ARTÍCULO
ECOLOGICAL ASPECTS OF Dasypus pastasae IN THE COLOMBIAN ALTILLANURA, WITH COMMENTS ON ACTIVITY PATTERNS AND ITS IDENTIFICATION VIA CAMERA TRAPS
ASPECTOS ECOLÓGICOS DE DASYPUS PASTASAE EN LA ALTILLANURA COLOMBIANA, CON COMENTARIOS SOBRE PATRONES DE ACTIVIDAD E IDENTIFICACIÓN EN CÁMARAS TRAMPA
ECOLOGICAL ASPECTS OF Dasypus pastasae IN THE COLOMBIAN ALTILLANURA, WITH COMMENTS ON ACTIVITY PATTERNS AND ITS IDENTIFICATION VIA CAMERA TRAPS
Mastozoología Neotropical, vol. 28, núm. 2, 2021
Sociedad Argentina para el Estudio de los Mamíferos
Recepción: 31 Diciembre 2020
Aprobación: 26 Agosto 2021
Abstract: The Pastaza greater long-nosed armadillo (Dasypus pastasae) is a recently revalidated species and therefore poorly studied from South America. The natural history of this species is virtually unknown in the wild. Here, we describe ecological aspects of D. pastasae including activity patterns, behavioral notes, endoparasite records, and diet information retrieved from riparian forest populations in the eastern plains of Colombia. Additionally, we point out key morphological traits to differentiate D. pastasae from other congeneric armadillos present in Colombia based on camera trap records.
Keywords: Armadillos, Colombia, diet, eastern plains, endoparasites, riparian forest.
Resumen: El armadillo espuelón (Dasypus pastasae) es una especie recientemente revalidada y poco estudiada de América del Sur. La historia natural del espuelón es prácticamente desconocida y son muy escasos los datos de esta especie en vida silvestre. El presente trabajo describe diversos aspectos sobre la historia natural de esta especie, incluyendo patrones de actividad, notas sobre su comportamiento, endoparásitos, así como anotaciones sobre su dieta de poblaciones en bosques riparios de los Llanos orientales en Colombia. Además, destacamos aspectos de su morfología, útiles para diferenciar D. pastasae de otros armadillos presentes en Colombia basados en registros de cámaras trampa.
Palabras clave: Armadillos, bosques riparios, Colombia, dieta, endoparásitos, Llanos orientales.
INTRODUCTION
The Pastaza greater long-nosed armadillo, Dasypus pastasae (Thomas, 1901), is the second-largest armadillo that inhabits Colombia, with a total length up to one meter and a weight between 5.9 and 9.8 kg (Feijó et al. 2018). This species was recently revalidated (Feijó & Cordeiro-Estrela 2016; Feijó et al. 2018, 2019) and therefore its conservation status has not yet been evaluated at the national or international levels. Dasypus pastasae is restricted to the western Amazon region of South America, from the foothills of the eastern Andes in Peru, Ecuador, Colombia, and Venezuela, and south of the Orinoco River to the west of the Brazilian Amazon (Feijó & Cordeiro-Estrela 2016). In Colombia, the species is known from the Orinoquia and Amazonia regions in an elevational range from sea level to 1175 m a.s.l. About 80% of its records come from the Meta department (Aya-Cuero et al. 2019a), with additional records from the departments of Nariño, Amazonas, Caquetá, Casanare, Putumayo, and Vichada (Aya-Cuero et al. 2019a).
Natural history information about D. pastasae is scarce (Aya-Cuero et al. 2019b). Field studies are thus crucial to provide basic information on population size, habitat requirements, activity patterns, diet, and main threats to support proper conservation plans (Rodríguez-Mahecha et al. 2006; Superina et al. 2014). Previous studies on D. pastasae’s ecology in Colombian ecosystems have described that they use P. maximus’s burrows (Aya-Cuero et al. 2017), display a leaf-dragging behavior (Aya-Cuero 2016), and have a diet composed mainly of Hemiptera, Coleoptera, Hymenoptera, and Gymnophyona (Barreto et al. 1985). A specimen of D. pastasae was also found to be infected by Trypanosoma cruzi (D’alessandro et al. 1984).
