Artículos
Nesting habits of Centris (Hemisiella) dichrootricha (Hymenoptera: Apidae) in the Northern Cerrado of Brazil
Hábito de nidificación de Centris (Hemisiella) dichrootricha (Hymenoptera: Apidae) en el Cerrado Norte de Brasil.
Nesting habits of Centris (Hemisiella) dichrootricha (Hymenoptera: Apidae) in the Northern Cerrado of Brazil
Revista de Biología Tropical, vol. 64, no. 3, 2016
Universidad de Costa Rica
Received: 25 November 2015
Accepted: 04 April 2016
Abstract: The Neotropical bee Centris (Hemisiella) dichrootricha is a solitary bee that nests in pre-existing cavities that occur in the rain forest. This study describes the nesting biology of C. dichrootricha and its preference for nesting in Cerrado and gallery forest habitats. The study was conducted from January 2012 and December 2013, in Mirador State Park in the municipality of Formosa da Serra Negra, Maranhão State, Brazil. For this, wooden trap-nests of 6, 8, 10, 12, 14 and 16 mm in diameter were used; a total of 300 trap-nests were placed in the gallery forest and Cerrado areas, respectively. Traps were monitored monthly and all completed nests were collected and replaced with empty ones. The nests were then taken to the laboratory to analyze bee development and emergence, nests characteristics and parasites presence. The species used 29 of the trap-nests, which had diameters of 8, 10, 12 and 14 mm. A total of 87 C. dichrootricha specimens emerged. The nests were parasitized by two bee species, Mesocheira bicolor (Apinae) and Coelioxys sp. (Megachilinae), and one fly species, Antrax sp. (Diptera). The highest nesting incidence of 72.4 % was observed in the gallery forest, whereas only 27.6 % in the Cerrado; this difference in habitat use was significant (χ² = 5.56; p < 0.05; DF = 1). For the nests that were built in the gallery forest, 80.9% of the soil originated from the Cerrado. The females were significantly larger than the males (F1, 76 = 595.19; p < 0.001). There were 11 pollen types that belonged to six families. Pollen of the family Malpighiaceae was most frequently used, with four species represented (Byrsonima crassifolia, B. rotunda, B. spicata and Heteropterys sp.). C. dichrootricha showed a preference for nesting in cavities of various diameters in gallery forest sites. The present study provides a novel description of the nesting habits and biology of C. dichrootricha in habitats of Central/Southern Maranhão. C. dichrootricha primarily used resources from the Cerrado, including soil to build their nests, pollen and floral oils; we concluded that gallery forest and Cerrado areas are intrinsically related to the maintenance of local populations of this species. Rev. Biol. Trop. 64 (3): 1041-1056. Epub 2016 September 01.
Key words: gallery forest, solitary bee, Centris, pollen analysis, trap-nests.
Resumen: Centris (Hemisiella) dichrootricha es una especie de abeja solitaria neotropical, que anida en cavidades preexistentes y vive en la selva tropical. El presente estudio describe la biología de anidación de C. dichrootricha y su preferencia de nidificación por áreas en Cerrado y Bosque de Galería. El área de estudio se localiza en el Parque Estadual de Mirador, municipio de Formosa da Serra Negra, Maranhão. El método empleado fue el de nidos trampa de madera de diferentes diámetros, 6, 8, 10, 12, 14 y 16 mm, colocándose 300 nidos trampa por área. Las trampas fueron monitoreadas mensualmente y todos los nidos terminados fueron recogidos y sustituidos por otros vacíos. Los nidos completos fueron llevados al laboratorio para analizar el desarrollo de la abeja, la emergencia, las características de los nidos y la presencia de parásitos. La especie ocupó 29 nidos de los diámetros de 8, 10, 12 y 14 mm. Emergieron 87 individuos de C. dichrootricha. Los nidos fueron parasitados por tres especies: Mesocheira bicolor (Apinae), Coelioxys sp. (Megachilinae) y Antrax sp. (Diptera). En el Bosque de Galería se presentó la mayor nidificación (72.4 %) y en el Cerrado solamente 27.6 %, de esta forma, hubo diferencias significativas entre los hábitats (χ² = 5.56; p < 0.05; G. L.= 1). Los nidos fundados en el bosque de galería presentaron 80.9 % de sedimento originario del Cerrado. Las hembras fueron significativamente más grandes que los machos (F1, 76 = 595.19; p < 0.001). Hubo 11 tipos de polen pertenecientes a seis familias. Los tipos de polinización de la familia Malpighiaceae fueron los más representativos, con cuatro especies: Byrsonima crassifolia, B. rotunda, B. spicata y Heteropterys sp. C. dichrootricha mostró preferencia por nidificar en cavidades con diámetros variados y en el bosque de galería. El presente estudio contribuye con la descripción de los hábitos de nidificación y la biología de la especie para las áreas de Cerrado en la región centrosur de Maranhão. C. dichrootricha utilizó en su mayoría recursos provenientes del Cerrado, tanto sedimento para construir el nido, como material polínico y aceite floral. Estós datos indican que el bosque de galería y el Cerrado están intrínsecamente relacionados con el mantenimiento y conservación de C. dichrootricha.
Palabras clave: análisis de polen, abejas solitarias, Centris, bosque de galería, nidos trampa.
Bees of the tribe Centridini (Hymenoptera, Apidae) are solitary and distinguished by morphological adaptations for the collection of floral oils (Neff & Simpson, 1981), which makes them key pollinators of plant families that produce floral oils (Buchmann, 1987; Albuquerque & Rêgo, 1989; Vinson, William, Frankie, & Shrum, 1997; Mello, Bezerra, & Machado, 2013). Several plant families are important oil sources for this group of bees, including Calceolariaceae, Iridaceae, Krameriaceae, Malpighiaceae, Orchidaceae, Plantaginaceae and Solanaceae (Vogel, 1974; Simpson, Neff, & Seigler, 1977; Machado, 2004; Renner & Schaefer, 2010; Martins, Aguiar, & Alves-dos Santos, 2013; Murúa, Cisterna, & Rosende, 2014). Species of the genus Centris Fabricius, 1804 are distributed throughout the American continent and can be found from arid regions to equatorial rain forests (Zanella, 2002). These species are capable of nesting in different habitats and can build their nests on the ground (Aguiar & Gaglianone, 2003; Rêgo, Albuquerque, Ramos, & Carreira, 2006), in termite nests (Gaglianone, 2001; Ramos, Mendes, Albuquerque, & Rêgo, 2007b) and in pre-existing cavities (Krombein, 1967; Coville, Frankie, & Vinson, 1983).
There are three subgenera of Centridini that nest in pre-existing cavities: Hemisiella, Heterocentris and Xanthemisia (Coville et al., 1983). These bees can be divided into two groups based on their use of building material for nests (Vinson, Frankie, & Williams, 1996), and this feature is one of the taxonomic criteria used to verify the phylogenetic affinity of bees (Michener, 2007). The group that includes Heterocentris and Xanthemisia uses wood chips for the construction of cells, and the Hemisiella subgenus builds their nests with sand (Vinson et al., 1996). Floral oil is another resource used to build nests, and it functions as a coating material for internal and external surfaces of the cells of the nests (Neff & Simpson, 1981). In addition, species such as C. analis, C. bicornuta, C. tarsata, C. trigonoides and C. vittata use the oil to coat the nest plug (Vinson et al., 1996; Jesus & Garófalo, 2000; Aguiar & Garófalo, 2004; Aguiar, Garófalo, & Almeida, 2006) to prevent attacks by natural enemies (Vinson & Frankie, 2000; Jesus & Garofalo, 2000).
