Artículos
New data on Pyraloidea from the Canary Islands, Spain 2 (Lepidoptera: Pyralidae, Crambidae)
Nuevos datos sobre los Pyraloidea de las Islas Canarias, España 2 (Lepidoptera: Pyralidae, Crambidae)
 O. Karsholt okarsholt@snm.ku.dk
F. Slamka f.slamka@nextra.sk
O. Karsholt okarsholt@snm.ku.dk
F. Slamka f.slamka@nextra.sk
New data on Pyraloidea from the Canary Islands, Spain 2 (Lepidoptera: Pyralidae, Crambidae)
SHILAP Revista de lepidopterología, vol. 50, núm. 199, pp. 469-488, 2022
Sociedad Hispano-Luso-Americana de Lepidopterología

Esta obra está bajo una Licencia Creative Commons Atribución 4.0 Internacional.
Recepción: 17/02/2022
Aprobación: 05/03/2022
Publicación: 30/09/2022
Abstract: We describe six new species of Pyraloidea from the Canary Islands, Spain: Dioryctria mieella Falck, Karsholt & Slamka, sp. n., Euzophera mirumella Falck, Karsholt & Slamka, sp. n., Dalakia moyalis Falck, Karsholt & Slamka, sp. n., Peoria gigantesalis Falck, Karsholt & Slamka sp. n. (Pyralidae: Phycitinae), Metasia (Clasperia) baezi Falck, Karsholt & Slamka, sp. n. (Crambidae: Spilomelinae) and Cynaeda acutalis Falck, Karsholt & Slamka, sp. n. (Crambidae: Odontiinae). Eight species are recorded as new to the fauna of the Canary Islands: Achroia grisella (Fabricius, 1794) (Pyralidae: Galleriinae), Alophia combustella (Herrich-Schäffer, 1855), Epischnia prodromella (Hübner, [1799]), Epischnia illotella Zeller, 1839, Assara conicolella (Constant, 1884), Homoeosoma stenotea Hampson, 1926 (Pyralidae: Phycitinae), Loxostege sticticalis (Linnaeus, 1761) and Achyra nudalis (Hübner, 1796) (Pyralidae: Pyraustinae). Hypsopygia rubidalis ([Denis & Schiffermüller], 1775) should be removed from the list of Lepidoptera found in the Canary Islands and replaced by Hypsopygia costalis (Fabricius, 1775) (Pyralidae: Pyralinae). We argue that Pima tricolorella Falck, Karsholt & Slamka, 2019 is a bona species. Photographs of adults of all species are shown. Photographs of the genitalia of new species are shown. Five of the six new species are Barcode and DNA analyses support the identification as new species.
Keywords: Lepidoptera, Pyralidae, Crambidae, new species, new records DNA barcodes, Canary Islands, Spain.
Resumen: Describimos seis nuevas especies de Pyraloidea de las Islas Canarias, España: Dioryctria mieella Falck, Karsholt & Slamka, sp. n., Euzophera mirumella Falck, Karsholt & Slamka, sp. n., Dalakia moyalis Falck, Karsholt & Slamka, sp. n., Peoria gigantesalis Falck, Karsholt & Slamka sp. n. (Pyralidae: Phycitinae), Metasia (Clasperia) baezi Falck, Karsholt & Slamka, sp. n. (Crambidae: Spilomelinae) and Cynaeda acutalis Falck, Karsholt & Slamka, sp. n. (Crambidae: Odontiinae). Se registran ocho especies como nuevas para la fauna de las Islas Canarias: Achroia grisella (Fabricius, 1794) (Pyralidae: Galleriinae), Alophia combustella (Herrich-Schäffer, 1855), Epischnia prodromella (Hübner, [1799]), Epischnia illotella Zeller, 1839, Assara conicolella (Constant, 1884), Homoeosoma stenotea Hampson, 1926 (Pyralidae: Phycitinae), Loxostege sticticalis (Linnaeus, 1761) y Achyra nudalis (Hübner, 1796) (Pyralidae: Pyraustinae). Hypsopygia rubidalis ([Denis & Schiffermüller], 1775) debería ser removida de la lista de Lepidoptera encontrados en las Islas Canarias y remplazada por Hypsopygia costalis (Fabricius, 1775) (Pyralidae: Pyralinae). Argumentamos que Pima tricolorella Falck, Karsholt & Slamka, 2019 es una buena especie. Se muestra fotografías de los adultos de todas las especies. Se muestran fotografías de la genitalia de las nuevas especies. Cinco de las seis nuevas especies con Barcode y los análisis de ADN respaldan la identificación como nuevas especies.
Palabras clave: Lepidoptera, Pyralidae, Crambidae, nuevas especies, nuevos registros, ADN código de barras, Islas Canarias, España.
Introduction
In a previous paper (FALCK et al., 2019) we dealt with the Pyraloidea fauna of the Canary Islands, presenting an overview of the literature and raising the number of species found in these islands from 132 to 152. Subsequent field work by the first author has resulted in the discovery of additional species of Pyraloidea, which are dealt with below.
During the preparation of our previous paper, we were aware of that not all species known to us from the Canary Islands could be identified. Based on ongoing research we have been able to identify some of these, whereas others proved to be undescribed; we can now describe six new species, mainly from material collected by the first author.
One species, Pima tricolorella Falck, Karsholt & Slamka, 2019, described by us in our previous paper (FALCK et al., 2019: 35) has since been given new status as a subspecies (LERAUT, 2021a, b). We disagree in this and provide additional information to confirm its status as a separate species.
Material and methods
Most of the specimens were attracted to an 8 watt. super actinic light. Genitalia were dissected following ROBINSON (1976). Whole specimens were photographed with a Canon EOS 700D camera equipped with a Canon EF 100 mm objective. The genitalia slides were photographed using a Soptop CX40T Trinocular microscope in conjunction with a Toup Tek P10500A-E3 / E3ISPM05000KPA-E3 / 5.0MP USB3 camera.
DNA samples were prepared from dried legs according to the prescribed standards and processed at the Canadian Centre for DNA Barcoding (CCDB, Biodiversity Institute of Ontario, University of Guelph) to obtain the 658 base-pair long barcode fragment of the mitochondrial COI gene (cytochrome c oxidase I). Intra- and interspecific distances of DNA barcode fragment were calculated using analytic tools of BOLD with the Kimura 2-parameter model of nucleotide substitution. Genetic clusters are presented with their barcode index number (BIN; cf. RATMNASINGHAM & HERBERT, 2013).
We examined the morphology from all species and the DNA Barcode from new and cryptic species except M. baezi.
The sequence for the species treated below follows VIVES MORENO (2014).
Abbreviations used
FS Frantisek Slamka, Bratislava, Slovakia
GP Genitalia preparation
MB Collection of Marcos Báez, La Laguna, Tenerife, Spain
PF Collection of Per Falck, Neksø, Denmark
MNCN Collection of Antonio Vives, Museo Nacional de Ciencias Naturales, Madrid, Spain
NHMUK Natural History Museum, London, United Kingdom
ZMUC Zoological Museum, Natural History Museum of Denmark, Copenhagen, Denmark
Results
PYRALIDAE
GALLERIINAE
Achroia grisella (Fabricius, 1794) (Fig. 1)
![1. Achroia grisella (Fabricius, 1794), ♀, La Gomera, 20.5 mm. 2.Alophia combustella (HerrichSchäffer, 1855), ♀, Gran Canaria, 20.5 mm. 3.Dioryctria mieella Falck, Karsholt & Slamka, sp. n., ♂, Tenerife, 23 mm. 4.Epischnia prodromella (Hübner, [1799]), ♂, Gran Canaria, 28 mm. 5.Epischnia illotella Zeller, 1839, ♀, Gran Canaria, 20 mm. 6.Assara conicolella (Constant, 1884), ♂, Tenerife, 19.5 mm. 7.Euzophera mirumella Falck, Karsholt & Slamka, sp. n., ♂, Gran Canaria, 29.5 mm. 8.Homoeosoma stenotea Hampson, 1926, ♀, Lanzarote, 15 mm.](../45573669014_gf2.png)
Figs 1-8.–
1. Achroia grisella (Fabricius, 1794), ♀, La Gomera, 20.5 mm. 2.Alophia combustella (HerrichSchäffer, 1855), ♀, Gran Canaria, 20.5 mm. 3.Dioryctria mieella Falck, Karsholt & Slamka, sp. n., ♂, Tenerife, 23 mm. 4.Epischnia prodromella (Hübner, [1799]), ♂, Gran Canaria, 28 mm. 5.Epischnia illotella Zeller, 1839, ♀, Gran Canaria, 20 mm. 6.Assara conicolella (Constant, 1884), ♂, Tenerife, 19.5 mm. 7.Euzophera mirumella Falck, Karsholt & Slamka, sp. n., ♂, Gran Canaria, 29.5 mm. 8.Homoeosoma stenotea Hampson, 1926, ♀, Lanzarote, 15 mm.