One of the great challenges for ecological studies of D. pastasae is the low encounter rate with individuals in the field due to their nocturnal and evasive habits. For instance, this species was photographed only once in Colombia at Mururito reserve, Puerto Gaitán (Emilio Constantino, pers. comm.). The use of camera traps can thus be an important tool to generate better understanding of the natural history of D. pastasae. However, species identification from photographs and videos can be misleading due to morphological similarities among congeneric species (Johansson et al. 2020). In fact, the difficulty to distinguish species via camera trap records has led some authors to combine all long-nosed armadillos in one group (Dasypus spp.) (Aya-Cuero et al. 2019b).
In Colombia, in addition to D. pastasae, two other congeneric species can be found, the nine-banded armadillo Dasypus novemcinctus Linnaeus, 1758, and the Northern long-nosed Armadillo Dasypus sabanicola Mondolfi, 1968. In the Colombian Orinoquia, all three Dasypus species are present, while in the Amazon portion, D. pastasae and D. novemcinctus occur in sympatry (Feijó et al. 2018; Aya-Cuero et al. 2019a). Therefore, recognizing the diagnostic characteristics that allow proper identification of the Dasypus species from camera trap records is essential to reveal the ecology of the cryptic Pastaza greater long-nosed armadillo.
Here, we report novel information on the natural history of D. pastasae obtained from camera-trapping and field surveys carried out in the Colombian Altillanura. Data on activity patterns, diet, endoparasites, and some behavioral notes are described. Finally, we highlight the main characteristics that are useful to differentiate D. novemcinctus from D. pastasae in images and videos obtained from camera traps.
MATERIALS AND METHODS
Study Area
The study was carried out at six sites in the Colombian Altillanura, located in the eastern plains of Colombia. This region represents a vast landscape including the departments of Meta and Vichada, with floodplain and non-floodable riparian forest and small wooded formations that grow in the middle of the stational savannahs, covering an area of 86211 km2, about 7.5% of the Colombia territory (Serrato 2018). Three sites were in the department of Meta:
(1) Puerto López, near the Yucao River (4º00’N; -72º00’ W); Puerto Gaitán, near the Planas River (4º00’N; -71º49’W); and (3) La Macarena, near Playa Rica (2º03’N; -74º15’W). The remaining three sites were in the department of Vichada in the municipalities of: (1) La Primavera (5º00’N; -70º00’W); and Puerto Carreño, near the (2) Bita (5º48’N -68º55’W) and (3) Gavilán (5º00’N; -69º00’W; Fig. 1) rivers. The average elevation at all sites was 200 m a.s.l., with an average annual temperature of 26 ºC. There are well-marked dry (from December to March) and rainy seasons (Cicery et al. 2005). Soils have low fertility, with high acidity and aluminum saturation (Lavelle 2014). The study areas were characterized by soft slopes, with natural savannas and riparian forests as the predominant vegetation (Botero & Serrano 2019). There were relatively low anthropic impacts, primarily from livestock and a mosaic of crops.
Camera trap survey
Five camera trapping surveys were carried out in the Colombian Altillanura encompassing two departments (Fig. 1): Department of Meta: The first included ten camera traps installed for eight months from February to August 2014 in the Planas River, Puerto Gaitán, Meta. The second survey occurred in Playa Rica, Meta, with six cameras from July to December 2017 (rainy season). The third survey occurred at two sites simultaneously (Yucao River, Puerto López and Planas River, Puerto Gaitán, Meta; separated by almost 76 km) using 11 cameras at each site from July to October 2019, once again during the rainy season.
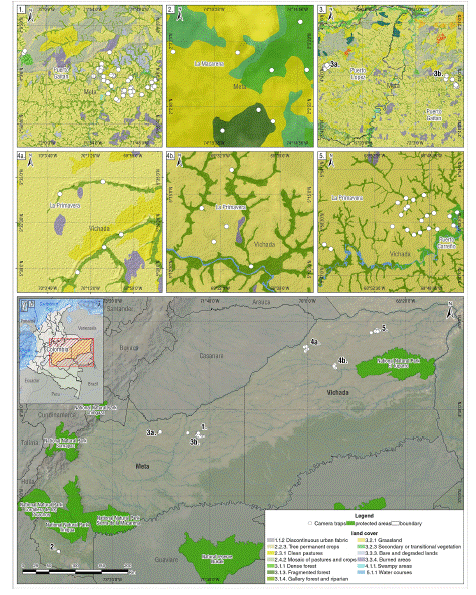
Fig. 1
Surveys in the five localities of the Colombian Altillanura. Circles represent the camera traps installed in the sampling station. The installation follows the riparian forest cover in the departments of Meta and Vichada.