In the Hemisiella subgenus, the nest cells are constructed horizontally (C. tarsata) or positioned vertically in a linear series (C. nitida and C. trigonoides) (Silva, Viana, & Neves, 2001; Aguiar et al., 2006; Buschini & Wolff, 2006; Mendes & Rêgo, 2007; Vinson, Frankie & Cônsoli, 2010). In C. tarsata nests, two parallel rows of cells may be found (Mendes & Rêgo, 2007; Mesquita, Vilhena, & Augusto, 2009). For larval feeding, the bees use pollen, nectar and oil (Aguiar & Garófalo, 2004; Aguiar et al., 2006; Vinson et al., 2010), and the pollen and oil resources mainly include pollen from Byrsonima (Mendes & Rêgo, 2007; Pemberton & Liu, 2008; Ramos, Albuquerque, & Rêgo, 2010; Dórea, Aguiar, Figueroa, Lima, & Santos, 2013).
Previous studies of the bees of the subgenus Hemisiella Moure, 1945, which nest in pre-existing cavities, have investigated five species: Centris (Hemisiella) dichrootricha (Moure, 1945), Centris (Hemisiella) nitida Smith, 1874, Centris (Hemisiella) tarsata Smith, 1874, Centris (Hemisiella) trigonoides Lepeletier, 1841 and Centris (Hemisiella) vittata Lepeletier, 1841 (Camillo, Garófalo, Serrano, & Muccillo, 1995; Morato, Garcia, & Campos, 1999; Garófalo, Martins, & Alves-dos Santos, 2004; Aguiar et al., 2006; Vinson et al., 2010). Of these, C. nitida occurs in Costa Rica (Vinson et al., 2010), C. tarsata is the most prevalent in Brazilian biomes (Viana, Silva, & Kleinert, 2001; Aguiar & Garófalo, 2004; Buschini, 2006; Mendes & Rêgo, 2007; Mesquita & Augusto, 2011), and C. vittata is the second-most prevalent in trap-nests (Camillo et al., 1995; Mesquita et al., 2009; Mesquita & Augusto, 2011) followed by C. trigonoides (Aguiar et al., 2006). However, only one study of C. dichrootricha in the Amazon rainforest has been reported (Morato et al., 1999).
Studies on the biology of solitary bees in the Cerrado areas in Northeastern Brazil are incipient, and they include the occurrence of C. vittata nesting in natural trunk cavities of Astronium sp. (Anacardiaceae) (Ramos, Rêgo, & Albuquerque, 2007a; Ramos et al., 2010) and the biology of C. tarsata nesting in artificial cavities (Mendes & Rêgo, 2007). In such studies, trap-nests are important for bee species within a given study location to determine bee biology and behavior (Camillo et al., 1995). The aim of this study was to provide data on the biology and nest architecture of C. dichrootricha, including its seasonality, sex ratio, parasitism rate, and food source origin, and to assess its preference for nesting in Cerrado and gallery forests.
Materials and methods
Study area:
The study area is located at Base Geraldina in Mirador State Park (Parque Estadual do Mirador), Formosa da Serra Negra Municipality, in the Central/Southern region of Maranhão State (6º 37’ 56.29’’ S - 45º 53’ 47.25’’ W). The park comprises an area of approximately 438 000 ha and predominantly presents vegetation of the Neotropical Savannas, including formations of Cerrado stricto sensu, cerradão and gallery forest. The terrain is flat, and the soil is a red-yellow latosol. The climate is subhumid and semiarid, with an annual rainfall of 1 200 mm. The rainy season lasts from October to May, and the dry season from June to September. The mean maximum temperatures range from 31.4 °C to 33 °C, and the mean minimum temperatures range from 19.5 ºC to 21 ºC (IBGE, 1998).
Sampling:
Wood trap-nests consisting of one cut piece (28 x 28 x 150 mm) drilled longitudinally were used to capture the bees. The cavities drilled in the trap-nests were 6, 8, 10, 12, 14 and 16 mm in diameter and 110 mm deep. The trap-nests were distributed in sampling stations. Each sampling station included 12 trap-nests, with two for each opening diameter. The sampling stations were distributed in two 1 ha areas, with five transects per area and five sampling stations per transect, for a total of 600 traps (300 trap-nests/area). One area was composed of Cerrado stricto sensu vegetation, and the other was composed of gallery forest. The sampling stations were tied to the tree trunks at a height of 1.5 m off the ground, and their entrance orientation was random.
The trap-nests were checked monthly from January 2012 to December 2013. All finished nests that had been constructed by the bees were removed and replaced by other nests of the same diameter, and the bee-constructed nests were taken to the laboratory to be monitored and photographed. Emerging individuals were killed and labeled with nest identifications, birth order of bees or parasites, sex and date of emergence. Some nests were monitored (n= 3) to observe the bee development and larval food composition characteristics. After the emergence of individuals, the nests were opened to determine the number of cells, presence or absence of vestibular cells, type of material that was used to built them, total length of the intranest cavity, and length and width of the cells. Pollen material was removed for analysis and identification. Climate data (rainfall, relative humidity and temperature) were measured with the help of a pluviometer and a datalogger.
The nest construction rate, which was calculated as the ratio of the number of constructed nests to the number of available nests (x 100), was determined to compare the abundance of constructed nests. The cell volume was calculated using the cylinder volume formula [(width of the cell)² x (length of the cell) x (π= 3.14)]. Cell measurements were performed with a digital caliper, and the maximum head width (HW) was measured using Axion-Vision software (Carl Zeiss, Thornwood, New York, United States), and a camera (ICc1) attached to a Zeiss Scope A1 stereomicroscope. The bees were deposited in the Bee Collection of the Laboratory of Bee Studies (LEACOL), Federal University of Maranhão (UFMA).
The residue from 20 C. dichrootricha nests was extracted for pollen analysis after the emergence of adults in 2012. Pollen material was extracted using Erdtman’s acetolysis method (Erdtman, 1960). Four slides were made for each nest after the samples had been processed. Counting was based on the positioning of four 5 mm² quadrants that were drawn on the cover slip with a permanent marker (CD marker). A total of 75 pollen grains were counted on each quadrant for a total of 300 grains for each slide and 1 200 grains for each nest sample. The pollen types were identified by comparison with a pollen reference collection of the local flora. All the plants in bloom found in the study area were collected monthly, exsiccates were subsequently produced and sent to the Brazilian Enterprise for Agricultural Research - Eastern Amazon (Empresa Brasileira de Pequisa Agropecuária-EMBRAPA Amazônia Oriental) for identification at the species level, and vouchers were stored in the Herbarium of the Federal University of Maranhão (MAR). In addition, microscope slides prepared by Erdtman’s acetolysis method (1960) were stored at the Palynotheca.
