ARTÍCULO
SOBRE LA ANATOMÍA CRANEANA DE MORENELAPHUS CARETTE, 1922 (MAMMALIA, CERVIDAE) Y FORMAS RELACIONADAS: DESCRIPCIÓN Y COMPARACIÓN MORFOLÓGICA DEL ÁREA DEL BASICRÁNEO
SOBRE LA ANATOMÍA CRANEANA DE MORENELAPHUS CARETTE, 1922 (MAMMALIA, CERVIDAE) Y FORMAS RELACIONADAS: DESCRIPCIÓN Y COMPARACIÓN MORFOLÓGICA DEL ÁREA DEL BASICRÁNEO
Mastozoología Neotropical, vol. 28, núm. 2, p. 579, 2021
Sociedad Argentina para el Estudio de los Mamíferos
Recepción: 09 Diciembre 2020
Aprobación: 01 Marzo 2021
Resumen: Entre los ciervos actuales y fósiles del Nuevo Mundo, Morenelaphus representa el género más conspicuo para el Pleistoceno de Argentina. Sin embargo, su registro fósil en América del Sur es conocido principalmente a través de sus astas, sin haber aportes sobre la anatomía de su cráneo. Con el fin de aclarar algunas cuestiones de la taxonomía y sistemática de los ciervos fósiles sudamericanos, en este caso se describen y analizan los principales rasgos anatómicos del basicráneo de Morenelaphus comparándolo con formas fósiles y vivientes de Cervinae Cervini (Goldfuss 1820), Capreolinae Rangiferini (Brookes 1828) y Odocoileini (Pocock 1923) procedentes de América. Varios rasgos identificados a nivel del basicráneo son únicos del género Morenelaphus, mientras que otros caracteres son compartidos con especies de la Tribu Cervini (Cervus), Odocoileini (Blastocerus, Odocoileus) y Rangiferini (Rangifer). Nuevos registros y futuras comparaciones entre más formas fósiles y vivientes (e.g., Alces alces), sumando más estructuras anatómicas a nivel del cráneo, dentición y postcráneo, servirán para esclarecer la sistemática del grupo dada la complejidad taxonómica y filogenética de las formas fósiles.
Palabras clave: Anatomía, Cervidae, Morenelaphus, Pleistoceno.
Abstract:
ON THE CRANIAL ANATOMY OF MORENELAPHUS (CARETTE 1922) (MAMMALIA, CERVIDAE) AND ITS RELATIVES: STUDY OF THE ANATOMY OF THE BRASICRANIUM. On the cranial anatomy of Morenelaphus (Carette 1922) (Mammalia, Cervidae) and its relatives: Study of the anatomy of the brasicranium. Among South American fossil deer, Morenelaphus is the most conspicuous genus from the Pleistocene beds of Argentina. However, its fossil record in South America is almost known only by antlers, the anatomy of the skull being poorly understood. In order to know the cranial morphologies in fossil and living forms useful to elucidate the systematic taxonomy of some South American deers, we describe and analyze the main anatomical features of the basicranium in Morenelaphus compared with fossil and living Cervinae Cervini (Goldfuss 1820), Capreolinae Rangiferini (Brookes 1828) and Odocoileini (Pocock 1923) forms from America. Several morphologies reported here are observed only in Morenelaphus. However other features are shared with some South American Cervini (Cervus), Odocoileini (Blastocerus, Odocoileus) and also with Rangiferini (Rangifer). Given the taxonomic complexity of South American fossil Cervidae, new records and future comparisons among fossil and living deer (e.g., Alces alces) using more anatomical structures from the braincase, dentition and postcranial bones, will clarify even more the systematic of the group.
Keywords: Anatomy, Cervidae, Morenelaphus, Pleistocene.
INTRODUCCIÓN
Los Cervidae representan una familia del orden Artiodactyla que ingresó en América del Sur durante la ola de migración GABI 2 del Gran Intercambio Biótico Americano (Woodburne 2010), con los primeros registros fósiles de la familia provenientes del Pleistoceno Medio de Argentina (ca. 500 ka AP; Cione et al. 2015). En este continente se registra una amplia diversificación, lo cual ha llevado a establecer varios géneros y distintas especies vi- vientes y fósiles (Mattioli 2011; Cassini et al. 2016). Sin embargo, la alfa taxonomía tradicional ha reconocido al menos cuatro géneros fósiles para el Cuaternario de América del Sur. Estos corresponden a AntiferAmeghino, 1889, EpieurycerosAmeghino, 1889, Paraceros Ameghino, 1889 y Morenelaphus (Carette 1922). Para cada uno de estos géneros se han creado varias especies junto a formas fósiles de los géneros vivientes Hippocamelus y Mazama (Ameghino 1889; Roth 1904; Carette 1922; Castellanos 1924; Cabrera 1929).
A diferencia de los taxones vivientes, los enfoques sistemáticos en ciervos fósiles sudamericanos, para obtener una mejor apreciación de la anatomía y alfa taxonomía, definieron taxones fundamentados principalmente en la morfología de las astas (Gervais & Ameghino 1880; Ameghino 1889; Carette 1922; Castellanos 1924; Kraglievich 1932), sin realizar un detallado análisis craneano o postcraneano, debido a la escasez de elementos óseos asociados (Menegaz 2000). A pesar de que las astas de los ejemplares fósiles suelen encontrarse muy fragmentadas en los afloramientos de América del Sur, el hallazgo de piezas completas puede aportar información sobre la variabilidad de su forma (e.g., Cabrera 1929; Churcher 1966; Churcher & Pinsof 1987; Alcaraz 2010). La morfología de las astas de ciervos vivientes es altamente variable intraespecíficamente, tanto en forma como tamaño, debido a factores tales como el desarrollo ontogenético, la dieta y el comportamiento social, lo cual las convierte en un carácter taxonómico no muy fiable (Brooke 1878; Pocock 1912; Carette 1922; Rusconi 1936; Churcher 1966; Popielno & Oberlaufing 1985; Churcher & Pinsof 1987; Duarte 1997; Maffei 2001; Azorit et al. 2002; Bubenik & Hundertmark 2002; González et al. 2002; Kaiser & Croitor 2004; Ungerfeld et al. 2008a,b; Breda 2008; Duarte et al. 2008; Flueck & Smith-Flueck 2011; Putman & Flueck 2011; Vezzosi 2015; Van Der Geer 2018). Mediante el empleo del actualismo, algunos autores intentaron esclarecer, de forma preliminar, las relaciones sistemáticas de los taxones extintos con respecto a las especies vivientes de América del Sur (e.g., Cabrera 1929; Castellanos 1945; Churcher 1966; Bianchini & Delupi 1979; Menegaz 2000; Kolb et al. 2015). Esto trajo aparejado una sistemática sumamente controvertida en los cérvidos sudamericanos, con taxones fósiles pobremente fundamentados y definidos según el sesgo de preservación presente en la familia (Carette 1922; Gervais & Ameghino 1880; Kraglievich 1932; Castellanos 1940, 1945, 1957; Bangalasta 1980; Alcaraz & Francia 2013).
Para el género MorenelaphusCarette, 1922 se han reconocido dos especies registradas en el Pleistoceno de América del Sur, los primeros registros son de Argentina: Morenelaphus brachyceros (Gervais & Ameghino 1880) y M. lujanensis (Ameghino 1888). La distinción sistemática de ambas especies de Morenelaphus se sustenta únicamente por la morfológica de las astas (Gervais & Ameghino 1880; Ameghino 1888; Cabrera 1929; Menegaz 2000; Alcaraz 2010), aunque ciertos rasgos craneales y dentarios permiten reconocer diferencias en las formas fósiles (Vezzosi et al. 2020; Rotti et al. 2021). Algunos autores justifican que tales especies están distribuidas en intervalos biocronológicos diferentes dentro del Pleistoceno (Alcaraz 2010; Cassini et al. 2016), aunque otros autores plantean que la familia se encuentra presente a partir del Plioceno en Argentina (Cione & Tonni 1999, 2005; Menegaz 2000; Woodburne 2010; Cione et al. 2015), pero sin registros fósiles que la sustenten. Por otra parte, estudios moleculares han propuesto una historia evolutiva para al menos ocho formas ancestrales de ciervos de América del Sur que habrían ingresado al continente durante el lapso Plio-Pleistoceno (Duarte et al. 2008). A su vez, existen escasos restos inéditos de astas asociados a fragmentos craneales (Menegaz 2000; Alcaraz 2010). En menor medida se han comentado elementos poscraneales con fragmentos de cráneos y astas (Ubilla 1996; Alcaraz 2010; Vezzosi et al. 2019). Existen actualmente estudios que han intentado resolver parcialmente la sistemática del grupo, mediante el uso de otras estructuras morfológicas a nivel craneal y postcraneal (Vezzosi 2015; Orcesi et al. 2019).
