Research Programme.
Nathan, R., & Muller-Landau, H. C. (2000). Spatial patterns of seed dispersal, their determinants and consequences for recruitment. Trends in Ecology and Evolution, 15(7), 278-285.
Nöske, N. (2008). Lichens. En S. Liede-Schumann & S. W. Breckle (Eds.), Provisional checklist of Flora and Fauna of the San Francisco Valley and its Surroundings (Reserva Biológica San Francisco, Province Zamora-Chinchipe, southern Ecuador). Ecotropical Monographs (vol. 4, pp. 101-117). GTÖ.
Ojeda, D. (2001). Ecosistemas. En P. Leyva (Ed.), El Medio Ambiente en Colombia (pp. 279-346). IDEAM.
Ordóñez-Blanco, J. C. (2017). Las orquídeas de un sector de La Calera, Cundinamarca, Colombia. Orquideología, 34(1), 52-64.
Ordóñez-Blanco, J. C. (2021). Distrito de Manejo Integrado (DMI), Cundinamarca, Colombia. Orquídeas del D.M.I. Cuchilla de Peñas Blancas y del Subia. Environmental & Conservation Programs, The Field Museum.
Orejuela-Gartner, J. E. (2010). La conservación de orquídeas en Colombia y un caso en proceso en la cuenca del río Cali, municipio de Santiago de Cali, Valle del Cauca, Colombia. El Hombre y la Máquina, 35, 53-66.
Ortiz, P., & Uribe, C. (2017). Orquídeas, Tesoro de Colombia (tomos 1 y 2). Editorial Da Vinci Publicidad y Medios y Cía. S. En. C.
Padilla, V., Cuevas, G., & Solís, M. (2005). Inga colimana (Leguminosae), una especie nueva del occidente de México. Acta Botánica Mexicana, 72, 33-38.
Parra-Sánchez, E. (2013). Efecto de los patrones del paisaje sobre la diversidad de orquídeas de bosques nublados del Valle del Cauca (Tesis, Universidad Nacional de Colombia).
Parra-Sánchez, E., Pérez-Escobar, O. A., & Edwards, D. P. (2023). Neutral-based processes overrule niche-based processes in shaping tropical montane orchid communities across spatial scales. Journal of Ecology, 111(8), 1614-1628. https://doi.org/10.1111/1365-2745.14140
Parra-Sánchez, E., Freckleton, R. P., Hethcoat, M. G., Ochoa-Quintero, J. M., & Edwards, D. P. (2024). Transformation of natural habitat disrupts biogeographical patterns of orchid diversity. Biological Conservation, 292, 110538. https://doi.org/10.1016/j.biocon.2024.110538
Parrales, H. (2015). Diversidad y distribución altitudinal de orquídeas terrestres del cerro Uyuca (Tesis de licenciatura, Escuela Agrícola Panamericana).
R Core Team (2020). R: A language and environment for statistical computing. R Foundation for Statistical Computing. https://www.R-project.org/
Reina, M., Medina, R., Ávila, F. A., Ángel, S. P., & Cortés, B. R. (2010). Catálogo preliminar de la flora vascular de los bosques subandinos de la Reserva Biológica Cachalú, Santander (Colombia). Colombia Forestal, 13(1), 27-54. https://doi.org/10.14483/udistrital.jour.colomb.for.2010.1.a02
Riofrío-Guamán, M. L. (2016). Estudio de los factores que condicionan, a escala local, la presencia de Epidendrum rhopalostele (Orchidaceae): implicaciones para su conservación (Tesis de doctorado, Universidad Politécnica de Madrid).
Rojas Flórez, C. B., & Sánchez Montaño, L. R. (2015). Estructura espacial de epífitas vasculares en dos localidades de bosque altoandino, Pamplona, Colombia. Caldasia, 37(1), 15-30. https://doi.org/10.15446/caldasia.v37n1.50819
Romero, C., & Alba-López, A. (2005). Taxonomía del género Inga Mill. Estudios en Leguminosas Colombianas, 25, 111.
