Botany
Morfología de semillas y desarrollo post-seminal de especies de Comanthera (Eriocaulaceae)
Seed morphology and post-seminal development in species of Comanthera (Eriocaulaceae).
Morfología de semillas y desarrollo post-seminal de especies de Comanthera (Eriocaulaceae)
Revista de Biología Tropical, vol. 63, núm. 4, 2015
Universidad de Costa Rica
Recepción: 17 Noviembre 2014
Aprobación: 23 Julio 2015
Financiamiento
Nº de contrato: 5821/11-8
Financiamiento
Nº de contrato: 301692/2010-6
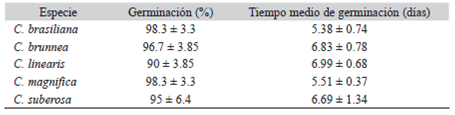
Resumen: Comanthera brasiliana, C. magnifica y C. suberosa (Eriocaulaceae) forman parte de la lista de especies amenazadas de extinción en Brasil, debido a su alto grado de endemismo y a la recolecta indiscriminada. Estas tres especies junto con C. brunnea y C. linearis constituyen el clado de Comanthera magnifica. Se estudió la germinación, la morfología de semillas y el desarrollo post-seminal de las cinco especies, buscando caracterizar morfológica y anatómicamente las semillas y estadios posteriores a la germinación, así como generar datos que ayuden en la conservación de estas especies. Se recolectaron semillas maduras en los "campos rupestres" de Minas Gerais, Brasil. Fueron puestas a germinar cuatro repeticiones de 15 semillas para cada especie en cajas de Petri con papel filtro humedecido con agua destilada, a 25 °C y luz fluorescente. Para el estudio morfológico de las semillas fue usado microscopía electrónica de barrido. Para la anatomía del desarrollo post-seminal, cada estado del desarrollo fue incluido en historresina, cortado con un micrótomo rotativo y analizado con microscopía de luz. Forma y tamaño de las semillas, así como las estructuras de la cubierta seminal permiten diferenciar las especies; a partir de estos caracteres desarrollamos una clave de identificación. Las semillas germinan en altos porcentajes (> 90 %) y rápidamente (< 7 días). La germinación ocurre a través de la protrusión del eje embrionario. Aproximadamente cuatro días después de germinar aparece la primera hoja, seguida por la raíz adventicia; después de 10-15 días se desarrollan la segunda hoja y nuevas raíces adventicias. En condiciones naturales estas especies habitan en suelos derivados de cuarcita y poseen semillas diminutas (tipo polvo) que germinan y se desarrollan rápidamente, incluso más rápido que en otras Eriocaulaceae de suelos mésicos. Estos datos pueden ayudar en la obtención de plántulas de las especies estudiadas, contribuyendo en la conservación de las mismas.
Palabras clave: Comanthera, Eriocaulaceae, morfología de semillas, germinación, siemprevivas, Brasil..
Abstract: Comanthera brasiliana, C. magnifica and C. suberosa (Eriocaulaceae) are included in the Brazilian list of endangered species, due to intensive harvest and micro-endemism. These species form the clade of Comanthera magnifica, along with C. brunnea and C. linearis. Germination, seed morphology and post-seminal development were studied aiming to characterize all five species and to provide information for their conservation. Seeds were collected in "campos rupestres" of Minas Gerais, Brazil. Seed morphology was examined using scanning electron microscope. For germination tests, mature seeds were placed in Petri dishes lined with filter paper dampened with distilled water at 25 °C and fluorescent light; four repetitions with 15 seeds were performed for each species. For the anatomical analysis of post-seminal development, seedlings at different developmental stages were included in historesin, cut with a rotatory microtome and examined under a light microscope. The structure of the seed coat and the shape and size of the seeds were characteristics that allowed species differentiation, and based on the seeds morphological characteristics, an identification key is provided. The seed germination was high (> 90 %) and fast (< 7 days). Germination occured through the axis protrusion of the embryo. Approximately four days after germination, the first leaf develops and is followed by the adventitious roots. After 10-15 days, the second leaf and the new adventitious roots develop. In field conditions, the studied species occur in quartzite soils and their small seeds (dust-like type) germinate faster than the other Eriocaulaceae species (from mesic soils). These results provide reliable information that may contribute to the species management and conservation.
