Abstract:
The subtribe Pronophilina Reuter, 1896 is a group of Papilionoidea that is particularly useful for assessing biodiversity and human-made disturbance, especially in cloud forests. This is because of their high diversity, abundance, distribution patterns, ecological fidelity, and usefulness in formulating conservation policies. A rapid assessment (RAP) was conducted on the Pronophilina in two locations in the upper montane area of Manu National Park, southeastern Peru. The assessment included an evaluation of species richness, faunal composition, similarity, and community structure of Lepidoptera in the area. The two locations were only 30 km apart but in different elevations, ranging from 2800 to 3200 m above sea level. The study recorded a total of 1941 specimens of 45 species, with 18 species (40%) belonging to the genus Pedaliodes Butler, 1867, and as many as 1356 individuals (70%) were found in this genus. Some local species were found to be restricted to very narrow elevation ranges at the forest-puna grassland ecotone. The community structure showed good adjustment with the RAD log-normal model in Trocha Unión and with the Zipf-Mandelbrot model in Qurqurpampa, indicating that the sample obtained is representative of well-preserved uppermost cloud forests. The Sørensen similarity index was relatively low, despite the short distance between the two sites and no apparent geographic isolation. Sampling by “Van Someren-Rydon” traps was confirmed as a very effective method for studying cloud forest species, Satyrinae in particular, to evaluate community structure, diversity, and altitudinal distribution.
Keywords: Lepidoptera, Nymphalidae, Satyrinae, conservation policies, forest disturbance, RBA, RAD, Peru.
Resumen:
La subtribu Pronophilina Reuter, 1896 es un grupo de Papilionoidea particularmente útil para evaluar la biodiversidad y las perturbaciones causadas por el hombre, especialmente en los bosques nubosos. Esto se debe a su gran diversidad, abundancia, patrones de distribución, fidelidad ecológica y utilidad para formular políticas de conservación. Se llevó a cabo una evaluación rápida (RAP) de la Pronophilina en dos localidades de la zona montañosa superior del Parque Nacional del Manu, al sureste de Perú. La evaluación incluyó una valoración de la riqueza de especies, la composición faunística, la similitud y la estructura comunitaria de los Lepidoptera de la zona. Las dos localidades estaban separadas por sólo 30 km pero en elevaciones diferentes, entre 2.800 y 3.200 m sobre el nivel del mar. El estudio registró un total de 1941 ejemplares de 45 especies, de las que 18 especies (40%) pertenecían al género Pedaliodes Butler, 1867, y se encontraron hasta 1356 individuos (70%) de este género. Algunas especies locales se encontraron restringidas a rangos de elevación muy estrechos en el ecotono bosquepradera de puna. La estructura de la comunidad mostró un buen ajuste con el modelo log-normal RAD en Trocha Unión y con el modelo Zipf-Mandelbrot en Qurqurpampa, lo que indica que la muestra obtenida es representativa de bosques nubosos superiores bien conservados. El índice de similitud de van Sørensen fue relativamente bajo, a pesar de la corta distancia entre los dos lugares y de no existir un aislamiento geográfico aparente. El muestreo mediante trampas “Van Someren-Rydon” se confirmó como un método muy eficaz para estudiar las especies de los bosques nubosos, Satyrinae en particular, para evaluar la estructura de la comunidad, la diversidad y la distribución altitudinal.
Palabras clave: Lepidoptera, Nymphalidae, Satyrinae, políticas de conservación, perturbación forestal, RBA, RAD, Perú.
Carátula del artículo
A Rapid Assessment of Pronophilina Reuter, 1896 community structure and diversity in the uppermost cloud forests near timberline in Manu National Park, Peru (Lepidoptera: Nymphalidae, Satyrinae)
Una evaluación rápida de la estructura y diversidad de la comunidad de Pronophilina Reuter, 1896 en los bosques nublados más altos cerca del límite maderero en el Parque Nacional del Manu, Perú (Lepidoptera: Nymphalidae, Satyrinae)
 José Cerdeña1 cerdenajoseal@yahoo.es
José Cerdeña1 cerdenajoseal@yahoo.es
Universidad Nacional de San Agustín de Arequipa, Perú
Oscar Mahecha-J.2 oscarmahecha23@gmail.com
Jagiellonian University, Polonia
Universidad Distrital Francisco José de Caldas, Colombia
Rafal Garlacz3 rafal.garlacz@uj.edu.pl
Jagiellonian University, Polonia
Jackie Farfán4 jjackie4u@gmail.com
Universidad Nacional de San Agustín de Arequipa, Perú
José Arivilca-Pari5 joalarp@gmail.com
Universidad Nacional de San Agustín de Arequipa, Perú
Klaudia Florczyk6 klaudia.florczyk@uj.edu.pl
Jagiellonian University, Polonia
Marco Delgado-Coila7 markro321@gmail.com
Universidad Nacional de San Agustín de Arequipa, Perú
Tomasz W. Pyrcz8 tomasz.pyrcz@uj.edu.pl
Jagiellonian University, Polonia
Jagiellonian University, Polonia
Vanessa Díaz-S.9 diazsvane@gmail.com
Jagiellonian University, Polonia
Universidad Distrital Francisco José de Caldas, Colombia
Shilap Revista de Lepidopterología, vol. 52, no. 206, pp. 235-250, 2024
Sociedad Hispano-Luso-Americana de Lepidopterología
Received: 10 June 2022
Accepted: 29 January 2024
Published: 30 June 2024
Introduction
Tropical regions around the world are known for their high biodiversity and endemism of flora and fauna (Hamer & Hill 2004; Lodh & Agarwala, 2016; Meseguer et al. 2022). However, they are under threat of species extinction due to deforestation and habitat loss caused by increasing human demands (Lodh & Agarwala, 2016; Moreira et al. 2023). Among these regions, the Andean tropical montane forest (ATMF) is considered the principal hotspot of global biodiversity, and it has extremely high species richness in most taxonomic groups of plants and animals and contains sites with high speciation rates (Pennigton & Lavin, 2010; Toledo-Aceves et al. 2011; Ledo et al. 2012; Horwath et al. 2019; Bax et al. 2021; Meseguer et al. 2022), and it is also home to many endemic species with restricted geographic ranges (Luebert & Weigend, 2014; Hutter et al. 2017; Pérez-Escobar et al. 2017; Meseguer et al. 2022). However, the ATMF is among the most threatened ecosystems on the planet, with a speedy rate of loss of its natural vegetation cover due to human land-use change, such as agricultural expansion, urbanization, and deforestation (Armenteras et al. 2011; Toledo-Aceves et al. 2011; Quitián et al. 2018; Aide et al. 2019; Horwath et al. 2019; Moreira et al. 2023). This has put many endemic species at risk of extinction due to the bottleneck process in their populations (ToledoAceves et al. 2011; Mahecha-Jiménez et al. 2011; Soh et al. 2019; Bax et al. 2021). Moreover, due to their restricted elevation range, many ATMF plant and animal species have naturally limited distribution in the cloud belt of tropical mountains, ATMF distribution is often considered analogous to an archipelago or islands, with deforestation enhancing the isolation of remaining ATMF fragments (Toledo-Aceves et al. 2011; Mahecha et al. 2019).
