Resumen: O. niloticus se ha introducido en todo el mundo en reservorios de agua dulce y actualmente representa un alimento alternativo para las poblaciones. Dos marcas anuales en las escamas de tilapias se producen en estos ambientes, una de ellas se da por la época de reproducción mientras que la segunda se sugiere que se forma debido a bajas temperaturas, reducción en la oxigenación del agua y altas densidades entre otros estresores. Con el fin de explorar las causas de esta segunda marca se llevó a cabo un estudio experimental que analiza la influencia de cuatro factores en el crecimiento y en consecuencia en la producción de marcas del crecimiento en escamas: reducción de la cantidad de alimento, aumento en la densidad, disminución de la concentración de oxígeno y reducción de la temperatura. El experimento se realizó usando 5 tinas plásticas con 215 L de capacidad, cada una se subdividió en dos secciones para obtener una réplica por tratamiento. Se usaron 20 juveniles por réplica. Las condiciones control fueron las óptimas para la especie: 5 % del peso corporal como alimento, densidad de 3 peces /L, saturación de oxígeno de 4.98 ± 0.5 mg/L y temperatura de 25.0 ± 1.0 oC, mientras que los tratamiento variaron como sigue: Alimento (2.5 % del peso corporal como alimento); densidad (6 peces/L); oxígeno (2.8 ± 1.2 mg/L) y temperatura (19.6 ± 1.0 oC), donde la respuesta se estimó a través de las variaciones en longitud, peso y distancias inter-circulli. Cada tercer día se midió la concentración de fosfatos, amonio y nitratos con un fotómetro Hanna® HI8203. El experimento se dividió en tres etapas de mediciones de talla, peso y toma de escamas. 1) Etapa inicial: Antes de someter los organismos a experimentación. 2) Etapa final: al final del experimento y 3) Etapa compensatoria: donde todos las condiciones experimentales fueron llevadas a las condiciones del control. Todos los tratamientos causaron reducción en longitud, peso y distancias inter-circuli con respecto al control (6.7 ± 0.5 cm; 5.2 ± 1.2 g; 5.4 ± 0.5 mm), pero solo la temperatura y alimento fueron significativos (5.6 ± 0.4 y 5.8 ± 0.5 cm; 3.4 ± 0.7 y 3.3 ± 0.9 g; 3.4 ± 0.3 y 4.1 ± 0.6 mm para los tratamientos de temperatura y alimento, respectivamente) (P < 0.05). La experimentación tuvo una duración de tres semanas, y posteriormente los tratamientos fueron revertidos a condiciones control durante otras tres semanas donde se observó crecimiento compensatorio en los tratamientos de oxígeno y la densidad pero no de la misma manera para los estresores: temperatura y alimento. Las distancias inter-circuli fueron un buen indicador de períodos de estrés.
Palabras clave: Oreochromis niloticusOreochromis niloticus,marcas en escamasmarcas en escamas,crecimientocrecimiento,distancias inter-circulidistancias inter-circuli,juvenilesjuveniles,estudio experimentalestudio experimental.
Abstract: Oreochromis niloticus has been introduced worldwide into freshwater reservoirs and represents an alternative food for local populations. Two annual marks on fish scales have been reported for fish in these environments, one of them is allocated to reproduction, while the other is thought to be formed following low temperatures, a reduction in oxygen concentration and/or high densities, among others. In order to explore the causes of this second ring, we designed an experimental study to analyze the influence of four factors: a reduction in food amount, an increase in density, a decrease in oxygen concentration and temperature. The experiment was carried out using five 215 L plastic tanks, each one divided into two sections, so as to have treatment replicates (each with 20 juveniles). The control conditions were optimal for the species: 5 % of body weight in food, a density of 3 fish/L, a total oxygen saturation of 4.98 ± 0.5 mg/L, and a temperature of 25.0 ± 1.0 oC; the treatments varied as follows: food of 2.5 % of body weight, density of 6 fish/L, oxygen of 2.8 ± 1.2 mg/L and a temperature of 19.6 ± 1.0 oC. Fish responses to stressors were estimated through variations in length, weight and inter-circuli spaces or circuli spacing in fish scales. Every third day it was measured the concentrations of ammonium, phosphates and nitrates with a photometer. Three stages for length/weight fish measurements and scales sampling were considered: 1) Initial stage: before experiment started, 2) Final stage: at the end of the experiment, and 3) Compensatory stage: when all the experimental treatments were adjusted to the Control conditions; the experiments lasted for three weeks. The results indicated that all the treatments caused a reduction in length, weight and inter-circuli spaces when compared to the controls (6.7 ± 0.5 cm; 5.2 ± 1.2 g; 5.4 ± 0.5 mm, respectively), but only the variables of temperature and food resulted significant (5.6 ± 0.4 and 5.8 ± 0.5 cm; 3.4 ± 0.7 and 3.3 ± 0.9 g; 3.4 ± 0.3 and 4.1 ± 0.6 mm for temperature and food treatments, respectively) (P < 0.05). When treatments were reversed to the control conditions for another three weeks compensatory growth was observed in the oxygen and density treatments, but not in the same way for the temperature and food treatments. We concluded that inter-circuli distances are a good indicator to stress periods in this species.
