SECCIÓN ESPECIAL - NOTA
UNA NUEVA ESPECIE DE Philander (MARSUPIALIA, DIDELPHIDAE) DEL MIOCENO TARDÍO DEL NOROESTE ARGENTINO
A new species of Philander (Marsupialia, Didelphidae) from the late Miocene of northwestern Argentina
UNA NUEVA ESPECIE DE Philander (MARSUPIALIA, DIDELPHIDAE) DEL MIOCENO TARDÍO DEL NOROESTE ARGENTINO
Mastozoología Neotropical, vol. 28, núm. 1, p. 540, 2021
Sociedad Argentina para el Estudio de los Mamíferos
Recepción: 28 Septiembre 2020
Aprobación: 18 Noviembre 2020
Resumen: Se describe una nueva especie de Philander (Marsupialia, Didelphimorphia, Didelphidae) procedente de niveles de la Formación Andalhuala (Mioceno tardío) de la localidad de Tio Punco, provincia de Tucumán (Argentina). Comparada con las especies vivientes del género, la nueva especie es de tamaño relativamente grande y sus premolares y molares son de aspecto robusto; el cráneo tiene las crestas frontales cortas y más transversales al eje craneano; la diastema entre los P1-2 es corta; el cíngulo labial del P3 es incompleto y vestigial; los molares superiores tienen los protoconos más robustos y la postmetacrista del M3 está menos desarrollada.
Palabras clave: Marsupialia, Didelphidae, Philander, Mioceno, Tucumán.
Abstract: A new species of Philander (Marsupialia, Didelphimorphia, Didelphidae), from levels of the Andalhuala Formation (late Miocene) at Tio Punco locality, Tucumán Province (Argentina) is described. Compared to the living species of the genus, the new species is relatively large and has stronger premolars and molars; the skull has short frontal crests which are more transversally placed regarding the cranial axis; the diastema between P1 and P2 is short; the labial cingulum of P3 is vestigial and incomplete; upper molars have more robust protocones, and the posmetacrista of M3 is less developed.
Keywords: Marsupialia, Didelphidae, Philander, Miocene, Tucumán.
Entre los Didelphidae (Metatheria, Marsupialia, Didelphimorphia), las especies de Philander Brisson, 1762 se cuentan entre las más pequeñas de la tribu Didelphini (240 a 600 gramos de masa corporal; Patton & da Silva 2008). Son similares a las especies de Didelphis en el aspecto general del cráneo y la morfología dentaria, aunque más carnívoras y arborícolas (Vieira & Astúa de Moraes 2003; pero véase Vieira & Camargo (2012) sobre la locomoción en estos marsupiales). Habitan todo tipo de ambientes tropicales y subtropicales forestados (incluyendo sistemas antropizados como plantaciones), desde el sudeste de México hasta el noreste de Argentina, con una mayor diversidad en la región periamazónica noroeste y oeste (este de Perú, sur y este de Venezuela, zonas bajas de Ecuador y Colombia, Guyanas; Voss et al. 2018). Algunas especies, como P. canus y P. quica, alcanzan una distribución más meridional (Fig. 1).
La sistemática de las especies vivientes de Philander ha sido objeto de varias revisiones parciales en las últimas décadas (Hershkovitz 1997; Patton & Da Silva 1997; Patton & da Silva 2008; Castro-Arellano et al. 2000; Gardner 2005; Lew et al. 2006; Flores et al. 2008; Chemisquy & Flores 2012; Voss et al. 2018; Voss & Giarla 2020). En la más reciente publicada, Voss & Giarla (2020) reconocieron nueve especies: P. andersoni Osgood, 1913, P. canus Osgood, 1913 (sinónimos junior: crucialis Thomas, 1923; mondolfiiLew et al., 2006; olrogiFlores et al., 2008), P. mcilhennyi Gardner y Patton, 1972, P. melanurus Thomas, 1899 (sinónimos junior: fuscogriseus Allen, 1900; grisescens Allen, 1901; melantho Thomas, 1923), P. opossum Linnaeus, 1758 (sinónimos junior: frenatus Olfers, 1818; superciliaris Olfers, 1818), P. pebas Voss et al., 2018, P. quica Temminck, 1824 (sinónimo junior: azaricus Thomas, 1923), P. vossi Gardner y Ramírez-Pulido, 2020 (sinónimo junior: pallidus Allen, 1901) y P. nigratusVoss y Giarla, 2020. Una décima especie sería P. deltaeLew et al., 2006, no evaluada por Voss et al. (2018) o Voss & Giarla (2020).
Muy distintos análisis coinciden en señalar las mayores afinidades de Philander con los representantes del género Didelphis Linnaeus, 1758 (e.g., Reig et al. 1987; Patton et al. 1996; Jansa & Voss 2000; Jansa et al. 2013; Voss & Jansa 2003, 2009). Algunos autores, como Simpson (1972) y Pine (1973), han señalado incluso que no existen argumentos de peso como para considerar a Philander más que un subgénero de aquél (pero véase Reig et al. 1987). Sobre las relaciones de estos géneros con otros didelfinos extintos y vivientes, distintos análisis filogenéticos coinciden en señalar que Lutreolina Thomas, 1910 constituye el grupo hermano de Didelphis + Philander (e.g., Voss & Jansa 2009). Goin & Pardiñas (1996) argumentaron en contra de las supuestas afinidades entre el extinto Thylophorops Reig, 1952 con Lutreolina. Por el contrario, Thylophorops podría constituir, junto con Didelphis y Philander, un conjunto de didelfinos estrechamente emparentados entre sí, siendo Lutreolina su grupo hermano (Goin & Pardiñas 1996:fig. 12).