Over the last two decades, there has been a gradual loss of tropical forests in Latin America, specifically the ATMF, that have been cleared and replaced with pastures and crops, leading to a permanent loss of forest cover, and due to their high population density, the ATMF has been subjected to intense agricultural activity, which has led to their reduction (Castellanos-Mora & Agudelo-Hz, 2020; Moreira et al. 2023). Extensive areas of cloud forests are being cut down for timber, cleared for agriculture at mid-elevations, and burnt for grazing at high elevations and the act of burning is likely the primary reason behind the widespread and dramatic disappearance of vast portions of lowland and montane tropical forests in South America for centuries (Arroyo-Kalin, 2012; Aide et al. 2019). It is increasingly clear that the impact of human activity in the tropical Andes has been intense and longlasting, with evidence dating back to the pre-Columbian era, and this activity has greatly contributed to the deforestation of the region, particularly at high elevations near timberline, and on a larger scale than what we see today (Loughlin et al. 2018). For the above reasons, ATMF is widely regarded as a conservation priority worldwide because provide important ecosystem services to millions of people (Ivory & Misrachi, 2014; Moreira et al. 2023) due to its critical role in the maintenance of hydrological and nutrient cycles, carbon sequestration and storage, and prevention of erosion and its high biodiversity (Toledo-Aceves et al. 2011; Soh et al. 2019; Bax et al. 2021). For that reason, there is an immediate need for conservation in highly diverse tropical ecosystems, especially in remote areas like mountain habitats. However, the lack of data has been a barrier to effective planning and implementation of conservation efforts (Lodh & Agarwala, 2016).
Conservation International introduced the Rapid Assessment Program (RAP) in 1990 for speciesrich areas, to create baseline data which includes species checklists, records of habitat structure, and of disturbance and anthropogenic pressures in a short time, and vegetation type, etc. (Alonso et al. 2011; Lodh & Agarwala, 2016). To address this, Rapid Biodiversity Assessment (RBA) is a cost- and timeeffective method for exploring the biodiversity of tropical habitats and it is considered a good method for predicting species richness using indicator taxa (Pearson, 1994; Lodh & Agarwala, 2016; Oliveira et al., 2020; Tettey et al. 2020). In addition, the RBAs only use a taxonomic group to indicate the site’s status (Tettey et al. 2020). Butterflies are commonly used to assess the health of both pristine and anthropogenic ecosystems. They have also been utilized to study various aspects of tropical forest ecology in natural, managed, and degraded ecosystems (Tettey et al. 2020). Moreover, butterflies are a promising group of insects for rapid assessment because they are widespread, conspicuous, and easily recognizable, taxonomically well-known, and effective indicators of forest health (Lodh & Agarwala, 2016; Henao-Bañol & Gantiva-Q., 2020; Tettey et al. 2020; Sharma & Sharma, 2021). Also, Lepidoptera diversity is an indicator of biodiversity because they depend on plant diversity and different habitats according to ecosystem ecology (Ferrer-Paris et al. 2013; Lodh & Agarwala, 2016; Tettey et al. 2020; Sharma & Sharma, 2021).
The subtribe Pronophilina Reuter, 1896 is one of the groups of Lepidoptera almost entirely restricted to ATMF region (Casner & Pyrcz, 2010; Mahecha et al. 2019; Pyrcz et al. 2019). Thanks to numerous contributions, over the last two decades, in all aspects of their taxonomy, phylogeny and natural history it turned from one of the least known into one of the better known, if not the best-known group of Andean Lepidoptera, and they present a number of characteristics which make them a group particularly suitable for researching biodiversity (Pyrcz et al. 2009; Casner & Pyrcz, 2010; Pyrcz et al. 2019; Díaz-Suárez et al. 2022). In particular, Pronophilina are one of the most abundant and arguably the most species-rich group among AMTF with well over 600 known species (Lamas et al. 2004; Pyrcz, 2010). They are highly habitat-specific, restricted to well-defined ranges of altitude, an issue which has been studied with considerable focus, and are extremely sensitive to habitat degradation which affects their elevational ranges (Mahecha-Jiménez et al. 2011; Pyrcz & Garlacz, 2012; Díaz-Suárez et al. 2022). Species are often geographically limited, confined to single ranges or valleys, with only a few having wider distributions (Pyrcz et al. 2020). Even though their highly complicated alphataxonomy still require some research, it can be considered as fair, and fast species identification only occasionally presents any serious obstacles (Marín et al. 2017; Padrón et al. 2021). In addition, the spatial distribution patterns of Pronophilina are well-known and fine-tuned both geographically and altitudinally, and this is fundamental from the perspectives of biogeography and population ecology. (Padrón et al. 2021). The presence of Pronophilina is generally high in bamboo thickets, where the adults remain close to their host plants, which are mainly Chusquea Kunth bamboos (Poaceae) (Pyrcz et al. 2009; Mahecha et al. 2019; Pyrcz et al. 2020; Padrón et al. 2021). Finally, and importantly, the use of van Someren-Rydon traps baited with decomposing animal matter guarantees the obtaining of a numerically large, statistically viable sample, frequently representative of local Pronophilina communities within a limited time frame (Pyrcz et al. 2009; Díaz-Suárez et al. 2022).
On a global scale, the number of species in most animal groups, including Lepidoptera, either decreases steadily with increasing elevation or has a peak at mid-elevation (Brehm et al. 2007; Grytnes & Maccain, 2007; Despland et al. 2018). However, recent studies conducted throughout the Andes yield a somewhat different pattern, with several groups of Lepidoptera whose diversity increases with altitude, sometimes reaching the highest values close to timberline at some 2600-3000 m, for example some taxa of the Larentiinae subfamily of Geometridae (Brehm et al. 2003), or to that matter the Pronophilina (Pyrcz, 2010). In this study, we did an RBA of Pronophilina butterfly community structure and diversity along two transects in the uppermost cloud forests near timberline in Manu National Park, Cusco department in SE Peru with the aim to confirm that Pronophilina are good habitat preservation indicators in the Andes.
Material and methods
Study Area
Study area is situated in the upper Madre de Dios watershed, in the upper montane area of the Manu National Park, located in the department of Cusco in southern Peru (Figure 1). The Manu NP covers 17163 km2 of Amazonian rainforest, montane forests and Andean grasslands between 150 m and 4200 m. The Park was created in 1973 in order to preserve natural habitats of the upper River Manu in the basin of the Madre de Dios. It was recognized as Biosphere Reserve by the UNESCO in 1977, and it is the second largest national park of Peru and corresponds to the one of the top diversity terrestrial areas of the World (Myers et al. 2000).
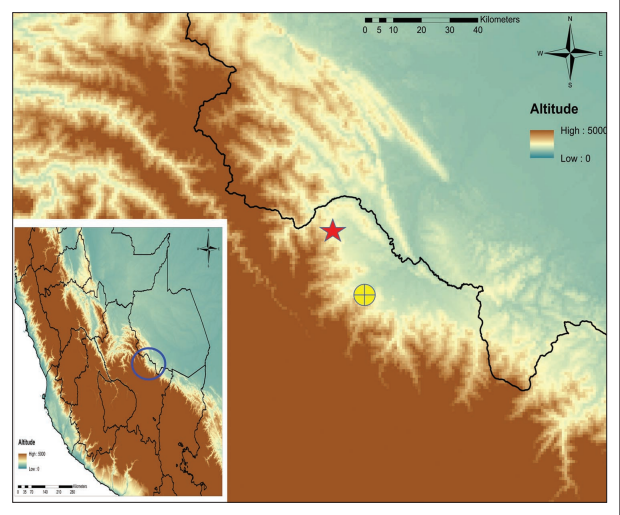 Figura 1.