Material examined: Spain, La Gomera, Arure, 830 m, 1 ♀, 24-X-12-XI-2021, leg. P. Falck, genitalia slide 3559PF (PF). New to the Canary Islands.
Distribution: Cosmopolitan (SLAMKA, 2006: 59).
Biology: The larva lives in webs and feeds on wax in hives and can be a pest there, occasionally also on vegetal detritus and dead insects (SLAMKA, 2006: 59).
Remarks: The species is probably a resident in the Canary Islands.
PYRALINAE
Hypsopygia costalis (Fabricius, 1775)
Correction. In our last paper (FALCK et al., 2019: 35) this species was erroneously recorded as Hypsopygia rubidalis ([Denis & Schiffermüller], 1775). H. rubidalis should be removed from the list of Lepidoptera found in the Canary Islands and replaced by H. costalis.
PHYCITINAE
Pima tricolorella Falck, Karsholt & Slamka, 2019, sp. rev.
The status of this species, described by us from Lanzarote and Fuerteventura in our earlier paper on Pyraloidea of the Canary Islands (FALCK et al., 2019: 35-37) was recently changed by LERAUT (2021a:184, 318; 2021b: 89) into a subspecies of Pima aureliae Leraut, 1914 from Tunisia. We disagree in this for the following reasons:
Adult: P. tricolorella differs from P. aureliae in the shape of the forewing, which is narrow, having the costa more rounded in the apical half and the termen oblique and in the nature of the white streak at the costa, which is well-developed only in the basal third.
Male genitalia: The two species are distinguished by the shape of the two large cornuti. Both species display one cornutus that is slender and pointed; the other cornutus is broad at the base, tapering towards the apex and ends in a strong spike in P. tricolorella, whilst in P. aureliaea this cornutus is broad at the base but is almost triangular tapering equally on both sides towards apex.
Female genitalia of P. tricolorella are difficult to compare with the published figures of P. aureliae (LERAUT 2014: 331, fig.122c; LERAUT 2021b: 89, fig. 2; SLAMKA 2019: 324, fig. 147b), because the difference between the female genitalia of the three specimens is quite large, especially in the left angle between the ductus bursae and the corpus bursae and in the sclerotized structures in corpus bursae. In our opinion the specimens don’t belong to the same species. P. tricolorella is distinguished in the female genitalia by the relatively shorter anterior apophysis, the barrel-shaped antrum having the lateral edges well sclerotized and tergite VIII with a small u-shaped excavation posteriorly. In P. aurelia tergite VIII has a deep v-shaped incision posteriorly. Additionally, the left angle between the ductus bursae and the corpus bursae is greater than that figured in a paratype by P. Leraut, but smaller than in the Algerian specimen figured by G. Leraut.
Alophia combustella (Herrich-Schäffer, 1855) (Fig. 2)
Material examined: Spain, Gran Canaria, Barranco de Azuaje, 270 m, 1 ♂, 3 ♀♀, 8-20-VIII-2020, leg. P. Falck, genitalia slide 3528PF; Carreteria, 455 m, 1 ♂, 2 ♀♀, 8-20-VIII-2020, leg. P. Falck; Fontanales 1100 m, 1 ♂, 8-20-VIII-2020, leg. P. Falck; La Gomera, Hermigua, 250 m, 1 ♂, 1 ♀, 9-12- VIII-2021, leg. P. Falck; Tenerife, Arona, 670 m, 1 ♀, 21-V-3-VI-2019, leg. P. Falck; 8 km S Aguamansa, 1400 m, 1 ♀, 21-V-3-VI-2019, leg. P. Falck (all PF). New to the Canary Islands.
Distribution: From southern Europe and North Africa (Morocco, Algeria, Tunisia) to the Middle East and Russia (LERAUT, 2014: 119; SLAMKA, 2019: 87).
Biology: The larva lives in aphid (Aphidoidea) galls on leaves of Pistacia lentiscus L., P. terebrinthus L., and P. atlantica Desf. (LERAUT, 2014: 119; SLAMKA, 2019: 87).
Remarks: The species is a resident of the Canary Islands.
Dioryctria mieella Falck, Karsholt & Slamka, sp. n. (Fig. 3)
Holotype ♂: Spain, Tenerife, Arona, 670 m, 21-V-3-VI-2019, leg. P. Falck, genitalia slide 3526PF, DNA sample Lepid Phyl 0938PF/CILEP0937-21 (MNCN).
Paratypes: Spain, La Gomera, Hermigua, 250 m, 1 ♀, 24-X-12-XI-2021, leg. P. Falck, genitalia slide 3560PF (PF); Tenerife, Güimar, 500 m, 1 ♂, 3-16-VIII-2021, leg. P. Falck, DNA sample Lepid Phyl 0937PF/CILEP0936-21 (PF).
Description: Male. Wingspan 22-23 mm. Head and neck light reddish-brown. Antenna about 2/3 length of forewing, light grey ringed with blackish grey, the first 1/3 pectinated, the rest of antenna distinctly serrate, scapus and flagellum light reddish brown with a few black scales edging sinus. Labial palps slightly upturned, segment 2 reddish brown, medially whitish, segment 3 brown with a whitish tip. Thorax and tegulae light reddish brown. Forewing narrow, elongate with rather pointed apex; ground colour light-reddish brown; basal streak white; ante-medial line white, zigzagging; post-medial line sinous, white, bordered by black scales medially in costal 2/3; discal spot distinct, white; medial part with two longitudinal black streaks combining the lines, upper black streak divided by the discal spot; marginal line black; fringes light reddish brown. Hindwing dark grey; fringes grey. Abdomen reddish grey.
Female: Externally similar to male, antenna filiform with short cilia.
Male genitalia (Figs 17, 17a): Uncus vertically oval, apically rounded with short setae; gnathos short and narrow, spicate, strongly sclerotized, bilaterally with small rounded projections on the posterior part of the short tegumen; valva long and slender, partially sclerotized, apex with long and thin spicate process about ⁄ of the length of valva; fibula (harpe) oblong, edges setose, situated above the sacculus; dorsal part of valva weakly sclerotized covered densely with long setae; juxta doublearmed, U-shaped, apically weakly spicate and shortly setose; saccus long and narrow, well sclerotized. Phallus of moderately length, straight, apically with long spicate cornutus placed on the sclerotized structure and coated by the quantity of fine spines (part of vesica); additional short and thin cornuti (about 50) situated below the main cornutus.