Key words: Comanthera, Eriocaulaceae, seed morphology, germination, everlasting plants, Brazil.
Comanthera Smith es un género recientemente reestablecido (Parra, Giulietti, Gomes de Andrade, & van den Berg, 2010), cuyos representantes pertenecían a las secciones Eulepis y Thysanocephalus de Syngonanthus Ruhland (Giulietti & Hensold, 1991). Actualmente Comanthera subg. Comanthera consta de 26 especies y Comanthera subg. Thysanocephalus de 15 especies (Parra et al., 2010).
Comanthera es el género de mayor importancia económica dentro de Eriocaulaceae (siemprevivas), debido a que sus especies son utilizadas y comercializadas en la artesanía local y de exportación, constituyendo una fuente de ingresos importante en la región de Diamantina, Estado de Minas Gerais, Brasil (Giulietti, Giulietti, Pirani, & Menezes, 1988). La recolecta indiscriminada ha disminuido las poblaciones de varias especies del género (Giulietti & Pirani, 1988), poniendo en peligro de extinción a las que presentan distribución geográfica reducida (Terra Brasilis, 1999).
Estudios sobre morfología de desarrollo post-seminal de Eriocaulaceae (Hare, 1950; Scatena, Menezes, & Stützel, 1993; Kraus, Scatena, Lewinger, & Trench, 1996) muestran que las semillas tienen embrión reducido e indiferenciado y abundante endospermo amiláceo (Dalhgren, Clifford, & Yeo, 1985).
Datos sobre germinación de semillas de representantes de la familia, como Syngo-nanthus spp. (Oliveira & Garcia, 2005; Oliveira & Garcia, 2011; Oliveira, Cruz, Sousa, Morei-ra, & Tanaka, 2014), Paepalanthus Kunth spp. (Kraus et al., 1996) y Comanthera spp. (Scatena, Lemos-Filho, & Lima, 1996) indican que estas semillas requieren de luz para germinar y que la temperatura óptima varía según el microhábitat de la especie.
Los caracteres de la cubierta seminal en Eriocaulaceae son de utilidad taxonómica a nivel genérico (Nair, 1987; Giulietti, Monteiro, Mayo, & Stephens, 1988; Coan, Stützel, & Scatena, 2010; Zona, Davis, Gunathilake, Prince, & Horn, 2012). También diferencian Comanthera subg. Comanthera de C. subg. Thysanocephalus (Barreto, Echternacht, & García, 2013). Los datos embriológicos y de desarrollo de semillas no permiten diferenciar entre los géneros (Ramaswamy, Arekal, & Raju, 1983; Scatena & Bouman, 2001; Coan et al., 2010), pero la ornamentación de la cubierta seminal es específica (Coan et al., 2010). De esa forma, son necesarios estudios de morfología de semillas y desarrollo post-seminal con representantes de Eriocaulaceae dada su importancia taxonómica. Como Comanthera presenta semillas muy pequeñas, el estudio con microscopia de barrido de la cubierta seminal podrá ser útil para su clasificación.
Comanthera brasiliana (Giul.) L.R. Parra & Giul., C. brunnea Echtern., C. linearis (Ruhnland) L.R. Parra & Giul., C. magnifica (Giul.) L.R. Parra & Giul. y C. suberosa (Giul.) L.R. Parra & Giul. constituyen el clado de Comanthera magnifica y tres de ellas están en la lista de especies brasileñas amenazadas de extinción (Biodiversitas, 2007; MMA, 2008). Por tanto, información sobre germinación y desarrollo post-seminal de estas especies es importante para su conservación. De esta forma, se estudió la morfología, la germinación de semillas y el desarrollo post-seminal de las cinco especies del clado Comanthera, teniendo como objetivo caracterizar morfológica y anatómicamente las semillas y los estadios posteriores a la germinación, así como generar datos que ayuden con la conservación de estas especies.