Location map of study sites in the Manu National Park, Peru. The red star is the Qurqurpampa station and the yellow circle the Trocha Union station.
Figura 1.
Location map of study sites in the Manu National Park, Peru. The red star is the Qurqurpampa station and the yellow circle the Trocha Union station.
RBA methodology
Two altitudinal transects were set at 2800-3500 m.a.s.l. One of them situated 4 km north of the park rangers’ station Qurqurpampa (12º54’ S 71º47’W), along the trails with links it with the village of Callanga. The second transect was set along the so-called trail Trocha Union (13º06’ S 71º36’ W), which links the sector of Tres Cruces with the road going down the Valley of Kosñipata. Both transects covered three kinds of habitats: puna grassland, transitional shrubby forest (tree-line ecotone), and montane forest. Papilionoidea were sampled using entomological nets and 20 Van Someren-Rydon traps, baited with decomposing fish. Traps were separated by 50 m each, starting at grassland forest ecotone at 3200 m and going down to 2800 m. Transect walks and traps were surveyed for five consecutive days each sampling period. Traps were revised daily. Only entomological nets were used in the habitat of puna grassland for four consecutive days, traps were not installed due to poor sampling efficiency demonstrated in previous studies (Pyrcz et al. 2013). Field work was carried out by two people. Field work/sampling took place three times in October (2012), April (2013) and August (2013).
Taxonomy
All the material was set and examined in the laboratory of entomology at the Museo de Historia Natural de la Universidad Nacional de San Agustín (MUSA), Arequipa, Perú, and crossed-checked, also against the type specimens kept at the Nature Education Centre of the Jagiellonian University, Kraków, Poland (CEPUJ) and the Museo de Historia Natural de la Universidad Nacional Mayor San Marcos (MUSM), Lima, Perú. Additionally, the types illustrated at Butterflies of America website were examined. Most of the material is deposited at MUSA and voucher specimens in the collections of CEPUJ and MUSM. In the case of specimens requiring more sophisticated methods of taxonomic analysis, male and female genitalia were dissected and compared to type specimens, using standard procedures (Pyrcz et al. 2019).
Data analysis
As a quantitative measure both the abundance, the number of species, doubletons and singletons recorded at each station were given. For the evaluation of alpha diversity three diversity measures: Fisher alpha, Shannon-Entropy and Simpson-Inv. estimative were calculated. As a measure of dominance, the Berger-Parker, and of evenness the Pielou estimative were used (Mahecha-Jiménez et al. 2011; Pyrcz & Garlacz, 2012; Urbano et al. 2018). Moreover, the Hill numbers (q= 0, 1, 2) were estimated for each site as a complementary measure of alpha diversity (Hill, 1973; Moreno et al. 2011; Chao et al. 2014; Marín et al. 2014; Hsieh et al. 2016). The sample coverage and species diversitybased rarefaction/extrapolation (RE) sampling curves were performed by site sampling, in which computes the Hills numbers for rarefied and extrapolated samples with sample completeness up to the coverage value of double the reference sample size (Hsieh et al. 2016). Moreover, the sample-size and coverage-based RE of Hill numbers as a unified framework for estimating species diversity. Likewise, a sample coverage RE curve can compare sites that have different sizes in their samples (Hsieh et al. 2016). Sørensen similarity index was computed to assess the similarity among both sites. Sørensen’s coefficient of similarity was calculated based on presence/absence data for each species. It varies from 0 to 1, and a value close to 1 indicates greater similarity between sites, hence a low diversity ß (Magurran, 2013; Ganamé et al. 2020).
The Relative Abundance Distribution (RAD) was calculated to describe the sampled community (Chao et al. 2015; Cusack et al. 2015). The Akaike’s Information Criteria (AIC) was carried out to select the best RAD model for each site, with subsequent inferences based on the model with the lowest AIC value (Burnham & Anderson, 2002; Cusack et al. 2015). However, the fitting of models to RADs is an intuitive way of representing and comparing community structures, it does not in itself provide information on the relative rank each species occupies within the observed communities (Cusack et al. 2015). All analyses were carried out in R version 3.6.3 (R DEVELOPMENT CORE TEAM, 2020). Species accumulation curves were plotted using the package iNEXT (HSIEH et al. 2013), and analyses of community composition and structure were carried out in package diverse (Guevara et al. 2016) and BiodiversityR (Kindt & Coe, 2005).
Results
Community structure
Table I.
Abundance of the Pronophilina butterflies in each sampling site.
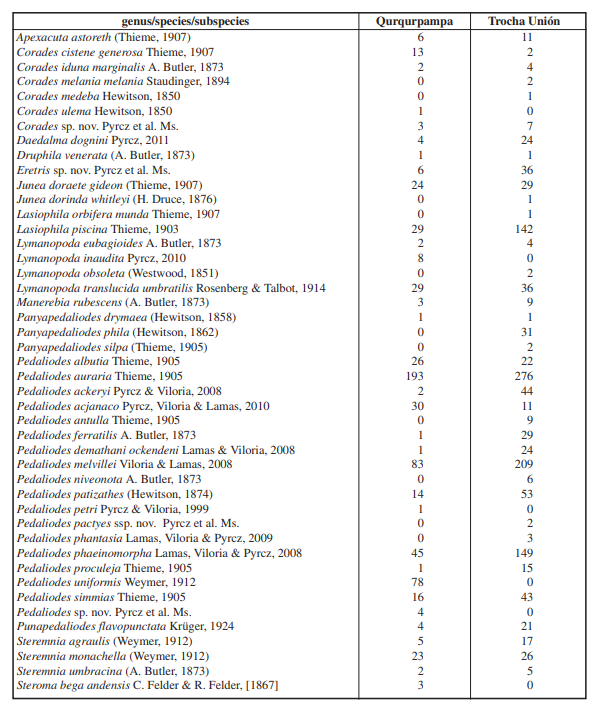
A total of 1941 specimens of the subtribe Pronophilina were collected, 664 along the Qurqurpampa, and 1310 along the Trocha Unión transects respectively. They belong to 15 genera and 45 species. Overall, 18 species (40%) belong to the genus Pedaliodes Butler, 1867. All the species of Pedaliodes account for a total of 1358 individuals which constitutes as much as 70% of the sample. The most abundant species in both transects is Pedaliodes auraria with 469 individuals overall, representing 24% of the sample. The second and third most abundant species also belong to the genus Pedaliodes (P. melvillei - 293, P. pheinomorpha - 194 specimens). Four species singletons and six doubletons were reported (Table I).
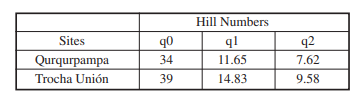
The sample of Trocha Unión presents slightly higher values of all principal diversity estimates (in particular Shannon, Simpson, and Fisher alpha) (Table II), and significant difference between the two samples was evidenced (U- Mann-Whitney test: p-value = 0.039). These results are supported by Hill numbers of values for both sites (Table III). On the other hand, there is a major value of dominance in the Qurqurpampa sample as shown by Berger-Parker index.
Table II.
Diversity estimates.