All the continuous variables were tested for normality using the Shapiro-Wilk test (p > 0.05) to investigate parametric test assumptions. A multivariate analysis of variance (MANOVA) was performed to determine if significant differences occurred in the maximum head width and cell volume in relation to sex and habitat. The non-parametric KruskalWallis test was used to test if significant differences occurred in the maximum head width and volume in relation to nest diameter. The MannWhitney test was used to identify the significance of differences between the diameter and cell volume for each sex. Spearman’s correlation (r) was used to assess the degree of association between the number of nests that were constructed in each month and climate data for each month. This correlation was also used to examine the relationship between the total length of the cavity of utilized nests and the number of cells that were constructed (brood cells and vestibular cells). The Chi-square (χ²) test was calculated to determine whether nest construction frequencies were similar in the Cerrado and gallery forest habitats, and whether the observed male / female sex ratio differed from the expected proportion of 1:1. Statistical analyses were performed using Systat, version 10.0 (Wilkinson, 2000) and PAST (Hammer, Harper & Ryan, 2001).
Results
Nesting activity and seasonality:
A total of 29 nests of C. dichrootricha were collected, and this number represented a construction rate of 4.83 % of the total available nests (Table 1). Nest construction occurred more frequently in the gallery forest (72.41 %) compared with the Cerrado habitat (27.59 %). The difference between the number of nests constructed in each habitat was significant (χ²= 5.56; p < 0.05; DF= 1).
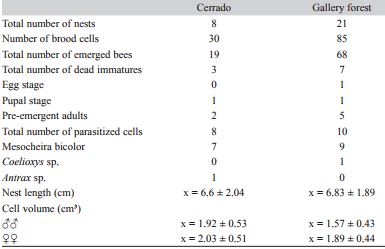
The species’ seasonal nesting activity began at the onset of the rainy season (most likely in October), and most nest building ended in November. This seasonal pattern was maintained in the second year of the study for both the gallery forest and the Cerrado, but the number of nests built was lower in the second year than in the previous year (Fig. 1). Significant correlations were not observed between the number of nests that were constructed and climate factors.
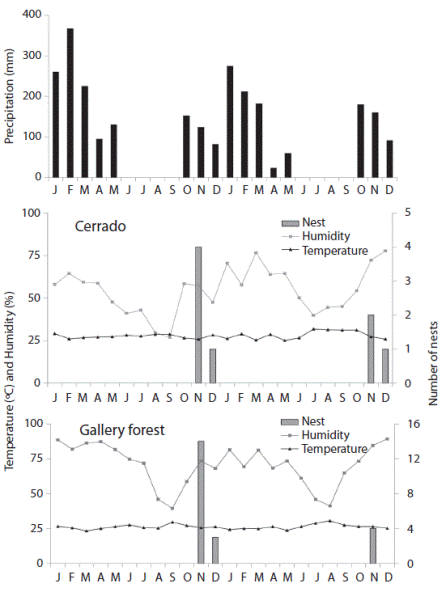
Fig. 1
Climatic factors (precipitation, temperature and humidity) and number of Centris (Hemisiella) dichrootricha nests collected in the cerrado and gallery forest habitats between January 2012 and December 2013 in Formosa da Serra Negra Municipality, Maranhão, Brazil.
Nest architecture:
The nest cells were primarily constructed of sand and an adhesive substance that bound the sand particles together. The soil in the nests differed in origin, and soils of two different colors were observed: red-yellow latosol that is characteristic of the Cerrado, and gray-colored soil from the gallery forest (Fig. 2A, Fig. 2B). For the nests that were constructed in the gallery forest, 80.9 % of the soil originated from the Cerrado and 19.0 % originated from the gallery forest. However, the Cerrado nests were made entirely from the red (Cerrado) soil.
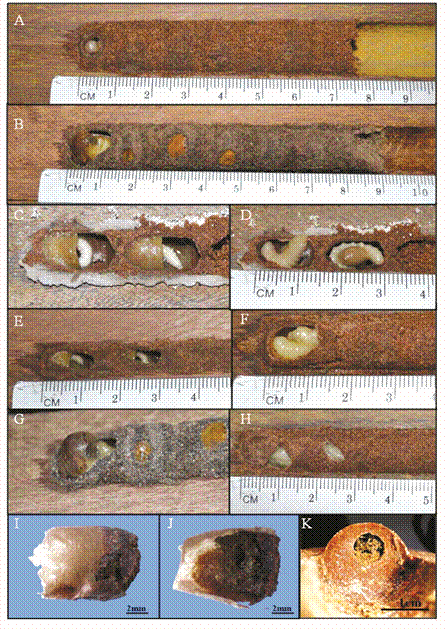
Fig. 2
Characteristics of nests and larval development of Centris (Hemisiella) dichrootricha: (A) red soil; (B) gray soil; (C) and (D) larvae of nest after removal from the area (11/09/2012) and after four days of development (11/13/2012), respectively; after this stage, one individual of Mesocheira bicolor emerged at 25 days (cell C2) and one female of C. dichrootricha emerged at 27 days (cell C1); (E) larvae feeding from nectar (individuals emerged after 35 days); (F) larva in cell C1 (one individual emerged after 27 days); (G) larva finishing the cocoon (individual emerged after 28 days); (H) cocoon of light yellow color (individuals emerged after 22 and 24 days); (I) and (J) cocoon; and (K) nest plug after individuals had emerged.
Females constructed their nests in 8 mm (27.6 %), 10 mm (31 %), 12 mm (27.6 %) and 14 mm (13.8 %) diameter trap-nests, but did not in 6 and 16 mm diameter traps. The intranest cavity length varied from 1.9 to 9.8 cm, and a nest contained one to seven cells. The length of a utilized nest was directly related to the number of built cells, and there was a strong correlation between nest space and cell number (r= 0.82; p < 0.001; n= 29). C. dichrootricha built 115 cells in nests from both habitats. Ten vestibular cells were observed, with four from the Cerrado (x= 1.01 ± 0.56 cm³) and six from the gallery forest (x=1.39 ± 0.44 cm³). Cells were compacted and placed linearly, and the internal surface of each cell consisted of a polished and smooth wall. The nest entrance plug had the same consistency as the cells and varied from 2.29 to 19.86 mm in thickness. The external side of the entrance ended in a concave shape with the same smooth and polished aspect as the internal wall of the brood cell, and did not show evidence of material deposition (Fig. 2A, Fig. 2B).
The multivariate analysis indicated that significant differences did not occur between the Cerrado and gallery forest habitats with regard to maximum head width and cell volume (F2, 74= 2.454; p= 0.093), but significant differences were observed between sexes with regard to maximum head width and cell volume (F2, 74= 298.76; p < 0.001). The mean female cell volume was higher than the male cell volume (Table 1), and significant differences were observed for the diameters 10, 12 and 14 mm (Manny-Whitney♂ x ♀, U10 mm= 46, p10 mm= 0.033; U12 mm= 13.5, p12 mm= 0.005; and U14 mm= 11, p14 mm= 0.031). Significant differences were not observed for the female cell volume for diameters of 8, 10, 12 and 14 mm (Kruskal-Wallis= 5.421; p♀= 0.143) or for the male cell volume for different diameters (Kruskal-Wallis= 7.726; p♂= 0.052).
