Artículos
A morphological comparative study of proboscis sensilla between Pierinae and Coliadinae (Lepidoptera: Pieridae)
Estudio morfológico comparativo de la sensila de la probóscide entre Pierinae y Coliadinae (Lepidoptera: Pieridae)
 Yağmur Mengi yagmurmengi@gmail.com
Yağmur Mengi yagmurmengi@gmail.com
A morphological comparative study of proboscis sensilla between Pierinae and Coliadinae (Lepidoptera: Pieridae)
Shilap Revista de Lepidopterología, vol. 52, no. 206, pp. 261-267, 2024
Sociedad Hispano-Luso-Americana de Lepidopterología
Received: 11 September 2023
Accepted: 29 October 2023
Published: 30 June 2024
Abstract: Pontia edusa (Linnaeus, 1758) and Colias croceus (Geoffroy, 1785, in Fourcroy), which are in different subfamilies within Pieridae, feed on nectar. In this study, we tested the hypothesis that sensilla differ in the subfamily category. For this purpose, we investigated the ultrastructural structure of proboscis and sensilla in Pontia edusa (Pierinae) and Colias croceus (Coliadinae) by scanning electron microscopy (SEM). The composition of the wall of proboscis, their surface structure, the shape and distribution of different types of sensilla were compared. Three sensillum types are located on the proboscises of both species, sensilla chaetica, sensilla basiconica, and sensilla styloconica being most common in Lepidoptera. It was determined that the size and distribution of sensilla and especially the structure of sensilla styloconica differ in two species. The results will contribute to studies on food and proboscis morphology of Lepidoptera, and will also be beneficial in further studies on the significance of proboscis and sensilla structure in classification.
Keywords: Lepidoptera, Pieridae, scanning electron microscopy, sensory organ, proboscis, sensilla.
Resumen: Pontia edusa (Linnaeus, 1758) y Colias croceus (Geoffroy, 1785, in Fourcroy), que pertenecen a subfamilias diferentes dentro de Pieridae, se alimentan de néctar. En este estudio, pusimos a prueba la hipótesis de que las sensilas difieren en la categoría de subfamilia. Para ello, investigamos la estructura ultraestructural de la probóscide y la sensila en Pontia edusa (Pierinae) y Colias croceus (Coliadinae) mediante microscopía electrónica de barrido (SEM). Se compararon la composición de la pared de la probóscide, su estructura superficial, la forma y la distribución de los distintos tipos de sensila. Tres tipos de sensilla se localizan en las probóscides de ambas especies, siendo la sensilla chaética, la sensilla basicónica y la sensilla estilocónica las más comunes en los Lepidoptera. Se determinó que el tamaño y la distribución de la sensilla y, especialmente, la estructura de la sensilla estilocónica, difieren en las dos especies. Los resultados contribuirán a los estudios sobre la alimentación y la morfología de la probóscide de los Lepidoptera y también serán beneficiosos en estudios posteriores sobre la importancia de la probóscide y la estructura de las sensilas en la clasificación.
Palabras clave: Lepidoptera, Pieridae, microscopía electrónica de barrido, órgano sensorial, probóscide, sensilla.
Introduction
Most adult lepidoptera feed on liquid foods through their proboscis. The proboscis consists of two extremely long galeas (Faucheux, 2013). These surround the food canal, interlocking with the cuticular processes on the dorsal and ventral sides called legulae (Bourgogne, 1951; Charlanes & Gaumont, 1960). The morphology of the proboscis changes depending on food-seizing habits (Büttiker et al. 1996; Krenn & Kristensen, 2000; Molleman et al. 2005; Faucheux, 2013). Sensilla in the proboscis of Lepidoptera plays a major role in host location and feeding behavior (Zaspel et al. 2013; Gilbert, 1972; Monaenkova, et al. 2012). The most common sensilla can be subdivided into six groups based on their external morphology (Krenn, 2010; Faucheux, 2013). All of them are located on the outer wall of the hose. The food canal contains only one-pore sensilla basiconica (Faucheux, 2013). Sensilla styloconica consists of a variety of differently shaped styli and shorter terminal sensory cones (Krenn, 1998, 2010; Kvello et al. 2006). There is an astonishing variety of Sensilla styloconica lengths and shapes in different families. The plesiomorphic features of the proboscis of Papilionoidea are that it contains vertically elongated exocuticular ribs forming the galeal wall, cuticular spines limited to the ventral side of the proximal galea, and 2-rows of grooved sensilla styloconica limited to the apex (Paulus & Krenn, 1996). Paulus & Krenn (1996) studied 71 species with a light microscope and 20 species with SEM. In the study, the length, surface structures, number, distribution, and shape of Sensilla styloconica were analyzed. In the study in which plesiomorphic and apomorphic characters were given for the Pieridae family, it was stated that the species were very similar to each other and show slight differences, but comparative sensilla photographs were not given for the species. Evaluation results were mostly limited by the length, surface structures, number, distribution, and shape of Sensilla styloconica. Dismorphinae (Leptidea sinapis (Linnaeus, 1758)) is in a separate group from Pierinae with the 2-row arrangement of sensilla styloconica. The phylogeny of Papilionidae and the monophyletic group was considered uncertain.
