ARTÍCULOS
AGGRESSIVE BEHAVIOR OF PREGNANT FEMALES OF Akodon azarae: A STRATEGY TO REDUCE INFANTICIDE RISK OF THEIR YOUNGS NEXT TO BE BORN?
AGGRESSIVE BEHAVIOR OF PREGNANT FEMALES OF Akodon azarae: A STRATEGY TO REDUCE INFANTICIDE RISK OF THEIR YOUNGS NEXT TO BE BORN?
Mastozoología Neotropical, vol. 26, no. 2, pp. 255-262, 2019
Sociedad Argentina para el Estudio de los Mamíferos
Received: 13 February 2019
Accepted: 10 June 2019
Abstract: Infanticide by males is an adaptive behavioral strategy to improve the reproductive success of the perpetrator reproductive success of perpetrator by increasing their chances to reproduce with the victim female by shortening the inter-birth interval. Thus, females are expected to evolve different counterstrategies towards infanticide by males. We study nest site defense, or future offspring protection, through the direct attack of pregnant females close to give birth against potentially infanticidal males in the Pampean grassland mouse (Akodon azarae). We experimentally test the prediction that pregnant females are more aggressive against unfamiliar intruder males than non-pregnant females, and that the intensity of this aggressive response is independent of females residence durations in their home territory. We conducted 46 behavioral trials between resident females (pregnant or non-pregnant) and unfamiliar intruder males, considering different residence durations of females in their own individual enclosure (48 and 72 hours). We found that pregnant females were always more aggressive than non-pregnant females, independently of residence duration. Our research provides evidence that aggressive behavior of future mothers of A. azarae to defend the nest site would reduce infanticide risk of their youngs next to be born; keeping away potential infanticidal males could reduce the greater cost of avoiding infanticide of their vulnerable offspring (i.e., when they become lactating females). In this way, even though aggression is risky, it would be advantageous for pregnant females to use aggression even when there are not yet any pups to protect.
Keywords: aggressive behavior, pampean grassland mouse, reproductive strategies, resident-intruder test, sexual conflict.
Resumen: Comportamiento agresivo de las hembras preñadas de Akodonazarae: ¿una estrategia para reducir el riesgo de infanticidio de sus crías próximas a nacer? El infanticidio por parte de machos es una estrategia adaptativa que incrementa el éxito reproductivo del agresor al aumentar sus posibilidades de reproducirse prontamente con la víctima. Así, se espera que en las hembras evolucionen contraestrategias hacia el infanticidio por parte de machos. Nosotros estudiamos la defensa de nidos, o protección de futuras crías, a través de la agresión de hembras preñadas, próximas a parir, hacia machos potencialmente infanticidas en el ratón de pastizal Pampeano (Akodon azarae). Probamos experimentalmente la predicción que las hembras preñadas son más agresivas hacia los machos intrusos que las no preñadas, y que la intensidad de esta respuesta agresiva es independiente del tiempo de residencia de las hembras en sus clausuras individuales. Realizamos 46 enfrentamientos comportamentales entre hembras residentes (preñadas y no preñadas) y machos intrusos desconocidos, considerando el tiempo de residencia de las hembras (48 y 72 horas). Independientemente de la duración de la residencia de las hembras, las preñadas siempre fueron más agresivas que las no preñadas. Nuestra investigación proporciona evidencia de que el comportamiento agresivo de las futuras madres de A. azarae para defender el sitio del nido reduciría el riesgo de infanticidio de sus crías próximas a nacer; mantener alejados a potenciales machos infanticidas podría reducir el mayor costo de evitar el infanticidio de sus crías vulnerables (es decir, cuando son lactantes). De esta manera, aunque la agresión es riesgosa, sería ventajoso para las futuras madres usar la agresión incluso cuando todavía no hay crías para proteger.
Palabras clave: comportamiento agresivo, conflicto sexual, estrategias reproductivas, prueba de intrusoresidentes, ratón de pastizal pampeano.
INTRODUCTION
Infanticide or the killing of infants by conspecific is an adaptive behavior that increases individual fitness of the perpetrator and constitutes a prominent example of the evolutionary conflict between the reproductive interests of males and females (Hrdy 1979; Parker 1979; Ebensperger 1998). Although infanticide is widely accepted as an adaptive strategy in both sexes (Ebensperger & Blumstein 2007), one of the earliest recognized forms of sexual conflict was infanticide by males, which imposes serious costs on female reproductive success, while increases the fitness of infanticidal males (Palombit 2015). Infanticide committed by males occurs in a wide variety of mammal taxa (Ebensperger et al. 2000; Ebensperger & Blumstein 2007; Opperbeck et al. 2012; Lukas & Huchard 2014). In vole and mice, in which polygyny and promiscuity are the predominating mating systems, fathers typically do not provide parental care, and adult conspecifics males can display a wide range of responses towards neonates such as indifference, avoidance or infanticide (Clutton-Brock 1989; Waterman 2007).