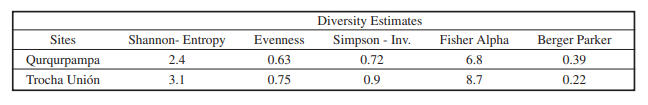
Table III.
Hill numbers or the effective number of species.
The analysis of sample coverage (Figure 2) and species diversity based on the RE curve (Figure 3) indicate to the sampling effort was appropriate for both sites. Additionally, the species diversity-sample coverage curves allowed to corroborate the good sampling effort (Figure 4). Although it is noteworthy that no curve reached an asymptote, this suggests that a more extensive sampling could potentially increase the number of species in the study area.
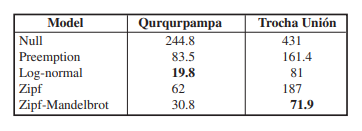
Five models of RAD were tested (null, preemption, lognormal, Zipf and Zipf-Mandelbrot models) (Table IV). According to the AIC criterion, the best adjustment for the sample of Trocha Unión turned out to be the Zipf-Mandelbrot model, whereas for Qurqurpampa the log-normal model (Table IV) (Figure 5). Our results are evidence of a similar pattern from the abundance distribution between dominant and rare species of butterflies in the study area.
Table IV.
RAD models with AIC values.
Faunal similarities
Overall, 13 species were found in one of the localities only. Significantly, however, as many as nine of the species reported in only one of the localities were either singletons or doubletons. Among five species which were reported in more individuals, four apparently indicate some faunal dissimilarities between the two sites. In fact, two species were reported in Trocha Unión only, the most southerly site of the two are known to occur most southwards as well, in the well-sampled Acjanaco area, in particular Pedaliodes antulla Thieme, 1905 and Pedaliodes niveonota Butler, 1873. On the other hand, Lymanopoda inaudita Pyrcz, 2004 reported in Qurqupampa only has never been collected in Acjanaco but, on the other hands, it was known from the Abra Malaga-San Luis area situated just northwards of the study area. The most significant absentee in Trocha Unión is Pedaliodes uniformis Weymer, 1912 one of the co-dominant species in Qurqurpampa. The absence of Panyapedaliodes phila (Hewitson, 1862) in Qurqupampa is hardly understandable since it is a very widely distributed, nearly Panandean species found frequently above 2800 m.
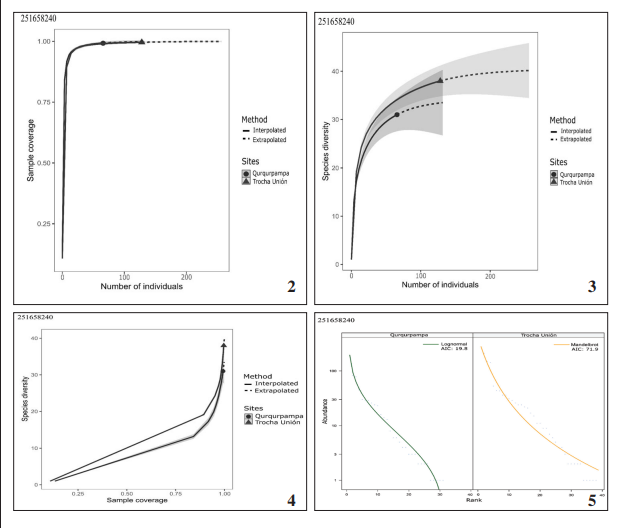 Figures 2-5.
2. Sample coverage based RE curve. 3. Species diversity based RE curve for each site. 4. Species diversity vs. Sample coverage based RE curve. 5. Relative Abundance Distribution (RAD) fitted to each site according to AIC criterion.
Figures 2-5.
2. Sample coverage based RE curve. 3. Species diversity based RE curve for each site. 4. Species diversity vs. Sample coverage based RE curve. 5. Relative Abundance Distribution (RAD) fitted to each site according to AIC criterion.
Sørensen similarity index is 0.66, which is a rather low value considered that the distance between the two collecting sites is only 30 km, and both are situated on the eastern slopes of the Cordillera de Vilcanota.
Altitudinal patterns
An overwhelming majority of the species recorded in both transects are those known to be associated with the highest strata of cloud forests, occurring elsewhere at 2800-3200 m, and found in well-preserved cloud forest. In particular, some species reported here are known to present extremely narrow elevational ranges and are basically restricted to the forest-paramo ecotone. Among such species we should list Corades sp. nov., Apexacuta astoreth (Thieme, 1907), Daedalma dognini Pyrcz, 2011, Pedaliodes acjanacoLamas, Viloria & Pyrcz, 2010, Pedaliodes proculeja Thieme, 1905 and Pedaliodes phantasia Viloria, Lamas & Pyrcz, 2009. On the other hand, some species reported here are generally found at lower elevations than those covered by the transects, in particular Lasiophila orbifera Butler, 1868, Corades medeba Hewitson, 1850, Lymanopoda obsolete (Westwood, [1851]), Pedaliodes petri Pyrcz & Viloria, 1999 and Steroma bega Westwood, [1850] occurring elsewhere at lower elevations, generally at 1800-2600 m. They were recorded either as singletons, doubletons or in three individuals in the latter species, which shows the elevational band covered in this study was above their usual altitudinal range. All five are, however, widespread, and common species and occasionally can be found elsewhere above 2800 m. One species collected on both transects as singletons, Panyapedaliodes drymaea (Hewitson, 1858), is an indicator of secondary areas. Punapedaliodes flavopunctata (Staudinger, 1894) is the only species in the sample which is associated with puna grasslands and is found usually at considerably higher elevations than 3200 m. Its presence in both transects indicates the sampling occasionally took place slightly above the upper forest limit.
Discussion
This is the first RBA study for Pronophilina butterflies diversity in a sanctuary protected by law in Manu National Park, Cusco department in SE Peru, and our study proves once again that sampling with van Someren-Rydon baited traps is an extremely efficient method in carrying out quick surveys of Lepidoptera diversity and community structure in both lowland and montane rainforest habitats for local or regional studies (De Vries & Walla, 2001; Maicher et al. 2020), as well as in ecological assessments of habitat preservation versus anthropogenic activity within a limited time frame (Aduse-Poku et al. 2012; Whitworth et al. 2016). Such a method is efficient not only by attracting large numbers of individuals which enables to produce viable samples for statistical analysis, but it is also relatively specifically unbiased being equally attractive for all the species of a butterfly community, at least true for cloud forest Satyrinae (Pyrcz et al. 2009; Pyrcz & Garlacz, 2012). Biased sampling method is frequently a problem in similar studies, thus the use a wide array of baits is necessary (Maicher et al. 2020).
This study is, in our opinion, also valuable because it is one of the few to offer a comprehensive comparison of two ecologically similar and geographically marginally set apart localities, allowing a fine-tuned insight into intricate zoogeography of one of topographically most complex areas of the Andes. In fact, the two sampling transects are only 30 km away, both situated on the eastern slopes of the Vilcanota range and are not separated by any noticeable topographical event. And yet, their Sørensen similarity index is surprisingly low, and several interesting differences can be pointed out. For example, the absence of Pedaliodes uniformis the fifth most abundant species overall, and Lymanopoda inaudita along the Trocha Unión transect most likely reflects their south-eastern distribution limit, which is relevant from a zoogeographical perspective as it uncovers a discreet geographical pattern. On the other hand, the absence of Panyapedaliodes phila, a widespread Andean species common at 2600-3000 m along the Qurqurpampa transect is difficult to explain. For comparison, the faunas of Pronophilina of two sites separated by 50 km in the Venezuela Cordillera de Merida were also studied (Pyrcz & Wojtusiak, 2002; Pyrcz & Garlacz, 2012). In that case, the Sørensen similarity index was significantly higher, reaching 0.96. The two samples in Venezuela differed by one species only. Our results indicate much higher beta diversity, understood as species turn-over on a geographical scale, in the Peruvian Andes. Also, the species richness in Venezuela, whereas the altitudinal span covered was much wider, 700 m compared to 400 m in this study, was considerably lower with 22 and 23 species respectively. Higher diversity of two Peruvian localities was also reflected by Shannon and Fisher alpha indices.