Larval food consisted of a paste of pollen mixed with floral oil that occupied approximately half the volume of the cell, and the other half was filled with nectar (Fig. 2C, Fig. 2D, Fig. 2E). The larvae fed initially from the nectar portion and then from the pollen paste (Fig. 2C, Fig. 2D). When they were finished feeding, the larvae built cocoons for their transformation into the pupal stage. A translucent yellow membrane covered approximately two-thirds of the cocoon, which was overlaid with a fused rigid and strong crust of feces (Fig. 2I, Fig. 2J).
Adult emergence, parasitism and mortality:
A total of 68 C. dichrootricha specimens emerged from the gallery forest nests and 19 emerged from the Cerrado nests. C. dichrootricha had only one generation per year in both habitats, and these offspring had a short activity cycle that lasted from November to December. Bees emerged at an interval of 14 to 35 days after the nests had been removed from the field, and parasite species emerged at a shorter interval (9 to 32 days) than the host species. No diapauses were observed in any bee development phase.
Parasitism occurred in 14 nests (48.3 %), which contained cells that were parasitized by two bee species, Mesocheira bicolor (Fabricius, 1804) (Apinae) and Coelioxys sp. (Megachilinae), and one fly species, Antrax sp. (Diptera). M. bicolor was the most abundant parasite (16 specimens), and the other parasites species were represented by only one individual each. In addition, parasitized cells, 10 C. dichrootricha individuals died during the egg, larval or pre-adult phases (Table 1).
Adult size and sex ratio:
The females were significantly larger than the males (F1, 76= 595.19; p < 0.001), although significant differences were not observed between males and females in head width and trap-nest diameter (Kruskal-Wallis♀= 5.936, p♀ = 0.115; KruskalWallis♂= 0.319, p♂= 0.956).
Protandry is the emergence of males before females, and it was observed for C. dichrootricha in both Cerrado and gallery forest. Males were found in all the constructed cells (cells one to seven), females were found in the first four cells, and parasites were found from the second to the sixth cells (Fig. 3). The female-biased sex ratio was 1.17:1, which was not significantly different from the expected proportion of 1:1 (χ²= 0.04; p= 0.83; DF= 1).
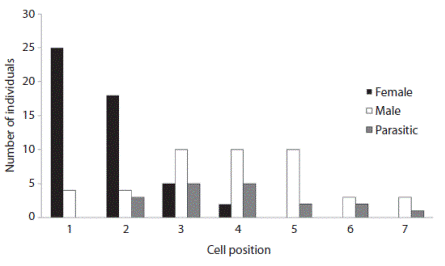
Fig. 3
Distribution of males, females and parasites with regard to the position of cells, demonstrating protandry in Centris (Hemisiella) dichrootricha.
Pollen analysis of the nests:
An analysis of the pollen in C. dichrootricha nests showed that there were 11 pollen types that belonged to 10 plant species from six families, with one undetermined pollen type (Table 2, Fig. 4). The average occurrence frequencies of the different pollen types revealed that Malpighiaceae (85.3 %) was the most represented throughout the sampling period, followed by Moraceae (6.75 %) and Myrtaceae (5.48 %). The most common sources of pollen and floral oil for C. dichrootricha in this study were three species of the genus Byrsonima, which represented 82.9 % of the samples, especially Byrsonima rotunda (79.2 %). The genus Byrsonima Rich. ex Kunth was predominant for the nests in November and December for the two habitats; however, the second-most frequent pollen type was Eugenia sp. (Myrtaceae) for the Cerrado, and Brosimum sp. (Moraceae) for the gallery forest in December (21.1 % and 26.8 %, respectively) (Table 2).
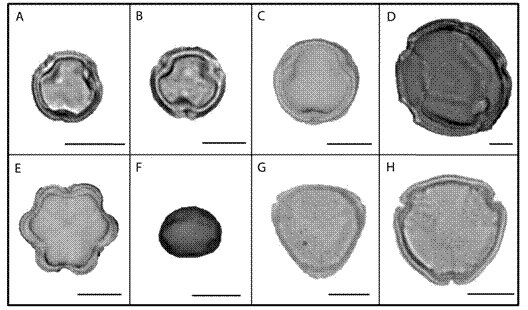
Fig. 4
The pollen types used most frequently by Centris (Hemisiella) dichrootricha in November and December 2012: (A) Byrsonima rotunda (Malpighiaceae); (B) Byrsonima crassifolia (Malpighiaceae); (C) Byrsonima spicata (Malpighiaceae); (D) Heteropterys sp; (Malpighiaceae); (E) Mouriri elliptica (Melastomataceae); (F) Brosimum sp. (Moraceae); (G) Eugenia sp. (Myrtaceae); and (H) Ouratea castaneifolia (Ochnaceae) (scale: 10 μm).
Discussion
C. dichrootricha is a univoltine species with a tendency to nest in forests in cavities of various diameters. Most of its food sources (pollen, nectar and floral oil) and soils used for nest building originated in the Cerrado. Quantitatively, few food sources were used, which characterizes the species as oligolectic, and qualitatively, C. dichrootricha showed a preference for species of the genus Byrsonima (82.9 %).
C. dichrootricha’s nest construction rate was similar to that found by Morato et al. (1999) in Amazon rainforest fragments. Compared with other species of the subgenus Hemisiella, the nest construction rate of C. dichrootricha was lower in the Cerrado and gallery forest, 2.67 % and 7 %, respectively, compared with that of C. tarsata, 5 % and 35.5 %, respectively (Aguiar & Garófalo, 2004; Mendes & Rêgo, 2007; Mesquita et al., 2009; Pires, Pompeu & Souza-Silva, 2012), C. trigonoides, 10 % (Aguiar et al., 2006) and C. vittata, 10.7 % (Mesquita et al., 2009). There is only one record of the species nesting in pre-existing cavities in Brazil (Morato et al., 1999). C. dichrootricha males were observed patrolling trap-nests in Costa Rica, although females were not observed nesting in this location (Thiele, 2005).
The low abundance of C. dichrootricha reported in the present study is consistent with the results of a faunal survey of flowers performed in the same study area (total effort of 624 h) in which only one C. dichrootricha individual was collected (data unpublished). The nesting behavior of C. dichrootricha has not been extensively studied, and prior to the present study, nesting had only been recorded in forest environments (Morato et al., 1999). The present study found that this species also nests in the Cerrado. Only a limited number of studies have been conducted for this species in forested areas, and priority should be given to studies documenting the biogeography of C. dichrootricha, especially those using trapnests and other bee sampling methodologies, because there are limited records of its occurrence in traditional faunal inventories and only a few reports of its occurrence in Brazil (see Silveira, Melo, & Almeida, 2002).
C. dichrootricha used cavities of 8 to 14 mm in diameter. This behavior expands the range of its nesting preferences because there are previous records of it nesting in cavities of 9.5 mm and 12.7 mm (Morato et al., 1999). The choice of a certain cavity size is related to the size of the species (Krombein, 1967). For example, C. tarsata is a medium-sized species and nests in cavities of 8 mm and 10 mm (Silva et al., 2001) and 7 mm to 13 mm in diameter (Buschini & Wolff, 2006). Another large species, C. vittata, prefers diameters that vary from 9 mm to 19 mm (Pereira, Garófalo, Camillo, & Serrano, 1999; Vinson et al., 2010).