Considering that infanticide is costly to females whose offspring is lost, it should be a sufficient evolutionary force to cause behavioral or physiological adaptations against it (Agrell et al. 1998; Lukas & Huchard 2014; Palombit 2015). A great variety of mechanisms have been proposed as females counterstrategies to infanticide by males, including choosing a dominant male, multi-male mating, female aggression against potential perpetrators, and pregnancy interruption, among others (Bruce 1960; Parmigiani et al. 1988a; Agrell et al. 1998; Wolff & Peterson 1998; Wolff & Dunlap 2002; Solomon & Keane 2007; Lopuch & Matula 2008). Agrell et al. (1998) and Wolff & Peterson (1998) describe maternal aggression as the specific type of aggressive behavior exhibited by pregnant or nursing females in proximity of their pups or the nest, against unfamiliar intruder males. The frequency and intensity of female aggressive behavior typically increases during late gestation, peaks sharply after the pups are born (during lactation), and gradually decreases towards weaning (Ostermeyer 1983; Parmigiani et al. 1988a;b; Maestripieri 1992; Koskela et al. 1997). In several small rodent species, keeping males far away from the nest site or young through aggressive behavior would be a common strategy against male infanticide exhibited by females (Wolff 1985; Maestripieri 1992).
While most experimental studies about female aggression against potentially infanticidal males have been performed in laboratory conditions (Huck et al. 1982; Mclean 1983; Parmigiani et al. 1989; Wilson et al. 1993; Coulon et al. 1995; Ylönen et al. 1997; Suárez & Kravetz 2001; Coda et al. 2011), our study in Akodon azarae (Cricetidae: Sigmodontinae) was developed in semi-natural conditions. This species, commonly known as the Pampean grassland mouse, is the numerically dominant rodent species in the Pampean agrarian ecosystems of central Argentina (Gomez et al. 2015). Akodon azarae is a habitat specialist species (Cavia et al. 2005; Fraschina et al. 2012) that prefers to inhabit stable or relatively undisturbed habitats (Martínez et al. 2014; Coda et al. 2015), and is as good indicator of habitat quality in agricultural systems (Coda et al. 2015). This species has a polygynous mating system that operates through female defense, in which a minority of males in the population (40% ) monopolizes several fertile females leaving other males without access to them (Bonatto et al. 2012; 2013a). However, depending on reproductive females availability some excluded males can trespass reproductive area bounds and have access to the females (Bonatto 2013; Bonatto et al. 2013b). Based on the hypothesis that aggressive behavior of A. azarae pregnant females against unfamiliar males reduce infanticide risk of their young next to be born, the aim of this study was to test the prediction that pregnant females are more aggressive against males than non-pregnant females. On the other hand, because aggressive behavior of A. azarae females towards unfamiliar males would have evolved as an adaptive response against infanticide, the fundament of the value-asymmetry hypothesis proposed by Maynard Smith & Parker (1976) would not apply in this case. Thus, we also hypothesized that the aggression of future mothers will be independent of the period of residence in their territory. Hence, we predict that pregnant females housed into individual enclosures during a shorter residence period invest the same time in aggressive interactions towards unfamiliar males than those housed a longer residence period. In addition, we described the behavioral responses of males against pregnant and non-pregnant females.
MATERIALS AND METHODS
The study species
Akodon azarae is a small (adult average weight 25 g) and opportunistic omnivore rodent (Suárez & Bonaventura 2001). This species shows continuous activity, being mainly active during daytime and crepuscular hours (Priotto & Polop 1997). This species is found in a great variety of stable habitats in the agroecosystems, characterized by keeping remnant native flora and fauna, with high gramineous cover, including natural pastures, road borders, borders between cultivated fields or pastures, and railway banks (Busch et al. 1997). Akodon azarae populations turnover annually and the individual lifespan is about 12 months (Hodara et al. 2000). During the breeding season A. azarae reproductive males have larger home ranges than females that overlap with two or more home ranges of breeding females (Bonatto et al. 2012; Bonatto 2013). Both female and male of A. azarae reach sexual maturity between 52 and 60 days of age, with a mean weight close to 15.5 g in females and 16.5 g in males (Bonatto 2013). The reproductive period of this species begins in spring (mid-September) and finishes in autumn (May). Akodon azarae has a gestation length of 23 days, each female can produce a maximum of 4 litters, with a mean of 4.6 pups per litter, only females provides parental care and young are weaned at 15 days old (Bonatto 2013).