Department of Vichada: The fourth survey took place at Matiyure and Buenavista, separated by almost 62 km in the municipality of La Primavera near to Bita River and Gavilán River respectively, Vichada, for eight months (November 2018 to July 2019; dry and rainy season), using five cameras at each locality. Finally, the fifth survey took place in another location along the Bita River on private properties of Puerto Carreño and La Primavera, Vichada. This survey lasted five months (November 2018 to March 2019: dry and rainy season) and used 21 camera traps (Fig. 1; Table 1).
At all sampled sites, camera traps were programmed in hybrid mode to take sequences of three pictures and one video of 30 sec and operated 24 hours a day. In total there were 116 sampling stations (Table 1). The camera traps were installed mainly at the entrances of armadillo burrows (n=73) and to a lesser extent along trails (n=43), with exception for the second and fourth surveys where the cameras were installed on trails only.
The sampling effort was calculated by multiplying the number of camera traps by the number of effective sampling days (Lira-Torres & Briones-Salas 2012). It is noteworthy that in some cases the sampling effort was lower than expected because some camera traps stopped working due to electronic failures such as in the first survey. In the first and second surveys the camera traps were translocated among different sites of the sampling stations to cover a major area. Capture effort was measured individually for each camera during the time each was operational.
Armadillo species were identified to species according to the characters described by Feijó & Cordeiro-Estrela (2016) and Feijó et al. (2018). Some of the external characters that allowed us to differentiate D. pastasae from D. novemcinctus (the two most similar species that occur in sympatry) were: (1) D. pastasae has well-developed scales at the knee (approximately 3 cm long), (2) the number of movable bands varies from 7-8, while D. novemcinctus in Colombia usually has 9 bands (ranging from 8-10), (3) the snout of D. pastasae is longer and more robust than of D. novemcinctus (4) a small portion of the unsegmented part of the tail, (5) the occipital lobe of the cephalic shield is absent in D. pastasae, leaving a noticeable gap between the cephalic shield and ears, (6) the cephalic shield does not cover the anterior part of the snout, and (7) the tail is more robust in relation to D. novemcinctus (Fig. 3). When observed, the penis of D. pastasae shows a distinct terminal blunt-ended projection; both D. novemcinctus and D. sabanicola have a terminal trifid form (Wetzel & Mondolfi 1979).
Activity patterns and relative abundance
The activity pattern for D. pastasae was determined from the camera trap records and calculated via circular kernel probability density with 1000 bootstrap replicates using the fitact function in activity R package (Rowcliffe 2021). We consider as independent records those obtained after at least a 2-hour interval.
The relative abundance index (RAI) was estimated using the number of independent records of D. pastasae per survey divided by the total number of records of mammal species obtained in each trap survey (Díaz-Pulido & Payán-Garrido 2012). RAI was used to compare abundance across sampled sites.
Ethnozoology
In a complementary approach, to obtain additional information about local knowledge and use on armadillos in our study area, we conducted semi-structured interviews with 45 people from the Planas River trails between February 8 and August 14, 2014. Some of the questions include: Do you know armadillos? When was the last time you saw an armadillo? Which of the five armadillo species present in the eastern plains have you seen or know? What uses do they give to armadillos? What do you consider to be the main threats to armadillos? Have you ever hunted an armadillo for consumption? In addition, we also took into account our own observations in the field to make inferences about the local use of D. pastasae and threats to this species.
Opportunistic collections
During the field surveys we encountered four dead Pastaza greater long-nosed armadillos. Three were killed by local hunters using dogs and “trampero” (automatic guns triggered by the movement of animals) installed along trails, and the other one was road killed. Parts of these specimens (stomach content, carapace, skull) were collected and housed in different Colombian natural history museums (see results). It was possible to preserve (in 70% alcohol) the stomach contents from one individual found in Playa Rica, La Macarena. The contents were then completely examined to determine the diet and presence of endoparasites. Identification of invertebrates in the diet was conducted in the Entomology Laboratory of the Kumangui Arthropod Research Group, Universidad Distrital Francisco José de Caldas, using a Zeiss Stemi 305 stereoscopic microscope, following the taxonomic keys and descriptions provided in Constantino (2002), Johnson & Triplehorn (2005), Hoffman (2005) and Fernández (2006). The items of diet were presented by percentage of individuals consumed following Barreto et al. (1985) and Szeplaki et al. (1988). Endoparasites were processed with Aman’s lactophenol and washed out through changes of ethanol (Kennedy 1979).