Faunal structure of Pronophilina evidenced in this study is similar with other sampled Andean localities, such as Monte Zerpa and El Baho in the Venezuelan Cordillera de Merida (Pyrcz & Wojtusiak, 2002; Pyrcz & Garlacz, 2012), Alto del Zuque-Bogotá (Mahecha-Jiménez et al. 2011) and Frailejonal- La Calera (Díaz-Suárez et al. 2022) in the Colombian Eastern Cordillera; Belmira (Pyrcz et al. 2018) in the Colombian Central Cordillera; Tambito (Pyrcz & Wojtusiak, 1999), and Golondrinas (Pyrcz et al. 2009) on the Pacific slopes in Colombia and Ecuador, and Chachapoyas in northern Peru (Pyrcz, 2004). All the communities of Pronophilina are characterized by the dominance, in terms of species-richness and abundance, of Pedaliodes¸ with one to three strong dominant or co-dominant species, and a comparative under-representation of other genera in the sample. Such a faunal structure is not an artefact of the method of sampling nor seems to be related with the geographic setting of sampling locality or altitudinal span covered. The species dominance of Pedaliodes in the sample is not surprising given that it is also the most species-rich genus of Pronophilina accounting for nearly 50% of all known species in this subtribe (Pyrcz, 2004). However, the dominance of Pedaliodes in terms of abundance is less straightforward to interpret. Also, in the less diverse communities the dominance of a single species of Pedaliodes is more accentuated, which is particularly striking in the case of El Baho sample where Pedaliodes minabilisPyrcz, 2008 constitutes as much as 53% of the sample (Pyrcz & Garlacz, 2012). Interestingly, dominant species of Pedaliodes in each locality belong to completely unrelated clades of the genus (Pyrcz et al. in prep.).
Our results suggest good preservation of the patches of uppermost forest where both samplings were carried out. This is shown, first, by the presence of species indicators of well-preserved habitats, in particular Corades sp. nov., Apexacuta astoreth (Thieme, 1907), Lymanopoda inauditaPyrcz, 2010, or Pedaliodes phaeinomorpha Viloria, Lamas & Pyrcz, 2008, the latter one of the co-dominant species in the study area, elsewhere extremely localized and rare, for example along the nearby Acjanaco - Manu Road due to high disturbance of forest cover at 3000-3200 m. At the same time, very few species related with disturbed forest habitats were found in the study area, in particular only one individual of Panyapedaliodes drymaea (Hewitson, 1858) which is one of the most prominent indicators of high anthropogenic disturbance, very few specimens of Steremnia umbracina (Butler, 1873), another species related with secondary habitats, and none of Pedaliodes palaepolis (Hewitson, 1878), again a secondary forest indicator species (Pyrcz, 2004; Mahecha-Jiménez et al. 2011; Díaz-Suárez et al. 2022). Second, the RAD models fit in Trocha Unión (Zipf-Mandelbrot) and Qurqurpampa (log-normal) are also indicative of undisturbed, natural, varied and mature communities (may, 1975; Hill et al. 1995; Hill & Hamer, 1998; Marimon et al. 2015; Harterreiten-Souza et al. 2020; Nallis, 2021). In fact, the Zipf-Mandelbrot model assumes that, in the succession processes, late colonizing species have a specialized niche and thus are more difficult to find, while early colonizing species are more generalist and dominate in abundance regardless of the time of succession (Magurran, 2013; Harterreiten-Souza et al. 2020; Nallis, 2021). This model also assumes that colonization is initially stochastic depending on the regional abundance of generalist species, but that mature communities are determined according to the niche of late species (Nallis, 2021). The log-normal model assumes that ecological communities will have many species with intermediate abundance and few rare or common species (Schluter & Ricklefs, 1993; Harterreiten-Souza et al. 2020; Nallis, 2021). It has been argued that the use of RAD is a much more reliable method in comparing communities than non-parametric indices of diversity (Kim et al. 2013; Sæther et al. 2013). The latter, frequently used in the studies of biodiversity are shown to oversimplify the complex structural nature of communities and are not independent of sampling intensity, thus on sample size as well (Mouillot & Lepretre, 2000; Dornelas et al. 2011). Interestingly, this does not seem to be the case in our study. Most indices show closely similar values despite the fact that the Trocha Unión sample contains two times as many records as Qurqurpampa.
In sum, our study confirms that Pronophilina, thanks to their ecological and also behavioural qualities, such as habitat fidelity coupled with low adult vagility, and high sensibility to human made disturbance, which immediately express in their community structure, can be explored in the assessment of cloud forest state of preservation, and thus in formulating future conservation policies. In addition, to their other features, such as easy high species richness, standard sampling procedures, rapid data gathering protocols, and good knowledge of taxonomy among others, they surpass as potential habitat preservation indicators any other group of Andean Lepidoptera.
Acknowledgments
This research was supported by NCN grant Harmonia-10 2018/30/M/NZ8/00293 “Evolutionary biogeography and diversification of the predominantly Andean butterfly subtribe Pronophilina (Nymphalidae, Satyrinae) based on phylogenetic data generated using modern molecular methods”. Taxonomical studies, genital dissections and barcoding was carried out in the laboratory of the Nature Education Centre, Jagiellonian University, Kraków, Poland (CEPUJ). Field work was performed under authorization R. J. Nº 0010-2012 SERNANP-SNM issued by the “Servicio Nacional de Áreas Naturales Protegidas por el Estado” (SERNANP), Ministerio del Ambiente, Peru.
References
Aduse-Poku, K., William, O., Oppong, S. K., Larsen, T., Ofori-Boateng, C., & Molleman, F., (2012). Spatial and temporal variation in butterfly biodiversity in a West African forest: lessons for establishing efficient rapid monitoring programmes. African Journal of Ecology, 50(3), 326-334. https://doi.org/10.1111/j.13652028.2012.01328.x
Aide, T. M., Grau, H. R., Graesser, J., Andrade-Núñez, M. J., Aráoz, E., Barros, A. P., & Zimmerer, K. S. (2019). Woody vegetation dynamics in the tropical and subtropical Andes from 2001 to 2014: Satellite image interpretation and expert validation. Global change biology, 25(6), 2112-2126. https://doi.org/10.1111/gcb.14618 PMid:30854741 PMCid:PMC6849738
Alonso, L. E., Deichmann, J. L., Mckenna, S. A., Naskrecki, P., & Richards, S. J. (2011). Still Counting: Biodiversity Exploration for Conservation - The First 20 Years of the Rapid Assessment Program. Conservation International.