El registro fósil de los representantes de Philander es escaso. Winge (1893) señaló la presencia de ejemplares fósiles de P. opossum en depósitos de cavernas de Lagoa Santa, Brasil, de edad Cuaternario lato sensu. Ferreira Lino et al. (1979) mencionaron la presencia de Philander sp. en niveles de caverna, de edad Pleistoceno tardío, en la región del Vale do Ribeira (Estado de San Pablo, Brasil). Más recientemente, varios ejemplares de Philander sp. fueron listados para los sitios arqueológicos con secuencias holocénicas de Garivaldinho y Sangao, en el sur de Brasil (Estado de Rio Grande do Sul; Hadler et al. 2009). Hasta el momento, el único taxón pre-Cuaternario referible a Philander es P. entrerianus Ameghino, 1899, de niveles “mesopotamienses” (Mioceno tardío) de los alrededores de Paraná, provincia de Entre Ríos, Argentina. La especie fue originalmente asignada al género Zygolestes (Ameghino 1898), quien también incluyó en el mismo a Z. paranensis (pero véase Goin 1997). Reig (1957) refirió entrerianus al género Philander. P. entrerianus era conocida solamente a partir de un único molar inferior; Goin et al. (2013) describieron un nuevo ejemplar referible a la misma (una mandíbula derecha con el p2 y la serie molar completa), confirmando su ubicación genérica.
En este trabajo se da a conocer, de manera concisa, una segunda especie de Philander para el Mioceno tardío de la Argentina, procedente de la provincia de Tucumán. Se deja para una futura contribución el análisis pormenorizado de sus afinidades, su significación en la evolución del clado Didelphis + Philander así como también la información que aporta desde el punto de vista paleoambiental y paleobiogeográfico.
Abreviaturas Institucionales
FCP-V-M, Fundación Casa del Pueblo, Firmat (Provincia de Santa Fe, Argentina); MACN, Colección Paleontología Vertebrados, Museo Argentino de Ciencias Naturales “Bernardino Rivadavia” (Buenos Aires, Argentina); PZVL, Colección Paleontología Vertebrados, Instituto Miguel Lillo, Tucumán (Provincia de Tucumán, Argentina). Otras abreviaturas: Ma, Megannum, un millón de años en la escala radioisotópica del tiempo; P, M, p, m, premolares y molares superiores e inferiores, respectivamente. Se asume la siguiente fórmula dentaria generalizada para los marsupiales: I/i 5/4, C/c 1/1, P/p 3/3, M/m 4/4. La terminología de las cúspides y crestas de los molares se detalla en Goin et al. 2016: fig. 1.2). Todas las medidas que aquí se brindan son en milímetros (mm).
Género Philander Brisson, 1762
Philander massoiai sp. nov.
(Figs. 2, 3B, 3D)
LSID: urn:lsid:zoobank.org:act:CF71BFFE-F511- 4423-AF28-F932436293E4
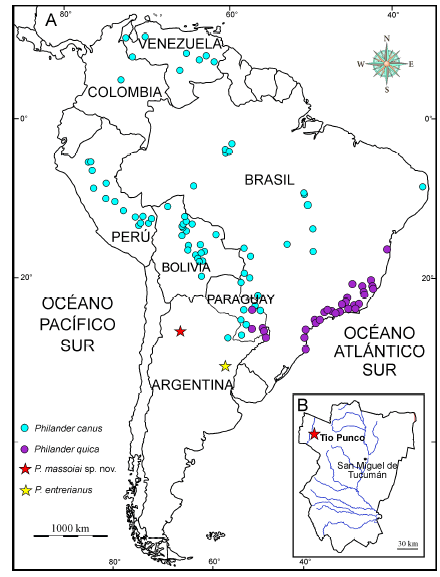
Fig. 1
A. Mapa de América del Sur indicando la localidad de procedencia del holotipo de Philander massoiai sp. nov. (estrella roja), del holotipo de P. entrerianus (estrella amarilla), como así también la distribución de dos de las especies vivientes del género: P. canus y P. quica. B. Detalle de la Provincia de Tucumán, Argentina, indicando la localidad tipo de Philander massoiai sp. nov., Tio Punco.
Holotipo
PZVL 3118, un fragmento de la porción dorsal del cráneo incluyendo parte del rostro y del techo craneano, una parte del hemimaxilar derecho con los alvéolos y raíces del P1 y los P2-M4 completos y varios huesos postcraneanos asociados, extremadamente fragmentarios; todos los restos presumiblemente pertenecientes al mismo individuo (Figs. 2, 3B,D).
Hipodigma
El holotipo solamente.
Distribución
Tio Punco (o Tiu Punco, o Tiopunco; Georgieff et al. 2014; Nasif et al. 2019), Departamento de Tafí del Valle, Provincia de Tucumán, Argentina. Más probablemente, Formación Andalhuala, Mioceno tardío (Messiniano; Edad-mamífero Huayqueriense). La localidad de Tio Punco se ubica en extremo noroeste de la Provincia de Tucumán, inmediatamente al este del río Santa María (Fig. 1B). Sus coordenadas geográficas son 26° 28’ S y 65° 57’ W.