Key words: Oreochromis niloticus, scales marks, growth, circuli spacing, juveniles, experimental study.
Artículo
Influencia de la temperatura, densidad, oxígeno y alimento en la formación de marcas en las escamas de tilapia
The influence of temperature, density, oxygen and food in the mark formation on the scales of tilapia
Universidad de Costa Rica
Recepción: 09 Agosto 2016
Aprobación: 24 Enero 2017
Las tilapias constituyen un grupo de varias especies de la Familia Cichlidae introducidas en países en desarrollo de regiones tropicales y subtropicales con el propósito de obtener una fuente de proteína de bajo costo de producción. Estas especies muestran una tasa de crecimiento acelerada y excelente conversión de alimento. Son resistentes a condiciones adversas del medio como variaciones de temperatura, sobreviven en aguas duras, soportan condiciones de anoxia, sobrevivien en ambientes con exceso de materia orgánica en suspensión y soportan periodos largos de ayuno (Morales, 1991; Boyd, 2004). De acuerdo con Isaac (1990) presentan un crecimiento muy variable causado por varios factores intrínsecos y extrínsecos tales como la temperatura del agua, disponibilidad de alimento, actividad metabólica y reproductiva que producen diferencias en el metabolismo.
Algunas estructuras de los peces, como las escamas, otolitos y radios espinosos permiten identificar cambios, que se han correlacionado con la depositación de material calcáreo con los cambios en el metabolismo del individuo y con el tiempo en que estos cambios ocurren, proporcionando así información del crecimiento individual (Ehrhardt, 1981), de aquí que la edad puede estimarse en especies ícticas a partir de las discontinuidades que ocurren en estas estructuras esqueléticas. Dentro de las causas que modifican la depositación de material calcáreo en tilapias, se ha mencionado factores ambientales como variaciones de la temperatura, modificación del fotoperiodo, baja disponibilidad de alimento, entre otros (Martínez, Cristina, & Ross, 1996; Morales-Nin, 2000).
En la literatura, hay consenso sobre la formación de anillos de crecimiento anuales en escamas y huesos operculares en varias especies de tilapia, debido a desoves y a las exigencias de energía durante el proceso de reproducción (Garrod, 1959; Fagade, 1974; Payne & Collinson, 1983; Jiménez-Badillo, 2006). En climas tropicales y subtropicales, la formación de anillos de Oreochromis andersonii (Castelnau, 1861) y O. mossambicus (Peters, 1852) se ha atribuido a la disminución en la actividad metabólica somática, resultado de las actividades reproductivas como el cortejo y la construcción de nidos (Booth, Merron, & Buxton, 1995; Bruton & Allonson, 1974). La maduración de las hembras de O. mossambicus y el cuidado parental de incubación bucal, es otro de los factores reportados por Tachihara, Tokunaga y Chimura (2002) en el río Genka en Okinawa Japón, la incubación bucal depriva a las hembras de alimento con la consecuente formación de marcas en las escamas (GómezMárquez, Peña-Mendoza, Salgado-Ugarte, & Arredondo-Figueroa, 2008). El mismo comportamiento fue reportado para Tilapia esculenta (Graham, 1928) en el Lago Victoria por Garood (1959).