RESULTS
With a total sampling effort of 7971 camera trap/days, we obtained 2407 records of mammals (Table 2). Of 185 Dasypus individuals recorded in this study, 119 could be identified to the species level, including 102 (4.23%) D. pastasae, 7 D. sabanicola, and 10 D. novemcinctus. Of the D. pastasae records, 83 were from the department of Meta and 19 from Vichada.
Activity pattern
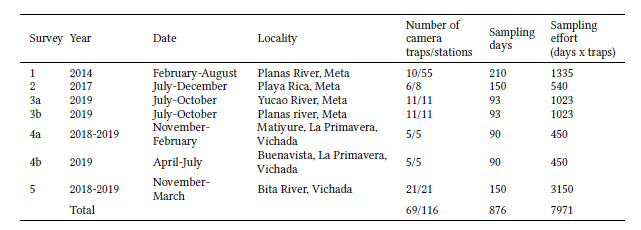
The 102 records of D. pastasae reveal that they are nocturnal (Fig. 2a), with higher activity between 20:00 and 3:00 hours. The only record obtained during daylight (at 14:47 on May 11, 2014, at the beginning of the rainy season) was of two individuals inspecting a burrow of P. maximus at the same time. Additionally, most of the records occurred in May and July during the middle of the rainy season, while in the driest months (November to March) records were scarce (Fig. 2b).
Relative abundance
The relative abundance index of D. pastasae was low for all sampling sites and varied from 0.025 at La Primavera, Vichada, to 0.208 at Puerto Gaitán, Meta (Table 2).
Diet and endoparasites
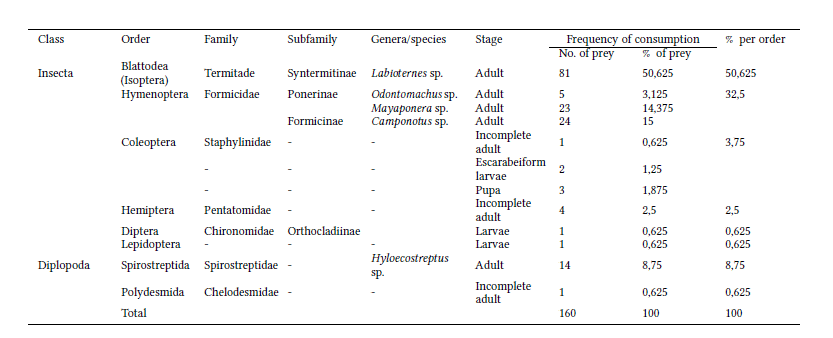
The stomach contents of the D. pastasae specimen from Playa Rica included only arthropods (160 individuals), most of them belonging to the Insecta (76%) and Diplopoda (24%) classes. The main food items are termites (representing 50.6% of the individuals consumed) of the genus Labiotermes, followed by ants (32.5%) of the genera Camponotus, Mayaponera, and Odontomachus. Millipedes (8.7%) of the orders Polydesmida and Spirostreptida were also found. Millipedes of the medium-sized genus Hyloecostreptus (length up to 6 cm) were the largest ingested arthropods. Other insects in smaller numbers included bedbugs Pentatomidae (2.5%), adult Staphylinidae beetles (0.6%), larval and pupal beetles (3.1%), chironomids of the Orthocladiinae subfamily (0.6%), and caterpillars (0.6%; Table 3). While analyzing the stomach contents, we also found seven nematode individuals of the genus Spirurida, Physalopteridae family.