En este contexto, la presente contribución aporta novedades anatómicas que permitirán reconocer el género Morenelaphus a partir de rasgos morfológicos descriptos en el basicráneo, distinguirlo de los demás ciervos de América, y avanzar en su posicionamiento sistemático y filogenético dentro de la familia Cervidae.
MATERIALES Y MÉTODOS
Para la descripción de los caracteres craneanos (forámenes, procesos e inserciones musculares) se utilizó la terminología propuesta por Matthew (1908), Frick (1937), Loponte (2005), NAV (2005) y Nicolay & Vaders (2006). Para las estructuras timpánicas se siguió a Delupi & Bianchini (1995a,b), Norris (1999), O’leary (2010), Valli (2010), Onuk et al. (2013), Shi et al. (2014) y Vezzosi et al. (2020). Para las comparaciones se utilizaron cráneos de cérvidos de colecciones científicas de Argentina y de otros países (e.g., Chile, EE. UU.). Todos los especímenes examinados y las abreviaturas institucionales son detalladas en la Información Suplementaria (IS). Se compararon y analizaron las morfologías del basicráneo de los especímenes más completos de Morenelaphus con cráneos y astas de diferentes especies vivientes de ciervos neotropicales (Blastocerus dichotomus, Hippocamelus antisensis, Hippocamelus bisulcus, Mazama gouazoubira, Mazama nana, Mazama americana, Odocoileus virginianus, Ozotocerus bezoarticus, Pudu puda, Pudu mephistophiles), y de América del Norte y Eurasia (Axis axis, Cervus elaphus, Dama dama, Rangifer tarandus). Materiales no disponibles para comparaciones directas (i.e., Candiacervus ropalophorus, Megaloceros giganteus, Navahocerus fricki, etc.) fueron comparados usando ilustraciones y descripciones publicadas (Croitor 2006; Webb 1992; Vislobokova 2013), además de materiales de colecciones oficiales disponibles online.
Abreviaturas anatómicas. ali, alisfeonides; be, basiesfenoides; bo, basioccipital; bu, bula timpánica; ca, canal auditivo; cc, canal carótido; co, cóndilo occipital; cra, cresta articular; ct, cresta temporal; fal, fosa anterolateral; fa, fosa articular; fc, foramen condilar; fo, foramen oval; for, foramen orbitorotundum; fesm, foramen estilomastoide; fh, foramen hipogloso; fl, foramen lacerum; fj, foramen jugular; fm, foramen magnum; fr, frontal; fpg, foramen postglenoide; ft, foramen temporal; mae, meato acústico externo; mt, meato del temporal; p, pedicelo; pa, parietal; pes, proceso estiliforme; pm, proceso del mastoides; poe, proceso occipital externo; ppa, proceso paraoccipital; ppo, proceso postglenoide; pt, proceso pterigoides; pr, protuberancia, ta, tuberosidad articular; ti, timpanohial; tm, tubérculo muscular; spo, sutura parieto-occipital; so, supraoccipital; tup, tuberosidad posterior; tua, tuberosidad articular.
RESULTADOS
Sistemática Paleontológica
Orden Artiodactyla (Owen 1848) Suborden Ruminantia Scopoli, 1777 Infraorden Pecora Linnaeus, 1758 Familia Cervidae (Goldfuss 1820) Morenelaphus Carette, 1922
Material craneal referido: MACN-Pv 11408, MACN-Pv 11479, MACN-Pv 11564, MACN-Pv 11722, MACN-Pv 11730, MACN-Pv 12082, MACN-Pv12630 partim (Cotipo de Nothropus carcaranensis), MACN- Pv 15697, MACN-Pv 16578, MACN-Pv 16580; MARC- Pv 2390164; MLP-Pv 9-10, MLP-Pv 51-IV-11-69, MLP- Pv 63-V-31-1; MPA-85-189-I-D, MFA-G-Pv1311, MFA- G-Pv 1312, MFA-G-Pv 1681, MFA-G-Pv 1722, MFA-G-Pv 1723, MFA-G-Pv 1724, MFA-G-Pv 1725, MFA-G-Pv 1726; MUFYCA 1027, MUFYCA 1432 (Figs. 1-2 y S1-S2).
Descripción anatómica
Occipital. En vista posterior, el plano nucal es de contorno subcircular, con el borde dorsal recto y contactando con el parietal (Fig. 1 y S1). La sutura parieto-occipital desciende lateral y distalmente, curvándose hacia el área postero-dorsal del escamoso (Fig. 1). Por detrás de la sutura parieto-occipital, la tuberosidad occipital dorsal limita con la cresta homónima. El proceso del mastoides del occipital se extiende ventrolateralmente a lo largo del borde posterior del escamoso (Fig. 2 y S2), hacia el límite que comunica el extremo superior del mastoides. El hueso mastoides está extendido en una placa vertical y angosta que se comprime entre la superficie posterior del escamoso y el borde anterolateral del occipital (Figs. 1a, 2a-c).
En algunos especímenes de Morenelaphus (MFA-G- Pv 1723, MFA-G-Pv 1311) se distingue una constricción lateral por encima del área en la que se ubica la cresta del mastoides y por encima del foramen mastoideo. Por encima del foramen magnum, se distinguen dos pequeñas protuberancias separadas por un pequeño surco que se comunica dorsalmente con la cresta occipital (Vezzosi et al. 2020; Fig. 1b). Estas protuberancias pueden estar ausentes (MFA-G- Pv 1311, MFA-G-Pv 1723) o incluso formar una única tuberosidad distal (MFA-G-Pv 1722, MFA-G-Pv 1726, MACN-Pv 16580; Fig. S1). No obstante, en algunos especímenes adultos de Ozotoceros bezoarticus (e.g., MACN-Ma 4297) suelen estar presentes. El foramen magnum es de contorno suboval.

Fig. 1
Caja craneana de ejemplares fósiles de Morenelaphus y taxones vivientes de Cervidae: a. Vista lateral del cráneo: 1. MFA-G-Pv 1312, 2. MFA-G-Pv 1722, 3. MACN-Pv 12630; b. Vista caudal de la región occipital: 4. MFA-G-Pv 1312, 5. MFA-G-Pv 1722, 6. MACN-Pv 12630; c. Vista ventral del basicráneo: 7. MFA-G-Pv 1312, 8. MFA-G-Pv 1722, 9. MACN-Pv 12630, 10. MPA-85-189-I-D, 11. Cervus elaphus (MACN s/n). 12. Rangifer tarandus (MACN 4331). Ver materiales y métodos para descripción de las abreviaturas anatómicas.
Los cóndilos occipitales son grandes, con una gran superficie de contacto para el atlas. El gran desarrollo y contorno reniforme se debe a que contribuyen a la función de mantener la postura y evitar una sobreflexión de la cabeza (Webb 1992). En vista ventral, los cóndilos occipitales son robustos, tienen un contorno subtriangular. La fosa condilar alberga el foramen condilar y el foramen del nervio hipogloso (Fig. 2). Anterior a cada fosa condilar están los forámenes jugulares. En varios ejemplares no se observan estos forámenes en vista palatal, debido al gran desarrollo que adquieren las tuberosidades (Fig. 2a). Los procesos paroccipitales se proyectan ventral y paralelamente a los cóndilos, aunque en ninguno de los ejemplares se preservaron completos.
Basioccipital. Se encuentra fusionado al basiesfenoides formando una superficie triangular (Fig. 1c). En contacto con ambos cóndilos presenta dos tuberosidades posteriores, las que se proyectan distalmente hacia ambos lados de los procesos paraoccipitales (Vezzosi et al. 2020). Estas tuberosidades muestran desarrollo similar como ocurren en B. dichotomus, Ozotoceros bezoarticus y Odocoileus virginianus. Sin embargo, en Hippocamelus spp., Mazama spp. y Pudu puda, ambas tuberosidades se encuentran proporcionalmente menos desarrolladas (Fig. 3). El surco mesiodistal en el basioccipital continúa en una cresta convexa y limitada por dos tubérculos musculares (=tuberosidades faríngeas según Croitor 2006; Shi et al. 2014) conspicuos y de tamaño variable, aunque menos desarrollados que aquellos posteriores (Fig. 1c).