In this study, sensilla styloconica images from previous studies and sensilla photographs of samples from Turkey were compared and discussed. The aim of this study is to examine the sensilla types in the Pieridae family and to reveal the similarities and differences between the taxa. For this purpose, we examined, defined and compared the morphology of proboscis sensilla of P. edusa and C. croceus using scanning electron microscopy (SEM).
Materıal and Methods
The specimens examined in this study are the collection samples of the second author and are kept in the Gazi University Zoology Museum. A total of 10 specimens belonging to two species were examined. The heads of some of the samples were cut off and imaged, and the proboscis of the other samples were separated from the head with the help of forceps. The proboscis was cleaned with an alcohol-moistened brush and photographed under the Olympus SZX7 stereomicroscope (SM). The cut and cleaned samples were air dried; and after drying, the hose was placed on the studs and gold plated in a Polaron SC 502 brand vacuum coating device. It was then examined with the JEOL JSM 6060 LV scanning electron microscope (SEM) and photographed. Then it was scanned under electron microscope (SEM) (JEOL JSM 6060 LV) at 10 kV. The study was produced from the master’s thesis of the first autho.
Pontia edusa (Türkiye, Antalya: Akseki, Çukurköy, Mahmutlu, 830 m, 19-V-2008 2 ex, leg.S. Seven Çalışkan; Türkiye, Adana, Kabasakal, 89 m, 28-IV-2008 leg. S. Seven Çalırkan 2 ex). Colias croceus (Türkiye, Gökçeada, 90 m, 20-V-2006 2ex., leg. S. S. Çalışkan & E. Çalışkan; Türkiye, Erzincan, Dutluca, 1182 m, 17-VII-2007 2ex., leg. S. S.Çalışkan; Türkiye, Balıkesir, Kaz Mountain, Hasanboguldu, 120 m, 06-VI-2005 2 ex., leg. S. S. Çalışkan.
Results
In Pieridae members, the proboscis is a tubular structure consisting of two long galeae (Figs.1a, 2a). The bodies of Pontia edusa and Colias croceus have typical sensillae commonly seen in Lepidopters (s.basiconica (sb), s.chaetica (sc) and s.styloconica (ss)).
Pontia edusa (Fabricius, [1777].
The proboscis forms 4-5 spirals in a curled state at rest (Figure 1b). The surface of the galea bears cuticular spines (Figures 1b, 1c). Cuticular spines, arranged parallel to each other on the dorsal of the galea, are scattered on the ventrally (Figure 1d). The dorsal legulae consists of finger-like, double rows of partially overlapping plates (Figure 1h). Sensilla are in regular rows. The sensilla basiconica of the outer proboscis surface, between the cuticular spines, consists of a short stylus and a long sensory cone with a perforated blunt tip (Figures 1e, 1f). Sensilla basiconica is arranged in an irregular, single row in the food canal. It consists of a long sensory cone resting on a bulging base (Figures 1h, 1ı). In the dorsal galea, there are many hair-shaped sensilla chaetica. They are arranged scattered on the surface of the galea (Figures 1c, 1d). Sensilla styloconicas, which is distributed over the distal part of the proboscis, consists of a short stylus with 5 spines and a peg embedded in the base (Figures 1g). The length of the peg is longer than the stylus’.