Sampling and housing
Between middle of December 2014 and beginning of January 2015 we collected sexually mature A. azarae along road borders of agricultural ecosystems in Chucul (64° 20’W, 32°21’S), Río Cuarto Department, Córdoba Province (Argentina), with Sherman-type live traps (23 x 8 x 9.5 cm). Animals were weighed and sexed in the field and taken to the GIEPCO laboratory located 30 km away from the capture site. Fifty females and 50 males were firstly housed individually in clear polycarbonate cages (29 x18 x 18 cm). Because females could have mated in the wild population, during 23 days we daily checked the shape of their bellies for the purpose of recording evidence of pregnancy. In this study none of the females became pregnant in the wild. While 25 females were mated with 25 males in opaque polycarbonate reproductive cages, 25 females and 25 males remained individually housed in opaque polycarbonate cages. Each group (couples and single males and females) were located in different rooms. Opaque cages ensured that animals were visually isolated from their immediate neighbors. Individuals were maintained at 21°C on a 16:8 light/dark photoperiod. Rodent Purina laboratory chow and water were provided ad libitum, and sunflower and maize seeds were provided as weekly supplements. Dry wood shavings of Pinus elliottii and strands of cotton were provided as bedding material. For the purpose of recording the occurrence of pregnancy in mated females we daily checked the shape of their belly. As soon as pregnancy was evident (14 days, approximately), siring males were removed from the reproductive cages and individually housed in males room under the same condition described above. Besides, each siring male were ear-tagged for posterior identification in order to ensure that pregnant females did not meet their reproductive partners in the behavioral trials.
Of the 25 females mated in the laboratory 21 became pregnant. Thus, 21 females with 16-17 days of pregnancy, approximately, and 25 non-pregnant females, were released into an individual outdoor enclosure. The forty-six males of A. azarae remained in the laboratory until the moment they were used in the behavioral trials. All the females and males were used only once.
Study design
We studied aggressive behavior of A. azarae females in 2 forty-six round individual enclosures of 0.79 m each one, placed in the Espinal Reservation in the National University of Río Cuarto Campus (64°14’W , 33°07’S), in Córdoba province, Argentina. The individual enclosures are arranged in two parallel and interspersed rows, separated 2 from each other by 4 m, in a fenced area of 500 m . Each enclosure was limited by a concrete circle of 1 m diameter and 0.5 m high, with an open bottom which allows individuals to scent ground odours. Due to the fact that the height of 0.5 m allowed individuals to escape from the enclosure, we covered each of them with an iron mesh. In this study we considered the enclosures as territories (Bonatto et al. 2013a;b; 2017). Each enclosure was provided with water, rodent Purina laboratory chow, and sunflower and maize seeds ad libitum. In addition, to provide refuge, one open and locked trap, similar to a Sherman live trap, was located within each enclosure. In all cases females were placed into their territories with the wood shavings and strands of cotton from their respective housing cages. According to Thomas (2002), Hurst & Beynon (2004) and Hurst (2009), scent marks allow obtaining specific information about characteristics of conspecific individuals. We assumed that the scent from urine and feces left by females during the residence period, plus the bedding material, would provide enough evidence to intruder males of female reproductive condition.
To test our predictions we recorded pregnant and nonpregnant female aggressive response toward unfamiliar intruder males in relation to their reproductive condition (RC) and residence duration in their respective territories (RD). In the Resident-Intruder paradigm an animal is allowed to establish a territory (the resident), subsequently another animal is placed into the resident territory and then the two animals are allowed to interact with each other for a fixed period of time (Kraak 2012). Considering the residentintruder paradigm we established different residence times in both pregnant and non-pregnant females. In this study females always registered the resident condition. The distribution of pregnant females between the different residence times into the territories was performed taking into account the probable date of delivery. Thus, 12 and 13 pregnant and non-pregnant females, respectively, were placed individually into 25 different enclosures 48 h prior to the behavioral trials (RD1); and 9 and 12 pregnant and non-pregnant females, respectively, were housed into 21 different enclosures 72 h prior to the trials (RD2).
Behavioral testing
During early and middle February 2015, we performed 46 behavioral trials: 21 with pregnant females (12 RD1 and 9 RD2) and 25 with non-pregnant females (13 RD1 and 12 RD2). To perform behavioral trials, we placed into the territories a movable polycarbonate circular opaque arena (COA), 70 cm high (20 cm higher than the enclosure border), 219.8 cm perimeter and 70 cm diameter, with an open ground area (Bonatto et al. 2013a;b; 2017) (Fig. 1). This open area allowed animals to scent conspecifics odours. A removable opaque partition was placed across the centre of the COA at the beginning of each trial, and animals (resident female and intruder male) were placed simultaneously on either side of the partition for a 1-minute acclimatization period. After this period, the separator was carefully removed so that interactions between opponents could occur, and then the behavioral trials started. Before the trial, to identify the opponents, one of them was marked on its forehead with an odorless yellow water color highlighter. This kind of marker does not affect individual behavior in small rodent species (Bonatto et al. 2013a;b; 2017). Trials were performed during one of the activity pick of the Pampean grassland mouse, between 09:00 and 11:00 hours (Priotto & Polop 1997). Observations lasted 5 minutes and were recorded using a full high definition (1080i) video camera. A tripod to stabilize the video camera to prevent blurred images was added. We measured each behavior per trial and for each opponent as duration (in seconds) of a determined behavior along the 5 minutes (300 seconds was the maximum duration value for a given behavior). Aggressive behaviors observed during trials were described according to criteria proposed by Bonatto et al. (2013a;b; 2017) (Table 1). Besides, we also recorded submissive, cautious, amicable and non-interactive behaviors (Table 1).