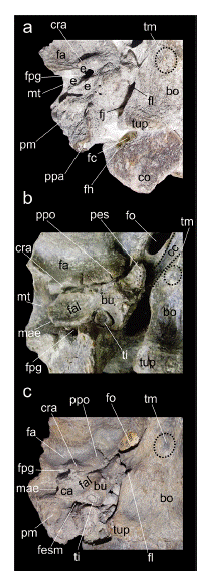
Fig. 2
Detalle del sector auditivo y estructuras asociadas en Morenelaphus: a. MACN-Pv 16578; b. MACN-Pv 11565; c. MACN-Pv 11730. Ver materiales y métodos para descrip- ción de las abreviaturas anatómicas.
Basiesfenoides. Es de contorno triangular, más angosto y largo que el basioccipital. La superficie es convexa y presenta una cresta convexa longitudinal que continúa al surco antes mencionado.
Sector auditivo. La bula timpánica de sección sub-triangularse ha preservado parcialmente en escasos ejemplares (Fig. 2). A su vez, el petroso se conserva completo en pocos ejemplares (MACN-Pv 12630,MFA-Pv 1312, Orcesi et al. 2019). Es proporcionalmente pequeño y levemente alargado. La fosa articular cóncava y el proceso postglenoide se proyectan transversalmente, aunque este proceso se posiciona transversalmente al foramen oval (Fig. 1). Ambas estructuras delimitan el borde posterior de la faceta articular, que articula con la mandíbula. Detrás de este foramen, la bula timpánica es menos inflada en comparación con Mazama spp., Od. virginianus, P. dichotomus, C. elaphus, Oz. bezoarticus, P. mephistophiles e Hippocamelus spp. (Vezzosi et al. 2020; Fig. 3). Dependiendo del estadio de desarrollo en los ejemplares de Morenelaphus (Vezzosi et al. 2020), la depresión para el receso del timpanohial forma una cavidad bien definida y profunda o puede representar una superficie plana y circular. En aquellos ejemplares de Morenelaphus en los que la bula no se ha preservado, se reconoce el foramen postglenoide circular y limitado por el hueso escamoso. Lateralmente, este foramen se posiciona lateral y anteriormente al meato acústico externo. El meato presenta un diámetro mayor de apertura y se proyecta latero-distalmente hacia el área occipital (Fig. 1). La bula muestra una fosa anterolateral bien desarrollada que limita posteriormente con una cresta transversal al basioccipital (Fig. 2). El hueso timpanohial presenta sección circular y posición ventral. Este hueso se ubica dentro de una invaginación o receso del timpanohial (Norris 1999; Vezzosi et al. 2020) con profundidad variable (Figs. 1 y 2). El extremo anteromedial del área preservada de la bula muestra el proceso estiliforme, robusto con el borde anterior redondeado y sección subtriangular. El foramen lacerum se posiciona posteriormente al canal carótido que limita, a su vez, con el foramen oval grande y de contorno suboval. El foramen orbitorrotundo es circular en sección y su dimensión es similar al oval. Inmediatamente posterior al borde caudal de la bula hay un foramen estilomastoideo, pequeño y redondeado.

Fig. 3
Vista ventral del basicráneo de ejemplares machos adultos de Cervidae vivientes de América y el Viejo Mundo: Blastocerus dichotomus (MACN-Ma 13-1); b. Hippocamelus bisulcus (SGO.Ma 1024); c. H. antisiensis (MACN-Ma 5362); d. Ozotoceros bezoarticus (MACN-Ma 49183); e. Odocoileus virginianus (AMNH 100289); f. Mazama gouazoubira (CFA-MA 06345); g. M. nana (MACN-Ma 24691); h. Pudu puda (SGO.Ma 559); i. P. mephistophiles (AMNH 181506); j, Rangifer tarandus (MACN); k, Dama dama (MACN-Ma 4397); L, Cervus elaphus (MACN-Ma s/n). Sin escalar.
Escamoso. El proceso postglenoide en vista palatal se observa bien desarrollado y expandido lateralmente (Fig. 1), mientras que la cresta articular, con un grueso reborde distal, se ubica transversalmente al eje longitudinal del cráneo. La fosa articular es cóncava y transversal al eje longitudinal del cráneo y está delimitada anteriormente por la tuberosidad anterior. En vista lateral el escamoso es alargado, con el eje mayor orientado en forma oblicua, desarrollando una dirección anteroventral-posterodorsal. La sutura parieto-escamosal es convexa del lado del escamoso y paralela a los ceratóforos de las astas. Sobre el borde ventral hay una marcada cresta temporal, que forma el borde lateral de la faceta articular. Dorsal a la cresta temporal está el foramen temporal, de tamaño variable, y en algunos ejemplares dividido en dos orificios, uno anterior pequeño y uno posterior más grande. Incluso en algunos ejemplares de Morenelaphus este foramen puede no estar presente. Interesa destacar que estas variaciones en el carácter (i.e., presencia/ausencia, foramen simple/doble) se han podido observar dentro de la muestra de todas las especies vivientes comparadas en esta investigación.
Cabe mencionar que, en los diferentes taxones vivientes, la bula adquiere volumen y tamaño variable, desde formas bien globosas con mayor volumen (Mazama gouazoubira, Odocoileus virginianus), a levemente convexas con menor volumen (Rangifer tarandus, Hippocamelus spp.) o muy re ducidas (Blastocerus dichotomus, Cervus elaphus, Ozotoceros bezoarticus, Pudu spp.; Figs. 1 y 3).
Comparaciones
El plano nucal de Morenelaphus presenta el occipital bajo dorsoventralmente y extendido transversalmente (Fig. 1), similar a B. dichotomus y H. antisensis, aunque estos dos tienen occipitales menos bajos dorsoventralmente (Fig. S3). En los demás cérvidos observados (i.e., H. bisulcus, Oz. bezoarticus, Od. virginianus, D. dama) los occipitales son más equidimensionales, es decir, casi tan altos como anchos (Fig. S3). En adición, el contorno general del occipital es redondeado y los bordes laterales son marcadamente convexos, con una constricción lateral muy leve. Estos rasgos asemejan a Morenelaphus con H. antisiensis, mientras que en los demás ciervos observados los bordes laterales son rectos y cóncavos, marcando más notoriamente la constricción lateral.
En vista ventral los cóndilos occipitales son robustos y cortos anteroposteriormente, el basioccipital es corto anteroposteriormente y ancho transversalmente y con las tuberosidades posteriores muy robustas y proyectadas posteriormente (Vezzosi et al. 2020). Los tubérculos musculares sobresalen de la superficie de contacto entre el basiesfenoides y el basioccipital con tamaño variable (Vezzosi et al. 2020), caracteres observados también en ejemplares machos de C. elaphus y B. dichotomus en menor medida (Fig. 3). Morenelaphus muestra similitud con R. tarandus en poseer un basioccipital muy ancho y con tuberosidades posteriores robustas, pero se diferencia en que este último presenta los tubérculos musculares poco marcados. En los demás ciervos comparados, los cóndilos son más largos, los basioccipitales no son tan anchos y no presentan tuberosidades posteriores tan robustas (Fig. S3). De las especies sudamericanas, en B. dichotomus, M. nana y M. americana se observa un basioccipital con proporciones similares a las de Morenelaphus, aunque sin las tuberosidades tan pronunciadas como este último. Los huesos basioccipital y basiesfenoides de Morenelaphus son muy distintos de Pudu spp., Od. virginianus, Hippocamelus spp. y M. gouazoubira, siendo largos anteroposteriormente. En todas las demás especies comparadas, las tuberosidades musculares están muy poco marcadas (Fig. 3).
En aquellos ejemplares de Morenelaphus que preservan parte de la región timpánica (Fig. 2c), la bula presenta una morfología general menos inflada que en los restantes taxones, con un tubo corto y cóncavo anteromedialmente que desemboca en el meato acústico externo (Fig. 2c). Dado el estado de preservación que poseen los ejemplares estudiados, no fue posible identificar la cresta ventral en el canal auditivo, como se reconoce claramente en B. dichotomus y en menor medida en Oz. bezoarticus. No obstante, en A. axis y R. tarandus la mencionada cresta se posiciona levemente en sentido mesial.