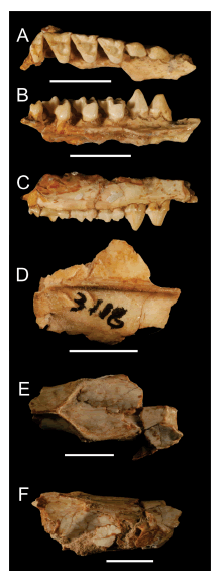
Fig. 2
Philander massoiai sp. nov. Ejemplar PZVL 3118 (holotipo): A-C. Hemimaxilar derecho en vistas oclusal (A), lingual (B) y lateral (C); D. Fragmento del techo craneano en vista dorsal; E-F, fragmento craneano a la altura de la constricción interorbital, en vistas dorsal (E) y lateral (F). El extremo rostral de los huesos se ubica en el margen derecho de la figura. Escala =10 mm.
Nota
El holotipo fue encontrado entre los materiales de la “Colección Peirano” del Instituto Miguel Lillo de Tucumán (Sección Paleozoología Vertebrados), aunque sin rótulo asociado. Sin embargo, el número de colección que presenta escrito sobre uno de los restos corresponde al de una serie de ejemplares colectados por Abel Peirano tanto en la localidad de Tio Punco como de niveles huayquerienses (Mioceno tardío) de Chiquimil, en la provincia de Catamarca. En una nota remitida al autor por las investigadoras de la Universidad Nacional de Tucumán Dras. Norma Nasif y Graciela Esteban se indica que el ejemplar (y otros de un mismo lote) fue coleccionado en Tio Punco, por lo que esta es considerada aquí la localidad tipo de la nueva especie. En el área de Tio Punco afloran niveles de edad Huayqueriense (Mioceno tardío) correspondientes a la Formación Andalhuala (Esteban & Abdala 1993; Esteban et al. 1997, 2014). Otros materiales colectados por Peirano en el área de Tio Punco también se corresponden con taxones de edad Huayqueriense procedentes de la Formación Andalhuala. Finalmente, consultas realizadas por N. Nasif y G. Esteban a baqueanos que acompañaron a Peirano en el área de Tio Punco permitieron confirmar que en las localidades específicas prospectadas por él afloran solamente sedimentos de la Fm. Andalhuala. En síntesis, si bien no es posible, con la información obtenida, precisar el sitio y niveles exactos de donde fue extraído el holotipo, el conjunto de evidencias disponibles sugiere que proviene de niveles de la Fm. Andalhuala (Mioceno tardío, Edad-mamífero Huayqueriense).
Geología, paleontología y edad
En el Noroeste argentino, los afloramientos correspondientes al Mioceno tardío (Tortoniano-Messiniano; 11.6-5.3 Ma) aflorantes en la Provincia de Catamarca y Tucumán son relativamente abundantes, siendo consecuencia de la depositación de sedimentos en valles intermontanos. En el noroeste de Tucumán afloran en el Valle de Santa María, componiendo precisamente el denominado Grupo Santa María. Dentro de este paquete sedimentario, la Fm. Andalhuala suprayace a la Fm. Chiquimil e infrayace a la Fm. Corral Quemado, siendo la más potente del grupo con 1 200 metros de espesor. Está constituida por limolitas, areniscas y conglomerados con niveles intercalados de tobas; el paleoambiente ha sido interpretado como perteneciente a un sistema fluvial entrelazado de baja sinuosidad (Georgieff et al. 2014). En diversas localidades fosilíferas de este valle (e.g., Entre Ríos) se ha exhumado una importante fauna de vertebrados referida a la edad Huayqueriense. Del área de Tio Punco se han recuperado diversos fósiles de mamíferos, en particular xenartros: dasipódidos de mediano tamaño (Chasicotatus, Paraeuphractus, Proeuphractus, Macroeuphractus), un pampaterino (Vassallia) y varios gliptodontes de tamaño grande (Eosclerocalyptus, Lomaphorus, Phlyctaenopyga, Pseudoeuryurus y Neothoracophorus; Nasif et al. 2019 y bibliografía ahí citada). La base de la Fm. Andalhuala fue datada en 6.02 Ma (Marshall & Patterson 1981); la edad del paquete sedimentario en esta área es referida al Messiniano (Mioceno tardío; Nasif et al. 2019).
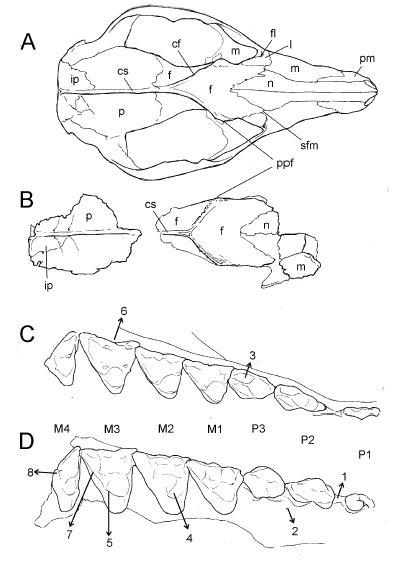
Fig. 3
A-B. Dibujos esquemáticos comparativos del cráneo de Philander canus (A) y de P. massoiai sp. nov. (B); C. Detalle del hemaxilar derecho del mismo ejemplar de P. canus mostrando la serie P1-M4 en vista oclusal; D, detalle del maxilar del holotipo de P. massoiai mostrando las raíces del P1 y la serie P2-M4 en vista oclusal. A y C corresponden al mismo individuo (AMNH 210413) figurado en Voss et al. (2018: figs. 12 B y E). Los dibujos esquemáticos no están en escala. Referencias: cf, cresta frontal; cs, cresta sagital; f, frontal; fl, foramen lacrimal; ip, interparietal; l, lacrimal; M1, M2, M3, M4, molares superiores; m, maxilar; n, nasal; P1, P2, P3, premolares superiores; p, parietal; pm, premaxilar; ppf, proceso postorbital del frontal; sfm, sutura frontomaxilar. Los números que se muestran en los esquemas C-D indican las diferencias más importantes entre la dentición de P. massoiai y la de las especies vivientes: 1, la diastema entre P1 y P2 es proporcionalmente menor en P. massoiai; 2, los premolares en esta especie son más robustos; 3, el cíngulo labial en el P3 de las especies vivientes está más desarrollado; 4, los protoconos en P. massoiai son más anchos y robustos; 5, en esta especie se aprecia el metacónulo en los M2-3; 6, el ectoflexo del M3 en las especies vivientes es más profundo; 7, la postmetacrista en P. massoiai es proporcionalmente más corta; 8, el metacono del M4 de esta especie es más reducido.