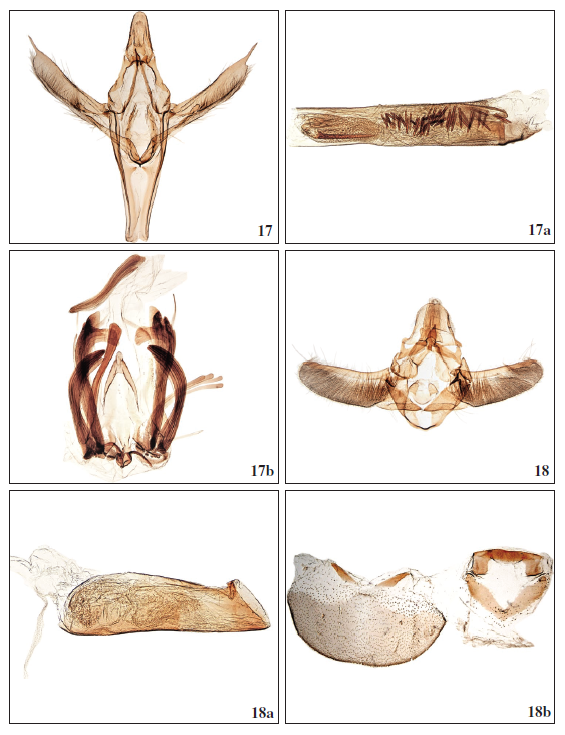
Figs 17-18b.–
17.Dioryctria mieella Falck, Karsholt & Slamka, sp. n., ♂, Tenerife, GP3526PF. 17a. Phallus, Tenerife, GP3526PF. 17b. Culcita, Tenerife, GP3526PF. 18.Euzophera mirumella Falck, Karsholt & Slamka, sp. n., ♂, Gran Canaria, GP3562PF. 18a. Phallus, Gran Canaria, GP3556PF. 18b. Segment VIII (tergit and sternit), Gran Canaria, GP3562PF
Culcita (modified sternum VIII) (Fig. 17b) paired and bearing latero-ventral scale tufts.
Female genitalia (Fig. 23): Papillae anales short, densely covered by setae; posterior and anterior apophysis of the same length; segment VIII well sclerotized, square-shape, posteriorly with long setae; ostium rounded, strongly sclerotized, wrinkled, bilaterally with stronger sclerotized longitudinal folds; ductus bursae long, strongly sclerotized, in the central part longitudinally ribbed, inferiorly vertically Sshaped; corpus bursae oblong, generally membranous, posteriorly with the quantity of short spies situated in irregular arcs and clusters.

Figs 22-25.–
22.Cynaeda acutalis Karsholt & Slamka, sp. n., ♂, Gran Canaria, GP3578PF. 22a. Phallus, Gran Canaria, GP3578PF. 23.Dioryctria mieella Falck, Karsholt & Slamka, sp. n., ♀, La Gomera, GP3560PF. 24.Euzophera mirumella Falck, Karsholt & Slamka, sp. n., ♀, Gran Canaria, GP3557PF. 25.Dalakia moyalis Falck, Karsholt & Slamka, sp. n., ♀, Gran Canaria, GP3548PF.
DNA barcodes: We obtained full length DNA barcode (658 bp) from one specimen and DNA barcode fragments of 654 bp from one specimen. The barcodes fall within Barcode Index Number (BIN) BOLD: AEM6898. The maximum intraspecific distance is 0.00%. The minimum p-distance to nearest neighbour Dioryctria nivaliensis Rebel, 1892 is 6.14%, and the minimum p-distance to D. reniculelloides Mutuura & Munroe, 1973 is 6.38%.
Diagnosis: Externally D. mieella only resembles a few reddish North American Dioryctriaspecies, e. g. D. auranticella (Grote, 1883) and D. disclusa Heinrich, 1953. It is distinguished by the narrow and pointed forewing and by the two longitudinal black streaks in the medial area. From all European species of Dioryctria it is distinguished by the red colour and shape of the forewing. In the male genitalia the long spicate process at the apex of the valva and small rounded harpe are characteristic. In the female genitalia, the large rounded ostium, and the S-shaped ductus bursae are characteristic.
Biology: Early stages unknown. The specimens were attracted to light.
Distribution: Only known from the islands of Tenerife and La Gomera, Spain.
Etymology: The species name is dedicated to the first author’s youngest daughter Mie.
Remarks: D. mieella is probably endemic to the Canary Islands as it is the case with D. nivaliensis, these two species being each other’s closest relatives according to their DNA barcodes.
Epischnia prodromella (Hübner, [1799]) (Fig. 4)
Material examined: Spain, Gran Canaria, Pie de la Cuesta, 500 m, 4 ♂♂, 4-23-III-2019, leg. P. Falck, genitalia slide 3538PF, DNA samples 0941PF/CILEP0940-21, 0942PF/CILEP0941-21; Ayacata, 1400 m, 1 ♂, 4-23-III-2019, leg. P. Falck (all PF). New to the Canary Islands.
Distribution: South- and Central Europe eastwards to Russia. North Africa (Morocco, Algeria, Tunisia), The Middle East and eastwards to Turkey and Iran (LERAUT, 2014: 315).
Biology: Larva hostplants are Centaurea sp. and Knautia arvensis (L.) Coult. (LERAUT, 2014: 315).
Remarks: The species is a resident of the Canary Islands.
Epischnia illotella Zeller, 1839 (Fig. 5)
Material examined: Spain, Gran Canaria, Pie de la Cuesta, 500 m, 3 ♂♂, 2 ♀♀, 11-24-VI-2018, leg. P. Falck, same data but 3 ♂♂, 1 ♀, 17-30-IX-2018, leg. P. Falck, same data but 3 00, 4-23-III-2019, leg. P. Falck, DNA sample 0946PF/CILEP0945-21; El Sao, 110 m, 1 ♀, 11-24-VI-2018, leg. P. Falck; Ayacata, 1400 m, 1 ♂, 1 ♀, 17-30-IX-2018, leg. P. Falck; Teror, 500 m, 1 ♀, 24-X-13-XI-2020, leg. P. Falck; Tenerife, Arona, 500 m, 2 ♂♂, 1-20-III-2017, leg. P. Falck, genitalia slide 2694PF, same data but 1 ♂, 1 ♀, 21-V-3-VI-2019, leg. P. Falck; Las Manchas, 1050 m, 1 ♂, 1-20-III-2017, leg. P. Falck; Erjos, 1000 m, 1 ♀, leg. P. Falck, genitalia slide 2553PF; Las Mercedes, 750 m, 2 ♀♀, 21-V-3-VI-2019, leg. P. Falck; 8 km N Vilaflor, 1700 m, 2 ♂♂, 21-V-3-VI-2019, leg. P. Falck, DNA sample 0945PF/CILEP0944-21 (all PF). New to the Canary Islands.
Distribution: In southern Europe from Portugal eastwards to Russia (LERAUT, 2014: 317).
Biology: The larva hostplants are Inula montana L., Inula crithmoides L. and Asteriscus spinosus (L.) Cass. (LERAUT, 2014: 317).
Remarks: The species is a resident of the Canary Islands.
Assara conicolella (Constant, 1884) (Fig. 6)
Material examined: Spain, Tenerife, Arona, 670 m, 1 ♂, 21-V-3-VI-2019, leg. P. Falck, genitalia slides 3314PF, 3570PF, DNA sample 0617PF/CILEP0616-20; Las Manchas, 1050 m, 1 ♂, 3-16-VIII2021, leg. P. Falck (PF). New to the Canary Islands.
Distribution: The southern part of Portugal, Spain and France (LERAUT, 2014: 382).
Biology: The larva lives in shrivelled cones of Pinus halepensis Mill. (LERAUT, 2014: 382). P. halepensis is widespread in the Canary Islands at altitudes ranging from sea level to 1500 m (MUER et al., 2016: 87).
Remarks: The species is probably a resident of the Canary Islands. We obtained DNA barcode fragments of 580 bp from one specimen. The barcodes fall within Barcode Index Number (BIN) BOLD: AEE8286. The minimum intraspecific p-distance to European specimens of A. conicolella is 6.2% with Barcode Index Number (BIN): ADR6757. Despite the significant divergence in COI we consider the Canary Islands specimens to be conspecific with A. conicolella, because we did not observe any morphological differences in either the external features in or the genitalia.
Euzophera mirumella Falck, Karsholt & Slamka, sp. n. (Fig. 7)
Holotype ♂: Spain, Gran Canaria, Pie de la Cuesta, 500 m, 11-24-VI-2018, leg. P. Falck (ZMUC).