At the end of this behavioral study all animals were taken to the laboratory. Juveniles born in captivity and adults male and female were kept in the laboratory in order to be used in other study. Our research protocol was approved by the Ethics Committee of Animal Research of Universidad Nacional de Río Cuarto, Argentina. During the study, animals were treated in humane manner according to current Argentinean Laws (National Law 14346).
Statistical Analyses
We used a generalized linear model (GLM) approach to examine the effects of reproductive condition of female (fixed factor with two levels: pregnant or non-pregnant female) and residence duration (fixed factor with two levels: RD1 or RD2) on time invested in aggressive interactions by the females (response variable). During this procedure, we fit time invested by females in aggressive interactions to a negative binomial distribution (with variance greater than the mean). Statistical analyses were carried out using the R software, version 3.2.2, MASS library (R Core Team 2015).

RESULTS
As pursuits never ended in injuries, we never had to interrupt the behavioral trials before the time limit. Thus, we analyzed 460 minutes of filming of behaviors exhibited by individuals. On average, pregnant females exhibited aggressive behavior towards males for more than the third part of each trial (127.67±37.79 s). Contrarily, this behavior was practically not exhibited by non-pregnant females during behavioral trials (0.60±2.20 s). The result of GLM for aggressive behavior showed that only female reproductive condition was statistically significant. Thus, female residence duration into their respective territories was not statistically significant. Pregnant females were always more aggressive towards males than non-pregnant females (β(SE) = 4.48 (0.43); z=10.37; p < 0.05) (Fig. 2), displaying aggressive approach, aggressive posture and pursuit; these behaviors were observed in 100%, 90.48% and 71.43% of behavioral trials, respectively. Fighting, the only aggressive behavior registered in this study that includes the two opponents, was never observed during encounters.

Fig. 1
Movable polycarbonate Circular Opaque Arena (COA).
Submissive behavior was not exhibited by females against males, independently of their reproductive condition and residence duration (RD1and RD2) (Fig. 2). In relation to caution behavior, only pregnant females remained vigilant in presence of nonsiring males (Fig. 2). Amicable behavior towards males was exhibited only by non-pregnant females (60.20±18.63 s) (Fig. 2). Respect to non interactive behaviors both pregnant and non-pregnant females exhibited them during the behavioral encounters. However, non-pregnant females exhibited both exploratory and self-grooming behaviors much more frequently than pregnant females (237.00±18.40 s and 111.62±45.75 s, respectively) (Fig. 2).
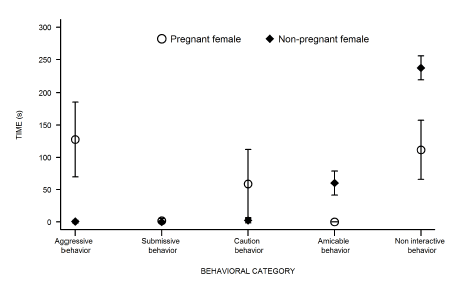
Fig. 2.
Mean (± SE) duration (in seconds) of behavioral response of resident females against intruder males for each behavior per female reproductive condition (pregnant or non-pregnant female) in intersexual trials of Akodon azarae.
In relation to the behavioral response of A. azarae males against females, aggressive behavior was not exhibited neither in presence of pregnant nor of nonpregnant females. On the other hand, submissive and alert behaviors only were displayed in presence of pregnant females (106.38±57.11 s and 72.95±40.12 s, respectively). Within submissive behaviors, escape was the most commonly exhibited by males, and this behavioral response was in relation with aggressive approaches by females.
DISCUSSION
In this study we predicted that pregnant females of A. azarae perform direct aggressive behaviors towards non-siring males associated with nest site defense. Our results showed that pregnant females were always more aggressive against non-siring males than non-pregnant females. Besides, this behavioral dominance of expectant mothers was supported by the greater values of submissive behavior that exhibited the intruder males in their presence. Therefore, high levels of intersexual aggression by pregnant A. azarae females may be the primary defensive mechanism for keeping away potential infanticidal males from the nest site. In a laboratory study, Suárez (1996) and Suárez & Kravetz (2001), also observed aggressive behaviors exhibited by A. azarae pregnant females against males, and proposed that this behavior would favor the exclusive use of its nest areas, avoiding disturbances in nearby areas. The goal of Suárez & Kravetz (2001) study was to compare the social behavior of A. azarae during the breeding and non-breeding seasons and, unlike our study in which continuous behavioral records were obtained, these authors measured malefemale interactions through instantaneous sampling. Besides, while Suárez & Kravetz (2001) registered intersexual interactions without distinguishing between familiar and unfamiliar males, in our study -siring (familiar) and non-siring (unfamiliar) males were clearly identified.