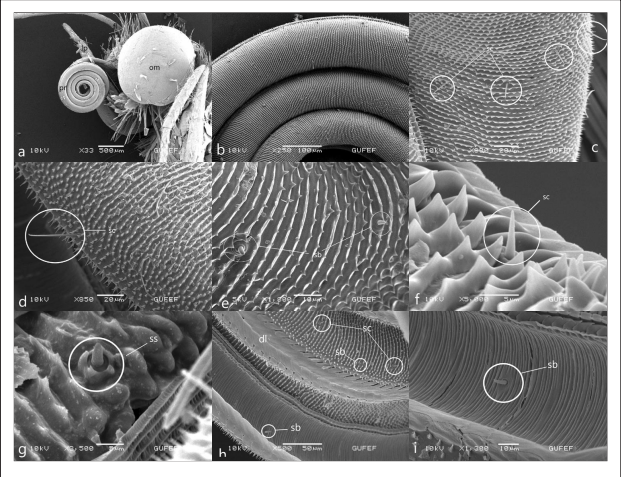
Figures 1.
Head and mouthparts of Pontia edusa, a. proboscis (pr), complex eye (om), labial palpus (lb) b. Coil count of proboscis c,d. galea surface and sensilla chaetica (sc) e,f. cuticuler spine on galea surface, external sensilla basiconica (sb) g. sensilla styloconica (ss) with 5 short spines h. dorsal surface of the galea and foodcanal on the inner galeal wall of the proboscis. i. Internal sensillum basiconicum in food canal (sb).
Colias croceus (Geoffroy, 1785, in Fourcroy.
The proboscis forms five rings at rest (Figure 2a). The surface of the galea bears cuticular spines (Figure 2b). These cuticular spines are arranged in parallel rows dorsally and independently from each other ventrally (Figures 2c, 2d). Sensilla are in regular rows. The external sensilla basiconica of the proboscis surface consists of a short stylus between the spines and a short sensory cone with a blunt tip (Figures 2e, 2g, 2j). The internal sensilla basiconica are arranged in a single row at irregular intervals in the food canal (Figures 2i, 2l). It consists of a short stylus and a long sensory cone (Figure 2k). Sensilla chaetica of varying lengths are numerous and are distributed over the surface of the galea (Figures 2c, 2e, 2f). Sensilla styloconica is concentrated in the distal part of the galea (Figure 2g). It consists of a relatively long five finger-shaped ribs stylus and a peg (Figures 2h). The length of the peg is close to the length of the stylus spines or may be slightly shorter.
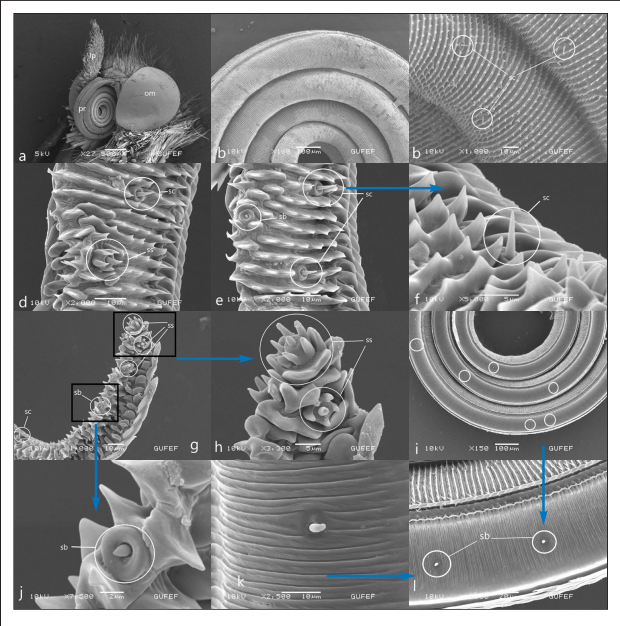
Figures 2.
Head and mouthparts of Colias croceus, a. proboscis (pr), complex eye (om), labial palpus (lb) b. Coil count of proboscis c. galea surface and sensilla chaetica (sc) d,e,f. Cuticular spines on galea surface, external sensilla basiconica (sb), sensilla styloconica (ss) sensilla chaetica (sc) g, h, j. distal part of the galea, sensilla chaetica (sc), sensilla basiconica, sensilla styloconica (ss), star-shaped sensilla styloconica with 5 long spines i. Distribution of internal s.basiconica in the food canal k, l. Sensillum basiconicum (sb) of the food canal.