En vista lateral se observan varios rasgos para destacar. Morenelaphus muestra un escamoso similar al de Od. virginianus y H. antisensis (Fig. 1a). En las demás especies, las principales diferencias con Morenelaphus están presentes en el borde anterodorsal del escamoso, el cual está más proyectado anterodorsalmente, con un proceso anteroventral del parietal más estrecho (e.g., Oz. bezoarticus, H. bisulcus, B. dichotomus, D. dama y A. axis, Fig. S4). En otras especies, la diferencia con Morenelaphus se observa en que el proceso anteroventral del parietal se vuelve muy ancho, como en Mazama spp, P. puda y P. mephistophiles. Contrariamente, R. tarandus presenta escamoso mucho más bajo y proceso anteroventral conectado al resto del parietal, en el extremo posterior del cráneo. Los ceratóforos, en esta especie, salen de los frontales mucho más atrás que en los demás taxones, y por esto, los parietales están más restringidos posteriormente (Fig. S4). El foramen postglenoide, presente en los ejemplares de Morenelaphus estudiados, varía en tamaño y se ubica por detrás de la cresta articular (Fig. S2). Este foramen, en las demás especies vivientes de Cervinae y Capreolinae, es proporcionalmente mayor, aunque con tamaño variable; se ubica por detrás de la cresta articular rodeando anteroventral a anterodorsalmente el escamoso (Fig. 3). Distalmente, además de estar en contacto con el escamoso, limita con la paredsuperior (tegmen tympani, O’leary 2010) y proceso anterior del timpánico.
En comparación con otros ciervos extinguidos, Morenelaphus comparte con Navahoceros la presencia de un basioccipital transversalmente ancho y anteroposteriormente corto, tubérculos musculares marcados, foramen condilar y foramen hipogloso separados, pero cercanos entre sí. Sin embargo, se diferencian marcadamente en que Navahoceros presenta cóndilos occipitales muy extendidos anteroposteriormente, basiesfenoides largo y con dos crestas subparalelas, foramen oval semicubierto ventromedialmente por una repisa del basiesfenoides (Webb 1992). Esta repisa también está presente en ejemplares de Rangifer por lo cual el foramen oval casi no se observa en vista ventral (Fig. 3j).
Teniendo en cuenta la morfología craneana descrita para otros ciervos del Viejo Mundo, se observa que Morenelaphus se diferencia de Capreolus en que este último presenta los cóndilos occipitales muy extendidos anteroposteriormente en el plano ventral, los tubérculos posteriores y los tubérculos musculares están muy poco marcados, el basiesfenoides es largo y el foramen oval es mucho mayor al foramen orbitorotundo, entre otros rasgos (Onuk et al. 2013).
Morenelaphus comparte con los ciervos pliopleistocénicos de Europa y Asia, Eucladoceros, Orchonoceros y Praemegaceros, la forma del plano nucal del occipital (bajo dorsoventralmente y de contorno subcircular, sin las constricciones laterales cercanas al proceso mastoides), un foramen oval grande y subcircular, un basioccipital con forma de campana, ensanchado al nivel de los tubérculos musculares, los tubérculos posteriores bien desarrollados y proyectados posteriormente y un marcado surco medio entre los tubérculos musculares (Croitor 2006; Vislobokova 2012, 2013; Baygusheva & Titov 2013; Vezzosi et al. 2020); pero se diferencia en la forma de los cóndilos occipitales en el plano ventral, en el escaso desarrollo de las tuberosidades musculares, y en la posición del foramen postglenoide. Para otros ciervos euroasiáticos, como Candiacervus, Sinomegaceros, Arvernoceros, entre otros, Morenelaphus muestra una morfología mucho más disímil (Vislobokova 2012, 2013).
La forma del plano nucal del occipital, el foramen magnum, los cóndilos occipitales en vista ventral, el gran desarrollo de las tuberosidades posteriores y los tubérculos musculares son caracteres que comparte Morenelaphus con Megaloceros giganteus. No obstante, se diferencia de este último en que el basioccipital tiene bordes rectos y el basiesfenoides es más largo, el canal auditivo presenta fosa anterolateral muy pequeña y, además, el foramen oval se abre lateralmente (Croitor 2006; Vislobokova 2013).
Por lo tanto, la totalidad de los ejemplares analizados permitió establecer varios caracteres morfológicos que son constantes, es decir, están representados en el basicráneo de todos los ejemplares aquí asignados a Morenelaphus.
Dadas las comparaciones hechas, se mencionan y detallan las siguientes morfologías presentes en Morenelaphus y se identifican los taxones de Cervidae que comparten tales características con el taxón bajo estudio:
1- Cóndilos occipitales bien extendidos transversalmente y con morfología reniforme a sigmoidea como en Mazama spp., Hippocamelus antisiensis, Blastocerus dichotomus y Rangifer tarandus.
2- Plano nucal del occipital bajo, de contorno subcircular y sin constricciones laterales cercanas a los procesos mastoideos (Vezzosi et al. 2020), como en los ciervos plio-pleistocénicos de Eurasia (Eucladoceros, Orchonoceros, Praemegaloceros, Megaloceros, etc.).
3- Cóndilos occipitales muy cortos anteroposteriormente, como en C. elaphus.
4- Procesos paraoccipitales cortos y robustos, con orientación ventral como en B. dichotomus, Cervus elaphus y R. tarandus.
5- Fosa condiloidea con el foramen hipogloso y condiloideo separados, como en Navahoceros.
6- Basioccipital muy ancho y de contorno acampanado como en C. elaphus, B. dichotomus, Mazama nana, M. americana y R. tarandus.
7- Surco medio longitudinal bien marcado entre los tubérculos musculares, como en A. axis, C. elaphus, M. gouazoubira, M. nana y Od. virginianus.
8- Tubérculos posteriores del basioccipital muy robustos, proyectados posteriormente, solapando ventralmente el borde anterior de los cóndilos occipitales, como en C. elaphus.
9- Tubérculos musculares proyectados caudalmente, en dirección hacia el foramen magnum, como en C. elaphus, H. bisulcus, Od. virginianus, M. nana y P. puda. Para B. dichotomus y Oz. bezoarticus la orientación es caudo-lateral, dada la posición marginal de ambos. Mientras que en A. axis y D. dama ambos tubérculos están orientados cranealmente, y en sentido ventral en M. gouazoubira.
10- Basiesfenoides con cresta media claramente bien desarrollada como en D. dama y R. tarandus.
11- Meato acústico externo de sección circular y canal auditivo proyectándose distomedialmente como en Cervinae y Capreolinae, pero el canal forma un tubo corto como H. antisensis, Mazama spp., Pudu spp y R. tarandus.
12- Foramen estilomastoideo pequeño similar a todos los taxones comparados.
13- Foramen yugular pequeño y ubicado en una estrecha fosa que lo relaciona al foramen lacerum.
14- Timpanohial pequeño de sección circular como en las formas de Cervidae neotropicales y R. tarandus, aunque de posición claramente ventral.
15- Proceso estiliforme corto y robusto con el borde anterior redondeado y sección subtriangular, alcanzando el borde posterior del foramen oval, similar a H. antisiensis.
16- Foramen oval grande y de contorno redondeado a ovalado, con apertura ventral a ventrolateral, como en Mazama spp., P. mephistophiles y P. puda.
17- El foramen orbitorrotundo es muy grande y circular, con dimensiones y proporciones similares al foramen oval; aunque anteriormente orientado.
18- El foramen lacerum es pequeño y angosto, con apertura hacia el canal carotideo. Este canal se aprecia angosto y largo.
19- El foramen postglenoide de tamaño variable se conecta a un corto canal postglenoide y rodea al escamoso como en Cervinae y Capreolinae.
20- Cresta articular grande y alargada.
21- Proceso postglenoide expandido anterior y ventrolateral. La orientación de este proceso en B. dichotomus, M. gouazoubira y P. puda, como así también en algunos ejemplares de Morenelaphus, suele continuar en dirección hacia la apertura del foramen oval.
22- Fosa articular amplia y expandida lateralmente como en B. dichotomus, C. elaphus, H. bisulcus, Oz. bezoarticus.
23- Tuberosidad articular bien expandida lateralmente, con el margen anterior de dirección curva como en ejemplares machos de B. dichotomus, C. elaphus, H. bisulcus.
24- Borde anterodorsal del escamoso levemente convexo y proceso anteroventral del parietal de la misma forma que en Od. virginianus e H. antisensis.