Paratypes: Spain, Gran Canaria, Pie de la Cuesta, 500 m, 24 ♂♂, 4 ♀♀, 11-24-VI-2018, leg. P. Falck, genitalia slides 3356PF, 3557PF, 3562PF; Barranquillo Andrés, 700 m, 7 ♂♂, 3 ♀♀, 11-24-VI2018, leg. P. Falck; La Gomera, Hermigua, 250 m, 1 ♂, 24-X-12-XI-2021, leg. P. Falck; La Palma, Las Toscas, 150 m, 2 ♂♂, 1 ♀, 17-23-I-2019, leg. P. Falck, DNA sample Lepid Phyl 0948PF/CILEP0947- 21; Los Cancajos, 20 m, 2 ♂♂, 17-23-I-2019, leg. P. Falck, genitalia slide 3563PF; Tenerife, Los Gigantes, 50 m, 3 ♂♂, 8-22-XI-2016, leg. P. Falck, genitalia slide 2601PF; Tamaimo, 550 m, 3 ♂♂, 4 ♀♀, 8-22-XI-2016, leg. P. Falck, genitalia slide 2603PF; Playa Paraiso, 50 m, 1 ♂, 1-20-III-2017, leg. P. Falck; Las Mercedes, 750 m, 1 ♀, 18-XI-8-XII-2018, leg. P. Falck; Arona, 670 m, 1 ♂, 21-V-3-VI2019, leg. P. Falck, DNA sample Lepid Phyl 0947PF/CILEP0946-21 (PF, MNCN).
Description: Male. Wingspan 19-30 mm. Head and neck with grey white-tipped scales. Antenna about 2/3 length of forewing, yellowish, ciliate-setose with very short ciliae; scapus whitish. Labial palps upturned, approximately 1.5 diameter of the eye, grey, mottled with light-tipped grey scales, more whitish medially. Thorax and tegulae with grey mottled scales. Forewing narrow, lanceolate with blunt apex; ground colour grey, mottled with black and white scales; ante-medial line slightly oblique, white, edged distally with black and towards dorsum reddish brown scales; discal spot indistinctly white, edged by black scales in lower half; post-medial line sinous, white, edged by black medially; marginal dots black; fringes with white-tipped grey scales. Hindwing grey; fringes grey. Abdomen greyish.
Female: Externally similar to male, but antenna filiform with short cilia, weakly ringed light grey / grey.
Male genitalia (Figs 18, 18a): Uncus robust, broad, moderately tapering towards apex with short setae; gnathos strongly sclerotized, short, pear-shaped; labides well sclerotized, apically rounded, with small depression in the middle; tegumen short; valva broad, corrugated and densely covered by long setae, costa well-sclerotized with few short setae, longer towards apex, apically with short, blunt projection; cucullus rounded; juxta well sclerotized, U-shaped, bilaterally oval enlarged; sacculus straight, broad; saccus sclerotized, short, U-shaped. Phallus short and wide, moderately tapering towards apex, apically with fine transverse sclerotization, vesica with quantity of fine spines. Culcita (modified sternum VIII) (Fig. 18b) simple, composed of a V-shaped and a N-shaped plate.
Female genitalia (Fig. 24): Papillae anales of moderate length, densely covered by long setae; posterior and anterior apophysis of the same length; segment VIII well sclerotized, square-shaped, covered by setae (mainly posteriorly); ventrally U-shaped, medially with broad vertical sclerotization, anteriorly double-armed; ostium broad, triangular and membranous; ductus bursae narrow, membranous; corpus bursae oval, membranous, posteriorly with a quantity (more than 100) of fine spines; ductus seminalis situated posteriorly.
DNA barcodes: We obtained full length DNA barcodes (658 bp) from two specimens, one from the island of La Palma and one from the island of Tenerife. The barcodes fall within Barcode Index Number (BIN) BOLD: AEN4203. The intraspecific p-distance is 2.00%. The minimum p-distance to nearest neighbour, an unidentified species from Gabon, Africa is 7.85%, with the Barcode Index Number (BIN) BOLD: ADJ4456. The minimum p-distance to E. pinguis (Haworth, 1811) is 9.68%, with the Barcode Index Number (BIN) BOLD: AAC0332.
Diagnosis: Based on the male and female genitalia this new species belongs to the genus Euzophera Zeller, 1867. Externally E. mirumella sp. n. is somewhat similar to E. pinguis (Haworth, 1811), but it is larger, its colour is light greyish (yellowish brown in E. pinguis), cross-lines on the forewings are smoother and hindwings are darker. The male genitalia are characteristic, particularly by shape of valva with an apical projection and labides and the short, pear-shaped gnathos; culcita has a different and characteristic shape. Female genitalia are characteristic by the combination of the shape of segment VIII, absence of typical signum and posteriorly situated ductus seminalis. Similar species were not found among the described species (cf. ROESLER, 1973; LERAUT, 2014; LEPIFORUM, 2008- 2022).
Biology: Early stages unknown. The specimens were attracted to light during late October to June.
Distribution: Known from La Gomera, Gran Canaria, La Palma and Tenerife, Spain, at altitudes ranging from sea level to 750 m.
Etymology: The specific epithet is derived from the Latin mirum (= surprise) and reflects the surprise discovery a rather large and characteristic unknown species, in abundance, from several of the Canary Islands.
Remarks: A high intraspecific variation in COI between the specimens from separate islands of the Canary Islands, in this case La Palma and Tenerife, is quite common (FALCK et al., 2021). It has been observed in endemic species of several families e.g. Tineidae, Scythrididae, Cosmopterigidae, Tortricidae (P. Falck, unpublished). E. mirumella sp. n. is not closely related to other Palaearctic Euzophera.
Homoeosoma stenotea Hampson, 1926 (Fig. 8)
Material examined: Spain, Lanzarote, Mojón Blanco, Orzola, 20 m, 4 ♂♂, 3 ♀♀, 21-X-10-XI2019, leg. P. Falck, genitalia slides 3264PF, 3265PF, 3266PF, DNA samples Lepid Phyl 0619PF/CILEP0618-20, 0620PF/CILEP0619-20, 0621PF/CILEP0620-20 (all PF). New to the Canary Islands.
Distribution: South Africa, Lesotho, Zimbabwe, Mozambique, Madagascar (DE PRINS & DE PRINS, 2011-2022), Morocco (AUDEOUD & ROCH, 1938: 369)
Biology: The larvae is reported to live in the flower-head of Senecio madagascariensis Poir. (Asteraceae), eating it out (EGLI, 2017). S. madagascariensis is not know from the Canary Island, and the larva probably feeds on other Senecio species there.
Remarks: The genitalia of both sexes are described in detail and figured by BALINSKY (1991: 26-27).
Dalakia moyalis Falck, Karsholt & Slamka, sp. n. (Fig. 9)

Figs 9-16.–
9.Dalakia moyalis Falck, Karsholt & Slamka, sp. n., ♀, Gran Canaria, 16 mm. 10.Peoria gigantesalis Falck, Karsholt & Slamka, sp. n., ♂, Tenerife, 20 mm. 11.Metasia (Clasperia) baezi Falck, Karsholt & Slamka, sp. n., ♂, Gran Canaria, 13.5 mm. 12.Loxostege sticticalis (Linnaeus, 1761), ♀, Gran Canaria, 21 mm. 13.Achyra nudalis (Hübner, 1796), ♂, Gran Canaria, 19 mm. 14.Cynaeda acutalis Falck, Karsholt & Slamka, sp. n., ♂, Gran Canaria, 24 mm. 15.Cynaeda acutalis Falck, Karsholt & Slamka, sp. n., ♂, Gran Canaria, 21 mm. 16.Cynaeda acutalis Falck, Karsholt & Slamka, sp. n., ♀, Gran Canaria, 23 mm.
Holotype ♂: Spain, Gran Canaria, Barranco Moya, 80 m, 8-20-VIII-2020, leg. P. Falck, genitalia slide 3525PF (ZMUC).