Discussion
The surface of the galea differs between families. In many species, the outer surface of the galea may have distinct ribs, cuticular spines, hair-like cuticular processes, or scales that may appear convex (Krenn, 1990; Krenn & Kristensen, 2000; Paulus & Krenn, 1996; Speidel et al. 1996). Vertical, often branched galea wall ridges, which are densely covered with triangular cuticle spines all around, are the characteristics of all Pieridae (Paulus & Krenn, 1996). In P. edusa and C. croceus, the outer surface of the galeal wall is similar. Both have parallel outstretched cuticular spines.
Sensilla chaetica, which has nutritional localizations and mechanical functions, is found throughout the proboscis, but is longer in the proximal region than in other regions (Faucheux, 2013; Xue et al. 2016). In both studied species, sensilla chaetica is distributed on the outer surface of the proboscis and is more abundant proximally. Sensilla chaetica of P.edusa is significantly longer than that of C. croceus. Surrounded by a flexible shallow slot, sensilla basiconica has a sensory cone in the form of a dome or peg of varying lengths (Ma et al. 2019). It is of two types, inner and outer sensilla basiconica. The outer sensilla basiconica forms a single line on the entire surface of the galea, while the inner sensilla basiconica forms a single line in the digestive tract (Zhang et al. 2021). The external sensilla basiconica consists of a short sylus and a blunt-tipped sensory cone in C. croceus, whereas P. edusa consists of a short sylus and a long cone. The morphology and size of the inner sensilla are similar to that of the outer sensilla, but are usually blunt tipped (Fauchex, 2013). In C. croceus, s.basiconica, consisting of a short stylus and a long cone, are arranged in a single row in the food canal, but are not evenly spaced. On the other hand, it consists of a long sensory cone resting on a bulging base and is arranged in a single row (middle and side) irregularly in P. edusa. Internal sensilla basiconica is longer and less numerous than the external sensilla basiconica (Ma et al. 2019). Similarly, the internal sensilla basiconica are longer than the external sensilla basiconica in both studied species.
The sensilla styloconica in various forms are the characteristic sensilla of the Lepidopteran proboscis (Börner, 1939; Sellier, 1975; Altner & Altner, 1986). In some groups, the stylus is smooth and ellipsoid, or longitudinally spine-like cuticular ribs and is star-shaped. Some species have pale spines around the sensory cone, some have serrated ribs, some have nothing at all (Faucheux, 1991; Walters et al. 1998; Krenn, 1998; Faucheux, 1999; Krenn & Kristensen, 2000; Krenn et al. 2001; Kvello et al. 2006; Paulus & Krenn, 1996; Petr & Stewart, 2004). It is thought that sensilla styloconica can be used as a tool to distinguish taxa from each other in the lepidopteran systematics (Ma et al. 2019).
The Papilionidae and Pieridae are relatively uniform, and each can be clearly defined as a monophyletic group (Paulus & Krenn, 1996). The similarity between the sensilla styloconica of Pieridae members and Papilioninae members supports their closeness.
Pieridae is divided into three subfamilies. The slender, short, and very few sensilla styloconica in the family Pieridae have 4-5 strongly protruding ribs (Paulus & Krenn, 1996).
Paulus & Krenn (1996) stated that there were minor differences between the sensilla of the Pieridae species in their study, and only the distribution of the sensilla styloconica in Leptidea sinapis was different. Due to the difference in the distribution of sensilla, Dismorphiinae is given as a separate branch from Pierinae.
Pierinae and Coliadinae are closer to each other compared to Dismorphiinae due to the similar distribution of sensilla styloconica. Differences were detected in the sensilla of the two species examined in our study. In both cases, s.styloconica is of the excoronate type with 5 protruding short pegs and a smooth stylus. The sensilla of Colias croceus are similar to the sensilla of the Colias fieldii species in Ma et al. (2019) study. In Colias croceus, s.styloconica is not embedded in the pedestal. The ribs on the peg are more prominent, the spines on the ribs are longer, and the stylus is shorter than the peg.