Materiales y Métodos
Se recolectaron en lo mínimo seis capítulos de por lo menos tres individuos diferentes de cada especie que contenían semillas totalmente desarrolladas y libres de los frutos (cápsulas) de Comanthera brasiliana (Dugarte, B. et al. 173), C. brunnea (Dugarte, B. et al. 176), C. linearis (Dugarte, B. et al. 169), C. magnifica (Dugarte, B. et al. 174; Costa, Sam-paio, Ramos, & Fonseca 1 543) y C. suberosa (Dugarte, B. et al. 172, 174) ), en los meses de junio y octubre 2013 en los "campos rupestres" de la región de Ibitipoca (21°42'52.6" S
- 43°53'41" W), Serra do Ambrosio (18°08'10" S - 43°02'46.3" W) y Serra Negra (18°00'15" S
- 42°43'1.2" W), en el Estado de Minas Gerais, sudeste del Brasil. El material de referencia está depositado en el Herbario Rioclarense (HRCB) del Instituto de Biociencias de la Universidad Estadual Paulista.
Los "campos rupestres", son un tipo de vegetación predominantemente herbáceo-arbustiva que pertenece al bioma Cerrado, presenta una estación seca y fría de abril a septiembre y una estación lluviosa y caliente el resto del año (Oliveira-Filho & Ratter, 2002). El suelo del "campo rupestre", derivado de cuarcita, es excesivamente drenado y está expuesto a alta radiación solar y fluctuación diaria de temperatura, con baja disponibilidad hídrica durante la estación seca (Jacobi, Carmo, Vincent, & Stehmann, 2007).
En el laboratorio, para la germinación y el desarrollo post-seminal, fueron utilizadas 60 semillas de cada especie, extraídas de los capítulos bajo el estereoscopio. Se usaron cuatro repeticiones de 15 semillas; puestas a germinar en cajas de Petri, con doble capa de papel de filtro humedecido con agua destilada, bajo las mismas condiciones: en sala climatizada con temperatura de 25 ± 2 °C y luz fluorescente (=32 Limol/m2.s PAR) constantes. El criterio de germinación fue la protrusión del eje embrionario. Las fases de desarrollo fueron acompañadas diariamente con ayuda de un estereoscopio. Se calcularon porcentaje y tiempo medio de germinación (Ranal & Santana, 2006) para cada especie y se compararon mediante Anovas de un factor.
Para el análisis anatómico, las diferentes fases de desarrollo post-seminal fueron fijadas en FAA50 y almacenadas en alcohol al 70 % (Johansen, 1940). Las plántulas fueron deshidratadas y sometidas a infiltración en historre-sina, seguida de inclusión (Feder & OBrien, 1968). Se obtuvieron secciones de 5 uní de grosor con micrótomo rotativo, teñidas con ácido peryódico, reactivo de Schiff (PAS) y azul de toluidina (Feder & OBrien, 1968) y montadas en láminas permanentes con Entellan.
Para el análisis de microscopía de barrido (SEM), semillas maduras fijadas en FAA50 fueron deshidratadas en serie de acetona, secadas a punto crítico (Balzers CPD030 Critical Point) y metalizadas con oro (Balzer 5 CDO 50 Sputter Coater). Se obtuvieron imágenes con el programa "TM 3 000 Application Program".
Resultados
Las semillas son obovadas en C. brasiliana (Fig. 1A), C. linearis (Fig. 1I) y C. magnifica (Fig. 1M) y oblongas en C. brunnea (Fig. 1E) y C. suberosa (Fig. 1Q); miden 0.6-0.8 mm de longitud. La cubierta seminal es rugosa, con proyecciones que forman hendiduras y protrusiones, que son medianamente visibles en C. brasiliana (Fig. 1A, Fig. 1B, Fig. 1C, Fig. 1D) y C. brunnea (Fig. 1E, Fig. 1F, Fig. 1G, Fig. 1H), débilmente visibles en C. linearis (Fig. 1I, Fig. 1J, Fig. 1K, Fig. 1L) y fuertemente visibles en C. magnifica (Fig. 1M, Fig. 1N, Fig. 1O, Fig. 1P) y C. suberosa (Fig. 1Q, Fig. 1R, Fig. 1S, Fig. 1T).