En México, Gómez-Márquez (1998) reportó la formación de un anillo en escamas de tilapia para los meses de diciembre y enero en un lago tropical (Coatetelco, Morelos, México) y lo atribuyó a la disminución de la temperatura del agua a 21 °C durante la estación invernal. Esto concuerda con reportes de temperatura óptima para el crecimiento y reproducción de O. niloticus (Linnaeus, 1758) de entre 22 y 32 °C (Beamish, 1970). Así mismo GómezMárquez (1998) adjudica la segunda marca, cerca del borde de la escama, a la interrupción del crecimiento por la maduración de las gónadas y la actividad de desove a pesar de que esta especie desova todo el año.
Beltrán-Alvarez, Sánchez-Palacios, Valdez y Ortega-Salas (2010) reportaron marcas en los meses de junio a agosto, y lo relacionaron con altas temperaturas del agua y la presencia de intensa estratificación térmica, que restringe la solubilidad del oxígeno disuelto en los primeros seis metros de la columna de agua. Ellos también adjudican la marca a la competencia por espacio y alimento originada por la disminución del volumen de agua en el embalse debido al estiaje. En este sentido Kebus et al. (1992) reportan para la trucha arcoiris que el incremento de la densidad de siembra lleva a un “stress social” causando un “stress crónico”, lo cual puede resultar en efectos negativos en el crecimiento. En tilapias también se ha observado la reducción del crecimiento en embalses que reducen su volumen, lo que se adjudica a competencia por espacio y alimento entre los organismos (Beltrán-Álvarez, Sánchez-Palacios, Ramírez-Lozano, & Santiago-Amaya, 2014). Así mismo, otra causa de estrés agudo y estrés crónico en peces es la falta de oxígeno (Pickering, 1981).
El crecimiento de los peces es un asunto complejo resultado directo de procesos bioquímicos, osmóticos y otros factores que contribuyen al aporte de material en el organismo (Tresierra & Culquichicón, 1993) y que puede afectarse por diversas variaciones en el ambiente que interactúan con el metabolismo. Por lo tanto, llevar a cabo estudios experimentales que analicen el efecto en el crecimiento y en la formación de marcas en las estructuras duras sería un avance importante en el necesario entendimiento de estos procesos.
Con el propósito de explorar la influencia de cuatro factores ecológicos en el crecimiento de O. niloticus, se llevó a cabo un estudio experimental donde la hipótesis nula fue que la reducción de la temperatura, cantidad de alimento, oxígeno y volumen de agua no afecta el crecimiento de O. niloticus. Para evaluar el impacto de estos factores ecológicos en el crecimiento, se analizaron las variaciones de la talla, el peso y las distancias inter-circuli en las escamas, considerando que en condiciones de estrés ambiental las distancias inter-circuli de las escamas se reducen, al igual que disminuye el incremento de la talla y el peso en comparación con el control.
Antes del inicio del experimento y de la introducción de los organismos, se requirió estabilizar los sistemas para obtener las condiciones necesarias para cada tratamiento y el control. Esto se realizó durante treinta días, al regular las condiciones de temperatura y concentración de oxígeno disuelto y registrar puntualmente estos valores. Así, la concentración de oxígeno se mantuvo entre 4.3 a 5.6 mg/L (en promedio 4.98 ± 0.5 mg/L) para todos los tratamientos con excepción del tratamiento de la concentración de oxígeno; para disminuir su concentración, se desconectó el filtro del sistema por un periodo de siete horas (filtros Fluval modelo MS306 funcionando como un sistema de recirculación), y se estabilizó en 2.8 ± 1.2 mg/L. Para el tratamiento de disminución de la temperatura, se instaló un enfriador (Puresa, modelo DC-300), con lo cual se redujo la temperatura de 25.1 ± 1.0 hasta 19.6 ± 1 °C. Para regular la temperatura del agua se utilizaron calentadores de 100 y 150 W. Se realizaron reposiciones de agua en las tinas que presentaban mayor evaporación, y la limpieza de los filtros se realizó cada tercer día. Cada tercer día, se determinaron los fosfatos, amonio y nitratos con un fotómetro marca Hanna® HI83203.