The sensilla styloconica of Pontia edusa was compared and found to be similar to the sensilla styloconica of A. crataegi (Candan & Koçakoğlu, 2021) and P. rapae (Candan et al. 2018, unpublished). The common feature of all three species is that the ribs are very slightly convex and relatively short in length. In Pontia edusa, the sensilla styloconica consists of a short piece of nail embedded in the base and a relatively long piece of stylus.
The results support the hypothesis that the differences seen in the sensilla of members of different categories may be in the subfamily category. Comparative studies with more species are needed to reach a definitive conclusion. These morphological results likely suggest that the proboscis and related sensilla may provide potential value for systematic studies and phylogenetic analysis at the subfamily level within Pieridae.
Acknowledgments
We would like to thank Prof. Selami Candan and Doç. Dr. Nurcan Koçakog˘lu and for SEM photograph.
References
Atner, H., & Altner, I. (1986). Sensilla with both, terminal pore and wall pores on the proboscis of the moth, Rhodogastria bubo Walker (Lepidoptera: Arctiidae). Zoologischer Anzeiger, 216, 129-150.
Bourgogne, J. (1951). Ordre des Lépidoptères, In P.-P. Grassé editor. Traité de Zoologie, tome X. Insectes supérieurs et Hémiptéroïdes (pp. 174-448). Masson et Cie.
Börner, C. (1939). Die Grundlagen meines Lepidopterensystems. Verhandlungen VII. Internationaler Kongres für Entomologie, Berlin 1938 (Vol. 2, pp. 1372-1424). Druck Von G. Uschmann.
Büttiker, W., Krenn, H., & Putterill, J. F. (1996). The proboscis of eye-frequenting and piercing Lepidoptera (Insecta). Zoomorphology, 116(2), 77-83. https://doi.org/10.1007/BF02526872
Candan. S., & Koçakoğlu, N. Ö. (2022). The Morphology of the Sensilla on the Proboscis of Aporia crataegi (Linnaeus, 1758) (Lepidoptera: Pieridae). Commagene Journal of Biology, 6(2), 184-189. https://doi.org/10.31594/commagene.1187494
Candan, S., Çalışkan, S. S., & Koçakoğlu, N. Ö. (2018). Morphology and Ecology of Large Cabbage White (Pieris brassicae Linnaeus, 1758) (Lepidoptera: Pieridae). IX International Agriculture Symposium AGROSYM 2018, Jahorina, 4-7 October 2018, Bosnia and Herzegovina.
Charlanes, M., & Gaumont, R. (1960). La coaptation des pièces buccales de quelques Lépidoptères. Comptes Rendus de l’Académie des Sciences, Paris, 250(D), 1345-1346.
Gilbert, L. E. (1972). Pollen feeding and reproductive biology of Heliconius butterflies. Proceedings of the National Academy of Sciences, 69(6), 1403-1407. https://doi.org/10.1073/pnas.69.6.1403
Faucheux, M. J. (1991). Morphology and distribution of sensilla on the cephalic appendages, tarsi and ovipositor of the European sunflower moth, Homoeosoma nebulella Den. & Schiff. (Lepidoptera: Pyralidae). International Journal of Insect Morphology and Embryology, 20, 291-307. https://doi.org/10.1016/0020-7322(91)90018-5
Faucheux, M. J. (1999). Biodiversité et unité des organes sensoriels des Insectes Lépidoptères. Nantes: Société des Sciences Naturelles de l’Ouest de la France, 296 p. In French; full English summaries of all chapters and captions.20.
Faucheux, M. J. (2013). Sensillum types on the proboscis of the Lepidoptera: a review. Annales de la Société Entomologique de France, 49(1), 73-90. https://doi.org/10.1080/00379271.2013.767514
Krenn, H. W. (1990). Functional morphology and movements of the proboscis of Lepidoptera (Insecta). Zoomorphology, 110, 105-114. https://doi.org/10.1007/BF01632816
Krenn, H. W. (1998). Proboscis sensilla in Vanessa cardui (Nymphalidae, Lepidoptera): functional morphology and significance in flowerprobing. Zoomorphology, 118, 23-30. https://doi.org/10.1007/s004350050053
Krenn, H. W. (2010). Feeding mechanisms of adult Lepidoptera: structure, function, and evolution of the mouthparts. Annual Review of Entomology, 55(1), 307-327. https://doi.org/10.1146/annurev-ento-112408085338
Krenn, H. W., & Kristensen, N. P. (2000). Early evolution of the proboscis of Lepidoptera (Insecta): external morphology of the galea in basal glossatan moth lineages, with remarks on the origin of the pilifers. Zoologischer Anzeiger, 239, 179-196.