Infanticide committed by males has been reported for numerous species of vole and mice, such as Clethrionomys glareolus (Ylönen et al. 1997; Klemme et al. 2007), Dicrostonys groenlandicus (Mallory & Brooks 1978), Microtus ochrogaster (Mahady & Wolff 2002), Myodes glareolus (Opperbeck et al. 2012), Peromyscus leucopus (Wolff & Cicirello 1991), and Calomys musculinus (Coda et al. 2011). The context in which infanticide committed by males occurs most frequently in these species supports the sexual selection hypothesis that proposed that males kill unrelated infants to gain early reproductive access to the mothers (Agrell et al. 1998; Ebensperger & Blumstein 2007; Opperbeck et al. 2012; Palombit 2015).
Female counterstrategies may include multiples behavioral responses like multi-male mating (Cicirello & Wolff 1991; Wolff & Cicirello 1991; Wolff & MacDonald 2004; Sommaro et al. 2015), avoiding infanticidal males by moving to another area without these type of individuals (Mclean 1983; Coulon et al. 1995), and choosing a dominant male (Horne & Ylönen 1996; Lopuch & Matula 2008). Even when Huck et al. (1982) proposed that choosing a dominant male is a common counterstrategy in small rodents, this would not have evolved in females of A. azarae, since in an experimental study Contreras et al. (2016) found that they were unable to discriminate between dominant and subordinates males. Regarding to mating with multiple males (uncertain paternity hypothesis), since A. azarae has been described as a polygynous species (Bonatto et al. 2012; 2013a), that counterstrategy neither would have evolved in this species. On the other hand, to our knowledge, the behavioral response of moving to another area in order to avoid infanticidal males has not been studied yet in A. azarae. Another strategy against infanticide in pregnant or nursing females consists in direct attacks against potential male perpetrators (Parmigiani et al. 1988a;b; Maestripieri 1992; Coda et al. 2011). Wolff (1998) and Ylönen & Horne (2002), claim that this represent an effective tactic to defend the young or the area around the nest even when pups are not born yet.
The value asymmetry hypothesis proposes that, often with less fighting abilities, residents almost invariably defeat challengers as a result of their greater investment and local experience in comparison with the challengers (Maynard Smith & Parker 1976; Maynard Smith 1979). However, in our study we assumed that A. azarae mothers aggressive behavior evolve as an antagonistic adaptation to avoid infanticidal males and is not time-dependent. According to this, residence duration would not have to affect the intensity of pregnant females behavioral response, but only the presence of a strange male in the vicinity of the nest site. Indeed, our results showed that residence duration of females into their respective territories did not affect pregnant females aggressive response towards unfamiliar intruder males. However, the absence of differences in female behavioral response in relation to both residence times could be due to methodological issues. Because our estimate of the time of pregnancy could have a bias of one or two days, we were concerned to double the period of short residence (2 days) to a long residence (4 days); if mothers of A. azarae had started to give birth just before or during intersexual trials, this would have strongly affected our behavioral tests.
In summary, our results support the predictions that, regardless of the residence times considered in this study, A. azarae pregnant females are more aggressive against intruder males than non-pregnant females. Aggressive behavior of A. azarae future mothers could reduce the greater cost of avoiding infanticide of their vulnerable offspring (i.e., when they become lactating females). In this way, even though aggression is risky, it would be advantageous for pregnant females to use aggression even when there are not yet any pups to protect. Due to the great relevance of A. azarae as good indicator of habitat quality in agricultural systems, deepening the knowledge on reproductive strategies of this species assumes particular significance.
Acknowledgements
We thank Simón Emmanuel Gutierrez Brida for his assistance in producing the English version. This research was made possible by grants from the Secretaría de Ciencia y Técnica (SECyT), Universidad Nacional de Río Cuarto.
Authors declare no conflict of interest.
References
Agrell, J., J.O. Wolff, & H Ylönen. 1998. Counter strategies to infanticide in mammals: costs and consequences. Oikos 83:507¬517. https://doi.org/10.2307/3546678
Bonatto, F. 2013. Sistema de apareamiento y efecto de la proporción de sexos sobre el comportamiento de espaciamiento en roedores: Akodon azarae como modelo comportamental. Tesis de Doctorado. Universidad Nacional de Río Cuarto, Córdoba, Argentina
Bonatto, F., D. Gomez, A. R. Steinmann, & J. Priotto. 2012. Mating strategies of Pampean mouse males. Animal Biology 62:381-396. https://doi.org/10.1163/157075612X634102
Bonatto, F., J. Coda, D. Gomez, J. Priotto, & A.r. Steinmann. 2013a. Inter-male aggression with regard to polygynous mating system in Pampean grassland mouse, Akodon azarae (Cricetidae: Sigmodontinae). Journal of Ethology 31:223-231. https://doi.org/10.1007/s10164-013-0370-4
Bonatto, F., J. Coda, J. Priotto, & A.r. Steinmann. 2013b. Interfemale territorial behavior in pampean Grassland mouse, Akodon azarae (Cricetidae: Sigmodontinae). Mastozoología Neotropical 20:367-372.