Diagnosis
Se diferencia del resto de las especies del género Philander por la siguiente combinación de caracteres: tamaño relativamente grande; cráneo con las crestas frontales cortas y orientadas más transversalmente al eje craneano; procesos postorbitales del frontal más gruesos y rugosos; premolares superiores robustos y anchos; diastema entre los P1-2 muy corto; P3 con el cíngulo labial incompleto y vestigial; molares superiores con los protoconos más anchos y robustos y la postmetacrista del M3, proporcionalmente, máscorta.
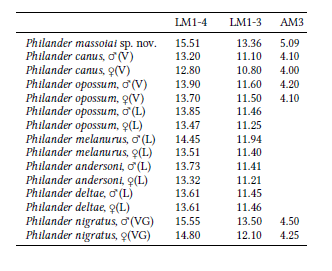
Medidas (en mm)
El hemimaxilar preservado en el holotipo (PZVL 3118) muestra las siguientes medidas dentarias (L, largo; A, Ancho): LP1-M4: 26.04*; LP1-P3: 11.07*; LM1-M4: 15.51; LM1-3: 13.36; LP2: 4.01; AP2: 2.38;LP3: 3.77; AP3: 2.43; LM1: 4.74; AM1: 3.94; LM2: 4.66;AM2: 4.50; LM3: 4.63; AM3: 5.09; LM4: 1.96; AM4:5.12. El asterisco en las medidas LP1-M4 y LP1-P3 indica que las mismas son aproximadas debido a la ausencia del P1 (se tomó, en lugar del extremo anterior de este diente, el borde mesial del alvéolo anterior). En la Tabla 1 se comparan tres medidas de P. massoiai con las homólogas de seis especies vivientes de Philander (véase Lew et al. 2006; Vosset al. 2018; Voss & Giarla 2020).
Descripción
El ejemplar PZVL 3118 ha conservado parte de la porción rostral incluyendo los nasales en su mitad posterior, la casi totalidad de los frontales y parte del hemimaxilar derecho (Figs. 2E, F, 3B). Por otra parte, existe un fragmento del techo craneano, del mismo individuo, conservando casi toda la porción dorsal de los parietales (Figs. 2D, 3B). Los nasales son de aspecto típicamente didelfino y alcanzan su ancho máximo cerca de la sutura fronto-maxilar. Hacia atrás se adelgazan progresivamente hasta terminar en punta, por lo que estos huesos conforman una figura romboidal en su parte posterior. En la cara lateral derecha del cráneo se ha preservado parte del lacrimal; se aprecia un único foramen lacrimal relativamente grande (tal como ocurre en varias especies de Philander; e.g., P. mondolfii) que abre no en el borde sino por fuera de la órbita. Los frontales tienen un aspecto rugoso a lo largo de casi todo el borde latero-dorsal de las órbitas (Fig. 2E, F), alcanzando la máxima rugosidad a la altura de los procesos postorbitales. Estos procesos son gruesos y robustos, mucho más que en los ejemplares observados de las especies vivientes de Philander, recordando más bien la condición presente en las especies de Didelphis. Sin embargo, este rasgo puede haber sido exagerado por el desgaste (¿por rodamiento?) que muestra el cráneo en esta región. A lo largo de su unión en la línea media dorsal del cráneo, por delante de las crestas, los frontales no son de aspecto plano sino que muestran una depresión longitudinal (Fig. 2E). Por detrás de los procesos postorbitales se desarrollan las crestas frontales (o “líneas temporales” en la terminología de Wible 2003), las que convergen para unirse en la cresta sagital más anteriormente que en la mayor parte de las especies vivientes del género, por lo que son comparativamente más cortas (esto es, son más transversales al eje craneano; Figs. 2E, 3A-B). El rasgo debe tomarse con cautela, ya que la orientación de las crestas varía según el desarrollo ontogenético de los individuos: son menos transversales al eje craneano en las formas juveniles y más transversales en los individuos adultos. De todos modos, cuando se comparan estas crestas con las de individuos adultos de las especies vivientes de Philander, queda claro que las de P. massoiai son más transversales que las de aquellas. Sólo en P. opossum y en P. andersoni esta configuración asemeja más a la de P. massoiai, si bien las crestas en estas especies se unen más posteriormente (véase, e.g., Lew et al. 2006; Flores et al. 2008; Voss et al. 2018; Voss & Giarla 2020). Esta condición también recuerda más a aquella presente en las especies de Didelphis que a la de las especies vivientes de Philander.