Behavioral notes
During camera trapping, we obtained nine independent records of mud bathing between August and October (end of the rainy season), mainly at the entrance of a P. maximus burrow, but also in a small hole in the ground. During these baths, the armadillo laid on its side or back and began to shake the body with small abdominal contractions. The baths lasted an average of 67 seconds and occurred between 19:00 and 02:00. Other behaviors observed during these events included scratching the forearm with the rear leg and shaking the ears https://www.youtube.com/watch?v=PRt56S3VSjY; https://www.youtube.com/watch?v=gRdPQosgbw4.
Opportunistic collections
In 2014, we found a female D. pastasae individual that had been hunted for human consumption in vereda Alto Manacacías. Its carapace is now housed in the Museo de la Universidad Distrital FJC under catalog number MUD 807. In the same year, another dead individual was a roadkill on a tertiary road that connects Puerto Gaitán city with Campo Rubiales. This animal was found in a savanna area close (200 m distance) to a riparian forest. Its skull is housed in the Museo de la Salle, Bogotá under catalog number MLS2430.
In 2017 and 2018, as part of fauna monitoring with a group of Farc ex-combatants (Aya-Cuero et al. 2020), two individuals of D. pastasae were opportunistically found to have been hunted for illicit commercial trade and subsistence consumption in the Municipality of La Macarena. The first one was being exhibited for sale in La Julia, department of Meta (2º56’N, -74º13’W) along with another armadillo (D. novemcinctus). The second animal was hunted in the Playa Rica village (2 º00’N, -74 º17’W) by local inhabitants with dogs at night on March 14, 2018. From this individual, we collected the cephalic shield, now deposited at Museo de la Universidad de Caldas under catalogue number MHN-UCa 2694, and the stomach contents analyzed in this study.
Ethnozoological notes
Of the 45 people interviewed, only 4 (9%) recognized D. pastasae as a distinct species, making it the species of armadillo least known by the community. According to the interviewers, dogs, hunting and environmental contamination (e.g. burning and dumping garbage in natural areas) are the main threats to armadillos in general. Among the main human uses of armadillos, consumption (52%) and medicinal uses (40%) were the most common, followed by sport hunting (5%) and pets (3%).
In general, through interviewer’s information, we documented that after being hunted, armadillos are eviscerated, and the epidermal scales are removed by exposing the carapace to fire or hot water until obtained a white appearance. The preparation of game meat varies from roasting in the shell and cooked with toppings. About 49% of the people interviewed had a carapace in their house. The carapace of D. pastasae is valued as a hunting trophy due to its size. We observed a carapace of a hunted animal in vereda Alto Manacacías had been prepared and dried with ash and was displayed on the wall of the hunter’s house. Other parts commonly kept at the hunter’s house is the unsegmented portion of the tail (33%).
DISCUSSION
Despite a considerable sampling effort, we obtained relatively few records of D. pastasae in relation to other mammals, which confirms the difficulty of recording this species even with camera traps (Pardo-Vargas & Payán-Garrido 2015). Previous studies with camera traps that focused on medium and large mammals in the Colombian Orinoquia either failed to detect D. pastasae or recorded it at a low rate. For instance, this species was not registered in Guayupes forest (Rodríguez-Bolaños et al. 2014), areas near the Planas and Tillavá rivers (Rodríguez-Bolaños et al.2015), savanna ecosystems (Castillo-Figueroa et al.2019), in oil palm cropland (Pardo-Vargas et al. 2019),or on the university campus of UNILLANOS (Alfonso & Sánchez 2020). In a recent study, Mosquera-Guerra et al. (2018) reported only one record of D. pastasae, obtained during the dry season in a location near the Bita River (Vichada), even with a sampling effort of 7700 camera-trap/nights. Their study was carried out in the principal gallery forest of the Bita River, which is more prone to flooding, in contrast to our survey, which sampled riparian forests along tributaries of the Bita River. Our data thus highlights the importance of riparian forests for the conservation of D. pastasae in the eastern plains. In fact, all our D. pastasae records were obtained in riparian forest except for one individual that was found road killed in an open savanna area. The latter may suggest that D. pastasae can use savannas to move between riparian forests. Nevertheless, because of its strong association with forested areas, D. pastasae may be vulnerable to ongoing land use change (Superina et al. 2019).
In the Orinoquia region, the Altillanura (departments of Meta and Vichada) is crucial for the conservation of the D. pastasae populations, because with the current information its presence in the department of Casanare is scarce and in the department of Arauca unrecorded. Some differences between the floodable savannas (Casanare and Arauca) and the Altillanura are the more flat relief and their major capacity for retaining water (Serrato 2018).