Armenteras, D., Rodríguez, N., Retana, J., & Morales, M. (2011). Understanding deforestation in montane and lowland forests of the Colombian Andes. Regional Environmental Change, 11(3), 693-705. https://doi.org/10.1007/s10113-010-0200-y
Bax, V., Castro-Nunez, A., & Francesconi, W. (2021). Assessment of Potential Climate Change Impacts on Montane Forests in the Peruvian Andes: Implications for Conservation Prioritization. Forests, 12(3), 375. https://doi.org/10.3390/f12030375
Brehm, G., Colwell, R. K., & Kluge, J. (2007). The role of environment and mid-domain effect on moth species richness along a tropical elevational gradient. Global Ecology and Biogeography, 16(2), 205-219. https://doi.org/10.1111/j.1466-8238.2006.00281.x
Burnham, K. P., & Anderson, D. R. (2002). Model selection and multimodel inference: a practical informationtheoretic approach, second edition. Springer-Verlag.
Castellanos-Mora, L., & Agudelo-Hz, W. (2020). Spatial Scenarios of Land-Use/Cover Change for the Management and Conservation of Paramos and Andean Forests in Boyacá, Colombia. Environmental Sciences Proceedings, 3(1), 87. https://doi.org/10.3390/IECF2020-08023
Chao A, Gotelli N. J., Hsieh T. C., Sander, E. L., Ma, K. H., Colwell, R. K., & Ellison, A. M. (2014). Rarefaction and extrapolation with Hill numbers: a framework for sampling and estimation in species diversity studies. Ecological Monographs, 84(1), 45-67. https://doi.org/10.1890/13-0133.1
Chao, A., Hsieh, T. C., Chazdon, R. L., Colwell, R. K., & Gotelli, N. J. (2015). Unveiling the species-rank abundance distribution by generalizing the Good-Turing sample coverage theory. Ecology, 96(5), 1189-1201. https://doi.org/10.1890/14-0550.1 PMid:26236834
Cusack, J. J., Dickman, A. J., Rowcliffe, J. M., Carbone, C., Macdonald, D. W., & Coulson, T. (2015). Random versus game trail-based camera trap placement strategy for monitoring terrestrial mammal communities. PloS one, 10(5), e0126373. https://doi.org/10.1371/journal.pone.0126373 PMid:25950183 PMCid:PMC4423779
Despland, E., Humire, R., & Martín, S. S. (2012). Species Richness and Phenology of Butterflies along an Altitude Gradient in the Desert of Northern Chile. Arctic, Antarctic, and Alpine Research, 44(4), 423-431. https://doi.org/10.1657/1938-4246-44.4.423
Díaz-Suárez, V., Mahecha-J., O., Andrade-C, M. G., & Pyrcz, T. W. (2022). Perturbación antrópica afectando a los patrones de diversidad en Satyrinae de montaña Pronophilina Reuter, 1896 en un bosque altoandino en Colombia (Lepidoptera: Nymphalidae). SHILAP Revista de lepidopterología, 50(200), 709-728. https://doi.org/10.57065/shilap.263
Dornelas, M., Soykan, C., & Ugland, K. I. (2011). Biodiversity and disturbance. Biological diversity. In A. E. Magurran & B. J. Mcgill. Frontiers in measurement and assessment. Oxford University Press.
Ferrer-Paris, J. R., Sanchez-Mercado, A., Viloria, A. L., & Donaldson, J. (2013). Congruence and diversity of butterfly host plant associations at higher taxonomic levels. PLoS ONE, 8, e63570. https://doi.org/10.1371/journal.pone.0063570 PMid:23717448 PMCid:PMC3662771
Ganamé, M., Bayen, P., Ouédraogo, I., Dimobe, K., & Thiombiano, A. (2020). Woody species composition, diversity and vegetation structure of two protected areas along a climatic gradient in Burkina Faso (West Africa). Folia Geobotanica, 54, 163-175. https://doi.org/10.1007/s12224-019-09340-9
Hamer, K. C., & Hill, J. K. (2004). Determining impacts of habitat modification on diversity of tropical forest fauna: the importance of spatial scale. Journal of Applied Ecology, 41(4), 744-754. https://doi.org/10.1111/j.00218901.2004.00926.x
Harterreiten-Souza, É. S., Pujol-Luz, J. R., Capellari, R. S., Bickel, D., & Sujii, E. R. (2020). Diversity and spatial distribution of predacious Dolichopodidae (Insecta: Diptera) on organic vegetable fields and adjacent habitats in Brazil. Florida Entomologist, 103(2), 197-205. https://doi.org/10.1653/024.103.0207
Henao-Bañol, E. R., & Gantiva-Q, C. H. (2020). Mariposas (Lepidoptera: Hesperioidea-Papilionoidae) del bosque seco tropical (BST) en Colombia. Conociendo la diversidad en un ecosistema amenazado. Boletín Científico del Museo de Historia Natural de la Universidad de Caldas, 24(1), 150-196. https://doi.org/10.17151/bccm.2020.24.1.10
Hill, J. K., Hamer, K. C., Lace, L. A., & Banham, W. M. T. (1995). Effects of selective logging on tropical forest butterflies on Buru, Indonesia. Journal of Applied Ecology, 32, 754-760. https://doi.org/10.2307/2404815
Horwath, A. B., Royles, J., Tito, R., Gudiño, J. A., Salazar Allen, N., Farfan-Rios, W., & Griffiths, H. (2019). Bryophyte stable isotope composition, diversity and biomass define tropical montane cloud forest extent. Proceedings of the Royal Society B, 286(1895), 20182284. https://doi.org/10.1098/rspb.2018.2284 PMid: 30963945 PMCid:PMC6364587
Hutter, C. R., Lambert, S. M., & Wiens, J. J. (2017). Rapid Diversification and Time Explain Amphibian Richness at Different Scales in the Tropical Andes, Earth’s Most Biodiverse Hotspot. The American Naturalist, 190(6), 828-843. https://doi.org/10.1086/694319 PMid:29166157
Ivory, S., & Misrachi, M. (2014). Commodities and biodiversity in the watersheds of the Andes impacts of commodity development on biodiversity and ecosystem services in the watersheds of the Andes. MacArthur Foundation, United Nations Environment Programme.
Kim, D. H., Cho, W. S., & Chon, T. S. (2013). Self-organizing map and species abundance distribution of stream benthic macroinvertebrates in revealing community patterns in different seasons. Ecological informatics, 17, 14-29. https://doi.org/10.1016/j.ecoinf.2013.06.006
Lamas, G., Viloria, A. L., & Pyrcz, T. W. (2004). Subtribe Pronophilini. In G., Lamas. Atlas of Neotropical Lepidoptera, Checklist: Part 4A, Hesperioidea-Papilionoidea. Association for Tropical Lepidoptera.
Lodh, R., & Agarwala, B. K. (2016). Rapid assessment of diversity and conservation of butterflies in Rowa Wildlife Sanctuary: An Indo-Burmese hotspot-Tripura, NE India. Tropical Ecology, 57(2), 231- 242.
Loughlin, N. J., Gosling, W. D., Mothes, P., & Montoya, E. (2018). Ecological consequences of post-Columbian indigenous depopulation in the Andean-Amazonian corridor. Nature ecology & evolution, 2, 1233-1236. https://doi.org/10.1038/s41559-018-0602-7 PMid:30013131
Mahecha-Jiménez, O. J., Dumar-Rodriguez, J. C., & Pyrcz, T. W. (2011). Efecto de la fragmentación del hábitat sobre las comunidades de mariposas de la tribu Pronophilini a lo largo de un gradiente altitudinal en un bosque andino en Bogotá (Colombia) (Lepidoptera: Nymphalidae, Satyrinae). SHILAP Revista de lepidopterología, 39(153), 117-126.