Los parietales son de aspecto liso y abovedado (Figs. 2D, 3B); en la zona medio-dorsal, cerca de la unión con el interparietal (o supraoccipital), se aprecia un par de surcos vasculares cortos y relativamente profundos a ambos lados de la cresta sagital. Un segundo par de surcos vasculares está presente también sobre el interparietal. Ambos pares de surcos son similares –aunque algo más cortos y profundos– a los observados en los ejemplares comparados, referibles a la especie viviente Philander canus. Posiblemente, estos surcos se correspondan con los forámenes emisarios descriptos para Monodelphis domestica por Wible (2003); los mismos constituyen aperturas de canales que conectan los senos intracraneanos con el sistema venoso extracraneano. Las crestas sagital y lambdoidea (o nucal según Wible 2003) alcanzan un desarrollo moderado, tal como ocurre en las especies vivientes de Philander (Fig. 2D).
El fragmento de hemimaxilar derecho que se ha preservado incluye las raíces del P1 y los P2-M4 completos (Figs. 2A-C,3D). El P3 y el M4 están completamente erupcionados y muestran poco desgaste, lo que sugiere que el ejemplar PZVL 3118 era un individuo adulto joven. El P1 muestra su raíz anterior algo más labial que la posterior, lo que sugiere que este premolar estaba oblicuamente orientado en la hilera dentaria. La diastema entre el P1 y el P2 es muy corta, más que en cualquiera de las especies vivientes de Philander. Los P2-3 son grandes y moderadamente anchos, comparativamente más que en P. canus. El P2 tiene cíngulos labial y lingual poco desarrollados en la mitad posterior del diente, los que culminan en una minúscula cúspide basal en su extremo posterior. Ambos cíngulos están menos desarrollados que en las especies vivientes del género (en P. nigratus, por ejemplo, el desarrollo de ambos cíngulos es casi completo alrededor del premolar). El P3 tiene el cíngulo labial incompleto y vestigial, menos desarrollado que en las especies vivientes (Fig. 2C); el cíngulo lingual, por su parte, tiene un moderado desarrollo (aunque menor al de P. canus, por ejemplo; Fig. 2B).
Los M1-3 tienen un aspecto didelfino generalizado y son más cortos y con las plataformas estilares más anchas que aquellos de las especies vivientes del género (Figs. 2A, 3D). En el M4 el metacono es menos saliente posteriormente que en P. canus, P. quica o P. pebas. A diferencia de las especies vivientes, la longitud y orientación de las postmetacristas varían muy poco del M1 al M3. Las preparacristas, por el contrario, aumentan sensiblemente de longitud del M1 al M4, conectándose labialmente en los dos primeros molares con la cúspide estilar B, mientras que en el M3 termina en la plataforma estilar sin conexión aparente. Los metaconos son claramente más grandes y altos que los paraconos (Fig. 2B). La cúspide estilar B es más pequeña que la D en el M1, algo más grande en el M2, muy reducida en el M3 y prácticamente ausente en el M4. La cúspide estilar D está bien desarrollada en los M1-2, es vestigial en el M3 y está ausente en el M4. Lingualmente a la cúspide estilar B en los M1-2 se desarrolla una pequeña cresta que, corriendo por la parte lingual de la plataforma estilar, termina conectándose con la cúspide estilar D. Los cíngulos anterobasales son cortos y se vuelven progresivamente más estrechos del M1 al M4. Hacia el extremo labial de este cíngulo se aprecia en los M1-3 una pequeña cúspide estilar A. Sobre la cara lingual de los M2-3 existen sendos metacónulos vestigiales. Los protoconos de los M1-4 carecen de cíngulos basales (Fig. 2A-B).
Etimología
En homenaje al notable mastozoólogo argentino Elio Massoia (1936-2001) y en agradecimiento a sus desinteresados consejos y orientación sobre la sistemática de los didélfidos argentinos, recibidos por el autor durante la década de 1990.
Comentarios
El conjunto de rasgos craneodentarios presentes en el holotipo de esta nueva especie sustentan su asignación al género Philander: las crestas sagital y lambdoideas son de desarrollo apenas moderado y la morfología dentaria es esencialmente similar a la de las especies vivientes de este género; el M4 está más comprimido anteroposteriormente que en las especies de Didelphis (pero menos que en Lutreolina), la cúspide estilar D del M3 está vestigialmente desarrollada, el P3 no es ancho y bulboso sino normalmente desarrollado y la proporción y posición de las cúspides y crestas de los molares es idéntica a la de las especies vivientes de Philander. Sin embargo, en dos rasgos craneanos el holotipo de P. massoiai concuerda más con la morfología característica de las especies de Didelphis que con aquella de Philander: (1) los procesos postorbitales del frontal son gruesos y rugosos y (2) el punto de intersección de las crestas frontales es relativamente anterior, motivo por el cual la cresta sagital es comparativamente más larga que en las otras especies de Philander. Junto con el mayor tamaño de esta nueva especie (véase la Tabla 1), los dos rasgos mencionados constituyen, precisamente, las mayores diferencias entre ésta y las otras especies reconocidas de Philander.