DISCUSIÓN
Varias de las morfologías identificadas a nivel del basicráneo (ítems 2, 5, 12, 13, 17, 18, 19, 21) no están representadas en las especies vivientes de los Cervidae de América. Tales morfologías podrían representar características propias de Morenelaphus en un análisis que integre la totalidad de las estructuras anatómicas a nivel craneal, para esclarecer mejor su posición sistemática entre los taxones estudiados y reconocer diferencias a nivel interespecífico.
Numerosos autores intentaron esclarecer las relaciones sistemáticas de los ciervos fósiles de América del Sur (e.g., Ameghino 1889; Carette 1922; Castellanos 1924; Cabrera 1929; Churcher 1966), aunque la mayoría de ellos basaron sus conclusiones únicamente en el estudio de la forma y dimensiones de las astas. Por otro lado, se ha pretendido reconocer diferencias osteológicas y morfométricas en el cráneo y la mandíbula de las especies de ciervos neotropicales de América del Sur, aunque sin incluir formas extintas en las comparaciones anatómicas (Cabrera 1941; Hershkovitz 1982; Delupi & Bianchini 1995a,b; Merino et al. 2005). Con relación a la anatomía del basicráneo en ciervos sudamericanos, Cabrera (1941) aporta rasgos morfológicos para diferenciar y reconocer cuatro géneros neotropicales vivientes (Blastocerus, Hippocamelus, Ozotoceros, Odocoileus) al considerar como rasgos característicos la forma del foramen oval, el foramen orbitorrotundo y las bulas timpánicas. De estos cuatro taxones, solamente B. dichotomus, Od. virginianus e incluso C. elaphus muestran una morfología y orientación semejante en Morenelaphus a nivel del foramen postglenoide y el foramen oval, aunque este último de proporciones más pequeñas. Más aún, Delupi & Bianchini (1995a) elaboran un análisis exhaustivo sobre la forma de la bula timpánica en Oz. bezoarticus comparándola únicamente con representantes vivientes de las especies de ciervos neotropicales y un extinto artiodáctilo europeo de la familia Amphimerycidae Stehlin, 1910 (Pearson 1927). A pesar de las diferencias morfológicas encontradas en lo que se refiere a esta estructura anatómica entre los taxones estudiados, los autores logran diferenciar dos patrones morfológicos, uno para referir a los Cervinae o ‘ciervos del Viejo Mundo’ y otro para los Odocoileinae o ‘ciervos del Nuevo Mundo’ (Delupi & Bianchini 1995a). Desafortunadamente, en los ejemplares de Morenelaphus analizados aquí no se ha preservado de manera íntegra la bula timpánica. De modo tal que no es posible distinguir con claridad esta morfología en el género estudiado para lograr ubicarlo en una de las dos categorías taxonómicas.
Por otro lado, las estructuras anatómicas que aquí han podido compararse para la región auditiva comprenden el área del timpanohial y aquellas relacionadas (Garrod 1877; Lydekker 1915; Carette 1922). En las comparaciones hechas se ha demostrado que los especímenes de Morenelaphus presentan ciertas características particulares que lo distinguen del resto de los taxones estudiados (ítems 2, 5, 12, 13, 17, 18, 19, 21). A su vez, es importante destacar que ciertas morfologías observadas en el basicráneo de algunas especies extintas de América del Norte (Navahocerus y Eocoileus; Webb 2000), podrían estar relacionadas con las formas neotropicales estudiadas (ítem 5).
Varias de las características anatómicas identificadas en Morenelaphus (ítems 1, 4, 10, 11, 14, 19) han sido reconocidas en R. tarandus, y algunas de ellas compartidas también con los ciervos neotropicales (ítems 1, 4, 6, 11, 14, 19) e incluso C. elaphus (ítems 4, 6, 11, 19). Desde el punto de vista filogenético, estos rasgos podrían representar caracteres compartidos entre Rangifer y los representantes neotropicales, como fuera planteado por Carette (1922), cuando establece el grupo Neocervinae, y parcialmente con Frick (1937) quien considera con dudas la pertenencia de algunos ciervos neotropicales al grupo Rangiferinae. En este aspecto, Hershkovitz (1982) opina que Morenelaphus podría estar vinculado a la línea evolutiva de Rangiferini (Brookes 1828), la cual se habría diferenciado tempranamente de Odocoileini (Pocock 1923) (Grubb 2000, Mattioli 2011), o bien Morenelaphus podría constituir por sí solo una tercera rama diferenciada tempranamente de los odocoilinos.
Por otro lado, muchos de los rasgos compartidos entre Morenelaphus y Rangifer, también están presentes en los ciervos del viejo mundo (ítems 4, 6, 10, 11, 19). Incluso, Morenelaphus presenta varios caracteres compartidos únicamente con C. elaphus y otros cérvidos del Viejo Mundo (ítems 2, 3, 8). En adición, el mayor número de morfologías en común se observan con C. elaphus (9 de los 24 ítems aquí analizados). Esto está en concordancia con la propuesta de Kraglievich (1932) quien incluyó a Morenelaphus en los Cervinae. En síntesis, estos rasgos anatómicos podrían estar indicando una relación taxonómica mayor con el grupo Cervinae (Goldfuss 1820) que con Capreolinae (Brookes 1828). Sin embargo, estas evidencias morfológicas necesitan ser testeadas en un estudio sistemático filogenético exhaustivo con el fin de integrar y comparar otras estructuras craneales de importancia sistemática (e.g., región rostral, premaxilar y maxilar, dientes, astas).
Recientemente, Orcesi et al. (2019) realizan la primera descripción anatómica del oído interno en Cervidae fósiles del Pleistoceno de América del Sur, con el fin de lograr elucidar ciertas cuestiones sistemáticas y filogenéticas entre ellos y las especies vivientes neotropicales. Entre los primeros resultados obtenidos, los autores encuentran que varias de las características estudiadas a nivel del petroso son únicas y más desarrolladas en Morenelaphus y otras formas fósiles (e.g., flanco posteromedial, meato acústico interno), en tanto que otras características son compartidas con B. dichotomus. La nueva evidencia aquí observada junto a lo reportado para el oído interno (Orcesi et al. 2019) estaría relacionando aún más a Morenelaphus con los Cervini (Goldfuss 1820) y secundariamente con Odocoileini, coincidiendo parcialmente con lo mencionado previamente por Hershkovitz (1982). No obstante, es necesario realizar una comparación en detalle con las estructuras anatómicas del oído interno de taxones holárticos y del Viejo Mundo (e.g., A. alces, C, canadensis, C. elaphus, Megalocerus giganteus) que permitan confirmar estas hipótesis.
Tradicionalmente, se han reconocido dos grupos de Cervidae, los Plesiometacarpalia y los Telemetacarpalia (Brooke 1878), los cuales tienen valor sistemático para la mayoría de los autores (Cronin 1991; Hassanin & Douzery 2003; Pitra et al. 2004; Gilbert et al. 2006; Immel et al. 2015). El primero incluye los ciervos del Viejo Mundo relacionados a Cervus (i.e., Elaphurus, Dama, Axis, Muntiacus, etc.), mientras que el segundo abarca a los corzos (Capreolus spp.), los alces (Alces alces) y la mayoría de los ciervos del Nuevo Mundo (Rangifer y todos los ciervos neotropicales vivientes). Las características morfológicas que distinguen estos dos grupos son principalmente tres: 1) metacarpianos vestigiales II y V solo desarrollados en su mitad proximal (Plesiometacarpalia) o en su mitad distal (Telemetacarpalia), 2) la presencia de un septo vomeriano completo (Telemetacarpalia) o incompleto (Plesiometacarpalia), y 3) un foramen postglenoide totalmente delimitado por el escamoso (Plesiometacarpalia), o por el escamoso y posteromedialmente por el petroso (Telemetacarpalia) (Brooke 1878; Groves & Grubb 1987; Bouvrain et al. 1989). Si bien no se conocen restos poscraneanos asociados con cráneos de Morenelaphus, las características craneanas podrían arrojar luz sobre su pertenencia a uno u otro grupo. Sin embargo, en ninguno de los cráneos analizados aquí se ha preservado el área para el septo vomeriano, por lo cual no se puede visualizar qué nivel de desarrollo óseo alcanza esta estructura. Por otro lado, el foramen postglenoide se encuentra limitando por el hueso petroso y rodeando el escamoso, de la misma manera que se aprecia en taxones Cervini como también Odocoileini (ítem 19), lo que demuestra que este rasgo morfológico a nivel del cráneo no resulta apropiado para considerar una u otra condición.