Mahecha, O., Garlacz, R., Andrade, M. G., Prieto, C., & Pyrcz, T. (2019). Island biogeography in continental areas: inferring dispersal based on distributional patterns of Pronophilina butterflies (Nymphalidae: Satyrinae) in the north Andean massifs. Revista Mexicana de Biodiversidad, 90, e902796. https://doi.org/10.22201/ib.20078706e.2019.90.2796
Magurran, A. E. (2013). Measuring Biological Diversity. John Wiley & Sons.
Maicher, V., Sáfián, S., Murkwe, M., Delabye, S., Przybylowicz, L., Potockx, P., & Tropek, R. (2020). Seasonal shifts of biodiversity patterns and species’ elevation ranges of butterflies and moths along a complete rainforest elevational gradient on Mount Cameroon. Journal of Biogeography, 47(2), 342-354. https://doi.org/10.1111/jbi.13740
Marimon, B. S., Colli, G. R., Marimon-Junior, B. H., Mews, H. A., Eisenlohr, P. V., Feldpausch, T. R., & Phillips, O. L. (2015). Ecology of floodplain campos de murundus savanna in southern Amazonia. International Journal of Plant Sciences, 176(7), 670-681. https://doi.org/10.1086/682079
Marín, M. A., Álvarez, C. F., Giraldo, C. E., Pyrcz, T. W., Uribe, S. I., & Vila, R. (2014). Mariposas en un bosque de niebla andino periurbano en el valle de Aburrá, Colombia. Revista Mexicana de Biodiversidad, 85, 200-208. https://doi.org/10.7550/rmb.36605
Marín, M. A., Cadavid, I. C., Valdés, L., Álvarez, C. F., Uribe, S. I., Vila, R., & Pyrcz, T. W. (2017). DNA barcoding of an assembly of montane Andean butterflies (Satyrinae): Geographical scale and identification performance. Neotropical Entomology, 46(5), 514-523. https://doi.org/10.1007/s13744-016-0481-z PMid:28116647
May, R. M. (1975). Patterns of species abundance and diversity. In M. L. Cody & J. M. Diamond. Ecology and evolution of communities. Cambridge University Press.
Meseguer, A. S., Michel, A., Fabre, P. H., Escobar, O. A. P., Chomicki, G., Riina, R., Antonelli, A., Pierre-Olivier A., Delsuc, F., & Condamine, F. L. (2022). Diversification dynamics in the Neotropics through time, clades, and biogeographic regions. ELife, 11, e74503. https://doi.org/10.7554/eLife.74503 PMid:36300780 PMCid: PMC9668338
Moreno, C. E., Barragán, F., Pineda, E., & Pavón, N. P. (2011). Reanalizando la diversidad alfa: alternativas para interpretar y comparar información sobre comunidades ecológicas. Revista Mexicana de Biodiversidad, 82, 1249-1261. https://doi.org/10.22201/ib.20078706e.2011.4.745
Moreira, H., Kuipers, K. J., Posthuma, L., Zijp, M. C., Hauck, M., Huijbregts, M. A., & Schipper, A. M. (2023). Threats of land use to the global diversity of vascular plants. Diversity and Distributions, 29, 689-697. https://doi.org/10.1111/ddi.13693
Myers, N., Mittermeier, R. A., Mittermeier, C. G., Da Fonseca, G. A. B., & Kent, J. (2000). Biodiversity hotspots for conservation priorities. Nature, 403, 853-858. https://doi.org/10.1038/35002501 PMid:10706275
Nallis, B. G. (2021). Fatores que estruturam metacomunidades em fitotelmos de restinga. (Defesa de Dissertação de Mestrado). Universidade Federal de São Carlos.
Oliveira, J. C., Santos, R. D., Lopes-Silva, M. L., Barros, L. D. P. V., Risse-Quaioto, B., Militão, C. M., & Rocha, C. F. D. (2020). Reptiles of the Serra das Torres Natural Monument: using the Rapid Assessment method to fill a knowledge gap in the Atlantic Forest of southeastern Brazil. Biota Neotropica, 20(2), e20190726. https://doi.org/10.1590/1676-0611-bn-2019-0726
Pennington, R. T., & Lavin, M. (2010). Contrasting plant diversification histories within the Andean biodiversity hotspot. Proceedings of the National Academy of Sciences, 107(31), 13783-13787. https://doi.org/10.1073/pnas.1001317107 PMid:20643954 PMCid:PMC2922214
Pearson, D. L. (1994). Selecting indicator taxa for the quantitative assessment of biodiversity. Philosophical Transaction of the Royal Society of London, 45,75-79. https://doi.org/10.1098/rstb.1994.0088 PMid:7972358
Pyrcz, T. W., & Wojtusiak, J. (1999). Mariposas de la tribu Pronophilini de la Reserva Forestal Tambito, Cordillera Occcidental, Colombia. Segunda parte. Patrones de distribución altitudinal (Lepidoptera: Nymphalidae: Satyrinae). SHILAP Revista de lepidopterología, 27(106), 203-213.
Pyrcz, T. W., & Wojtusiak, J. (2002). The vertical distribution of pronophiline butterflies (Nymphalidae, Satyrinae) along an elevational transect in Monte Zerpa (Cordillera de Mérida, Venezuela) with remarks on their diversity and parapatric distribution. Global Ecology & Biogeography, 11, 211-221. https://doi.org/10.1046/j.1466822X.2002.00285.x
Pyrcz, T. W. (2004). Pronophiline butterflies of the highlands of Chachapoyas in northern Peru: faunal survey, diversity, and distribution patterns (Lepidoptera, Nymphalidae, Satyrinae). Genus, 15, 455-622.
Pyrcz, T. W., Wojtusiak, J., & Garlacz, R. (2009). Diversity and Distribution Patterns of Pronophilini Butterflies (Lepidoptera: Nymphalidae: Satyrinae) along an Altitudinal Transect in North-Western Ecuador. Neotropical Entomology, 38(6), 716-726. https://doi.org/10.1590/S1519-566X2009000600003 PMid:20098916
Pyrcz, T. W. (2010). Wybrane zagadnienia z taksonomii, zoogeografii i ewolucji faun górskich na przyk.adzie grupy modelowej motyli z plemienia Pronophilini (Nymphalidae). Mantis.
Pyrcz, T. W., & Garlacz, R. (2012). The presence-absence situation and its impact on the assemblage structure and interspecific relations of Pronophilina butterflies in the Venezuelan Andes (Lepidoptera: Nymphalidae). Neotropical entomology, 41(3), 186-195. https://doi.org/10.1007/s13744-012-0031-2 PMid:23950042 PMCid:PMC3380249
Pyrcz, T. W., Cerdeña, J., & Huamani, E. (2013). Systematics, bionomics and zoogeography of high Andean pedaliodines. Part 14: Two new species of Pedaliodes Butler from the Huancabamba Deflection in southern Ecuador and northern Peru (Lepidoptera: Nymphalidae: Satyrinae). Genus, 24(2), 131-141.