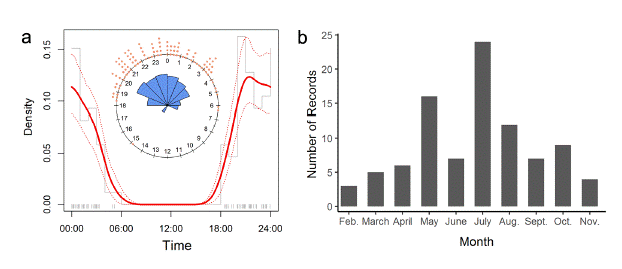
Fig. 2
Activity patterns of Dasypus pastasae in Colombian Altillanura. (a) Kernel density plot showing the daily activity. Dashed lines represent the confidence limits. Insert circular plot shows the raw records per time (orange dots) and the relative frequency (blue polygons). (b) Number of records across months of D. pastasae during the camera trap surveys in Meta (2014 and 2019) and Vichada (2018 and 2019) departments.
In the Armadillo Conservation Program of Eastern Plains, D. pastasae was the least frequently observed and reported in Meta and Casanare (Superina et al. 2019). On the other hand, in a field effort focused on armadillo species in the department of Arauca, Rodríguez-Durán et al. (2018) did not report any sightings of this species in a one-year field study, and this species was the least mentioned in interviews, indicating that it occurs, if at all, at low population densities.
An interesting result is that D. pastasae was the most recorded species of the genus Dasypus that could be confidently identified in our study area (see Table 2). This result contrasts with the large number of records of D. novemcinctus reported in previous studies of the Orinoco ecoregion and in which D. pastasae was little recorded (Trujillo & Superina 2013; Superina et al. 2019). This difference might be explained in two complementary ways: first, while D. novemcinctus can use both open and forested areas, D. pastasae seems to be more restricted to forested biomes, which might explain its relative higher abundance in riparian forests. Additionally, we hypothesized that D. pastasae requires more pristine areas and has low tolerance to anthropic alteration, as has been suggested for D. beniensis, a sister species of the D. kappleri complex that were less abundant in tourist areas (Rocha et al. 2012). Second, studies may have misidentified Dasypus species, leading to an inflation of D. novemcinctus records, the most widespread species of the genus. On the other hand, the overall low detectability of Pastaza armadillos might be related to their low density population, similarly to P. maximus whose sightings are inherently rare (Carter et al. 2015; Aya-Cuero et al. 2017).
Activity patterns
Our study is the first to describe the activity patterns of D. pastasae, revealing its nocturnal habits that are similar to those of other medium-to-large Dasypus species (e.g. D. kappleri, D. novemcinctus, Loughry & McDonough 2013; Aya-Cuero et al. 2019b). The nocturnal behavior of D. pastasae was also mentioned by the indigenous Matses people of northeastern Peru (Fleck & Voss 2016). Our only camera trap record of diurnal activity in D. pastasae may have been associated with the reproductive season because two individuals were recorded together. The same behavior (two individuals foraging together during daylight) was also observed for the nocturnal P. maximus by Carlos Aya in December of 2019 at Puerto Gaitán, Meta. Furthermore, the fact that the majority of our records of D. pastasae were obtained during the rainy season might be partially an artifact of our greater sampling effort during this season or because the duration of the rainy season is much longer (nine months) than that of the dry season (three months).
Relative abundance
The increase of records of D. pastasae through camera-trapping seems to be related to the installation of cameras in front of the entrance of large armadillo burrows and longtime surveys. In fact, a three-year monitoring focused on armadillos in Planas River obtained a higher RAI for D. pastasae than reported here (Aya-Cuero et al. 2019c). This relative higher abundance of D. pastasae in Planas River private areas might be related to the hunting restriction, limited use of fire, and well-conserved riparian forests. On the other hand, the area with the lowest RAI is located about 5 km of the rural town Playa Rica. This area has few conserved forest patches and is often used by local communities that have small crops, occasionally use fire, and actively hunting armadillos.