Bonatto, F., A. R. Steinmann, D. Gomez, & J. Priotto. 2015. Do polygynous males of Akodon azarae (Rodentia: Sigmodontinae) vary their mating tactics at low availability of females? Mammalia 79:159-168. https://doi.org/10.1515/mammalia-2013-0102
Bonatto, F., J. Priotto, J. Coda, & A. R. Steinmann. 2017. Female intrasexual territoriality and its potential adaptive significance: the Pampean grassland mouse as an ecological model species. Ethology 123:230-241. https://doi.org/10.1111/eth.12592
Bruce, H. M. 1960. A block to pregnancy in the mouse caused by proximity of strange male. Journal of Reproduction and Fertility 1:96-103. https://doi.org/10.1530/jrf.0.0010096
Busch, M., M. R. Alvarez, E. A. Cittadino, & F. O. Kravetz. 1997. Habitat selection and interspecific competition in rodents in Pampean agroecosystems. Mammalia 61:167-184. https://doi.org/10.1515/mamm.1997.61.2.167
Cavia, R., I. E. Gómez Villafañe, E. A. Cittadino, D. N. Bilenca, M. H. Miño, & M. Busch. 2005. Effects of cereal harvest on abundance and spatial distribution of the rodent Akodon azarae in central Argentina. Agriculture, Ecosystems and Environment 107:95-99. https://doi.org/10.1016/j.agee.2004.09.011
Cicirello, D. M. & J. O. Wolff. 1991. The effects of mating on infanticide and pup discrimination in white-footed mice. Behavioural Ecology and Sociobiology 26:275-279. https://doi.org/10.1007/BF00178320
Clutton-Brock, T. H. 1989. Mammalian mating systems. Proceedings of the Royal Society of London. Series B 236:339-372. https://doi.org/10.1098/rspb.1989.0027
Coda, J., J. Priotto, & A. R. Steinmann. 2011. Behavioral counterstrategies against infanticide in corn mouse females, Calomys musculinus. Mastozoología Neotropical 18:207-215.
Coda, J. A., D. Gomez, A. R. Steinmann, & J. Priotto. 2015. Small mammals in farm lands of Argentina: responses to organic and conventional farming. Agriculture, Ecosystems and Environment 211:17-23. https://doi.org/10.1016/j.agee.2015.05.007
Contreras, F., H. J. Escudero, F. Bonatto, J. Priotto, & A. R. Steinmann. 2016. Jerarquía de dominancia y elección de machos por parte de hembras en Akodon azarae (Cricetidae: Sigmodontinae): un estudio experimental. Mastozoología Neotropical 23:389-400.
Coulon, J., L. Graziani, D. Allainé, M. C. Bel, & S. Puderoux. 1995. Infanticide in the Alpine marmot (Marmota marmota). Ethology Ecology and Evolution 7:191-194. https://doi.org/10.1080/08927014.1995.9522965
Ebensperger, L. A. 1998. Strategies and counterstrategies to infanticide in mammals. Biological reviews 73:221-346. https://doi.org/10.1017/S0006323198005209
Ebensperger, L. A., & D. T. Blumstein. 2007. Nonparental infanticide. Rodent societies: an ecological and evolutionary perspective (J. O. Wolff & P. W. Sherman, eds.). University of Chicago Press, Chicago, Illinois.