Por otro lado, Rotti et al. (2016), al analizar ejemplares de Brasil de Morenelaphus, describen que estos especímenes presentarían rasgos característicos de los Plesiometacarpalia. En este aspecto, los rasgos anatómicos mencionados aquí como compartidos entre Morenelaphus y los ciervos del Viejo Mundo (i.e., Cervus, Dama, Axis, Megaloceros, etc.) podrían estar evidenciando una posible inclusión dentro de Plesiometacarpalia. Esto implicaría que el género Morenelaphus resultaría ser un representante basal de Cervidae que participó del Gran Intercambio Biótico Americano durante el Pleistoceno.
No obstante, un estudio más exhaustivo que contemple ejemplares más completos, junto con el análisis de la anatomía del oído interno y otras estructuras morfológicas de cráneos más completos asociadas a elementos postcreaneales, permitirá definir nuevos rasgos morfológicos para esclarecer aún más la sistemática de Morenelaphus, así como también intentar resolver la complejidad taxonómica y filogenética de los ciervos fósiles y vivientes de América.
Agradecimientos
Por asistirnos durante el acceso y estudio de los ma- teriales bajo su cargo se agradece a Rubén Bárquez y Mónica Díaz (Colección Mamíferos Lillo); Andrés Pautasso y Leonardo Leiva (Museo Provincial de Ciencias Naturales “Florentino Ameghino”); Pablo Teta y Sergio Lucero (Colección Mastozoología, Museo Argentino de Ciencias Naturales “Bernardino Rivadavia”); Alejandro Kramarz (Colección Paleontología de Vertebrados, Museo Argentino de Ciencias Naturales “Bernardino Rivadavia”); Itatí Olivares, Mariano Merino y Diego Verzi (División Mastozoología, Museo de La Plata, Universidad Nacional de La Plata, Buenos Aires); Marcelo Reguero (División Paleontología, Museo de La Plata, Universidad Nacional de La Plata, Buenos Aires); Enrique González (División de Mastozoología, Museo Nacional de Historia Natural de Montevideo, Uruguay); Matías Taglioreti, Fernando Scaglia y Victoria Sarasa (Laboratorio de Paleontología, Museo Municipal de Ciencias Naturales “Lorenzo Scaglia”), Silvia Cornero, Lucía Ragnone y Georgina Vaiana (Museo Florentino y Carlos Ameghino, Rosario, Santa Fe); Cristian Oliva y Rodrigo Tomassini (Museo Municipal Darwin de Punta Alta, Bahía Blanca). Un especial agradecimiento a Guillermo Cassini (MACN) por facilitarnos el uso de imágenes e información sobre especímenes de Cervidae vivientes procedentes del AMNH. Esta investigación fue parcialmente financiada por el CONICET (Argentina), PICT- ANPCyT 2017-0954 (Argentina) y PEDECIBA-UDELAR (Uruguay).
REFERENCIAS
Alcaraz, M. A. 2010. Sistemática de los ciervos (Mammalia, Artiodactyla) del Pleistoceno de las áreas extraandinas de Argentina. Tesis Doctoral (inédita). Universidad Nacional de La Plata. Buenos Aires. Argentina, 302 pp. https://doi.org/10.35537/10915/55174
Alcaraz, M. A., & A. Francia. 2013. Diversidad de Cervidae (Mammalia, Artiodactyla) en el Pleistoceno de la Provincia de Corrientes, Argentina. Revista Brasileira de Paleontologia 16:157-166. https://doi.org/10.4072/rbp.2013.1.12
Ameghino, F. 1888. Rápidas diagnosis de algunos mamíferos fósiles nuevos de la República Argentina. Obras Completas 5:471-480. https://doi.org/10.5962/bhl.title.14834
Ameghino, F. 1889. Contribución al conocimiento de los mamíferos fósiles de la República Argentina. Actas de la Academia Nacional de Ciencias de Córdoba 6:1-027. https://doi.org/10.5962/bhl.title.121288
Azorit, C., A. Mohamen, R. Carrasco, A. Carrasco, & A. Muñoz-Cobo. 2002. Astas, esqueletos y edad del ciervo (Cervus elaphus hispanicus) de Sierra Morena oriental: Estudio de correlación. Anales de Biología 24:195-200. https://doi.org/10.1007/bf02189987
Bangalasta, M. B. 1980, Algunas consideraciones sobre la validez del género Epieuryceros Ameghino, 1889. II Congreso Argentino Paleontología y I Congreso Latinoamericano Paleontología. Buenos Aires. 1978. T. III:183-192.
Baygusheva, V. S. & V. V. Titov. 2013. Large deer from the Villafranchian of Eastern Europe (Sea of Azov Region): Evolution and paleoecology. Quaternary International 284:110-122. https://doi.org/10.1016/j.quaint.2012.04.001
Bianchini, J. J. & J. H. Delupi. 1979. El estado sistemático de los ciervos Neotropicales de la tribu Odocoileinae Simpson, 1945. Physis C 38(94):83-89.
Bouvrain, G., D. Geraads, & Y. Jehenne. 1989. Nouvelles données relatives á la classification des Cervidae (Artiodactyla, Mammalia). Zoologischer Anzeiger 223:82-90.
Breda, M. 2008. Palaeoecology and palaeoethology of the Plio-Pleistocene genus Cervalces (Cervidae, Mammalia) in Eurasia. Journal of Vertebrate Paleontology 28:886-899. https://doi.org/10.1671/0272-4634(2008)28[886:papotp]2.0.co;2
Brooke, V. 1878. On the classification of the Cervidae, with a synopsis of the existing species. Proceedings of the Zoological Society of London 1878:883-928.
Brookes, J. 1828. A Catalogue of the Anatomical and Zoological Museum Joseph Brookes, Esp., part 1:61.
Bubenik, G. A., & K. J. Hundertmark. 2002. Accessory antlers in male Cervidae. Zeitschrift fur Jagdwissenschaft 8:10-21. https://doi.org/10.1007/bf02285353
Cabrera, A. 1929. Sobre los ciervos fósiles sudamericanos llamados Paraceros. Morenelaphus. Memorias de la real Sociedad Española de Historia Natural. 15:155. Madrid.
Cabrera, A. 1941. Cranial and dental characters of some South American Cervidae. Field Museum of Natural History, Chicago 27:125-135.
Cabrera, A. 1943. Sobre la sistemática del venado y su variación individual y geográfica. Revista del Museo de La Plata. Sección Zoología 3:5-41.
Carette, E. 1922. Cérvidos actuales y fósiles de Sudamérica. Revisión de las formas extinguida pampeanas. Revista del Museo de la Plata. 26:393-472.
Cassini, G. H., Muñoz, N. A., & M. L. Merino. 2016. Evolutionary History of South American Artiodactyla. Historia Evolutiva y Paleobiogeográfica de los vertebrados de América del Sur (F. L. Agnolín, G. L. Lio., F. Brissón Egli, N. R. Chimento & F. E. Novas, eds.). Contribuciones del MACN 6:311-322.
Castellanos, A. 1924. Ciervos vivientes y extinguidos de la República Argentina (Nota preliminar). Revista de la Universidad de Córdoba 11:111-134.
Castellanos, A. 1940. Breve reseña sobre estratigrafía pampeana de Santa Fe. Sus amigos a Lucas Kraglievich. Obras de geología y paleontología (L. Kraglievich, ed.). La Plata.
Castellanos, A. 1945. Nota sobre algunas cornamentas de ciervos de los géneros Epieuryceros, Antifer. Paraceros y descripción de la de Epieuryceros proximus n. sp. Publicaciones del Instituto de Fisiografía y Geología de la facultad de Ciencias Físico–Químicas y Naturales aplicadas a la Industria de la Universidad Nacional del Litoral 24:5-23. https://doi.org/10.22179/revmacn.6.71
Castellanos, A. 1957. Nuevos géneros de clamiterio y de ciervo y una nueva especie de Plohophorus. Nota paleontológica. Ameghiniana 1(3):5-14.
Churcher, C. S. 1966. Observaciones sobre el status taxonómico de Epieuryceros Ameghino, 1889 y sus especies E. truncus . E. proximus. Ameghiniana 10:351-362.