Rosero, H. M., & Benítez, N. P. (2010). Diversidad de orquídeas epífitas en un bosque húmedo tropical (BH-T) del departamento del Chocó, Colombia. Acta Biológica Colombiana, 15(2), 37-45.
Ruiz-Córdoba, J. P., Toledo-Hernández, V. H., & Flores-Palacios, A. (2014). The effect of substrate abundance in the vertical stratification of bromeliad epiphytes in a tropical dry forest (Mexico). Flora, 209(8), 375-384. https://doi.org/10.1016/j.flora.2014.06.003
Sáenz, E. C., & Camargo, J. C. F. (2003). Especies de los géneros Dracula y Masdevallia (Orchidaceae) en Colombia. Biota Colombiana, 4(2), 187-201.
San Martín, J., Espinosa, A., Zanetti, S., Hauenstein, E., Ojeda, N., & Arriagada, C. (2008). Composición y estructura de la vegetación epífita vascular en un bosque primario de Olivillo (Aextoxicon punctatum R. et P.) en el sur de Chile. Ecología Austral, 18(1), 1-11.
Solano-Gómez, R., Alonso-Hernández, N., Rosado-Ferrer, K., Aguilar-Hernández, M., & García, R. (2008). Diversidad, distribución y estrategias para la conservación de las Pleurothallidinae (Orchidaceae) en Oaxaca. Boletín de la Sociedad Botánica de México, 82, 41-52.
Testé, E., Palmarola, A., & González-Torres, L. R. (2018). Uso del microhábitat por Encyclia pyriformis (Orchidaceae) en la Reserva Ecológica Los Pretiles, Cuba. Lankesteriana: International Journal on Orchidology, 18(2), 93-101. https://doi.org/10.15517/lanke.v18i2.33673
UICN. (2023). Categorías y Criterios de la Lista Roja de la UICN: Versión 3.1. (2da ed). UICN.
Uribe, J. (2015). Cyrtochillums de Colombia. Orquideología, 25(2), 178-182.
Wiese, K. W. (2015). Riqueza de la familia Orchidaceae en la zona de visitantes del Parque Nacional Cerro Azul Meámbar de Honduras. Lankesteriana: International Journal on Orchidology, 15(3), 203-210. https://doi.org/10.15517/lank.v15i3.21667
Williams, L. G., Sosa, V., & Platas, T. (1995). La fragmentación del bosque de niebla, el destino de las orquídeas y el paisaje. En Instituto de Ecología (Eds.), Fragmentación del Bosque de Niebla (pp. 83-93). Instituto de Ecología.
Zelenko, H., & Chase, M. W. (2002). Orchids: The Pictorial Encyclopedia of Oncidium. ZAI Publications.
Zizka, A., Silvestro, D., Vitt, P., & Knight, T. M. (2021). Automated conservation assessment of the orchid family with deep learning. Conservation Biology, 35(3), 897-908. https://doi.org/10.1111/cobi.13616
Zotz, G. (2016). Plants on plants: The biology of vascular epiphytes. Springer.
Notes
Notes
Citación: Ordóñez-Blanco, J. C., López, M., López, A. N., Morales Liscano, G., & Parra Sánchez, E. (2025). Diversidad de orquídeas en diferentes tipos de vegetación en el DMI Cuchilla de Peñas Blancas, Cundinamarca, Colombia.
Colombia Forestal, 28(2), e22574.
https://doi.org/10.14483/2256201X.22574
Conflict of interest declaration
CONFLICTO DE INTERÉS Los autores declaran no tener conflicto de intereses.
Author notes
CONTRIBUCIÓN POR AUTOR J. O.-B.: diseño, toma de datos, identificación de especies, análisis de datos y escritura del documento. M. L.: toma y análisis de datos. A. N.-L.: procesamiento estadístico. G. M.-L.: identificación de especies y toma de datos en campo. E. P.-S.: contribuciones en la consolidación del manuscrito
 Juan Camilo Ordóñez-Blanco phaleno@gmail.com
Juan Camilo Ordóñez-Blanco phaleno@gmail.com