Paratypes: Spain, Gran Canaria, Barranco Moya, 80 m, 2 ♂♂, 1 ♀, 8-20-VIII-2020, leg. P. Falck, genitalia slide 3548PF, DNA samples Lepid Phyl 0741PF/CILEP0740-20, 0742PF/CILEP0741-20 (PF, MNCN).
Description: Male. Wingspan 14-16 mm. Frons white, light reddish brown in the middle and laterally; neck tufts light reddish brown. Antenna about 2/3 length of forewing, yellowish white, the first 1/3 pectinate the rest distinctly serrate with cilia shorter than diameter of the antenna, scapus yellowish, reddish dorsally. Labial palps long (approx. 2.5 diameter of the eye) and straight, reddish brown, more whitish medially. Forewing narrow, elongate with blunt apex; ground colour yellowish ochre, mottled with reddish scales in dorsal and apical part, more intensively reddish below the cell and in basal part; fringes light grey. Hindwing greyish; fringes light grey. Abdomen segment 1-3 orange, other segments yellowish white. Female: Externally similar to male, but antenna filiform with short cilia.
Male genitalia (Figs 19, 19a): Uncus slightly bilobed, laterally subtriangular with edges sclerotized, sparsely covered by short setae; gnathos short, narrow, well sclerotized, from lateral view anvil-shaped, dorsally broadening and covered by fine spikes, ventrally pointed; tegumen well developed and sclerotized; transtilla missing; valva at the base broad, evenly tapered towards apex, along costa covered by long setae, along dorsum shorter setae; cucullus narrowing, apically blunt, parallel with costa, near the middle a sclerotized flap, pointed spike-like apically, ending near cucullus, edge weakly dentate; juxta oval, sclerotized; saccus U-shaped, narrow and short, well sclerotized. Phallus slender and straight, apical-dorsally with long, slender and well sclerotized process, it is longitudinally ribbed, dorsally with 3 small parallel projections; vesica with fine sclerotized wrinkled structures. Culcita (modified sternum VIII) (Fig. 19b) simple, in shape of arcuate sclerotized lamella with paired lateral U-shaped sclerotization.
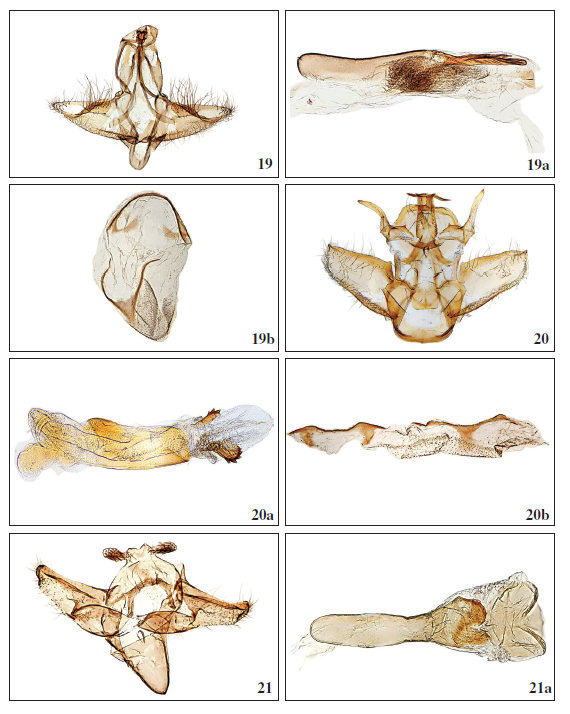
9Figs 19-21a.–
19.Dalakia moyalis Falck, Karsholt & Slamka, sp. n., ♂, Gran Canaria, GP3525PF. 19a. Phallus, Gran Canaria, GP3525PF. 19b. Segment VIII (tergit and sternit), Gran Canaria, GP3575PF. 20.Peoria gigantesalis Falck, Karsholt & Slamka, sp. n., ♂, Tenerife, GP2545PF. 20a. Phallus, Tenerife, GP2545PF. 20b. Segment VIII (tergit and sternit), Tenerife, GP3574PF. 21.Metasia (Clasperia) baezi Falck, Karsholt & Slamka, sp. n., ♂, Tenerife, GP3558PF. 21a. Phallus, Tenerife, GP3558PF.
Female genitalia (Fig. 25): Papillae anales (lateral view) strongly compressed (flattened), apically narrowing and covered by short setae; apophyses relatively short, anterior apophysis about 2/3 length of posterior apophysis; segment VIII well sclerotized; ostium area large and slightly sclerotized; ductus bursae narrow at base, broadening towards corpus bursae with fine sclerotization; ductus seminalis well developed and broad; corpus bursae oval, membranous; centrally an irregular, semioval and well sclerotized signum, covered with about 13 long and short spikes, laterally a smaller, irregular plate-like, sclerotized signum is present.
DNA barcodes: We obtained full length DNA barcodes (658 bp) from two specimens. The barcodes fall within Barcode Index Number (BIN) BOLD: AEH1203. The maximum intraspecific distance is 0.92%. The p-distance to nearest neighbour is 4.17%, with the Barcode Index Number (BIN) BOLD: ACH1411, probably an undescribed species from Pakistan.
Diagnosis: Externally, D. moyalis resembles some probably undescribed species from Pakistan and Australia, but the shape of the male genitalia are unique.
Biology: Early stages unknown. The specimens were attracted to light.
Distribution: Only known from the type-locality Barranco Moya in the northern part of the island of Gran Canaria, Spain.
Etymology: The species is named after the town Moya, situated near the type-locality.
Remarks: The placement of this new species in the genus Dalakia Amsel, 1961 is provisional. It is based on morphological similarity of male genitalia with the type species of that genus, Dalakia uniformella Amsel, 1961.
The Anerastiini is in need of revision and there is no published overview of its genera and species. Externally D. moyalis is similar to species of Hypsotropa Zeller, 1848 or Peoria Ragonot, 1887, but the forewings are more pointed, and the venation on the hindwing shows an abnormal (for Anerastiini) character with a bifid group (veins M2+M3 and Cu1 are fused and Cu2 is separate), which is shared with the tribus Cabniini (cf. SLAMKA, 2019: 10, fig. 1c). Normally, the venation of the hindwings in Anerastiini is characterized by a trifid group (veins M2+M3 are fused and Cu1 and Cu2 are separate) or rarely a quadrifid group (all 4 veins are separate) (cf. SHAFFER, 1968). Although the genus Dalakia shows characters of the trifid group, due to similarity of genitalia we place the new species moyalis into genus Dalakia.
Peoria gigantesalis Falck, Karsholt & Slamka, sp. n. (Fig. 10)
Holotype ♂: Spain, Tenerife, Los Gigantes, 50 m, 8-22-XI-2016, leg. P. Falck (ZMUC).
Paratypes: Spain, Tenerife, Los Gigantes, 50 m, 7 ♂♂, 1 ♀, 8-22-XI-2016, leg. P. Falck, genitalia slides 2545PF, 2547PF, 3574PF, DNA sample Lepid Phyl 0949PF/0948-21; Tamaimo, 550 m, 1 ♂, 1 ♀, 8-22-XI-2016, leg. P. Falck, genitalia slide 1958FS, DNA sample Lepid Phyl 0950PF/0949-21 (PF, MNCN).
Description: Male. Wingspan 14.5-20 mm. Frons straw-yellow with long thin scales, vertex strawyellow, neck tufts straw-yellow with admixture of brownish scales laterally. Antenna about 2/3 length of forewing, ciliate-setose, above whitish and beneath yellowish. Labial palps long (approx. 1.5 diameter of the eye) and straight with light brown and straw-yellow scales, more whitish medially. Thorax and tegulae light yellowish, more brownish laterally. Forewing narrow, elongate, apex blunt; ground colour white with fine straw yellow longitudinal streaks, edged with brown scales, in the middle and towards apex the admixture with brown scales in the streaks are more intense; fringes greyish. Hindwing greyish; fringes light grey. Abdomen greyish.
Female: Externally similar to male, but antenna filiform with short cilia.