Pyrcz, T. W., Clavijo, A., Uribe, S., Marín, M. A., Álvarez, C. F., & Zubek, A. (2016). Páramo de Belmira as an important centre of endemism in the northern Colombian Andes: New evidence from Pronophilina butterflies (Lepidoptera: Nymphalidae, Satyrinae, Satyrini). Zootaxa, 4179(1), 77-102. https://doi.org/10.11646/zootaxa.4179.1.3 PMid:27811691
Pyrcz, T. W., Lorenc-Brudecka, J., Zubek, A., Prieto, C., Boyer, P., Florczyk, K., Waclawik, B., & LachowskaCierlik, D. (2019). Considerations on the Taxonomy of the Genus Arhuaco Adams and Bernard 1977, and its Relationships with the Genus Pronophila Doubleday, [1849] (Nymphalidae, Satyrinae). Neotropical entomology, 48(2), 302-313. https://doi.org/10.1007/s13744-018-0641-4 PMid:30414019 PMCid: PMC6443605
Pyrcz, T. W., Zubek, A., Boyer, P., Nakamura, I., Waclawik, B., & Florczyk, K. (2020). Revisional notes on the cloud forest butterfly genus Oxeoschistus Butler in Central America (Lepidoptera: Nymphalidae: Satyrinae). Neotropical entomology, 49, 392-411. https://doi.org/10.1007/s13744-019-00757-7 PMid:32172389 PMCid: PMC7253525
Quitián, M., Santillán, V., Espinosa, C. I., Homeier, J., Böhning-Gaese, K., Schleuning, M., & Neuschulz, E. L. (2018). Elevation-dependent effects of forest fragmentation on plant–bird interaction networks in the tropical Andes. Ecography, 41(9), 1497-1506. https://doi.org/10.1111/ecog.03247
Pérez-Escobar, O. A., Chomicki, G., Condamine, F., Karremans, A., Bogarín, D., Matzke, N., Silvestro, D., & Antonelli, A. (2017). Recent origin and rapid speciation of Neotropical orchids in the world’s richest plant biodiversity hotspot. New Phytologist, 215(2), 891-905. https://doi.org/10.1111/nph.14629 PMid:28631324 PMCid:PMC5575461
R DEVELOPMENT CORE TEAM, VERSION 3.6.3. (2020). R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. http://www.R-project.org/
Sæther, B. E., Engen, S., & Grøtan, V. (2013). Species diversity and community similarity in fluctuating environments: parametric approaches using species abundance distributions. Journal of Animal Ecology, 82(4), 721-738. https://doi.org/10.1111/1365-2656.12068 PMid:23578202
Sharma, N., & Sharma, S. (2021). Assemblages and seasonal patterns in butterflies across different ecosystems in a sub-tropical zone of Jammu Shiwaliks, Jammu and Kashmir, India. Tropical Ecology, 62, 261-278. https://doi.org/10.1007/s42965-020-00139-w
Schluter, D., & Ricklefs, R. E. (1993). Species Diversity: An Introduction to the Problem. In R. E. Ricklefs & D. Schluter. Species Diversity in Ecological Communities: Historical and Geographical Perspectives. University of Chicago Press.
Soh, M. C., Mitchell, N. J., Ridley, A. R., Butler, C. W., Puan, C. L., & Peh, K. S. H. (2019). Impacts of habitat degradation on tropical montane biodiversity and ecosystem services: a systematic map for identifying future research priorities. Frontiers in Forests and Global Change. https://doi.org/10.3389/ffgc.2019.00083
Tettey, C. N. D., Anderson, R. S., & Kyerematen, R. (2020). Rapid Assessment of Butterfly Diversity of Two Proposed Community Resource Management Areas (CREMAs) in the Western North Region of Ghana: Implication for Conservation. Biodiversitas, 21, 1-36. https://doi.org/10.13057/biodiv/d210837
Toledo-Aceves, T., Meave, J. A., González-Espinosa, M., & Ramírez-Marcial, N. (2011). Tropical montane cloud forests: current threats and opportunities for their conservation and sustainable management in Mexico. Journal of environmental management, 92(3), 974-981. https://doi.org/10.1016/j.jenvman.2010.11.007 PMid:21144641
Urbano, P., Mahecha-J., O., Suárez, E., Izquierdo, V., & Díaz-S., V. (2018). Variación temporal del ensamblaje de mariposas asociadas a la Cuenca de la Calaboza, Yopal, Casanare, Colombia (Lepidoptera: Papilionoidea). SHILAP Revista de lepidopterología, 46(184), 533-550. https://doi.org/10.57065/shilap.745
Whitworth A., Villacampa, J., Brown, A., Pillco Huarcaya, R, Downie, R., & Macleod, R. (2016). Past Human Disturbance Effects upon Biodiversity are Greatest in the Canopy; A Case Study on Rainforest Butterflies. PLoS One, 11(3), e0150520. https://doi.org/10.1371/journal.pone.0150520 PMid:26950438 PMCid:PMC4780695
Notes
Author notes
1 Universidad Nacional de San Agustín de Arequipa, Museo de Historia Natural, Av. Alcides Carrión s/n, Arequipa. PERÚ / PERU. E-mail: cerdenajoseal@yahoo.es
2 Nature Education Centre, Jagiellonian University, Gronostajowa, 5, PL-30-387 Kraków. POLONIA / POLAND y / and Biogeografía y Ecología Evolutiva Neotropical BEEN, Universidad Distrital Francisco José de Caldas, Carrera 3 Nº 26A-40, Bogotá. COLOMBIA / COLOMBIA
3 Nature Education Centre, Jagiellonian University, Gronostajowa, 5, PL-30-387 Kraków. POLONIA / POLAND. E-mail: rafal.garlacz@uj.edu.pl
4 Universidad Nacional de San Agustín de Arequipa, Museo de Historia Natural, Av. Alcides Carrión s/n, Arequipa. PERÚ / PERU. E-mail: jjackie4u@gmail.com
5 Universidad Nacional de San Agustín de Arequipa, Museo de Historia Natural, Av. Alcides Carrión s/n, Arequipa. PERÚ / PERU. E-mail: joalarp@gmail.com
6 Nature Education Centre, Jagiellonian University, Gronostajowa, 5, PL-30-387 Kraków. POLONIA / POLAND. E-mail: klaudia.florczyk@uj.edu.pl
7 Universidad Nacional de San Agustín de Arequipa, Museo de Historia Natural, Av. Alcides Carrión s/n, Arequipa. PERÚ / PERU. E-mail: markro321@gmail.com
8 Nature Education Centre, Jagiellonian University, Gronostajowa, 5, PL-30-387 Kraków. POLONIA / POLAND y / and Department of Evolution of Invertebrates, Institute of Zoology and Biomedical Research, Jagiellonian University, Gronostajowa, 9, PL-30-387 Kraków. POLONIA / POLAND. E-mail: tomasz.pyrcz@uj.edu.pl
9 Nature Education Centre, Jagiellonian University, Gronostajowa, 5, PL-30-387 Kraków. POLONIA / POLAND y / and Biogeografía y Ecología Evolutiva Neotropical BEEN, Universidad Distrital Francisco José de Caldas, Carrera 3 Nº 26A-40, Bogotá. COLOMBIA / COLOMBIA. E-mail: diazsvane@gmail.com
* Autor para la correspondencia / Corresponding author: oscarmahecha23@gmail.com