Churcher, C. S., & D. Pinsof. 1987. Variation in the antler of North American Cervalces (Mammalia; Cervidae): review of new and previously recorded specimens. Journal of Vertebrate Paleontology 7:373-397. https://doi.org/10.1080/02724634.1988.10011671
Cione, A. L., & E. P. Tonni.1999. Biostratigraphy and chronological scale of uppermost Cenozoic in the Pampean Area, Argentina. Quaternary of South America and Antarctic Peninsula 12:23-52.
Cione, A. L., & E. P. Tonni. 2005. Bioestratigrafía basada en mamíferos del Cenozoico superior de la provincia de Buenos Aires, Argentina. Geología y Recursos Minerales de la Provincia de Buenos Aires. Relatorio del 16º Congreso Geológico Argentino (R.E. de Barrio, R.O. Etcheverry, M.F. Caballé & E. Llambías, eds.). Relatorio: La Plata. Asociación Geológica Argentina. https://doi.org/10.25145/j.pasos.2017.4.009
Cione, A. L., G. M. Gasparini, E. Soibelzon, L. H. Soibelzon, & E. P. Tonni (eds.). 2015. The Great American Biotic Interchange: A South American Perspective. Springer Briefs in Earth System Sciences. https://doi.org/10.1007/978-94-017-9792-43
Croitor, R. 2006. Taxonomy and systematics of large-sized deer of the genus Praemegaceros PORTIS, 1920 (Cervidae, Mammalia). Cour. Forsch.-Inst. Senckenberg 256:91–116.
Cronin, M. A., 1991. Mitochondrial-DNA phylogeny of deer (Cervidae). Journal of Mammalogy 72(3):553-566. https://doi.org/10.2307/1382139
Delupi, L. G. & J. J. Bianchini. 1995a. Morfología comparada de la bulla timpánica en Odocoileus bezoarticus (Linnaeus, 1758) y en otros ciervos sudamericanos. Physis 50:11-21.
Delupi, L. G., & J. J. Bianchini. 1995b. Análisis morfológico comparado de los caracteres craneo-faciales en el ciervo de las pampas Odocoileus bezoarticus y formas relacionadas. Physis 50:23-36.
Duarte, J. M. B. 1997. (Ed.). Biologia e conservação de Cervídeos sul-americanos: Blastocerus, Ozotoceros . Mazama. FUNEP, Jaboticabal 1-238.
Duarte, J. M. B., S. González, & J. E. Maldonado. 2008. The surprising evolutionary history of South American deer. Molecular Phylogenetics and Evolution 49:17-22. https://doi.org/10.1016/j.ympev.2008.07.009
Flueck, W. T., & J. M. Smith-Flueck. 2011. Osteological comparisons of appendicular skeletons: a case study on Patagonian huemul deer and its implications for conservation. Animal Production Science 51:327-339. https://doi.org/10.1071/an10174
Frick, C. 1937. Horned ruminants of North America. Bulletin of American Museum of Natural History 69(XXVIII):1-669.
Garrod, A. H.1877. Notes on the visceral anatomy and osteology of the ruminants, with a suggestion regarding a method of expressing the relations of species by means of formulae. Proceedings of the Zoological Society of London 2-18.
Gervais, H. F. P., & F. Ameghino. 1880. Los mamíferos fósiles de la América del Sud. Igon Hermanos, Buenos Aires.
Gilbert, C., A. Ropiquet, & A. Hassanin. 2006. Mitochondrial and nuclear phylogenies of Cerviae (Mammalia, Ruminantia): systematic morphology and biogeography. Molecular Phylogenetics and Evolution 40:101-117. https://doi.org/10.1016/j.ympev.2006.02.017
Goldfuss, G. A. 1820. Handbuch der Zoologie. Erste Abteilung. Schrag. Nürberg.
González, S., F. Álvarez-Valdin, & J. E. Maldonado. 2002. Morphometric Differentiation of Endangered Pampas Deer (Ozotoceros bezoarticus), with Description of New Subspecies from Uruguay. Journal of Mammalogy 83:1127-1140. https://doi.org/10.1644/1545-1542(2002)083<1127:mdoepd>2.0.co;2
Groves, C. P., & P. Grubb. 1987. Relationships of living deer. Biology and Management of the Cervidae (C. M. Wemmer, ed.). Smithsonian Institution, Washington.
Grubb, P. 2005. Valid and invalid nomenclature of living and fossil deer, Cervidae. Acta Theriologica 45(3):289-307. https://doi.org/10.4098/at.arch.00-30
Hassanin, A., & E. J. P. Douzery. 2003. Molecular and morphological phylogenies of Ruminantia and the alternative position of the Moschidae. Systematic Biology 52(2):206-228. https://doi.org/10.1080/10635150390192726
Hershkovitz, P. 1982. Neotropical deer (Cervidae) Part 1. Pudus, genus Pudu Gray. Fieldiana Zoology. Field Museum of Natural History. New Series 11:1-86. https://doi.org/10.5962/bhl.title.5080
Immel, A. et al. 2015. Mitochondrial Genomes of Giant Deers Suggest their Late Survival in Central Europe. Scientific Reports 5:10853. DOI: 10.1038/srep10853.
Kaiser, T. M., & R. Croitor. 2004. Ecological interpretations of early Pleistocene deer (Mammalia, Cervidae) from Ceyssaguet (Haute-Loire, France). Geodiversitas 26:661-674.
Kolb, C. et al. 2015. Growth in fossil and extant deer and implications for body size and life history evolution. BMC Evolutionary Biology 15:19. doi:10.1186/s12862-015-0295-3
Kraglievich, L. 1932. Contribución al conocimiento de los ciervos fósiles del Uruguay. Anales del Museo de Historia Natural de Montevideo 111:355-438. Montevideo. https://doi.org/10.1215/00182168-18.2.213
Kurtén, B. 1975. A new Plesitocene genus of American mountain deer. Journal of Mammalogy 56:507-508. https://doi.org/10.2307/1379377
Loponte, D. M. 2005. Atlas osteológico de Blastocerus dichotomus (ciervo de los pantanos) Ed. Los Argonautas. Buenos Aires.
Lydekker, R. 1915. Catalogue of the ungulate mammals in the British Museum. Vol. IV. Artiodactyla, families Cervidae (deer), Tragulidae (chevrotains), Camelidae (camels and llamas), Suidae (pigs and peccaries), and Hippopotamidae (hippopotamuses). London, British Museum 438 pp. (Rustees 4). https://doi.org/10.5040/9781472926968.part-0020
Maffei, L. 2001. Estructura de edades de la urina (Mazama gouazoubira) en el chaco boliviano. Mastozoología Neotropical 8:149-155.
Matthew, W. D. 1908. Osteology of Blastomeryx and phylogeny of the American Cervidae. Bulletin of the American Museum of Natural History 24:535-562.
Mattioli, S. 2011. Family Cervidae. Handbook of the Mammals of the World, Vol. 2: Hoofed Mammals (D.E. Wilson & R.A. Mittermeier, eds.). Lynx Edicions, Barcelona, Spain. https://doi.org/10.1002/zoo.21021
Menegaz, A. N. 2000. Los Camélidos y Cérvidos del Cuaternario del Sector Bonaerense de la Región Pampeana. Tesis Doctoral (inédita). Universidad Nacional de La Plata, Argentina. https://doi.org/10.35537/10915/4491
Merino, M. L., N. Mile, & S. F. Vizcaíno. 2005. A cranial morphometric study of deer (Mammalia, Cervidae) from Argentina using three-dimensional landmarks. Acta Theriologica 50(1):91-108. https://doi.org/10.1007/bf03192622
Nicolay, C. W., & M. J. Vaders. 2006. Cranial Suture Complexity in White-Tailed Deer (Odocoileus virginianus). Journal of Morphology 267:841-849.
Nomina Anatomica Veterinaria (NAV). 2005. Prepared by the Internacional Committee on Veterinary Gross Anatomical Nomenclature (ICVGAN) and authorized by the General Assembly of the World Association of Veterinary Anatomists (WAVA), Knoxville, U.S.A., Fifth Edition. http://www.wava-amav.org/Downloads/nav2005.pdf
Norris, C. A. 1999. The cranium of Bunomeryx (Artiodactyla: Homacodontidae) from the upper Eocene Uintia deposits of Utah and its implications for tylopod systematics. Journal of Vertebrate Paleontology 19:742-751. https://doi.org/10.1080/02724634.1999.10011186
O’leary, M. A. 2010. An anatomical and phylogenetic study of the osteology of the petrosal of extant and extinct artiodactylans (Mammalia) and relatives. Bulletin of the American Museum of Natural History 335:1-206. https://doi.org/10.1206/335.1
Onuk, B., M. Kabak, & K. Atalar. 2013. Anatomic and craniometric factors in differentiating roe deer (Capreolus capreolus) from sheep (Ovis aries) and goat (Capra hircus) skulls. Archives of Biological Sciences 65(1):133-141. https://doi.org/10.2298/abs1301141m
Orcesi, M., R. I. Vezzosi, & G. H. Cassini. 2019. Anatomía comparada de los petrosos de Antifer . Morenelaphus (Mammalia, Cervidae). Publicación electrónica de la APA 19(2):64.