La especie viviente P. canus muestra una distribución amplia en América del Sur, extendiéndose fundamentalmente por regiones periamazónicas hasta el sur de Bolivia y el noreste de la Argentina (Fig. 1A). En virtud de su proximidad geográfica, vale la pena profundizar las comparaciones morfológicas entre esta especie y P. massoiai. Lo primero que llama la atención es el tamaño claramente mayor de P. massoiai (Tabla 1). En vista dorsal los nasales son, en su extremo posterior, de aspecto similar (contorno romboidal), si bien el extremo posterior de estos huesos, en relación con los procesos postorbitales del frontal, se ubica más anteriormente en P. massoiai (Fig. 3A-B). Ya se mencionó que las crestas frontales son más perpendiculares al eje anteroposterior que en las especies vivientes del género. En vista dorsolateral se aprecia que existe un único foramen lacrimal en P. massoiai (en P. canus se observa uno o dos forámenes, según los individuos). Si bien el lacrimal está roto, aparentemente el contacto fronto-maxilar es de similar extensión en ambas especies. El aspecto de los parietales y del interparietal es también muy similar. En la dentición superior se destaca el aspecto más robusto de los premolares y molares de P. massoiai. La diastema entre el P1 y el P2 es más corta (Fig. 3D). A juzgar por el tamaño de las raíces, el P1 de P. massoiai era proporcionalmente más ancho que el de P. canus y se orientaba más oblicuamente, con la raíz anterior implantada más labialmente que la lingual. El P2 es un diente de aspecto más simétrico en P. massoiai, con la cúspide mayor orientada más centralmente, la cresta porterior a esta cúspide menos desarrollada y con el cíngulo labial más reducido. En P. canus las cúspulas posterobasales de los P2-3 están más desarrolladas. En la serie molar llama la atención el metacono del M4, más desarrollado y saliente posteriormente en P. canus que en P. massoiai (Fig. 3C-D). En el M3 se aprecia un vestigio de paracónulo y en los M2-3 aparece un metacónulo apenas desarrollado; ninguna de estas cúspides se observa en P. canus. El ectoflexo está muy poco desarrollado en ambas especies, si bien en el M3 de P. canus es algo más profundo (Fig. 3C-D). En este molar la StA está más próxima a la altura de la StB en P. massoiai (en P. canus es proporcionalmente más pequeña y se ubica en un nivel más bajo). La longitud relativa de la postmetacrístida del M3 es mayor en P. canus. En el M1 de esta última especie (y, en menor medida, en el M2) se aprecia una StE pequeña pero diferenciada, mientras que en P. massoiai es apenas distinguible. La plataforma estilar del M3 de esta especie está más desarrollada que en P. canus y muestra más rugosidades en el esmalte.
Los materiales hasta ahora referidos a la especie extinta P. entrerianus corresponden a elementos de la dentición inferior y a un dentario: el holotipo (MACN 8888) es un molar inferior derecho aislado, mientras que el ejemplar FCP-V-M 055 constituye un dentario derecho con el p2 y la serie m1-4 completa (Goin et al. 2013:figura 1). Es imposible, en consecuencia, una comparación directa entre estos y el tipo de P. massoiai, cuyas únicas piezas dentarias preservadas son dos premolares y los molares superiores del hemimaxilar derecho. Sin embargo, es evidente que ambos no pertenecen a la misma especie. Téngase en cuenta que, en general, la longitud de la serie molar inferior de los marsupiales es mayor que la de los molares superiores. En el caso de P. entrerianus, la longitud de la serie molar inferior del ejemplar FCP-V-M 055 es de 13.7 mm, mientras que el holotipo de P. massoiai tiene una serie molar superior de 15.51 mm de largo, lo que permite suponer un tamaño significativamente mayor.
A partir del Mioceno medio la región de los Valles Calchaquíes comenzó un proceso de aridificación, con el establecimiento de vegetación xerofítica, como consecuencia de una nueva fase del Levantamiento Andino, el volcanismo asociado y cambios en la circulación oceánica y atmosférica (Barreda et al. 2007). Se sugiere que el origen de P. massoiai podría haber ocurrido en este complejo escenario. Concordantemente, Jansa et al. (2013) postularon que la diversificación de los didélfidos característicos de hábitats de bosque seco habría ocurrido recién a partir del Mioceno tardío. En particular para este lapso temporal los autores postularon la cladogénesis del linaje Lutreolina + (Philander + Didelphis).
Agradecimientos
Agradezco a Jaime Powell (1953- 2016), de la Sección Paleozoología Vertebrados, Instituto Miguel Lillo (Tucumán) por el préstamo del material objeto de este estudio. A N. Nasif y G. Esteban por la información provista al autor sobre la procedencia geográfica y estratigráfica del ejemplar. A Bruno Pianzola, del Laboratorio de Fotografía del Museo de Ciencias Naturales de La Plata, por las imágenes fotográficas que ilustran este trabajo. A Juana Yáñez que elaboró los dibujos esquemáticos de la Fig. 3 y a Marcela Tomeo quien colaboró en la realización de las ilustraciones. A dos revisores anónimos que permitieron mejorar la redacción y contenidos de este trabajo. Finalmente, a la Fundación Alexander von Humboldt (Alemania).
REFERENCIAS
Ameghino, F. 1898. Sinopsis geológico-paleontológica de la República Argentina. Segundo Censo de la República Argentina, Vol. 1. Buenos Aires. https://doi.org/10.5962/bhl.title.14834
Barreda, V. et al. 2007. Diversificación y cambios de las angiospermas durante el Neógeno en Argentina. Ameghiniana 50º Aniversario. Publicación Especial 11, Asociación Paleontológica Argentina, Buenos Aires: 173–191.