El experimento se realizó en cinco tanques de plástico de 420 L, que se llenaron hasta 215 L de volumen efectivo, cada tina se dividió por medio de una cortina plástica para formar un duplicado por cada condición experimental; cuatro de las tinas se utilizaron para los tratamientos experimentales y una para el control. Las condiciones control fueron las óptimas reportadas para la especie de acuerdo con el Programa de información de especies acuáticas (FAO, 2006): 5 % de su peso total en alimento; 3 peces/L de densidad; saturación total de O2 (5.03 ± 0.2 mg/L) y 25.1 ± 1.0 oC de temperatura. Los tratamientos variaron como sigue: 1) El alimento fue reducido a 2.5 % del peso total; 2) La densidad se duplicó a 6 peces/L, reduciendo a la mitad el volumen de agua; 3) el oxígeno se redujo a 2.8 ± 1.2 mg/L y, 4) la temperatura se redujo a 19.6 ± 1.0 oC, como se explicó anteriormente. El alimento suministrado a todos los tratamientos y control, consistió en pelets Nutripec marca Purina para desarrollo con 44 % de proteína, el cual se suministró en dos dosis, por la mañana a las 10 am y por la tarde a las 17 hrs. El oxígeno fue determinado con el método de Winkler y la temperatura con un termómetro de mercurio de escala de -20 a 110 oC. A cada organismo se le extrajeron cuatro escamas cerca de la zona del opérculo, estas fueron enjuagadas con una solución jabonosa y una vez limpias fueron colocadas entre dos portaobjetos sujetados con cinta y etiquetadas con el nombre del tratamiento al que pertenecían. Los espacios intercirculi fueron empleados como una variable en representación de la tasa de crecimiento (Fisher & Pearcy, 1990, 2005; Barber & Walker, 1988). Para realizar esta medida, se usó un microscopio invertido Invertoscop-D, Karl Zeiss, con ocular micrométrico Kpl W 8x/20, calibrado con una reglilla micrométrica de 1 mm con 100 subdivisiones (0.01 mm cada división). Se midieron los cuatro espacios inter-circuli externos más alejados del foco de la escama en un ángulo de 45º, considerados éstos como indicadores de las últimas condiciones ambientales.
El experimento se dividió en tres etapas que se describen a continuación.
Para esta etapa se tomaron, sin haber sido sometidos a las condiciones experimentales, la talla y el peso de 40 juveniles de O. niloticus (20 por cada duplicado) por tratamiento y el control. Así mismo, se obtuvieron escamas de cuatro de los organismos por tratamiento y control. En el día uno, se midieron y pesaron los ejemplares que fueron elegidos al azar de un lote de aproximadamente 300 individuos, con una longitud total de entre 4 a 6 cm con promedio de 4.6 ± 0.3 cm, y que previamente habían sido aclimatados durante una semana. La longitud total se midió con un ictiómetro con precisión de ± 1 mm, mientras que el peso se obtuvo con una balanza digital (± 0.1 g) Ohaus modelo CT1200-S, con capacidad de 1 200 x 0.1 g. Para la obtención de escamas, se eligieron azarosamente ocho organismos por tratamiento (cuatro por duplicado); esto representó el 20 % del total de los ejemplares, lo cual se consideró una muestra suficiente para representar el total. De cada ejemplar se obtuvieron cuatro escamas que fueron removidas de la región posterior al opérculo.
Al término del período experimental, luego de 24 días de duración de cada etapa, se obtuvo nuevamente una muestra de cuatro escamas de la serie lateral de los peces, con el objeto de analizar las distancias inter-circuli y así observar el efecto del tratamiento en la formación de marcas del crecimiento, así mismo los peces fueron medidos y pesados.
Seguidamente, los cuatro tratamientos fueron revertidos a las condiciones del control; se consideró observar crecimiento compensatorio una vez que se regresara a las condiciones óptimas sin estrés. Esta etapa tuvo una duración de 24 días adicionales. Al término de la etapa compensatoria, se obtuvo nuevamente una muestra de cuatro escamas de la serie lateral de los peces, con el objeto de analizar las distancias intercirculi, y así observar el efecto del posible crecimiento compensatorio en las medidas de talla y peso y escamas analizadas.