Ebensperger, L. A., C. Botto-Mahan, & R. H. Tamarin. 2000. Nonparental infanticide in meadow voles, Microtus pennsylvanicus: The influence of nutritional benefits. Ethology Ecology and Evolution 12:149-160. https://doi.org/10.1080/08927014.2000.9522810
Fraschina, J., V. A. León, & M. Busch. 2012. Long-term variations in rodent abundance in a rural landscape of the Pampas, Argentina. Ecological Research 27:191-202. https://doi.org/10.1007/s11284-011-0888-2
Gomez, M. D., J. Coda, I. Simone, J. Martínez, F. Bonatto, A. R. Steinmann, & J. Priotto. 2015. Agricultural land-use intensity and its effects on small mammals in the central region of Argentina. Mammal Research 60:415-423. https://doi.org/10.1007/s13364-015-0245-x
Hodara, K., M. Busch, M. J. Kittlein, & F. O. Kravetz. 2000. Density-dependent habitat selection between maize cropfields and their borders in two rodent species (Akodon azarae and Calomys laucha) of Pampean agroecosystems. Evolutionary Ecology 14:571-593. https://doi.org/10.1023/A:1010823128530
Horne, T., & H. Ylönen. 1996. Female bank voles (Clethrionomys glareolus) prefer dominant males; but what if there is no choice. Behavioural Ecology and Sociobiology 38:401-405. https://doi.org/10.1007/s002650050257
Hrdy, S. B. 1979. Infanticide among mammals: a review, classification, and examination of the implications for the reproductive strategies of females. Ethology and Sociobiology1:13-40. https://doi.org/10.1016/0162-3095(79)90004-9
Huck, U. W., R. L. Soltis, & C. B. Coopersmith. 1982. Infanticide in male laboratory mice: effects of social status, prior sexual experience, and basis for discrimination between related and unrelated young. Animal Behaviour 30:1158-1165. https://doi.org/10.1016/S0003-3472(82)80206-6
Hurst, J. L. 2009. Female recognition and assessment of males through scent. Behavioural Brain Research 200:295-303. https://doi.org/10.1016/j.bbr.2008.12.020
Hurst, J. L., & R. J. Beynon. 2004. Scent wars: the chemobiology of competitive signalling in mice. BioEssays 26:1288-1298. https://doi.org/10.1002/bies.20147
Klemme, I., J. A. Eccard, & H. Ylönen. 2007. Why do female bank voles, Clethrionomys glareolus, mate multiply? Animal Behaviour 73:623-628. https://doi.org/10.1016/j.anbehav.2006.07.010
Koskela, E., T. Mappes, & H. Ylönen. 1997. Territorial behaviour and reproductive success of bank vole Clethrionomys glareolus females. Journal of Animal Ecology 66:341-349. https://doi.org/10.2307/5980
Kraak, L. W. 2012. The Resident-Intruder model: Evaluation of the possible factors and mechanisms that control the display of aggressive behavior in rodents. Ph.D. Thesis. Faculty of Veterinary Medicine, Utrecht University, Netherlands.
Lopuch, S., & B. Matula. 2008. Is there a relationship between dominance rank and condition in captive male bank voles, Clethrionomys glareolus? Acta Ethologica 11:1-5. https://doi.org/10.1007/s10211-007-0035-9
Lukas, D., & E. Huchard. 2014. The evolution of infanticide by males in mammalian societies. Science 346:841-844. https://doi.org/10.1126/science.1257226
Maestripieri, D. 1992: Functional aspects of maternal aggression in mammals. Canadian Journal of Zoology 70:1069-1077. https://doi.org/10.1139/z92-150
Mahady, S.j., & J.o. Wolff. 2002. A eld test of the Bruce effect in the monogamous prairie vole (Microtus ochrogaster). Behavioral Ecology and Sociobiology 52:31-37. https://doi.org/10.1007/s00265-002-0484-0
Mallory, F. F., & R. J. Brooks. 1978. Infanticide and other reproductive strategies in the collared lemming, Dicrostonyx groenlandicus. Nature 273:144-146. https://doi.org/10.1038/273144a0
Martínez, J. J., V. Millien, I. Simone, & J. Priotto. 2014. Ecological preference between generalist and specialist rodents: spatial and environmental correlates of phenotypic variation. Biological Journal of the Linnean Society 112:180-203. https://doi.org/10.1111/bij.12268
Maynard Smith, J. M. 1979. Game theory and the evolution of behaviour. Proceedings of the Royal Society of London. Series B 205:475-488. https://doi.org/10.1098/rspb.1979.0080
Maynard Smith, J. M., & G. A. Parker. 1976. The logic of asymmetric contests. Animal Behaviour 24:159-175. https://doi.org/10.1016/S0003-3472(76)80110-8
Mclean, W. J. 1983. Parental behaviour and killing of young in arctic ground squirrels. Animal Behaviour 31:32-44. https://doi.org/10.1016/S0003-3472(83)80171-7
Opperbeck, A., H. Ylönen, & I. Klemme. 2012. Infanticide and population growth in the bank vole (Myodes glareolus): the effect of male turnover and density. Ethology 118:178-186. https://doi.org/10.1111/j.1439-0310.2011.01998.x
Ostermeyer, M. 1983. Maternal aggression. Parental behaviour in rodents (R.W. Elwood, ed.). John Wiley and Sons Ltd., Chichester, New York.
Palombit, R. A. 2015. Infanticide as sexual conflict: coevolution of male strategies and female counterstrategies. Cold Spring Harbor Perspectives in Biology 7:a017640. https://doi.org/10.1101/cshperspect.a017640
Parker, G. A. 1979. Sexual selection and sexual conflict. Sexual Selection and Reproductive Competition in Insects (M.S. Blum & N.A. Blum, eds.). Academic Press. https://doi.org/10.1016/B978-0-12-108750-0.50010-0
Parmigiani, S., A. Sgoifo, & D. Mainardi. 1988a. Parental aggression displayed by female mice in relation to the sex, reproductive status and infanticidal potential of conspecific intruders. Monitore Zoologico Italiano 22:193-201.