A pattern was found in intranest cavity occupation that corroborates reports that bees usually do not use the entire cavity space (Pereira et al., 1999; Buschini & Wolff, 2006; Mesquita et al., 2009). This length variation is mainly related to the number of constructed cells, as demonstrated by Pereira et al. (1999) for C. vittata, in which there was a correlation between the number of constructed cells and the length of the cavity used. However, this correlation only applies for linearly placed cells because reduced lengths have been observed when the cells are placed in two parallel columns.
Bamboo cavities usually allow the bee to build a greater number of cells, especially when the species places its cells in two rows, as was observed for C. tarsata in the study of Aguiar and Garófalo (2004), who found that the number of constructed cells varied from 2 to 7 and 9 and 13. However, Mendes and Rêgo (2007) found 1 to 12 cells, and Mesquita et al. (2009) found a range of 1 to 10 cells. C. vittata requires larger cells and places its cells in only one row that has 1 to 9 cells (Pereira et al., 1999; Mesquita et al., 2009). For C. dichrootricha, the present study found that 1 to 7 cells were built, which corroborates the findings of Morato et al. (1999).
Although the present study did not analyze nest stratification, Morato et al. (1999) reported that C. dichrootricha nests were at heights of 8 m and 15 m (88.5 % of nests) and only found seven nests that were located at 1.5 m. The present study documented C. dichrootricha nesting at a height of 1.5 m. It has been suggested that due to the canopy size of the Alpercatas River gallery forest, if trapnests are placed at a greater height, the number of nests constructed by the species will most likely increase.
There was no correlation between the beginning of the rainy season and the beginning of species activity. This result is inconsistent with the observations of Aguiar and Garófalo (2004) for C. tarsata in a Caatinga area in Ipirá, Bahia, Brazil. In this study, the activity of C. dichrootricha most likely began in October because their peak activity occurred in surveys from November in both years and both forests (gallery and Cerrado). This result is similar to the seasonal activity that was observed in the Amazon rainforest by Morato et al. (1999), who found that nesting by C. dichrootricha in the Amazon rainforest occurred from September to October and was more abundant in the second year. This result is inconsistent with the results of the present study, in which nests were more abundant in the first year.
An obligatory characteristic of univoltine species is a long diapause period during certain development phases. However, diapauses were not observed for C. dichrootricha in the generations that were monitored in 2012 and 2013 in the present study. Even multivoltine species such as C. analis (Couto & Camillo, 2007) and C. tarsata (Aguiar & Garófalo, 2004) go into diapause during the immature phase after several consecutive generations. Authors who have observed the absence of diapause are skeptical that C. tarsata can go into diapause during the adult phase (Buschini & Wolff, 2006). However, newly emerged adults are believed to cease their activities under unfavorable conditions for their development, such as a lack of resources (Denlinger, 1986; Danks, 2002).
The two different soil colors used by C. dichrootricha in the present study may also be a pattern that is adopted by other species. Vinson et al. (2010) observed differences in the color and size of sand grains used by several Centris species. According to Vinson et al. (2010), Centris (Hemisiella) nitida Smith, 1874 prefers dark soil, whereas C. trigonoides does not, and C. vittata uses larger soil particles compared with those used by C. nitida and C. trigonoides. It is clear that local soil variations affect the type of soil used in nests. However, in this study, most of the soil used by C. dichrootricha to construct nests in the gallery forest originated from the Cerrado. Vinson et al. (2010) assumed that the light color of the soil used by C. trigonoides resulted from it nesting in arid areas, which corroborates the results of the present study, in which all C. dichrootricha nests constructed in the Cerrado were made of soil that originated from the Cerrado.
The closing walls of C. dichrootricha nests consisted of a molded coating of a consistency similar to that of the cell walls and were similar to those of C. trigonoides nests (Aguiar et al., 2006). Most of the materials used to build the nests (sand and floral oil) and for larval food originated from the Cerrado (90 %), despite C. dichrootricha constructing 72.4 % of its nests in gallery forest. There are no reports in the literature on the contributions of different ecosystems to the resources that are used by solitary bee populations, although information is available on nesting site preferences (Morato et al., 1999; Aguiar, Garófalo, & Almeida, 2005; Buschini, 2006; Vinson et al., 2010). Thus, future studies on other bee species are required to examine the relationships among the period of resource availability, species’ seasonal activity and preferences for certain food sources. The present study highlights the importance of such information when investigating the biology and behavior of C. dichrootricha. This is emphasized in the present study because of its implications for establishing priority areas for the conservation of the species, as C. dichrootricha depends on both Cerrado vegetation types to maintain their populations. Gallery forest most likely provides microclimatic conditions that are more favorable for the development of C. dichrootricha broods, perhaps because of the low exposure to light. According to Martins, Ferreira and Carneiro (2012), bees nest more frequently in shaded areas, although the biology of the species influences this preference. For example, C. tarsata prefers to nest in open areas (Viana et al., 2001; Buschini & Wolff, 2006; Mendes & Rêgo, 2007), although only one study has been conducted in Brazil in forest environments for other Centris species (Morato et al., 1999).
The present study is the first to describe the sources of larval food composition for C. dichrootricha. Nectar and a mixture of pollen and floral oil were found to be the components of C. dichrootricha larval food. Based on the type of larval food of solitary bees, Aguiar and Garófalo (2004) subdivided the species of the genus Centris according to whether they use floral oil (see the species list in Aguiar & Garófalo, 2004). Aguiar et al. (2006) later reorganized the classification for C. trigonoides. With the results of the present study, C. dichrootricha is included in this classification of floral oil users, which also includes C. vittata, C. tarsata and C. trigonoides. Thus, all species of the group Hemisiella that nest in trap-nests share the same behavior with regard to larval food (Pereira et al., 1999; Aguiar & Garófalo, 2004; Aguiar et al., 2006).
The sizes of males and females do not overlap in other species of the subgenus Hemisiella (Pereira et al., 1999; Silva et al., 2001; Mendes & Rêgo, 2007). Sexual dimorphism is reflected in the size of the brood cells, with the volume of female cells being greater than the volume of male cells. In the present study, females were placed in the first cells, and males occupied all positions, which confirm the observations of Silva et al. (2001), Buschini and Wolff (2006) and Mendes and Rêgo (2007), for C. tarsata and of Drummont, Silva and Viana (2008) for Centris (Heterocentris) terminata Smith, 1874. In addition, males are smaller than females in all Centris species (Pereira et al., 1999; Jesus & Garófalo, 2000; Silva et al., 2001; Buschini & Wolff, 2006; Mendes & Rêgo, 2007).