Los datos obtenidos de las mediciones realizadas durante las tres fases estudiadas, que incluyeron pesos, tallas y espacios intercirculi, se sometieron a pruebas de Omnibus de normalidad para cada una de las etapas del estudio; posteriormente, se realizaron una serie de ANOVAS para las distintas etapas del estudio y así estimar si había diferencias entre los tratamientos; finalmente, se hizo una prueba de comparación de Duncan para reconocer a los tratamientos que eran significativamente diferentes en cada una de las etapas.
El experimento se realizó entre septiembre y diciembre del 2015.
Debido a que no fueron detectadas diferencias significativas para la talla, peso y distancias inter-circuli entre los duplicados (P = 0.75), los resultados se muestran por tratamiento.
La talla no mostró diferencias significativas entre el control y los tratamientos para el inicio de la etapa experimental (Fig. 1). Sin embargo, al final de la etapa experimental las diferencias significativas (P < 0.05) se presentaron para los tratamientos de alimento y temperatura; igualmente, sucedió para la etapa compensatoria (Fig. 1). Se observa claramente en la figura 1 que los tratamientos de alimento y temperatura presentaron los ejemplares de menor talla, donde los valores promedio fueron de 5.8 ± 0.5 cm y 5.6 ± 0.4 cm, respectivamente, para el final de la etapa experimental, y de 8.0 ± 1.1 cm para ambos, en la etapa compensatoria.
Al final de la etapa experimental, los valores promedio mayores en la talla los presentó el tratamiento control (6.7 ± 0.5 cm), seguido por el de oxígeno (6.6 ± 0.6 cm) y el de densidad (6.4 ± 0.4 cm), P > 0.05. Para la etapa compensatoria, los valores promedio mayores en la talla los presentó el tratamiento oxígeno (9.3 ± 1.2 cm), seguido por el control (8.9 ± 1.1 cm), y posteriormente, la densidad (8.7 ± 1.0 cm), P > 0.05.
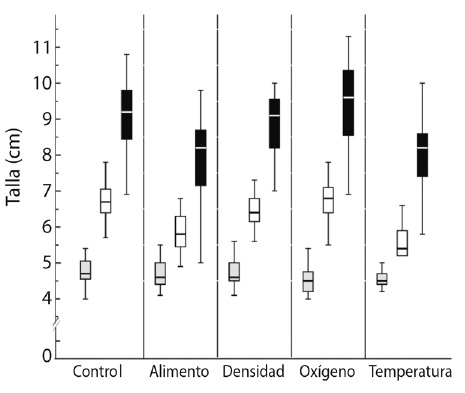
Fig. 1
Boxplot depicting size (cm) for Control and the treatments: initial (light grey) and end of experiment (white) and compensatory stage (black).
El peso mostró diferencias significativas (P < 0.05) para el inicio de la etapa experimental en los tratamientos de oxígeno (5.2 ± 1.4 g) y temperatura (3.4 ± 0.7 g) (Fig. 2). Al final de las etapas experimental y compensatoria, las diferencias significativas (P < 0.05) se presentaron entre los tratamientos alimento (9.7 ± 3.7 g) y temperatura (10.0 ± 3.9 g) (Fig. 2). Al final de la etapa experimental, los valores promedio mayores en peso los presentó el grupo control (5.2 ± 1.2 g), seguido por el tratamiento de oxígeno (5.2 ± 1.4 g) y el de densidad (4.6 ± 0.9 g). Al final de la etapa compensatoria, los valores promedio mayores en el peso los presentó el tratamiento de oxígeno (15.9 ± 6.2 g), seguido por el control (14.6 ± 5.2 g), y el de la densidad (13.6 ± 4.5 g).
No hubo diferencias en los espacios inter-circuli en la etapa experimental de inicio, es decir los espacios inter-circuli eran iguales al inicio del experimento mientras que al final del experimento todas las medidas de los espacios fueron distintas y en la etapa compensatoria solamente los tratamientos de alimento y temperatura difirieron (P < 0.5), es decir, éstos no promovieron un aumento de los espacios inter-circuli como lo hicieron el resto de los tratamientos (Fig. 3).