Castro-Arellano, I., H. Zarza, & R. A. Medellín. 2000. Philander opossum. Mammalian Species 638:1–8. https://doi.org/10.1644/1545-1410(2000)638<0001:po>2.0.co;2
Chemisquy, M. A., & D. A. Flores. 2012. Taxonomy of the southernmost populations of Philander (Didelphimorphia, Didelphidae), with implications for the systematics of the genus. Zootaxa 3481:60–72. https://doi.org/10.11646/zootaxa.3481.1.5
Esteban, G., & F. Abdala. 1993. Nuevos restos de Glossotheriopsis (Edentata-Tardigrada) de Tío Punco, Provincia de Tucumán. Análisis filogenético preliminar. Ameghiniana 30:328–329.
Esteban, G., N. Nasif, & S. Musalem. 1997. Nuevo registro de mamíferos fósiles en la región de Tio Punco (Formación Andalhuala - Mioceno tardío), provincia de Tucumán, Argentina. Ameghiniana 34:535.
Esteban, G. I., N. Nasif, & S. M. Georgieff. 2014. Cronobioestratigrafía del Mioceno tardío - Plioceno temprano, Puerta de Corral Quemado y Villavil, provincia de Catamarca, Argentina. Acta Geologica Lilloana 26:65–192.
Ferreira Lino, C.; C. M. Dias-Neto, E. Trajano, G. L. N. Gusso, I. Karmann, & R. Rodrigues. 1979. Paleontologia das Cavernas do Vale do Ribeira. Exploração I e Abismo do Fóssil (SP-145). Resultados parciais. Atas do 2 Simpósio Regional de Geologia, Rio Claro, Sao Paulo. https://doi.org/10.11606/t.44.1994.tde-22042013-163755
Flores, D. A., R. M. Bárquez, & M. M. Díaz. 2008. A new species of Philander Brisson, 1762 (Didelphimorphia, Didelphidae). Mammalian Biology 73:14–24. https://doi.org/10.1016/j.mambio.2007.04.002
Gardner, A. L. 2005. Order Didelphimorphia. Mammal Species of the World: A Taxonomic and Geographic Reference, 3º Ed, Vol. 1 (D. E. Wilson & D. M. Reeder, eds.). Johns Hopkins University Press, Baltimore. https://doi.org/10.1108/09504120610673024
Georgieff, S. M., Ibáñez, L. M., Vides, M. E., Anis, K. B., & Nieva, S. M. 2014. Paleógeno y Neógeno de Tucumán: estratigrafía y paleoambientes sedimentarios. Geología de Tucumán (Moyano, S., Puchulu, M. E., Fernández, D., Aceñolaza, G., Vides, M. E., Nieva, S. eds.). Colegio de Graduados en Ciencias Geológicas, Tucumán.
Goin, F. J. 1997. Sobre la edad y afinidades de Zygolestes paranensis Ameghino, 1898 (Marsupialia, Didelphidae, Marmosinae). Neotrópica 43:15–19.
Goin, F. J., & U. F. J. Pardiñas. 1996. Revisión de las especies del Género Hyperdidelphys Ameghino, 1904 (Mammalia, Marsupialia, Didelphidae). Su significación filogenética, estratigráfica y adaptativa en el Neógeno del Cono Sur Sudamericano. Estudios Geológicos 52:327–359. https://doi.org/10.3989/egeol.96525-6275
Goin, F. J., J. I. Noriega, & M. De Los Reyes. 2013. Los Metateria (Mammalia) del “Mesopotamiense” (Mioceno tardío) de la Provincia de Entre Ríos, Argentina, y una reconsideración de Philander entrerianus (Ameghino, 1899). El Neógeno de laMesopotamia Argentina (D. Brandoni & J. I. Noriega, eds.). Asociación Paleontológica Argentina, Publicación Especial 14,Buenos Aires.
Goin, F. J., M. O. Woodburne, A. N. Zimicz, G. M. Martin, & Chornogubsky. 2016. A brief history of South American metatherians. Evolutionary contexts and intercontinental Dispersals. Springer Earth System Sciences Series, Dordretch. https://doi.org/10.1007/978-94-017-7420-8
Hadler, P., J. Ferigolo, & F. J. Goin. 2009. Mamíferos de pequeno porte (Didelphimorphia, Chiroptera e Rodentia) do Pleistoceno Final/Holoceno do Brasil, com ênfase no Rio Grande do Sul. Quaternario do Rio Grande do Sul. Integrando conhecimentos (A. M. Ribeiro, S. G. Bauermann, & S. Saldanha Scherer, eds.). Monografías da Sociedade Brasileira de Paleontologia, Porto Alegre. https://doi.org/10.29289/259453942018v28s1059
Hershkovitz, P. 1997. Composition of the Family Didelphidae Gray, 1821 (Didelphimorphia, Marsupialia), with a review of the morphology and behavior of the included four-eyed pouched opossum of the genus Philander Tiedemann, 1808. Fieldiana, Zoology 86:1–103. https://doi.org/10.5962/bhl.title.3146
Jansa, S. A., & R. S. Voss. 2000. Phylogenetic studies on didelphid marsupials I. Introduction and preliminary results from nuclear IRBP gene sequences. Journal of Mammalian Evolution 7:43–77.