Parmigiani, S., P. F. Brain, D. Mainardi, & V. Brunoni. 1988b. Different patterns of biting attack employed by lactating females mice (Mus domesticus) in encounters with male and female conspecific intruders. Journal of Comparative Psychology 102:287-293. https://doi.org/10.1037//0735-7036.102.3.287
Parmigiani, S., P. Palanza, & P. F. Brain. 1989. Intraspecific maternal aggression in the house mouse (Mus domesticus): a counterstrategy to infanticide by males? Ethology Ecology and Evolution 1:341-352. https://doi.org/10.1080/08927014.1989.9525504
Parmigiani, S., P. Palanza, D. Mainardi, & P. F. Brain. 1994. Infanticide and protection of young in house mouse (Mus domesticus): female and male strategies. Infanticide and parental care (S. Parmigiani & F.S. von Saal, eds.). Harwood Academic Publishers, Chur, Switzerlan.
Priotto, J., & J. Polop. 1997. Space and time use in syntopic populations of Akodon azarae and Calomys venustus (Rodentia, Muridae). Mammalian Biology 62:30-36.
R Development Core Team. 2015. R: a language and environment for statistical computing. Vienna, Austria. R Foundation for Statistical Computing. URL http://www.R-project.org
Solomon, N. G., & B. Keane. 2007. Reproductive strategies in female rodents. Rodent societies: An ecological and evolutionary perspective (J.O. Wolff & P.A. Sherman, eds.). University of Chicago Press, Chicago, Illinois.
Sommaro, L. V., M. B. Chiappero, N. S. Vera, J. A. Coda, J. W. Priotto, & A. R. Steinmann. 2015. Multiple paternity in a wild population of the corn mouse: its potential adaptive signifcance for females. Journal of Mammalogy 96:908-917. https://doi.org/10.1093/jmammal/gyv091
Suárez, O. V. 1996. Estrategias reproductivas y cuidado parental en Akodon azarae (Rodentia, Muridae). Tesis Doctoral. Universidad Nacional de Buenos Aires, Buenos Aires, Argentina.
Suárez, O., & S. M. Bonaventura. 2001. Habitat use and diet in sympatric species of rodents of the low Paraná delta, Argentina. Mammalia 65:167-176. https://doi.org/10.1515/mamm.2001.65.2.167
Suárez, O., & F. Kravetz. 2001. Male-female interaction during breeding and non-breeding seasons in Akodon azarae (Rodentia, Muridae). Iheringia Série Zoologia 91:171-176. https://doi.org/10.1590/S0073-47212001000200023
Thomas, S. A. 2002. Scent marking and mate choice in the prairie vole, Microtus ochrogaster. Animal Behaviour 63:1121-1127. https://doi.org/10.1006/anbe.2002.3014
Waterman, J. 2007. Male mating strategies in rodents. Rodent Societies: an Ecological and Evolutionary Perspective (J. O. Wolff & P.W. Sherman, eds.). University of Chicago Press, Chicago, Illinois.
Wilson, W. L., R. W. Elwood, & W. I. Montgomery. 1993. Infanticide and maternal defense in the wood mouse Apodemus sylvaticus. Ethology Ecology and Evolution 5:365-370. https://doi.org/10.1080/08927014.1993.9523024
Wolff, J. O. 1985. Maternal aggression as a deterrent to infanticide in Peromyscus leucopus and P. maniculatus. Animal Behaviour 33:117-123. https://doi.org/10.1016/S0003-3472(85)80125-1
Wolff, J. O. 1998. An offspring-defence hypothesis for territoriality in female mammals. Etholology Ecolology and Evolution 10:227239. https://doi.org/10.1080/08927014.1998.9522854
Wolff, J. O., & D. M. Cicirello. 1991. Comparative paternal and infanticidal behavior of sympatric white-footed mice (Peromyscus leucopus noveboracensis) and deermice (P. maniculatus nubiterrae). Behavioural Ecology 2:38-45. https://doi.org/10.1093/beheco/2.1.38
Wolff, J. O., & J. A. Peterson. 1998. An offspring-defence hypothesis for territoriality in female mammals. Ethology Ecology and Evolution 10:227-239. https://doi.org/10.1080/08927014.1998.9522854
Wolff, J. O., & A. S. Dunlap. 2002. Multi-male mating, probability of conception, and litter size in the prairie vole (Microtus ochrogaster). Behavioural Processes 58:105-110. https://doi.org/10.1016/S0376-6357(02)00022-0
Wolff, J. O., & D. W. MacDonald. 2004. Promiscuous females protect their offspring. Trends in Ecology and Evolution 19:127-134. https://doi.org/10.1016/j.tree.2003.12.009
Ylönen, H., & T. J. Horne. 2002. Infanticide and effectiveness of pup protection in bank voles: does the mother recognise a killer? Acta Ethologica 4:97-101. https://doi.org/10.1007/s10211-001-0055-9
Ylönen, H., E. Koskela, & T. Mappes. 1997. Infanticide in the bank vole (Clethrionomys glareolus): occurrence and the effect of familiarity on female infanticide. Annales Zoologici Fennici 31:259-266.
Additional information
Editor asociado: P. Taraborelli