El opérculo es redondo, prominente en C. brasiliana (Fig. 1B), C. magnifica (Fig. 1N) y C. suberosa (Fig. 1R) y no prominente en C. brunnea (Fig. 1F) y C. linearis (Fig. 1J). El hilo es obtuso en C. brasiliana (Fig. 1C) y agudo en las otras especies (Fig. 1G, Fig. 1K, Fig. 1O, Fig. 1S). La cubierta seminal presenta estrías formadas por las paredes anticlinales de la endotesta (Scatena & Bouman, 2001), con distribución longitudinal conspicua en todas las especies (Fig. 1D, Fig. 1H, Fig. 1L, Fig. 1P, Fig. 1T); la distribución transversal es incons-picua en C. magnifica (Fig. 1N, Fig. 1O, Fig. 1P) y conspicua en las otras especies (Fig. 1B, Fig. 1C, Fig. 1D, Fig. 1F, Fig. 1G, Fig. 1H, Fig. 1J, Fig. 1K, Fig. 1L, Fig. 1R, Fig. 1S, Fig. 1T).
Las especies germinaron en altos porcentajes yenun corto período de tiempo (Cuadro 1), sin diferencias significativas entre ellas (Anova, P > 0.08). El porcentaje y el tiempo medio de germinación no fueron diferentes entre las especies (Anova de dos vías, P > 0.052).
Clave para identificar semillas de las especies de Comanthera del ciado de C. magnifica:
1. Semilla obovada
2. Proyecciones de la cubierta seminal fuertemente visibles, estrías longitudinales conspicuas y transversales inconspicuas, ligeramente sinuosas...................................................C. magnifica
2'. Proyecciones de la cubierta seminal visibles, estrías longitudinales y transversales conspicuas, rectas........3
3. Proyecciones de la cubierta seminal visibles, opérculo prominente, hilo obtuso............... C. brasiliana
3'. Proyecciones de la cubierta seminal débilmente visibles, opérculo no prominente, hilo agudo..... C. linearis
1'. Semilla oblonga
4. Proyecciones de la cubierta seminal medianamente visibles, opérculo no prominente............C. brunnea
4'. Proyecciones de la cubierta seminal fuertemente visibles, opérculo prominente................ C. suberosa
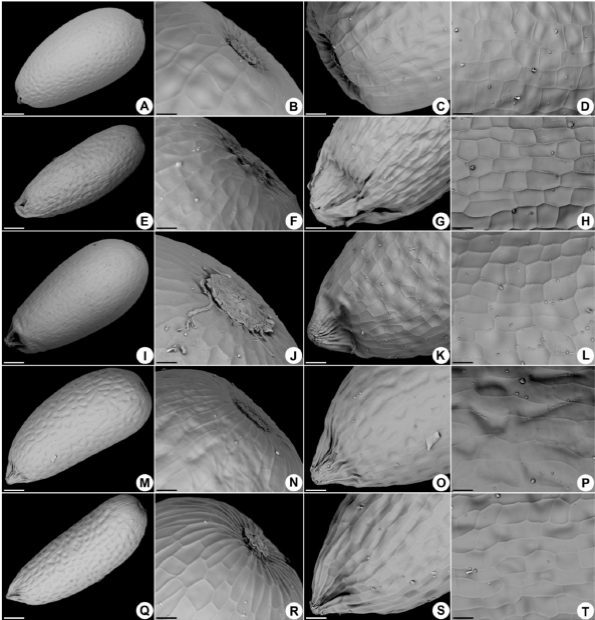
Fig. 1
Morfología de semillas de Comanthera. A-D. C. brasiliana. E-H. C. brunnea. I-L. C. linearis. M-P. C. magnifica. Q-T. C. suberosa. Barras: A, M = 63 um; E, I, Q = 79 um; B, F, J, N, R = 16 um; C, G, K, O, S = 26 um; D, H, L, P, T = 16 um.
La semilla embebida (Fig. 2A) presentó un embrión reducido en comparación con el endospermo (Fig. 2B). De uno a siete días después de la imbibición, se produce la germinación con la protrusión del eje embrionario (Fig. 2C, Fig. 2D), desplazando lateralmente el opérculo (Fig. 2C, Fig. 2E). El eje embrionario presenta células meristemáticas (Fig. 2D), que dan origen a la primera hoja y a la primera raíz adventicia, en dirección opuesta, haciendo la conexión vascular (Fig. 2F). Después de 3.68 ± 1.26 días de la germinación se desarrolla la primera hoja, seguida por el desarrollo de la raíz adventicia (Fig. 2E, Fig. 2F).