Male genitalia (Figs 20, 20a): Medial process of uncus with strong sclerotized T-shape projection in the middle; on the lateral parts of uncus a long, sclerotized spicate projection, at the base of the projection a minute well-sclerotized spike anteriorly; gnathos broad and strongly sclerotized, edge densely finely serrate, narrowing towards apex, apicolaterally with two spikes; tegumen short; valva short and broad, costa well sclerotized, sparsely covered with long setae, apex pointed and sclerotized; sacculus sclerotized, densely covered with short setae; juxta square, at its bottom and bilaterally more sclerotized, in the middle (inferiorly) with a triangular sclerotization; saccus short, U-shaped, strongly sclerotized. Phallus of a medium length, well-sclerotized, slightly bent; in the everted vesica two strongly sclerotized small oval plates, dorsally with 3-5 strong spikes, between them a cluster of fine cornuti. Culcita (modified sternum VIII) (Fig. 20b) simple, being a well sclerotized lamella with a paired lateral scleriotization.
Female genitalia (Figs 26, 26a): Papillae anales long and relatively slender (lateral view), setose. Apophyses relatively short, anterior apophysis about 2/3 the length of posterior apophysis. Segment VIII well sclerotized, ventrally in the middle with deep U-shaped notch and bilaterally with a blunt protuberance; ostium short, broad, membranous; colliculum a well-sclerotized short ring; ductus bursae very short, merging with corpus bursae (modified ductus), laterally with a sclerotized, irregular and semioval plate of variable shape with 15-28 small thorns of various length: corpus bursae membranous without any structures on the surface.

Figs 26-28.–
26.Peoria gigantesalis Falck, Karsholt & Slamka, sp. n., ♀, Tenerife, GP2547PF. 26a.Peoria gigantesalis Falck, Karsholt & Slamka, sp. n., ♀ (segment VIII unfolded), Tenerife, GP1958FS. 27.Metasia (Clasperia) baezi Falck, Karsholt & Slamka, sp. n., ♀, Gran Canaria, GP3573PF. 28.Cynaeda acutalis Falck, Karsholt & Slamka, sp. n., ♀, Gran Canaria, GP3461PF.
DNA barcodes: We obtained full length DNA barcodes (658 bp) from two specimens. The barcodes fall within Barcode Index Number (BIN) BOLD: AEN7864. The maximum intraspecific distance is 0.00%. The p-distance to nearest neighbour 4.81%, with the Barcode Index Number (BIN) BOLD: ADZ3170, probably an undescribed species from South Africa.
Diagnosis: Externally P. gigantesalis does not resemble any known species from Europe or Africa. The male and female genitalia are characteristic and unmistakable compared with the other European Anerastiini (cf. LERAUT, 2014).
Biology: Early stages unknown. The specimens were attracted to light.
Distribution: Only known from two localities in the north-western part of the island of Tenerife, Spain.
Etymology: The species is named after the town Los Gigantes, situated at north-west coast of the island of Tenerife, where most of the specimens were collected.
Remarks: This new species, according the morphological characters especially of the genitalia, belongs to the genus Peoria. In the male genitalia it is characteristic that the uncus consists of two, often bifid arms, which protrude along the tegumen; gnathos ends in a small median arm, the valva is simple, with or without costal tooth and saccus is broad. In the female genitalia the elongate papillae anales with oblique profile and a short weakly sclerotized ductus bursae are characteristic (LERAUT, 2014: 95).
P. gigantesalis sp. n. belongs, together with the previous species, Dalakia moyalis sp. n. in the tribe Anerastiini. We follow HORAK (2003: 89) in considering Peroiini as a synonym of Anerastiini.
CRAMBIDAE
SPILOMELINAE
Metasia (Clasperia) baezi Falck, Karsholt & Slamka, sp. n. (Fig. 11)
Holotype ♂: Spain, Gran Canaria, Pie de la Cuesta, 500 m, 11-24-VI-2018, leg. P. Falck (ZMUC).
Paratypes: Spain, Gran Canaria, 1.3 km N Mogan, 430 m, 1 ♂, 2-8-XI-2014, leg. B. Skule (ZMUC); Barranquillo Andrés, 700 m, 2 ♂♂, 1 ♀, 11-24-VI-2018, leg. P. Falck, genitalia slide 2734PF; Puerto Rico, 50 m, 5 ♂♂, 11-24-VI-2018, leg. P. Falck, same data but 8 ♂♂, 2 ♀♀, 17-30-IX2018, leg. P. Falck, genitalia slide 3572PF; Pie de la Cuesta, 500 m, 5 ♂♂, 1 ♀, 17-30-IX-2018, leg. P. Falck, same data but 1 ♂, 24-X-13-XI-2020, leg. P. Falck; El Sao, 110 m, 2 ♂♂, 1 ♀, 17-30-IX-2018, leg. P. Falck; Fataga, 500 m, 3 ♂♂, 1 ♀, 21-VIII-4-IX-2020, leg. P. Falck; Barranco Moya, 80 m, 1 ♂, 8-20-VIII-2020, leg. P. Falck; Barranco Guayadeque, 700 m, 2 ♂♂, 2 ♀♀, 21-VIII-4-IX-2020, leg. P. Falck, genitalia slide 3573PF; Barranco de Azuaje, 270 m, 1 ♂, 8-20-VIII-2020, leg. P. Falck; Ayacata, 1400 m, 1 ♂, 21-VIII-4-IX-2020, leg. P. Falck; Agaeta, 180 m, 3 ♂♂, 24-X-13-XI-2020, leg. P. Falck; Playa del Cura, 15 m, 1 ♂, 9-22-VI-2021, leg. P. Falck; Tenerife, Barranco Badajoz, 2 ♂♂, 1 ♀, 24-VI-1995, leg. M. Báez, genitalia slide 5099OK (MB, ZMUC); El Medano, 10 m, 1 ♂, 18-XI-8- XII-2018, leg. P. Falck, same data but 4 ♂♂, 1 ♀, 21-V-3-VI-2019, leg. P. Falck; Arona, 670 m, 4 ♂♂, 3-16-VIII-2021, leg. P. Falck, genitalia slide 3555PF; Las Manchas, 1050 m, 1 ♂, 3-16-VIII-2021, leg. P. Falck, genitalia slide 3558PF; Güimar, 500 m, 2 ♂♂, 3-16-VIII-2021, leg. P. Falck (PF, MNCN).
Description: Male. Head grey-brown mottled with yellowish scales. Neck yellowish, mottled with a few brownish scales. Antenna about 2/3 length of forewing, brown, filiform, with short cilia (approximately fl diameter of antenna). Labial palps length approximately 1.5 of eye diameter, straight, brown, dorsally and medially mottled with yellowish scales, segment 3 very short. Thorax and tegulae brown. Forewing relatively long and narrow, apex rounded and termen oblique; ground colour brown; antemedial line and post-medial line diffuse dark brown, not always visible; antemedial spot diffuse, small, dark brown; discal spot dark brown, medially a small diffuse yellowish spot; fringes greyish brown. Hindwing greyish brown with concolorous fringes. Abdomen greyish.
Female: Externally similar to male, but antenna with shorter ciliae.
Male genitalia (Figs 21, 21a): Uncus bilobed, lobes short, apically densely covered by short spines; tuba analis short, membranous; gnathos absent, tegumen short, weakly sclerotized; transtilla well developed, slightly sclerotized with long paired projections towards juxta; valva relatively wide (sparsely covered by setae), tapering towards apex, cucullus ± rounded, apically and dorsally with slight groove, a heavily sclerotized ridge parallel to costa not reaching costa; juxta rounded, well sclerotized, apico-bilaterally with arcuate folds or grooves; saccus well developed, long, strongly sclerotized, subtriangular. Phallus long and slender, apically with a sclerotized, often bilobed disc (plate) with or without minute teeth on the edge; cornuti absent.
Female genitalia (Fig. 27): Papillae anales short and quite broad, densely covered by ± short setae; posterior and anterior apophysis of the same length, robust and relatively long; sternum VIII well sclerotized, U-shaped; ostium bursae broad and finely sclerotized, ductus bursae broad, membranous, corpus bursae oval, membranous, without any spines; signum oval, weakly sclerotized with several very short spines, placed at the bottom of corpus bursae.