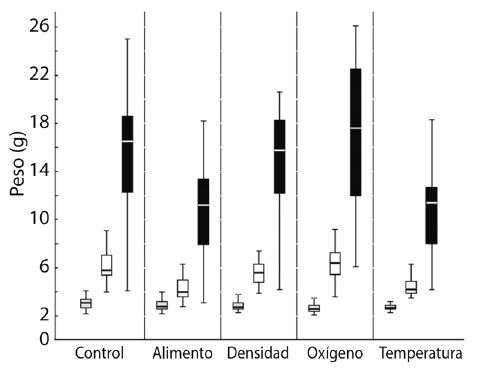
Fig. 2
Boxplot depicting weight (g) for Control and the treatments: initial (light grey) and end of experiment (white) and compensatory stage (black).
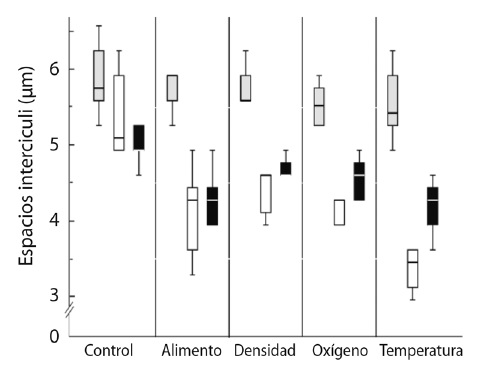
Fig. 3
Boxplot depicting intercirculi spaces (mm) for Control and the treatments: initial (light grey) and end of experiment (white) and compensatory stage (black).
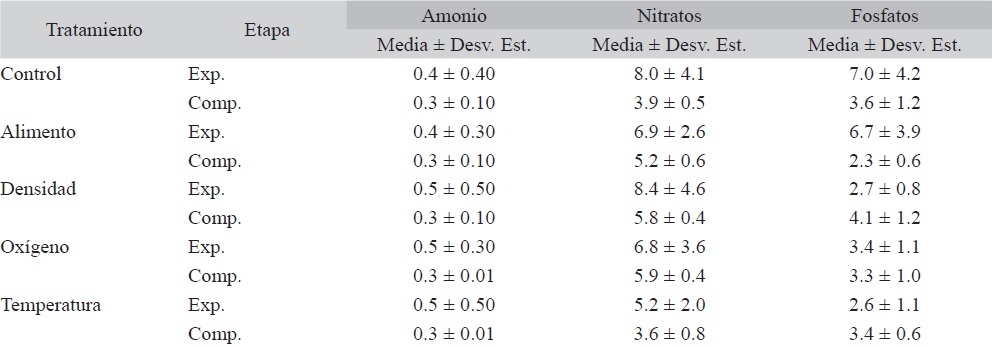
Valores promedio ± Desviación estándar de concentraciones en mg/L, para amonio, nitratos y fosfatos en la etapa experimental (Exp.) y compensatoria (Comp.) en el Control y los cuatro tratamientos
Al inicio de la etapa experimental los valores promedio para el espacio inter-circuli en todos los casos fue de 5.8 ± 0.5 mm y que, como se mencionó antes, no se presentaron diferencias significativas. Para el final de la etapa experimental, los valores mayores en promedio para el espacio inter-circuli los presentó el control (5.4 ± 0.5 mm), seguido por el tratamiento de la densidad (4.2 ± 0.5 mm), el del oxígeno y el del alimento (4.1 ± 0.5 mm; 4.1 ± 0.6 mm, respectivamente), mientras que los valores más bajos se presentaron para el tratamiento de la temperatura (3.4 ± 0.3 mm).
En la etapa compensatoria, los valores promedio mayores en el espacio inter-circuli los presentó el tratamiento control (4.9 ± 0.2 mm), seguido por el de la densidad (4.7 ± 0.2 mm), el del oxígeno y el alimento (4.6 ± 0.4 mm; 4.3 ± 0.4 mm, respectivamente). Los valores más bajos se presentaron para el tratamiento de la temperatura (4.2 ± 0.3 mm).