Aproximadamente 15 días después de la germinación, se desarrolla la segunda hoja (Fig. 2G), observándose con tejidos diferenciados (Fig. 2H, Fig. 2J), seguida por el desarrollo de la tercera hoja (Fig. 2I) y el desarrollo de nuevas raíces adventicias (Fig. 2I, Fig. 2J), también con los tejidos diferenciados. Durante el crecimiento de la plántula, las células haustoriales del cotiledón, que permanecen en contacto con el endospermo, son más grandes y con núcleos evidentes en comparación con las otras células (Fig. 2F, Fig. 2H).
Discusión
Las características de las semillas de Comanthera, como forma, tamaño, proyecciones de la cubierta seminal y distribución de las estrías en la endotesta, pueden diferenciar las especies del clado de C. magnifica, ayudando en su identificación. En otras Eriocaulaceae, cuyas semillas también son pequeñas, igual que en Comanthera, el levantamiento de caracteres de la cubierta seminal, a través del microscopio electrónico de barrido (SEM), ha demostrado ser útil en la identificación de las especies (Nair, 1987; Giulietti et al., 1988; Zona et al., 2012; Barreto, Echternacht, & García, 2013).
Las diferentes intensidades de las proyecciones de la cubierta seminal en las especies estudiadas están relacionadas con el grado de engrosamiento de las paredes periclinales de las células de la endotesta, carácter importante en su diferenciación. En Eriocaulaceae la endotesta se origina a partir de la capa interna del tegumento externo, el cual presenta células cuyas paredes periclinales y anticlinales se espesan formando diferentes ornamentaciones (Scatena & Bouman, 2001; Coan et al., 2010). Esta capa, junto con el endotegmen, que se origina de la capa interna del tegumento interno, es la cubierta seminal patrón en Eriocaulaceae
Las especies de Comanthera estudiadas mostraron un alto porcentaje de germinación y se clasifican como fotoblásticas positivas. El tamaño pequeño de las semillas, junto con la necesidad de luz para germinar, se consideran estrategias de adaptación para lugares abiertos, expuestos al sol, como en los "campos rupestres", como lo muestran Oliveira y García (2011) y Oliveira et al. (2014). El fotoblastismo positivo puede funcionar como un mecanismo que detecta la presencia o ausencia de luz, evitando que las semillas enterradas germinen, siendo vital para las semillas pequeñas, cuya cantidad de reservas es insuficiente para que alcancen la superficie una vez que germinen en la oscuridad (Vázquez-Yanes & Orozco-Sego-via, 1990; Pons, 2000; Fenner & Thompson, 2005). Las especies de Comanthera estudiadas desarrollan primero las hojas y luego las raíces adventicias, lo que ratifica que el fotoblastismo positivo es un mecanismo que sólo permite la germinación cuando las semillas están en la superficie del suelo, lo que sería ventajoso para las plántulas.
El comportamiento de germinación de las semillas de las especies de Comanthera estudiadas (porcentaje alto y rápida germinación) es similar al de otras especies del género, también típicas de los "campos rupestres", tales como C. elegans (Bong.) L.R.Parra & Giul. (= S. elegans) (Scatena et al., 1996; Oliveira & García, 2005), C. bisulcata (Kórn.) L.R.Parra & Giul. (=S. venustus) (Oliveira & García, 2005), C. nivea (Bong.) L.R.Parra & Giul. (=S. niveus) (Scatena et al., 1996); así como representantes de Syngonanthus (Oliveira et al., 2014) y Velloziaceae J.Agardh (Mercier & Gerreiro-Filho, 1989; García & Diniz, 2003) que crecen simpátricamente.

Fig. 2
Morfología y anatomía del desarrollo post-seminal de Comanthera. A-B. C. suberosa, semillas embebidas. C-D. C. linearis, semillas después de dos días de germinación. E. C. suberosa e F. C. linearis, plántulas después de aproximadamente 10 días de germinación. G. C. suberosa e H. C. brunnea, plántulas después de 15 días de germinación. I. C. magnifica y J. C. suberosa, plántulas cerca de 28 días después de germinar. (ch) células haustoriales, (ej) eje embrionario, (em) embrión, (en) endospermo, (h) hoja, (o) opérculo, (ra) raíz adventicia. Barras: A-C = 0.16 mm; E = 0.20 mm; G-I = 0.60 mm; B = 18.43 urn; D, J = 36.9 urn; F-H = 37.4 urn.