Diagnosis: The adult of Metasia (Clasperia) baezi is unique because of the shape of the almost uniform brown (except for the diffuse spots) forewing. In the male genitalia the sclerotized ridge parallel to costa and the sclerotized U-shaped fold (instead of clasper) are characteristic. In the female genitalia the long apophysis and the membraneous corpus bursae without spines are characteristic.
Biology: Early stages unknown. The specimens were attracted to light from June until the beginning of December.
Distribution: M. baezi is known from the islands of Gran Canaria and Tenerife at altitudes ranging from sea level to 1400 m. The species is probably an endemic species of the Canary Islands.
Etymology: The species name is dedicated to Marcos Báez of La Laguna, Tenerife who collected the first known specimens used for our study.
Remarks: According to external features it is similar to subgenus Clasperia Hartig, 1952 and Aristidea Slamka, 2013, but in the male genitalia is developed a ± sclerotized fold in shape of “U” or “V”, which continues dorsally towards sacculus, instead of short clasper (harpe) (typical for subgenus Clasperia). In the female genitalia the oval, membranous corpus bursae without any spines is a typical character for other species of the subgenus Clasperia.
PYRAUSTINAE
Loxostege sticticalis (Linnaeus, 1761) (Fig. 12)
Material examined: SPAIN, Gran Canaria, Barranco de Azuaje, 270 m, 2 ♂♂, 1 ♀, 8-20-VIII-2020, leg. P. Falck; Carretería, 455 m, 1 ♀, 8-20-VIII-2020, leg. P. Falck (all PF). New to the Canary Islands.
Distribution: L. sticticalis is a well-known migrant and it is widely distributed throughout the Palaearctic and Nearctic regions (SLAMKA, 2013: 26).
Biology: The larva lives on various herbaceous plants, such as Artemisia sp., Beta sp., Chenopodium sp., Trifolium sp., etc. (SLAMKA, 2013: 26).
Remarks: Probably a migrant to the Canary Islands.
Achyra nudalis (Hübner, 1796) (Fig. 13)
Material examined: Spain, Gran Canaria, Pie de la Cuesta, 500 m, 1 ♀, 17-30-IX-2018, leg. P. Falck, same data but 3 ♂♂, 2 ♀♀, 21-VIII-4-IX-2020, leg. P. Falck; El Sao, 110 m, 1 ♂, 11-24-VI-2018, leg. P. Falck, same data but 3 ♂♂, 1 ♀, 21-VIII-4-IX-2020, leg. P. Falck; Puerto Rico, 50 m, 2 ♂♂, 17-30-IX-2018, leg. P. Falck; Ayacata, 1400 m, 1 ♂, 1 ♀, 21-VIII-4-IX-2020, leg. P. Falck; Barranco Moya, 80 m, 1 ♂, 8-20-VIII-2020, leg. P. Falck (all PF). New to the Canary Islands.
Distribution: Southern Europe, Africa, Middle East, Turkey, Mongolia, Transcaucasia and India.
Biology: The larva feeds on leaves of Champhorosma sp., Echium sp., Chenopodium sp. and Amaranthus sp. (SLAMKA, 2013: 28).
Remarks: The species is a resident in the Canary Islands.
ODONTIINAE
Cynaeda acutalis Falck, Karsholt & Slamka, sp. n. (Figs 14, 15, 16)
Holotype ♂: Spain, Gran Canaria, Ayacata, 1400 m, 17-30-IX-2018, leg. P. Falck, genitalia slide 2810PF, DNA sample Lepid Phyl 0093PF/CILEP0093-19 (ZMUC).
Paratypes: Spain, Gran Canaria, Ayacata, 1400 m, 2 ♂♂, 2 ♀♀, 17-30-IX-2018, leg. P. Falck, genitalia slide 2806PF, DNA samples Lepid Phyl 0092PF/CILEP0092-19, 0094PF/CILEP0094-19, same data but 28 ♂♂, 2 ♀♀, 21-VIII-4-IX-2020, leg. P. Falck, genitalia slides 3461PF, 3577PF, 3578PF, DNA samples Lepid Phyl 0752PF/CILEP0751-20, 0753PF/CILEP0752-20, same data but 2 ♂♂, 1 ♀, 9-22-VI-2021, leg. P. Falck; Barranco de Guayadeque, 700 m, 5 ♂♂, 21-VIII-4-IX-2020, leg. P. Falck, DNA sample Lepid Phyl 0754PF/CILEP0753-20; 4 km N Pie de la Cuesta, 700 m, 1 1, 21-VIII-4-IX2020, leg. P. Falck (PF, MNCN).
Description: Male. Wingspan 17-25 mm. Head and neck white, mottled with light yellowish brown towards middle. Antenna about 2/3 length of forewing, white, with long cilia (approximately 1.5 diameter of antenna). Labial palps straight (approximately 1.5 diameter of the eye) light yellowish brown laterally and white medially. Thorax and tegulae light yellowish brown. Forewing triangular with pointed apex, ground colour light yellowish brown, whitish medially to the postmedial line; at costa about 1/9 a diffuse brownish oblique antemedial line with a blackish scale tuft at dorsum; postmedial line zigzagging, bordered by blackish brown scales, sometimes forming two irregular dark brown spots distally, and interrupted by a white area distally to the cell; subterminal area white; fringe chequered blackish brown and white. Hindwing whitish often with an admixture of dark grey scales distally to the zigzagging postmedial line; fringe white.
Female: Externally similar to male, but antenna filiform with shorter ciliae.
Variation: The species can vary from specimens almost without blackish brown admixture of wings (Fig. 15) to very contrasting specimens (Fig. 16). It is also highly variable in size, a phenomenon seen in many species of Lepidoptera from the Canary Islands, probably depending on access to fresh food-plants.
Male genitalia (Figs 22, 22a): Uncus rounded, in shape of two paired arcuate lobes, densely haired (except of apical parts); gnathos strongly sclerotized, triangle-shaped, apex ± pointed; tegumen short; valva rather broad, cucullus rounded, densely haired, costa well sclerotized, short, moderately arcuate; sacculus moderately convex, weakly haired; vinculum strongly sclerotized; saccus short, triangular with rounded tip, well sclerotized; juxta A-shaped towards saccus, long (as long as phallus), narrow and moderately sclerotized. Phallus slender, moderately bent, without cornutus.
Female genitalia (Fig. 28): Papillae anales short and broad, sparsely setose; posterior apophysis slightly shorter than anterior apophysis; segment VIII narrow, well sclerotized and sparsely covered by short setae (mainly posteriorly), anteriorly with broad square sclerotized projection; antrum narrow, membranous, colliculum well sclerotized, short and narrow; ductus bursae long, membranous, broadening towards corpus bursae; corpus bursa oval, membranous, without signum.
DNA barcodes: We obtained full length DNA barcodes (658 bp) from six specimens. The barcodes fall within Barcode Index Number (BIN) BOLD: ADV8645. The maximum intraspecific pdistance is 0.32% (mean 0.14%, n=6). The minimum p-distance to nearest neighbour Cynaeda dentalis ([Denis & Schiffermüller], 1775) is 3.09%, with the Barcode Index Number (BIN) BOLD: AAE7870. We obtained full length barcodes (658 bp) from three specimens of C. dentalis from the Canary Islands to compare their DNA with continental European specimens. The results corresponded 100%.
Diagnosis: C. acutalis resembles especially C. dentalis, which is also established in the Canary Islands, and C. gigantea Staudinger, 1880. It can be distinguished from C. dentalis by the pointed apex of the forewing, the dorsal part of the postmedial line is less zigzagging and often diffuse. Specimens with dark brown areas distally to the postmedial line are very characteristic. From C. gigantea it can be distinguished by the blurred wing pattern and by the lack of brownish stripes in the marginal area (between the veins). In the male genitalia C. acutalis sp. n. is similar to C. dentalis. It can be distinguished by the less broad valva, evenly curved costa (in C. dentalis costa is bent almost 90 medially), and the medial edges of the two uncus lobes are curved beneath apex. Both species lack cornutus in the phallus. From C. gigantea and C. forsteri de Lattin, 1951 it can be distinguished by the phallus without cornutus; in the phallus of C. forsteri is one small pointed cornutus, in C. gigantea two pointed cornuti. In the female genitalia C. acutalis can be distinguished by the lack of signum; in C. dentalis and C. gigantea a long narrow, serrated signum is present in corpus bursae. The female of C. forsteri is unknown.