Los valores promedio de amonio variaron de 0.3 a 0.5 mg/L con una media general de 0.4 ± 0.23 mg/L, y sin diferencias significativas. En el caso de los nitratos y fosfatos, los valores promedio variaron de 3.6 a 8.4 y 2.3 a 7.0 mg/L, con promedios generales de 6.0 ± 1.96 y 3.9 ± 1.57 mg/L, respectivamente (Cuadro 1).
Se presentó mortalidad únicamente en el período experimental para los tratamientos de oxígeno y temperatura, donde murieron cuatro y cinco ejemplares, respectivamente (Cuadro 2).
Número de organismos vivos en cada etapa del estudio
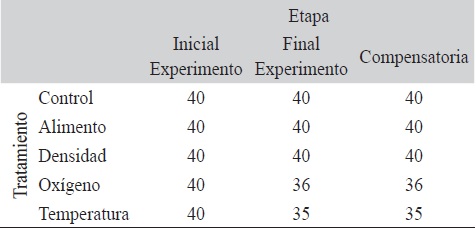
Sin embargo, en el caso del tratamiento del oxígeno, dos de los ejemplares murieron al introducirse por el tubo de aireación, por lo que se considera que solamente dos ejemplares murieron por efecto de la reducción del oxígeno. En ningún tratamiento se reemplazaron los organismos perdidos.
Rechazamos la hipótesis nula planteada, es decir, la reducción de la temperatura, cantidad de alimento, oxígeno y densidad sí afecta el crecimiento de O. niloticus. Los resultados indican que el efecto de los estresores parece ser claro, todos los probados aquí con el incremento de la densidad, la disminución del alimento, oxígeno y temperatura, produjeron menor talla, peso y espacios inter-circuli, es decir, los cuatro son potenciales formadores de marcas en las escamas. Sin embargo, solamente el tratamiento del alimento y el de la temperatura resultaron estadísticamente diferentes. Es posible que los niveles de densidad y oxígeno empleados en este estudio, no fueran suficientemente severos para los peces o posiblemente el tiempo de experimentación no fue el suficiente como para producir un efecto decisivo y estadísticamente significativo, como los observados en los estudios de Abdel-Tawwab, Hagras, Elbaghdady y Monier (2014); ellos reportaron que la baja concentración de oxígeno disuelto (1.0 a 3.0 mg/L), causó la reducción del crecimiento y ritmo de ingesta de alimento de O. niloticus. También probaron que el factor densidad, al incrementar aunado a la interacción de los dos factores, provocó un descenso en la tasa de crecimiento de forma significativa. Así mismo, la talla pequeña de nuestros ejemplares, pudo ser afectada por la reducción de oxígeno ya que en otro experimento AbdelTawwab, Hagras, Elbaghdady y Monier (2015) comprobaron que los peces pequeños fueron más susceptibles a retardar el crecimiento, ante la baja concentración de oxígeno disuelto. Así mismo, Tran-Duy, van Dam y Schrama (2012) concluyeron que O. niloticus crece menos a bajas concentraciones de oxígeno (1.1 a 2.6 mg/L) y también, disminuye la ingesta de alimento; en esta especie los peces casi adultos no modifican la ingesta, pero los peces pequeños pueden disminuir radicalmente la alimentación.
En cuanto al efecto del crecimiento en la etapa compensatoria, se esperaba que los tres indicadores, talla, peso y espacios inter-circuli regresaran a los niveles del control, pero sin embargo, esto no sucedió. Todos los ejemplares aumentaron en talla y peso durante esta etapa, aunque solo el tratamiento de oxígeno lo hizo al mismo nivel que el control, mientras que los tratamientos de alimento y temperatura, mostraron una recuperación menos vigorosa. Wang, Cui, Yang y Cai (2000) han reportado que el híbrido O. mossambicus X O. niloticus posee débil capacidad de crecimiento compensatorio cuando se expone a periodos de deprivación de alimento mayores a una semana, similares a los utilizados en nuestro experimento. Así mismo mencionan que este híbrido tarda hasta cuatro semanas en tener un índice de recuperación compensatorio similar a los organismos alimentados de manera regular; ellos creen que esto provocó la gran pérdida de peso durante los experimentos realizados. A similares conclusiones llegaron Yang, Ziwei, Junwook y Jeong-Yeol (2015) para O. niloticus.