Mesocheira bicolor has been identified as a parasite in nests of six species of the genus Centris: C. (Hemisiella) dichrootricha, C. (Hemisiella) nitida, C. (Hemisiella) tarsata, C. (Hemisiella) trigonoides, C. (Heterocentris) analis and C. (Heterocentris) terminata (RochaFilho, Morato & Melo, 2009). M. bicolor was the most numerous parasite of C. dichrootricha (87.5 %) in the present study. This differs from the results of the study by Morato et al. (1999), in which M. bicolor represented only 12.5 % of parasite specimens and C. dichrootricha was primarily parasitized by one coleopteran species (Tetraonyx sp., 78.1 %). The parasite Coelioxys sp. is common for Centris species and has been observed for C. vittata, C. tarsata and C. trigonoides (Morato et al., 1999; Pereira et al., 1999; Aguiar & Garófalo, 2004; Buschini & Wolff, 2006; Mendes & Rêgo, 2007; Ramos et al., 2010).
The activity of C. dichrootricha occurred during the flowering peak of B. rotunda, which is commonly known as “murici-branco” (white murici), and three other species of Malpighiaceae were in bloom during this period. Several authors have reported that the genus Centris has an affinity for Malpighiaceae species (Neff & Simpson, 1981; Buchmann, 1987; Albuquerque & Rêgo, 1989; Vogel, 1990; Vinson et al., 1997). Oil-collecting bees present morphological adaptations in their legs to collect oil (Vogel, 1974), and these adaptations promote a mutualistic evolutionary relationship between species of Malpighiaceae and Centridini in the Neotropical region (Rêgo & Albuquerque, 1989; Sigrist & Sazima, 2004; Davis, Schaefer, Baum, Donoghue, & Harmon, 2014).
Oil-collecting bees are strictly associated with oil flowers of the genera Byrsonima and Banisteriopsis (Vogel, 1974; Buchmann, 1987; Mello et al., 2013). Ramos et al. (2010) recorded C. vittata females visiting Byrsonima flowers and found pollen from Tetrapterys sp. (Malpighiaceae) in the scopae of a dead female inside a nest, and Mendes and Rêgo (2007) found pollen of Banisteriopsis sp. (Malpighiaceae) and Cassia sp. (Fabaceae) in nests of C. tarsata in a eucalyptus plantation and a mesophytic forest area. Chamaecrista ramosa (Vogel) H.S. Irwin & Barneby (Fabaceae) and Byrsonima vacciniifolia A. Juss. were reported to be the main sources of pollen for C. analis (Fabricius, 1804) (Dórea, Aguiar, Figueroa, Lima, & Santos, 2010). In addition, Centris (Ptilotopus) maranhensis and Centris (Centris) decolorata Lepeletier, 1841 (cited as Centris leprieuri) have been reported to exclusively use Byrsonima pollen as a nest food source (Ramos et al., 2007b; Ribeiro, Rêgo, & Machado, 2008).
Although there is evidence for the preference of Centridini for plant families that produce floral oil, other plant families were used by C. dichrootricha as pollen sources (Myrtaceae and Moraceae). Some studies, e.g., Dórea, Santos, Lima and Figueroa (2009) for C. tarsata, showed that other plant families, such as Leguminosae (49.3 %) and Solanaceae (43.2 %), were used as a main food source of pollen. These authors observed the importance of Solanum paniculatum L. and Senna rizzini H. S. Irwin & Barneby as pollen sources in a Caatinga area in Bahia despite the availability of oil plants. Gonçalves, Silva and Buschini (2012) reported results from Paraná State that are similar to those from the caatinga for C. tarsata, and they reported that two species of Solanaceae were the main sources of pollen (Solanum americanum Mill. and Solanum variabile Mart.). Chamaecrista ramosa and S. paniculatum were preferred by C. trigonoides in Bahia (Dórea et al., 2013).
Additional records for C. dichrootricha, different than those in the review by Rebêlo, Rêgo and Albuquerque (2003), on the diversity of bees in the state of Maranhão are not available. The present study provides a record of the presence of the species in the state and the first record of the bee in the Northern Cerrado of Brazil. Another unique contribution of the present study is the observation that two types of soil are used in the construction of nests, with soil from the Cerrado being used most frequently, especially in gallery forest nests. Although there was a significant difference between the number of nests that were constructed in the two studied habitats, the species requires products of both habitats, such as a suitable nest site, soil to build the nests and pollen and energy material for the brood. Thus, the gallery forest and the Cerrado are believed to be intrinsically related to the maintenance of local populations of C. dichrootricha. Additional studies are required to clarify the distribution of populations in other areas that have gallery forest vegetation and areas that are influenced by the Amazon rainforest.
C. dichrootricha is a species that produces only one generation per year and has floral preferences restricted to plants that bloom from October to December. These sources are abundant in the Cerrado area of Mirador State Park. The nesting sites of C. dichrootricha are at the margins of the Alpercatas River. The quality of the Cerrado and gallery forest ecosystems appears to be essential for the maintenance of C. dichrootricha populations. We propose that C. dichrootricha occurs only in areas of the Cerrado that are close to gallery forest. This hypothesis can be tested in future studies. In addition, the impact of ecosystem fragmentation on C. dichrootricha populations should be investigated for the conservation of species in other areas of the Northern Neotropical region.
Acknowledgments
The authors appreciate the valuable suggestions for this paper from Carlos Alberto Garófalo and José Manuel Macário Rebêlo. We thank Fernando Silveira for identifying the species, Catalina Angel for translating the abstract into Spanish and Felipe Rêgo for his motivational support. We would like to thank the anonymous reviewers for their valuable comments and suggestions. We also thank the field team at Mirador State Park: Ana Carolina Araújo, Denilson Martins, Fernanda Brito and Samara Serra. MCT/CNPq/FNDCT/CT-AçãoTransversal/CT-Amazônia/CT-Biotec-BIONORTE (Process 554318/2010-5) and FAPEMA (CBIOMA-02930/12) provided the financial support for the research project. CAPES and CNPq provided grants to G.C.A.C. and P.M.C.A., respectively. SEMA-MA authorized the collection of bees and plants in Mirador State Park
References
Aguiar, C. M. L., & Gaglianone, M. C. (2003). Nesting biology of Centris (Centris) aenea Lepeletier (Hymenoptera, Apidae, Centridini). Revista Brasileira de Zoologia, 20(4), 601-606.
Aguiar, C. M. L., & Garófalo, C. A. (2004). Nesting biology of Centris (Hemisiella) tarsata Smith (Hymenoptera, Apidae, Centridini). Revista Brasileira de Zoologia , 21(3), 477-486.
Aguiar, C. M. L., Garófalo, C. A., & Almeida, G. F. (2005). Trap-nesting bees (Hymenoptera, Apoidea) in areas of dry semideciduous forest and caatinga, Bahia, Brazil. Revista Brasileira de Zoologia , 22(4), 1030-1038.
Aguiar, C. M. L., Garófalo, C. A., & Almeida, G. F. (2006). Biologia de nidificação de Centris (Hemisiella) trigonoides Lepeletier nidificação (Hymenoptera, Apidae, Centridini). Revista Brasileira de Zoologia , 23(2), 323-330.
Albuquerque, P. M. C., & Rêgo, M. M. C. (1989). Fenologia de abelhas visitantes de murici (Byrsonima crassifolia, Malpighiaceae). Boletim do Museu Paraense Emílio Goeldi, 5, 163-178.
Buchmann, S. (1987). The ecology of oil flowers and their bees. Annual Review of Ecology, 18, 343-369.