Pearson, H. S. 1927. On the skull of early Tertiary Suidae. Together with an account of the otic region in some other primitive Artiodactyla. Philosophical Transactions of the Royal Society of London, serie B 215:389-460. https://doi.org/10.1098/rstb.1927.0009
Pitra, C., J. Fickel, E. Meijaard, & P. C. Groves. 2004. Evolution and phylogeny of old world deer. Molecular Phylogenetics and Evolution 33:880-895. https://doi.org/10.1016/j.ympev.2004.07.013
Pocock R. I. 1912. On the antler growth of the Cervidae, with special reference to Elaphus and Odocoileus .Dorcelaphus). Proceedings of the Zoological Society of London 193-303. Obras completas, II. Art. LXXXIX, 5–204. Londres. https://doi.org/10.1111/j.1469-7998.1912.tb07557.x
Pocock R. I. 1923. On the external characters of Elaphurus, Hydropotes, Pudu, and other Cervidae. Proceedings of the Zoological Society of London 1923:181-207. https://doi.org/10.1111/j.1096-3642.1923.tb02183.x
Popielno, Von Z. J., & C. O. Oberlaufing. 1985. Einiege bermerkungen über die internationalen formeln zur vermessung und berwertung von sumpfhirsch (Blastocerus dichotomus Illiger, 1815) und schomburgkhirsch (Cervus schomburgki Blyth, 1863). Zeitschrift fur Jagdwissenschaft 31:175-184. https://doi.org/10.1007/bf02265746
Putman, R., & W. T. Flueck. 2011. Intraspecific variation in biology and ecology of deer: magnitude and causation. Animal Production Science 51:365-374. https://doi.org/10.1071/an10168
Roth, S. 1904. Noticias preliminares sobre nuevos mamíferos fósiles del Cretácico superior y terciario de la Patagonia. (Apéndice). Revista del Museo de La Plata. XI:158. https://doi.org/10.24215/25456377e046
Rotti, A., N. R. Chimento, R. Pinheiro, G. M. Gasparini, & L. Avilla. 2016. A diversidade de Cervidae (Cetartiodactyla: Mammalia) na América do Sul e o caso de Morenelaphus: um estranho no ninho? X Simpósio Brasileiro de Paleontologia de Vertebrados, Boletim de Resumos: p. 208. https://doi.org/10.12660/gvcasosv5n2c9
Rotti, A., R. I. Vezzosi, D. Mothé, & L. Avilla. 2021. Rising from the ashes: The biggest South American deers (Cetartiodactyla: Cervidae) once roamed Northeast Brazil. Journal of South American Earth Sciences 108:103-154. https://doi.org/10.1016/j.jsames.2021.103154
Rusconi, C. 1936. Anomalías en las cornamentas del Huemul. Anales de la Sociedad Científica Argentina 288-296.
Shi, Q., W. He, & S. Chen. 2014. A new species of Shaanxispira (Bovidae, Artiodactyla) from upper Miocene of China. Zootaxa 3794(4):501-513. https://doi.org/10.11646/zootaxa.3794.4.1
Stehlin, H.G. 1910. Die Säugetiere des schweizerischen Eocaens, Sechster Teil: Catodontherium - Dacrytherium – Leptotheridium– Anoplotherium- Diplobune - Xiphodon - Pseudamphimeryx- Amphimeryx – Dichodon - Haplomeryx - Tapirulus - Gelocus - div. Nachträge.- Abhandlungen der Schweizerischen Palaeontologischen Gesellschaft 36:838-1164. https://doi.org/10.5962/bhl.title.14832
Ubilla, M. 1996. Paleozoología del Cuaternario continental de la Cuenca Norte del Uruguay: Biogeografía, Cronología y aspectos climático-ambientales. Tesis de Doctorado en Paleontología (inédita). Facultad de Ciencias, Universidad de la República, Montevideo, Uruguay. https://doi.org/10.14349/sumapsi.2019.v26.n2.6
Ungerfeld, R., U. T. González-Sierra, & A. Bielli.2008a. Seasonal antler cycle in a herd of pampas deer (Ozotoceros bezoarticus arerunguaensis) in Uruguay. Mammalian Biology 73:388-391. https://doi.org/10.1016/j.mambio.2007.08.006
Ungerfeld, R., A. Bielli, S. X. González-Pensado, S. X., M. Villagrán, & U. T. González-Sierra. 2008b. Antler size and weight in a herd of pampas deer (Ozotoceros bezoarticus). Mammalian Biology 73:478-481. https://doi.org/10.1016/j.mambio.2007.12.004
Valli, A. M. F. 2010. Dispersion of the genus Procapreolus and the relationships between Procapreolus cusanus and the roe deer (Capreolus). Quaternary International 212:80-85. https://doi.org/10.1016/j.quaint.2008.11.002
Van Der Geer, A. A. 2018. Uniformity in variety: Antler morphology and evolution in a predator-free environment. Palaeontologia Electronica 21.1.9A 1-31. https://doi.org/10.26879/834
Van Der Made, J. & H. W. Tong. 2008. Phylogeny of the giant deer with palmate brow tines Megaloceros from west and Sinomegaceros from east Eurasia. Quaternary International 179:135-162. https://doi.org/10.1016/j.quaint.2007.08.017
Vezzosi, R. I. 2015. Diversidad de los mamíferos del Pleistoceno de la Provincia de Santa Fe, Argentina: aspectos taxonómicos, bioestratigráficos y paleobiogeográficos relacionados. Tesis Doctoral (inédita), Universidad Nacional de La Plata. Buenos Aires. Argentina. https://doi.org/10.35537/10915/84888
Vezzosi, R. I., E. Brunetto, & M. C. Zalazar. 2019. El registro fósil de vertebrados del Cuaternario de Santa Fe (Argentina): Primera Actualización Mioceno al Pleistoceno del centro y norte de Argentina (N. Nasiff, G. Esteban, J. Chiesa, A. Zurita & S. Georgieff, eds.). Opera Lillioana 52. Capítulo 3: Pleistoceno. Tucumán, Argentina. https://doi.org/10.35537/10915/84888
Vezzosi, R. I., A. Rotti, & L. Avilla, L. 2020. ‘A very lazy deer’: Revision of the cotype of Nothropus carcaranensis (Mammalia, Xenarthra). Ameghiniana.
Vislobokova, I. A. 2012. Giant Deer: Origin, Evolution, Role in the Biosphere. Paleontological Journal 46(7):643-775. https://doi.org/10.1134/s0031030112070027
Vislobokova, I. A. 2013. Morphology, Taxonomy, and Phylogeny of Megacerines (Megacerini, Cervidae, Artiodactyla). Paleontological Journal 47(8):833-950. https://doi.org/10.1134/s0031030113080017
Webb, S. D. 1992. A cranium of Navahocerus and its phylogenetic place among New World Cervidae. Annales Zoologici Fennici 28:401-401.
Webb, S. D. 2000. Evoutionary history of new world deer. Anelopes, Deer, and Relatives. (E. S. Vrab & G. B. Shaller, eds.). Yale University Press, London.
Woodburne, M. O. 2010. The Great American Biotic Interchange: Dispersals, Tectonics, Climate, Sea Level and Holding Pens. Journal of Mammalian Evolution 17:245-264. https://doi.org/10.1007/s10914-010-9144-8
Información adicional
Editor asociado: Aldo Iván Vassallo
Citar como: Vezzosi, R. I. & N. R. Chimento. 2021. Sobre la anatomía craneana de Morenelaphus Carette, 1922 (Mammalia, Cervidae) y formas relacionadas: descripción y comparación morfológica del área del basicráneo. Mastozoología Neotropical, 28(2):e0579. https://doi.org/10.31687/saremMN.21.28.2.0.01.e0579