La germinación rápida y el desarrollo post-seminal de las especies de Comanthera estudiadas también son características comunes en otras especies de hábitats abiertos con clima estacional, en los que hay una gran disponibilidad de agua y luz durante la estación lluviosa, seguida por períodos de baja temperatura y poca agua (Swaine & Whitemore, 1988). El rápido desarrollo permite que las plántulas maximicen el tiempo favorable para su establecimiento, formando un sistema de raíces eficientes o acumulando reservas suficientes antes de la llegada de la estación seca, como lo señalan Franco (2002) y Oliveira (2008) para las especies del cerrado, bioma, del cual, el "campo rupestre" forma parte.
Las semillas de tipo polvo (Eriksson & Kainulainen, 2011) sufren adaptaciones que permiten parasitismo o interacción micorrízica por lo menos durante el reclutamiento, pero en las especies de Comanthera estudiadas, así como en Syngonanthus spp. (Oliveira & García, 2011), las reservas nutricionales de las semillas son suficientes para completar la germinación, ya que las hojas fotosintéticas se desarrollan rápidamente. La reducción de tamaño en este tipo de semilla y, por tanto, su rápido desarrollo permite aumentar el número de semillas producidas y también su capacidad de dispersión, favorecida en ambientes altamente heterogéneos que cambian sus características a escalas espaciales pequeñas (Coomes & Grubb, 2003; Turnbull, Coomes, Hector, & Rees, 2004; Eriksson, 2005), como ocurre en los "campos rupestres" (Silveira, Ribeiro, Oliveira, Fernandes, & Lemos-Filho, 2011). De este modo, el tamaño pequeño de las semillas de Comanthera puede aumentar sus posibilidades de encontrar lugares favorables para su establecimiento y reclutamiento, que es la principal limitación en especies del género, según Echternacht et al. (2014).
En las especies estudiadas la germinación de la semilla proyecta un cuerpo cilindrico que se alarga verticalmente, como en otras Erio-caulaceae ya estudiadas, que se denomina eje embrionario (Ramaswamy et al., 1983; Scatena et al., 1993). Poco después de la proyección comienza la diferenciación de la primera hoja y después la diferenciación de la primera raíz adventicia, formando un patrón para la familia (Hare, 1950; Scatena et al., 1993; Kraus et al., 1996; Scatena et al., 1996).
Si comparamos el desarrollo post-seminal de las especies de Comanthera estudiadas con el de otras Eriocaulaceae (Scatena et al., 1993; Scatena et al., 1996), se verifica que tanto la germinación como el desarrollo post-seminal son más rápidos. Las otras Erio-caulaceae crecen en suelos de arena/arcilla o en suelos húmedos; Comanthera en suelo de cuarcita pura. Quizás esta característica es una respuesta adaptativa al ambiente, debido a que estas especies están expuestas a alteraciones ambientales constantes. En el trabajo de campo se observó que donde ellas crecen con mucha frecuencia extraen arena de cuarzo para su uso en la construcción civil. Si esta actividad continúa, podrían extinguirse C. brasiliana y C. suberosa debido a que estas especies están geográficamente restringidas a la Serra do Ambrosio, municipio de Felicio dos Santos, Minas Gerais.
Se concluye que las semillas de las especies estudiadas presentan cubierta seminal con diferentes intensidades de las proyecciones y diferentes formas de las estrías longitudinales y transversales, así como forma del hilo y prominencia del opérculo, características importantes para la identificación de las especies. Además, los datos sobre el desarrollo post-seminal pueden ser útiles para cultivar las especies, su manejo controlado y conservación.
Agradecimentos
Al PEC-PG-CAPES (proc. n° 5821/11-8) y al Consejo Nacional de Desarrollo Científico y Tecnológico (CNPq proc. n° 301692/2010-6) por el financiamiento. Al Instituto Estadual de Florestas/IEF por los permisos de recolecta bajo los números 017/13 y 018/13.
Referencias
Barreto, L., Echternacht L., & García, Q. S. (2013). Seed coat sculpture in Comanthera (Eriocaulaceae) and its implications on taxonomy and phylogenetics. Plant Systematics and Evolution 2991461-1469.
Biodiversitas. (2007). Revisäo das listas das especies da flora e da fauna ameagadas de extingäo do Estado de Minas Gerais. Belo Horizonte: Fundacäo Biodiversitas & Fundacäo ZooBotänica de Belo Horizonte.