Buschini, M. L. T. (2006). Species diversity and community structure in trap-nesting bees in Southern Brazil. Apidologie, 37(1), 58-66.
Buschini, M. L. T., & Wolff, L. L. (2006). Nesting biology of Centris (Hemisiella) tarsata Smith in Southern Brazil (Hymenoptera, Apidae, Centridini). Brazilian Journal of Biology, 66(4), 1091-1101.
Camillo, E., Garófalo, C. A., Serrano, J. C., & Muccillo, G. (1995). Diversidade e abundância sazonal de abelhas e vespas solitárias em ninhos armadilhas (Hymenoptera, Apocrita, Aculeata). Revista Brasileira de Entomologia, 39(2), 459-470.
Couto, M. R., & Camillo, E. (2007). Influência da temperatura na mortalidade de imaturos de Centris (Heterocentris) analis (Hymenoptera, Apidae, Centridini). Iheringia, Série Zoológica, 97(1), 51-55.
Coville, R., Frankie, G. W., & Vinson, S. B. (1983). Nests of Centris segregata (Hymenoptera: Anthophoridae) with a Review of the Nesting Habitats of the Genus. Journal of the Kansas Entomology Society, 56(2), 109-122.
Danks, H. V. (2002). The ranges of insects dormancy responses. European Journal of Entomology, 99, 127-142.
Davis, C. C., Schaefer, H., Xi, Z., Baum, D. A., Donoghue, M. J., & Harmon, L. J. (2014). Long-term morphological stasis maintained by a plant-pollinator mutualism. Proceedings of the National Academy of Sciences of the United States of America, 111(16), 5914-5919.
Denlinger, D. L. (1986). Dormancy in tropical insects. Annual Review of Entomology, 31, 239-264.
Dórea, M. C., Santos, F. A. R., Lima, L. C. L., & Figueroa, L. E. R . (2009). Análise Polínica do Resíduo Pós-Emergência de Ninhos de Centris tarsata Smith (Hymenoptera: Apidae, Centridini). Neotropical Entomology, 38(2), 197-202.
Dórea, M. C., Aguiar, C. M. L., Figueroa, L. E. R., Lima, L. C. L., & Santos, F. A. R. (2010). Residual pollen in nests of Centris analis (Hymenopera, Apidae, Centridini) in an area of Caatinga vegetation from Brazil. Oecologia Australis, 14(1), 232-237.
Dórea, M. C., Aguiar, C. M. L., Figueroa, L. E. R., Lima, L. C. L., & Santos, F. A. R. (2013). A study of pollen residues in nests of Centris trigonoides Lepeletier (Hymenoptera, Apidae, Centridini) in the Caatinga vegetation, Bahia, Brazil. Grana, 52(2), 37-41.
Drummont, P., Silva, F. O., & Viana, B. F. (2008). Ninhos de Centris (Heterocentris) terminata Smith (Hymenoptera, Apidae, Centridini) em Fragmentos de Mata Atlântica Secundária, Salvador, BA. Neotropical Entomology , 37(3), 239-246.
Erdtman, G. (1960). The acetolysis method. A revised description. Svensk Botanisk Tidskrift, 54, 561-564.
Gaglianone, M. C. (2001). Nidificação e forrageamento de Centris (Ptilototopus) scopipess Friese (Hymenoptera, Apidae). Revista Brasileira de Zoologia , 18(1), 107-117.
Garófalo, C. A., Martins, C. F., & Alves-dos-Santos, I. (2004). The brazilian solitary bee species caught in trap nests. In B. M. Freitas, & J. O. P. Pereira (Eds.), Solitary Bees Conservation: Rearing and Management for Pollination (pp. 26-30). Fortaleza: Editora Universitária.
Gonçalves, L., Silva, C. I., & Buschini, M. L. T. (2012). Collection of Pollen Grains by Centris (Hemisiella) tarsata Smith (Apidae: Centridini): Is C. tarsata an Oligolectic or Polylectic Species? Zoological Studies, 51(2), 195-203.
Hammer, Ø., Harper, D. A. T., & Ryan, P. D. (2001). Paleontological Statistics software package for education and data analysis. Palaeontologia Electronica.
IBGE. (1998). Subsídios ao zoneamento ecológico-econômico da Bacia do Rio Itapecuru - MA: diretrizes gerais para ordenação territorial/ IBGE. Primeira Divisão de Geociências do Nordeste, Rio de Janeiro.
Jesus, B. M. V., & Garófalo, C. A. (2000). Nesting behaviour of Centris (Heterocentris) analis (Frabicius) in southeastern Brazil (Hymenoptera, Apidae, Centridini). Apidologie , 31, 503-515.
Krombein, K. V. (1967). Trap-nesting wasps and bees: Life histories, nests and associates. Washington, D.C.: Smithsonian.
Machado, I. C. (2004). Oil-collecting bees and related plants: a review of the studies in the last twenty years and case histories of plants occurring in NE Brazil. In B. M Freitas, & J. O. P. Pereira (Eds.), Solitary Bees Conservation: Rearing and Management for Pollination (pp. 255-280). Fortaleza: Editora Universitária .
Martins, C. F., Ferreira, R. P., & Carneiro, L. T. (2012). Influence of the orientation of nest entrance, shading, and substrate on sampling trap-nesting bees and wasps. Neotropical Entomology , 41(2), 105-111.
Martins, A. C., Aguiar, A. J. C., & Alves-dos Santos, I. (2013). Interaction between oil-collecting bees and seven species of Plantaginaceae. Flora-Morphology, Distribution, Functional Ecology of Plants, 208(7), 401-411.
Mello, M. A. R., Bezerra, E. L. S., & Machado, I. C. S. (2013). Functional Roles of Centridini Oil Bees and Malpighiaceae Oil Flowers in Biome-wide Pollination Networks. Biotropica, 45(1), 45-53.
Mendes, F. N., & Rêgo, M. M. C. (2007). Nidificação de Centris (Hemisiella) tarsata Smith (Hymenoptera, Apidae, Centridini) em ninhos-armadilha no Nordeste do Maranhão, Brasil. Revista Brasileira de Entomologia , 51(3), 382-388.
Mesquita, T. M., Vilhena, A. M. G. F., & Augusto, C. S. (2009). Ocupação de ninhos-armadilha por Centris (Hemisiella) tarsata Smith, 1874 e Centris (Hemisiella) vittata Lepeletier, 1841 (Hymenoptera: Apidae: Centridini) em áreas de Cerrado. Bioscience Journal, 25(5), 124-132.
Mesquita, T. M. S., & Augusto, S. C. (2011). Diversity of trap-nesting bees and their natural enemies in the Brazilian savanna. Tropical Zoology, 24, 127-144.
Michener, C. D. (2007). The Bees of the Word. Baltimore: The Johns Hopkins University Press.
Morato, E. F., Garcia, M. V. B., & Campos, L. A. D. O. (1999). Biologia de Centris Fabricius (Hymenoptera, Anthoporidae, Centridini) em matas contínuas e fragmentos na Amazônia Central. Revista Brasileira de Zoologia , 16(4), 1213-1222.