Jansa, S. A., K. Barker, & R. S. Voss. 2013. The early diversification history of didelphid marsupials: a window into South America’s “Splendid Isolation”. Evolution 68:684–695 https://doi.org/10.1111/evo.12290
Lew, D., R. Pérez-Hernández, & J. Ventura. 2006. Two new species of Philander (Didelphimorphia, Didelphidae) from Northern South America. Journal of Mammalogy 87:224–237. https://doi.org/10.1644/05-mamm-a-065r2.1
Marshall, L., & B. Patterson. 1981. Geology and geochronology of the mammal-bearing Tertiary of the Valle de Santa María and Río Corral Quemado, Catamarca Province, Argentina. Fieldiana Geology, New Series 9:1–80. https://doi.org/10.5962/bhl.title.3461
Nasif N. L., G. Esteban, S. M. Georgieff, L. Ibañez, & C. Spagnuolo. 2019. Biocronoestratigrafía y paleoambientes del Mioceno tardío de Catamarca, Tucumán y Santiago del Estero, Argentina. Mioceno al Pleistoceno del centro y norte de Argentina. (N. Nasif, G. Esteban, J. Chiesa, A. Zurita & S. Georgieff, eds.). Opera Lilloana 52:229–250.
Patton, J. L., & M. N. F. Da Silva. 1997. Definition of species of pouches four-eyed opossums (Didelphidae, Philander). Journal of Mammalogy 78:90–102. https://doi.org/10.2307/1382642
Patton, J. L., & M. N. F. da Silva. 2008. Genus Philander Brisson, 1762. Mammals of South America, vol. 1 - Marsupialia, Insectivora, Edentata, Chiroptera (A. L. Gardner, ed.). Chicago University Press, Chicago.
Patton, J. L., S. F. Dos Reis, & M. N. F. Da Silva.1996. Relationships among didelphid marsupials based on sequence variation in the mitochondrial cytochrome b gene. Journal of Mammalian Evolution 3:3–29. https://doi.org/10.1007/bf01454252
Pine, R. H. 1973. Anatomical and nomenclatural notes on opossums. Proceedings of the Biological Society of Washington 86:391–402.
Reig, O. A. 1957. Sobre la posición sistemática de Zygolestes paranensis Ameg. y de Zygolestes entrerianus Amegh., con una reconsideración sobre la edad y la correlación del “Mesopotamiense”. Holmbergia 5:209–226.
Reig, O. A., J. A. W. Kirsch, & L. G. Marshall. 1987. Systematic relationships of the living and Neocenozoic American opossum-like marsupials (suborder Didelphimorphia) with comments on the classification of these and of the Cretaceous and Paleocene New World and European metatherians. Possums and Opossums: Studies in Evolution, Vol. 1. (M. Archer, ed.). Surrey Beatty and Sons Pty. Ltd. & The Royal Zoological Society of New South Wales, Sydney.
Simpson, G. G. 1972. Didelphidae from the Chapadmalal Formation in the Museo Municipal de Ciencias Naturales of Mar del Plata. Publicaciones del Museo Municipal de Ciencias Naturales de Mar del Plata 2:1–40.
Vieira, E. M., & D. Astúa de Moraes. 2003. Carnivory and insectivory in neotropical marsupials. Predators with pouches: the biology of carnivorous marsupials (M. Jones, C. Dickman, & M. Archer, eds.). CSIRO Publishing, Collingwood. https://doi.org/10.1071/9780643069862
Vieira, E. M., & N. F. Camargo. 2012. Uso do espaço vertical por marsupiais brasileiros. Marsupiais do Brasil. Biologia, Ecologia e conservacao (N. Cáceres, ed.). Ed. UFMS, Universidade Federal de Mato Grosso do Sul, Campo Grande. https://doi.org/10.20873/uft.2179-3948.2018v9n3p290
Voss, R. S., & S. A. Jansa. 2003. Phylogenetic studies of didelphid marsupials II. Nonmolecular data and new IRBP sequences: separate and combined analyses of didelphine relationships with denser taxon sampling. Bulletin of the American Museum of Natural History 276:1–82. https://doi.org/10.1206/0003-0090(2003)276<0001:psodmi>2.0.co;2
Voss, R. S, & T. C. Giarla. 2020. A Revision of Philander (Marsupialia: Didelphidae), Part 2: Phylogenetic Relationships and Morphological Diagnosis of P. nigratus Thomas, 1923. American Museum Novitates 3955:1–16. https://doi.org/10.1206/3955.1
Voss, R. S., & S. A. Jansa. 2009. Phylogenetic relationships and classification of didelphid marsupials, an extant radiation of New World metatherian mammals. Bulletin of the American Museum of Natural History 322:1–177. https://doi.org/10.1206/322.1
Voss, R. S., J. F. Díaz-Nieto, & S. A. Jansa. 2018. A revision of Philander (Marsupialia: Didelphidae), Part 1: P. quica, P. canus, and a new species from Amazonia. American Museum Novitates 3891:1–70. https://doi.org/10.1206/3891.1
Wible, J. 2003. On the cranial osteology of the short-tailed opossum Monodelphis brevicaudata (Didelphidae, Marsupialia). Annals of Carnegie Museum 72:137–202.
Winge, H. 1893. Jorfundne og nulevende Gnawere (Marsupialia) fra Lagoa Santa, Minas Gerais, Brasilien: med udsigt over gnavares pungdyrdes slaegtskab. E Museo Lundii Band 1:1–133. https://doi.org/10.5962/bhl.title.14821
Información adicional
Editor asociado: U. F. J. Pardiñas & C. A. Galliari
Citar como: Goin, F. J. 2021. Una nueva especie de Philander (Marsupialia, Didelphidae) del Mioceno tardío del noroeste Argentino. Mastozoología Neotropical, 28(1):e0540. https://doi.org/10.31687/saremMN.21.28.1.0.10