Coan, A. I., Stützel, T., & Scatena, V. L. (2010). Comparative embryology and taxonomic considerations in Eriocaulaceae (Poales). Feddes Repertorium 1217-8), 268-284.
Coomes, D. A., & Grubb, P. J. (2003). Colonization, tolerance, competition and seed size variation within functional groups. Trends in Ecology and Evolution 18283-291.
Dalhgren, R. M. T., Clifford, H. T., & Yeo, P. F. (1985). The families of the monocotyledons: structure, evolution and taxonomy. Berlin: Springer-Verlag.
Echternacht, L., Sano, P. T., Bonillo, C., Cruaud, C., Couloux, A., & Dubuisson, J.-Y. (2014). Phylogeny and taxonomy of Syngonanthus and Comanthera (Eriocaulaceae): Evidence from expanded sampling. Taxon 63147-63.
Eriksson, O. (2005). Game theory provides no explanation for seed size variation in grasslands. Oecologia 11498-105.
Eriksson, O., & Kainulainen, K. (2011). The evolutionary ecology of dust seeds. Perspectives in Plant Ecology, Evolution and Systematics, 1373-87.
Feder, N., & O'Brien, T. (1968). Plant microtechnique: some principles and new methods. American Journal of Botany 55123-142.
Fenner, M., & Thompson, K. (2005). Germination. In M. Fenner & K. Thompson (Eds.), The Ecology of Seeds (pp. 110-135). Cambridge University Press.
Franco, A. C. (2002). Ecophysiology of woody plants. In P. S. Oliveira & R. J. Marquis (Eds.), The Cerrados of Brazil: ecology and natural history of a neotropical savanna (pp. 178-197). New York: Columbia University Press.
García, Q. S., & Diniz, I. S. S. (2003). Comportamento germinativo de très espécies de Vellozia da Serra do Cipó (MG). Acta Botanica Brasilica 17(4), 487-494.
Giulietti, N., Giulietti, A. M., Pirani, J. R., & Menezes, N. L. (1988). Estudos em sempre-vivas: importäncia econòmica do extrativismo em Minas Gerais, Brasil. Acta Botanica Brasilica 1(2), 179-193.
Giulietti, A. M., & Hensold, N. (1991). Synonymization of the genera Comanthera and Carptotepala with Syn-gonanthus (Eriocaulaceae). Annals of the Missouri Botanical Garden, 78460-464.
Giulietti, A. M., Monteiro, W. R., Mayo, S. J., & Stephens, J. (1988). A preliminary survey of testa sculpture in Eriocaulaceae. Beiträge zur Biologie der Pflanzen, 62189-209.
Giulietti, A. M., & Pironi, J. R. (1988). Patterns of geographic distribution of some plant species from the Espinhaco Range, Minas Gerais and Bahia, Brazil. In W. R. Heyer & P. E. Vanzolini (Eds.), Proceedings of a workshop on Neotropical Distribution Patterns (pp. 39-69). Rio de Janeiro: Academia Brasileira de Cièncias.
Hare, L. (1950). The structure and development of Erio-caulon septangulare With. Journal of the Linnean Society 53422-448.
Jacobi, C. M., Carmo, F. F., Vincent, R. C., & Stehmann, J. R. (2007). Plant communities on ironstone outcrops: a diverse and endangered Brazilian ecosystem. Biodiversity and Conservation 162185-2200.
Johansen, D. A. (1940). Plant Microtechnique. New York: Mc Graw Hill Book.
Kraus, J. E., Scatena, V. L., Lewinger, M. E., & Trench, K. U. S. (1996). Morfologia externa e interna de quatro espécies de Paepalanthus Kunth (Eriocaulaceae) em desenvolvimento pós-seminal. Boletim de Botànica da Universidade de Säo Paulo 1545-53.
Mercier, H., & Gerreiro-Filho, O. (1989). Germinacäo de Pleurostima fanniei Menezes, Pleurostima rogieri (Hort. ex Moore e Ayres) Menezes e Vellozia alata L.B. Smith (Velloziaceae) sob diferentes condicöes de luz e temperatura. Hoehnea 16195-202.
MMA. (2008). Lista official das espécies da flora Brasileira ameagadas de extingäo. Brasilia: Ministério do Meio Ambiente.