Murúa, M., Cisterna, J., & Rosende, B. (2014). Pollination ecology and breeding system of two Calceolaria species in Chile. Revista Chilena de Historia Natural, 87(7), 1-3.
Neff, J., & Simpson, B. (1981). Oil-Collecting Structures in the Anthophoridae (Hymenoptera), Morphology, Function, and Use in Systematics. Journal of the Kansas Entomological Society, 54(1), 95-123.
Pemberton, R. W., & Liu, H. (2008). Naturalization of the Oil Collecting Bee Centris nitida (Hymenoptera, Apidae, Centrini), a Potential Pollinator of Selected Native, Ornamental, and Invasive Plants in Florida. Florida Entomologist, 91, 101-109.
Pereira, M., Garófalo, C. A., Camillo, E., & Serrano, J. C. (1999). Nesting biology of Centris (Hemisiella) vittata Lepeletier in southeastern Brazil (Hymenoptera, Apidae, Centridini). Apidologie , 30, 327-338.
Pires, E. P., Pompeu, D. C., & Souza-Silva, M. (2012). Nidificação de vespas e abelhas solitárias (Hymenoptera: Aculeata) na Reserva Biológica Boqueirão, Ingaí, Minas Gerais. Bioscience Journal , 28(2), 302-311.
Ramos, M. C., Rêgo, M. M. C., & Albuquerque, P. M. C. (2007a). Ocorrência de Centris (Hemisiella) vittata Lepeletier (Hymenoptera: Apidae: Centridini) no Cerrado s. l. do Nordeste do Maranhão, Brasil. Acta Amazonica, 37(1), 165-168.
Ramos, M., Mendes, F., Albuquerque, P., & Rêgo, M. (2007b). Nidificação e forrageamento de Centris (Ptilotopus) maranhensis Ducke (Hymenoptera, Apidae, Centridini). Revista Brasileira de Zoologia , 24(4), 1006-1010.
Ramos, M., Albuquerque, P., & Rêgo, M. (2010). Nesting Behavior of Centris (Hemisiella) vittata Lepeletier (Hymenoptera: Apidae) in an Area of the Cerrado in the Northeast of the State of Maranhão, Brazil. Neotropical Entomology , 39(3), 379-383.
Rebêlo, J. M. M., Rêgo, M. M. C., & Albuquerque, P. M. C. (2003). Abelhas (Hymenoptera, Apoidea) da região setentrional do Estado do Maranhão, Brasil. In G. A. R. Melo, & I. Alves-dos-Santos (Eds.), Apoidea Neotropica: Homenagem aos 90 Anos de Jesus Santiago Moure (pp. 265-278). Criciúma: UNESC.
Rêgo, M. M. C., & Albuquerque, P. M. C. (1989). Comportamento das abelhas visitantes de murici, Byrsonima crassifolia (L.) Kunth, Malpighiaceae. Boletim Museu Paraense Emilio Goeldi, 5, 179-193.
Rêgo, M. M. C., Albuquerque, P. M. C., Ramos, M. C., & Carreira, L. M. (2006). Aspectos da Biologia de Nidificação de Centris flavifrons (Friese) (Hymenoptera: Apidae, Centridini), um dos Principais Polinizadores do Murici (Byrsonima crassifolia L. Kunth, Malpighiaceae) no Maranhão. Neotropical Entomology , 35(5), 579-587.
Renner, S. S., & Schaefer, H. (2010). The evolution and loss of oil-offering flowers: new insights from dated phylogenies for angiosperms and bees. Philosophical Transactions of the Royal Society of London. Series B, 365, 423-35.
Ribeiro, E. K. M. D., Rêgo, M. M. C., & Machado, I. C. S. (2008). Cargas polínicas de abelhas polinizadoras de Byrsonima chrysophylla Kunth. (Malpighiaceae): fidelidade e fontes alternativas de recursos florais. Acta Botanica Brasilica, 22(1), 165-171.
Rocha-Filho, L. C., Morato, E. F., & Melo, G. A. R. (2009). New host records of Aglaomelissa duckei and a compilation of host associations of Ericrocidini bees (Hymenoptera: Apidae). Zoologia, 26(2), 299-304.
Sigrist, M. R., & Sazima, M. (2004). Pollination and reproductive biology of twelve species of neotropical Malpighiaceae: stigma morphology and its implications for the breeding system. Annals of Botany, 94(1), 33-41.
Silva, F. O., Viana, B. F., & Neves, E. L. (2001). Biologia e Arquitetura de Ninhos de Centris (Hemisiella) tarsata Smith (Hymenoptera: Apidae: Centridini). Neotropical Entomology , 30(4), 541-545.
Silveira, F. A., Melo, G. A. R., & Almeida, E. A. B. (2002). Abelhas Brasileiras: Sistemática e Identificação. Belo Horizonte, Brasil: Editora Composição e Arte.
Simpson, B. B., Neff, J. L., & Seigler, D. (1977). Krameria, free fatty acids and oil-collecting bees. Nature, 267(56), 150-151.
Thiele, R. (2005). Phenology and nest site preferences of wood-nesting bees in a Neotropical lowland rain forest. Studies on Neotropical Fauna and Environment, 40(1), 39-48.
Wilkinson, L. (2000). SYSTAT: The system for statistics. SYSTAT: Evanston, Illinois.
Viana, B. F., Silva, F. O., & Kleinert, A. M. P. (2001). Diversidade e Sazonalidade de Abelhas Solitárias (Hymenoptera: Apoidea) em Dunas Litorâneas no Nordeste do Brasil. Neotropical Entomology , 30(2), 245-251.
Vinson, S. B., Frankie, G. W., & Williams, H. J. (1996). Chemical ecology of bees of the genus Centris (Hymenoptera: Apidae). Florida Entomologist , 79, 109-129.
Vinson, S., Williams, H., Frankie, G., & Shrum, G. (1997). Floral lipid chemistry of Byrsonima crassifolia (Malpigheaceae) and a use of floral lipids by Centris bees (Hymenoptera: Apidae). Biotropica , 29, 76-83.
Vinson, S. B., & Frankie, G. W. (2000). Nest Selection, Usurpation, and a Function for the Nest Entrance Plug of Centris bicornuta (Hymenoptera: Apidae). Entomological Society of America, 93(2), 254-260.
Vinson, S. B., Frankie, G., & Cônsoli, R. (2010). Description, Comparison and Identification of Nests of Cavity-Nesting Centris Bees (Hymenoptera: Apidae: Centridini) in Guanacaste Province, Costa Rica. Journal of the Kansas Entomological Society , 83(1), 25-46.
Vogel, S. (1974). Ölblumen und ölsammelnde Bienen. Tropische und Subtropische Pflanzenwelt, 7, 285-547.
Vogel, S. (1990). History of the Malpighiaceae in the light of pollination ecology. Memoirs of the New York Botanical Garden, 55, 130-142.
Zanella, F. C. V. (2002). Sistemática, filogenia e distribuição geográfica das espécies sul-americanas de Centris (Paracentris) Cameron, 1903 e de Centris (Penthemisia) Moure, 1950, incluindo uma análise filogenética do “grupo Centris” sensu Ayala, 1998 (Hymenoptera, Apoidea, Centridini). Revista Brasileira de Entomologia , 46(4), 435-488.