Krenn H. W., Zulka K. P., & Gatschnegg, T. (2001). Proboscis morphology and food preferences in nymphalid butterflies (Lepidoptera: Nymphalidae). Journal of Zoology, 254, 17-26. https://doi.org/10.1017/S0952836901000528
Kvello, P., Almaas, T. J., & Mustaparta, H. A. (2006). Confined taste area in a lepidopteran brain. Arthropod Structure & Development, 35(1), 35-45. https://doi.org/10.1016/j.asd.2005.10.003
Ma, L., Hu, K., Li, P., Liu, J., & Yuan, X. (2019). Ultrastructure of the proboscis sensilla of ten species of butterflies (Insecta: Lepidoptera). Plos One, 14(3), e0214658. https://doi.org/10.1371/journal.pone.0214658
Molleman, F., Krenn, H. W., Alphen, M. E. V., Brakefield, P. M, Devries, P. J., & Zwaan, B. J. (2005). Food intake of fruit-feeding butterflies: evidence for adaptive variation in proboscis morphology. Biological Journal of the Linnean Society, 86(3), 333-343. https://doi.org/10.1111/j.1095-8312.2005.00533.x
Monaenkova, D., Lehnert, M. S., Andrukh, T, Beard, C. E., Rubin, B., Tokarev, A., Wah-Keat Lee, Peter, H. A., & Konstantin, G. K. (2012). Butterfly proboscis: combining a drinking straw with a nanosponge facilitated diversification of feeding habits. Journal of the Royal Society Interface, 9(69), 720-726. https://doi.org/10.1098/rsif.2011.0392
Paulus, H. F., & Krenn, H. W. (1996). Vergleichende morphologie des schmetterlingsrüssels und seiner sensillen-ein beitrag zur phylogenetischen systematik der Papilionoidea (Insecta, Lepidoptera). Journal of Zoological Systematics and Evolutionary Research, 34(4), 203-216. https://doi.org/10.1111/j.1439-0469.1996.tb00826.x
Petr, D., & Stewart, K. W. (2004). Comparative morphology of sensilla styloconica on the proboscis of North American Nymphalidae and other selected taxa (Lepidoptera): systematic and ecological considerations. Transactions of the American Entomological Society, 130, 293-409.
Sellier, R. (1975). Etude ultrastructurale en microscopie electronique par balayage des organes sensoriels de la trompe des Iepidopteres Rhopalocires. Alexanor, 9, 9-15.
Speidel, W., Fanger, H., & Naumann, C. M. (1996). The phylogeny of the Noctuidae (Lepidoptera). Systematic Entomology, 21, 219-251. https://doi.org/10.1046/j.1365-3113.1996.d01-14.x
Walters, B. D., Albert, P. J., & Zacharuk, R. Y. (1998). Morphology and ultrastructure of sensilla on the proboscis of the adult spruce budworm, Choristoneura fumiferana (Clem.) (Lepidoptera: Tortricidae). Canadian Journal of Zoology, 76, 466-479. https://doi.org/10.1139/z97-218
Xue, S., Hu, Y.-Q., & Hua, B.-Z. (2016). Morphological comparison of proboscis sensilla between Plusiinae and Noctuinae (Lepidoptera: Noctuidae). Zoologischer Anzeiger, 263, 74-83. https://doi.org/10.1016/j.jcz.2016.05.002
Zhang, C.-M., Niu, Y., Hu, G.-L., & Lu, J.-Q. (2021). Ultramorphological Comparison of Proboscis and Associated Sensilla of Scotogramma trifolii and Protoschinia scutosa (Lepidoptera: Noctuidae). Insects, 12(11), 992. https://doi.org/10.3390/insects12110992
Zaspel, J. M., Coy, S., Habanek, K., & Weller, S. J. (2013). Presence and distribution of sensory structures on the mouthparts of self-medicating moths. Zoologischer Anzeiger, 253(1), 6-10. https://doi.org/10.1016/j.jcz.2013.06.001
Author notes
* Autor para la correspondencia / Corresponding author: selma@gazi.edu.tr, selmaseven@gmail.com