Nair, R. (1987). Taxon omic significance of seed coat morphology in Eriocaulon L. (Eriocaulaceae). Seed Science Technology 15297-310.
Oliveira, P. E. (2008). Fenologia e biologia reprodutiva das espécies de cerrado. In S. M. Sano & S. P. Almeida (Eds.), Cerrado: ambiente e flora (pp. 169-192). Planaltina: EMBRAPA-CPAC.
Oliveira, M. N. S., Cruz, S. M., Sousa, A. M., Moreira, F. da C., & Tanaka, M. K. (2014). Implications of the harvest time on Syngonanthus nitens (Bong.) Ruhland (Eriocaulaceae) management in the state of Minas Gerais. Brazilian Journal of Botany 37(2), 5-103.
Oliveira, P. G., & Garcia, Q. S. (2005). Efeitos da luz e da temperatura na germinacäo de sementes de Syn-gonanthus elegantulus Ruhland, S. elegans (Bong.) Ruhland e S. venustus Silveira (Eriocaulaceae). Acta Botanica Brasilica 193639-645.
Oliveira, P. G., & Garcia, Q. S. (2011). Germination characteristics of Syngonanthus seeds (Eriocaulaceae) in campos rupestres vegetation in south-eastern Brazil. Seed Science Research 2139-45.
Oliveira-Filho, A. T., & Ratter, J. A. (2002). Vegetation physiognomies and woody flora of the Cerrado biome. En P. S. Oliveira & R. J. Marquis (Eds.), The Cerrados of Brazil: ecology and natural history of a neotropical savanna (pp. 91-120).. New York: Columbia University Press
Parra, L. R., Giulietti, A. M., Gomes de Andrade, M. J., & van den Berg, C. (2010). Reestablishment and new circumscription of Comanthera (Eriocaulaceae). Taxon 59(4), 1135-1146.
Pons, T. L. (2000). Seed responses to light. En M. Fenner (Ed.), Seeds: The ecology of regeneration in plant communities (pp. 237-260). 2nd ed. Walligford: CABI Publishing.
Ramaswamy, S. N., Arekal, G. D., & Raju, M. V. S. (1983). Developmental anatomy of seed coat and pericarp in two species of Eriocaulon L. (Eriocaulaceae). Bulletin of the Torrey Botanical Club 110287-291.
Ranal, M. A., & Santana, D. G. (2006). How and why to measure the germination process? Revista Brasileira de Botànica, 291-11.
Scatena, V. L., & Bouman, F. (2001). Embryology and seed development of Paepalanthus sect. Actinocephalus (Körn.) Ruhland (Eriocaulaceae). Plant Biology 3341-350.
Scatena, V. L., Lemos-Filho, J. P., & Lima, A. A. A. (1996). Morfologia do desenvolvimento pós-seminal de Syn-gonanthus elegans e S. niveus (Eriocaulaceae). Acta Botanica Brasilica 1085-91.
Scatena, V. L., Menezes, N. L., & Stützel, T. (1993). Embriology and seedling development in Syngo-nanthus rufipes Silv. (Eriocaulaceae). Beitrage zur Biologie der Pflanzen 67333-343.
Silveira, F. A. O., Ribeiro, R. C., Oliveira, D. M. T., Fernandes, G. W., & Lemos-Filho, J. P. (2011). Evolution of physiological dormancy multiple times in Melastomataceae from Neotropical montane vegetation. Seed Science Research 22(01), 37-44.
Swaine, M. D., & Whitemore, T. C. (1988). On the definition of ecological species groups in tropical rain forests. Vegetatio 75(1-2), 81-86.
Terra Brasilis, Instituto. (1999). Projeto sempre-vivas: subsidios para seu uso sustentado. Publicado por Instituto Terra Brasilis.
Turnbull, L. A., Coomes, D., Hector, A., & Rees, M. (2004). Seed mass and the competition/colonization trade-off: a sowing experiment. Journal of Ecology 9297-109.
Vàzquez-Yànes, C., & Orozco-Segovia, A. (1990). Ecological significance of light controlled seed germination in two contrasting tropical habitats. Oecologia 832171-175.
Zona, S., Davis, P., Gunathilake, L. A. A. H., Prince, J., & Horn, J. W. (2012). Seeds of Eriocaulaceae of the United States and Canada. Castanea 77(1), 37-45.