Diversos autores han reportado que la temperatura del agua es uno de los factores ambientales con mayor impacto en las respuestas fisiológicas de crecimiento y alimentación (Xie, Zheng, Chen, Zhang, Zhu, & Yang, 2011). Kapetsky y Nath (1997) mencionan que la disponibilidad de alimento y la temperatura del agua, parecen ser factores limitantes para el crecimiento de O.niloticus. Se han determinado intervalos de mayores crecimientos de O. niloticus en temperaturas de 28 y 32 °C (Likongwe Stecko, Stauffer, & Carline, 1996), 31 a 36 °C (FAO, 2006) y 30.1 °C (Xie et al., 2011). La reducción de la temperatura detiene la reproducción cuando se registran temperaturas de 22 °C. La alimentación se detiene por debajo de los 20 °C (Wolhfarth, Hulata, Rothbard, Itzkowich, & Halevy, 1983), lo cual concuerda con la temperatura utilizada en nuestro experimento; sin llegar a las temperaturas que producen mortalidad (8 - 11 °C) reportadas por Stauffer, Boltz y Boltz (1988), Chervinski y Lahav (1976), y Kindle y Whitmore (1986) para O. aureus (Steindachner, 1864). Así mismo, se ha encontrado que el incremento de depositación en otolitos o estructuras calcáreas puede ser afectado por distintos factores ambientales, entre los más importantes están el fotoperiodo, temperatura y alimentación. Los efectos de estos factores, en la tasa de depositación de otolitos han sido investigados en condiciones extremas; como lo es la inanición o la baja temperatura (Campana & Neilson, 1985).
A pesar de haber revertido las condiciones de estrés a condiciones óptimas de cultivo o condiciones del control en la etapa compensatoria, los tratamientos de temperatura y alimentación siguieron significativamente diferentes respecto al Control y el resto de los tratamientos, ya que los organismos comúnmente reestructuran sus procesos de absorción cuando están expuestos a nuevas condiciones ambientales durante largos períodos de tiempo; también se ha observado que cambios sostenidos en la dieta a menudo también provocan ajustes crónicos en la cantidad de enzimas digestivas producidas por el organismo (Hill, Gordon, & Margaret, 2004).
Para el cultivo de tilapia se han mostrado rangos de tolerancia para amonio desde 0.001 a 0.2 mg/L por El-Sayed (2006) y de hasta 0.35 a 0.5 mg/L por Williams (2000). En el caso de nitratos Wicki (1997) reporta tolerancias a concentraciones de hasta 100 mg/L, mientras que para los fosfatos los valores de tolerancia son entre 0.6 a 1.5 mg /L (Nicovita, 2007). La calidad del agua tanto para los tratamientos como el control, se encontró dentro de los rangos de tolerancia para la especie con excepción de los valores elevados de fosfatos, sin embargo, al presentarse valores altos en todos los tratamientos y el control, no se considera que sea un estresor adicional.
La reducción de alimento disponible, así como la disminución en la temperatura del agua, retardaron significativamente el crecimiento de O. niloticus. La concentración de oxígeno disuelto y la reducción de la proporción biomasa/volumen no mostraron diferencias estadísticamente significativas con respecto al control. En estudios previos las distancias intercirculi han mostrado ser un buen indicador de la tasa de crecimiento (Fisher & Pearcy, 1990, 2005; Barber & Walker, 1988) en este estudio también se reducen las distancias inter-circuli en periodo de estrés de los organismos.
Los fondos del estudio fueron aportados por la Universidad Autónoma MetropolitanaIztapalapa (UAMI).
Fig. 1
Boxplot depicting size (cm) for Control and the treatments: initial (light grey) and end of experiment (white) and compensatory stage (black).
Fig. 2
Boxplot depicting weight (g) for Control and the treatments: initial (light grey) and end of experiment (white) and compensatory stage (black).
Fig. 3
Boxplot depicting intercirculi spaces (mm) for Control and the treatments: initial (light grey) and end of experiment (white) and compensatory stage (black).
Valores promedio ± Desviación estándar de concentraciones en mg/L, para amonio, nitratos y fosfatos en la etapa experimental (Exp.) y compensatoria (Comp.) en el Control y los cuatro tratamientos
Número de organismos vivos en cada etapa del estudio