Biology: Early stages and host-plant are unknown, but the larva probably feeds on an Echium species. There are several endemic Echium species in the Canary Islands (MUER et al., 2016: 741- 761). The specimens were attracted to light from June until the end of September.
Distribution: Known only from scattered localities in central part of Gran Canaria at altitudes between 700 and 1400 m. In one locality (4 km N Pie de la Cuesta) C. acutalis occurred sympatric with C. dentalis.
Etymology: The species is named after the pointed apex of the forewing, from the Latin acutus (= acute).
Discussion
The Pyraloidea is an important and diverse group of Lepidoptera in the Canary Islands. More than 150 species of Pyraloidea are currently known from the islands, yet in our previous paper (FALCK et al., 2019) we were able to increase the number of known species there by 15%. In the present paper we add another 14 additional species of this superfamily to the fauna of the Canary Islands.
Most of these species are members of the native fauna of the islands. The six newly described species can all appear to be endemic as they are neither known nor reasonably expected from elsewhere. Several species are shared with Europe and North Africa and one, Homoesoma stenotea, has an entirely African distribution. Only one species, Loxostege sticticalis is known to be migratory, and none of the species reported here can be traced back to importation from trade.
The Pyraloidea is a megadiverse superfamily of Lepidoptera, which is far from being revised at the global level. This is especially true for the subfamily Phycitinae which, according to LERAUT (2021a: 1, 5) includes 3452 described species in 641 genera. Nobody has a full overview of this subfamily, and several genera that contain species of economic importance have not yet been revised. We are well aware of these descriptions of new taxa should preferably, be made within taxonomic revisions, but it is also evident that such revisions will not be available for most groups of Pyraloidea within a foreseeable future and also not before many of the species have become extinct. With the declining interest from governments to support institutions dealing with insect taxonomy, and the problems with obtaining type material on loan from some of these institutions, it is becoming increasingly difficult to prepare revisions of diverse groups of insects. We have made serious efforts to check if the new species described here were already named and we feel confident that describing them, accompanied by illustrations of good quality and information on the DNA barcode, will make it possible for other researchers to recognise them.
Acknowledgments
We are grateful to Dr. Marcos Báez, La Laguna, Tenerife, Spain and to Bjarne Skule, Veksø, Denmark for loan and donation of specimens. We are moreover grateful to Dr. Antonio Vives, Madrid, Spain for translating the abstract into Spanish, for editing our manuscript, and for his kind help with obtaining permission to collect Lepidoptera in the Canary Islands into the Scientific Project of SHILAP. Colin W. Plant, Bishops Stortford, England, kindly corrected the English language and commented on the manuscript.
BIBLIOGRAPHY
AUDEOUD, G. E. & ROCH, M., 1938.– Chasses printanières aux Lépidoptères au Maroc.– Mitteilungen der Schweizerischen Entomologischen Gesellschaft, 17: 354-372.
BALINSKY, B. I., 1991.– On genitalia of some southern African Phycitinae (Lepidoptera: Phycitinae).– South African Journal of Zoology, 26: 11-35.
DE PRINS, J. & DE PRINS, W., 2011-2022.– Afromoths, online database of Afrotropical moth species (Lepidoptera). Available from http://www.afromoths.net/ (accessed 17 January 2022).
EGLI, D., 2017.– Native range studies on insect herbivores associated with fireweed (Senecio madagascariensis) in KwaZulu-Natal, South Africa, with prospects for biological control in invaded countries: 155 pp. PhD thesis, University of KwaZulu-Natal, Pietermaritzburg.
FALCK, P. & KARSHOLT, O., 2019.– New data on Praydidae, Oecophoridae, Stathmopodidae and Cosmopterigidae from the Canary Islands, Spain (Insecta: Lepidoptera).– SHILAP Revista de lepidopterología, 47(186): 325-340.
FALCK, P., KARSHOLT, O. & SLAMKA, F., 2019.– New data on Pyraloidea from the Canary Islands, Spain (Lepidoptera).– SHILAP Revista de lepidopterología, 47(185): 33-48.
FALCK, P., KARSHOLT, O. & SIMONSEN, T. J., 2021.– The genus Apatema Walsingham, 1900 in the Canary Islands and Madeira, with description of 13 new species (Lepidoptera: Autostichidae, Oegoconiinae).– SHILAP Revista de lepidopterología, 49(194): 273-318.
HORAK, M., 2003.– Reassesment of the Anerastiini and their status in the Phycitinae (Pyralidae): a century-long controversy.– Invertebrate Systematics, 17: 89-98.
LEPIFORUM, 2008-2022.– E. RENNWALD & J. RODELAND et al. (ed.): Lepiforum: Bestimmungshilfe für die in Europa nachgewiesenen Schmetterlingsarten. Lepiforum e.V. Available from http://www.lepiforum.de/lepiwiki.pl?Schmetterlingsfamilien_Europa (accessed 17 January 2022).
LERAUT, G. H. C., 2021a.– Species général des Phycitinae (Lep.: Pyraloidea, Pyralidae).– Revue Française d’Entomologie Générale, 2 (Supplement, 5-6): 1-500.
LERAUT, G. [H. C.], 2021b.– Short Note - A Phycitinae species new to the Algerian Sahara (Lepidoptera : Pyralidae).– Revue Française d’Entomologie Générale, Autun, 3(4): 88-92.
LERAUT, P., 2014.– Pyralids 2. Moths of Europe, 4: 440 pp. N. A. P. editions. Verrieres le Buisson.
MUER, T., SAUERBIER, H. & CABRERA CALIXTO, F., 2016.– Die Farn- und Blütenpflanzen der Kanarischen Inseln: 1310 pp. Margraf Publishers GmbH, Frankfurt am Main.
RATNASINGHAM, S. & HEBERT, P. D. N., 2013.– A DNA-based registry for all animal species: The Barcode Index Number (BIN) System.– PLOS ONE, 8(8): e66213. doi:10.1371/journal.pone.0066213.
ROBINSON, G. S., 1976.– The preparation of slides of Lepidoptera genitalia with special reference to the Microlepidoptera.– Entomologist’s Gazette, 27: 127-132.
SHAFFER, J. C., 1968.– A revision of the Peoriinae and Anerastiinae (auctorum) of America north of Mexico (Lepidoptera: Pyralidae).– Bulletin of the United States National Museum, Washington, D.C., 280: 1-124.
SLAMKA, F., 2006.– Pyralinae, Gallerinae, Epipaschiinae, Cathariinae, Odontiinae. Identification - Distribution - Habitat - Biologie.– Pyraloidea (Lepidoptera) of Europe, 1: 139 pp. Frantisek Slamka, Bratislava.
SLAMKA, F., 2013.– Pyraustinae & Spilomelinae. Identification - Distribution - Habitat - Biologie.– Pyraloidea (Lepidoptera) of Europe, 3: 357 pp. Frantisek Slamka, Bratislava.
SLAMKA, F., 2019.– Phycitinae - Part 1. Identification - Distribution - Habitat - Biologie.– Pyraloidea (Lepidoptera) of Europe, 4: 432 pp. Frantisek Slamka, Bratislava.
VIVES MORENO, A., 2014.– Catálogo sistemático y sinonímico de los Lepidoptera de la Península Ibérica, de Ceuta, de Melilla y de las islas Azores, Baleares, Canarias, Madeira y Salvajes (Insecta: Lepidoptera): 1184 pp. Suplemento de SHILAP Revista de lepidopterología, Improitalia, Madrid.
Notas de autor
per.falck@live.dk