Fig. 3
External diagnostic traits to differentiate D. pastasae (left) from D. novemcinctus (right) from camera trap records. (a) Large segmented/unsegmented portion of the tail ratio and number of movable bands of the carapace (b) robust and long face, (c) well-developed scales on the knee of the hind legs, (d) more robust tail, (e) less developed occipital lobe, leaving a clear gap between the cephalic shield and ears.
Dietary characterization
Dasypus pastasae is considered an opportunistic insectivore that feeds on a wide range of invertebrates (coleopterans, arachnids, earthworms, myriapods) and even some vertebrates, as caecilians have been recorded in its diet (Aya-Cuero et al. 2019b). However, this classification is based on the stomach contents of only one individual from Colombia (Barreto et al. 1985) and four individuals from Venezuela (Szeplaki et al. 1988). Reports from indigenous hunters in northeastern Peru mention palm fruits (Oenocarpus bataua and Mauritia flexuosa) as part of the diet (Fleck & Voss 2016). Aya-Cuero et al. (2019b) noted this species does not have preference for ants and termites, which is in clear contrast to our results where termites and ants dominated (∼83%) the stomach contents of the Playa Rica animal. Additional studies involving multiple individuals are necessary to better understand the diet preference of this species.
Behavior notes
Our study records for the first time the mud bathing behavior of D. pastasae, which has been mentioned by the indigenous Matses people (Fleck & Voss 2016). In D. novemcinctus, this behavior was recorded mainly on hot, dry summer days in North America, and thus associated with thermoregulation (Loughry & McDonough 2013). However, all nine records of mud bathing in D. pastasae occurred at the end of the rainy season, when the monthly temperature range varied from 24.2–27.5 °C (IGAC 2021). Loughry & McDonough (2013) also hypothesized that the mud could reduce insect biting, which seems a more reasonable explanation for our findings because the highest abundance of insects in tropical forests occurs during the rainy season (Abella-Medrano et al. 2015). On the other hand, a mud bath could also have negative effects. This behavior might expose armadillos to pathogens carried by wild pigs (Eckert et al. 2019), which were found occurring in sympatry with Pastaza armadillos at Planas River (Aya-Cuero et al. 2019c). Similarly, the use of giant armadillo burrows by D. pastasae and many other vertebrates might represent another source of potential pathogenic infection (Desbiez & Kluyber 2013).
The duration of mud baths can be longer than estimated in our study. In camera trap monitoring carried out at Mururito Natural Reserve in Puerto Gaitán, Meta, this behavior was observed in August 2013 and the estimated duration was 168 seconds (unpublished data).
Curiously, from the unique encounter known in Colombia with a live individual, Emilio Constantino observed an animal displaying a death-feigning or thanatosis behavior. According to Humphreys & Ruxton (2018), antipredatory defenses are crucial to many aspects of behavioral ecology and it is known that armadillos might remain immobile to reduce the risk of predator attack, and they also adopt a curled posture that protects vulnerable.
Ethnozoology notes
Communities play a key role in providing natural history information about rare species. For example, Fleck & Voss (2016) summarized novel information about D. pastasae natural history (e.g. behavior, reproduction, diet, and others) based on the Peruvian Matses people’s knowledge and showed that only 21% overlapped with the information available in the literature. According to the information we obtained from local Colombian communities, D. pastasae is probably one of the rarest armadillo species of the eastern plains. Long-nosed armadillos have a high value for local communities as a food source. Although D. pastasae is not widely recognized as a distinct species, it is hunted and may be particularly prized because it provides a greater amount of protein than other species of armadillos that are frequently consumed, such as the nine-banded armadillo and the llanos long-nosed armadillo. On the medicinal uses, the blood is considered as a medicine for respiratory diseases, and this perception has been documented for other congeneric species. According to Rodrigues et al. (2020) at least 11 species of armadillos have been reported for medicinal purposes. The carapace and the unsegmented portion of the tail kept by local people are important sources of information about the frequency of the hunting, species consumption and other topics. They can also be a valuable source of tissue samples for genetic studies (Trujillo & Superina 2013).
Endoparasites
Parasites are a biotic factor that affects the dynamics and density of host populations (Tompkins et al. 2002). The endoparasite present in D. pastasae, Spirurida, is commonly found in marsupials and other mammals such as Herpailurus yaguaroundi, Sapajus apella and Tamandua tetradactyla (Ramos et al. 2016), but not in Cingulata. More studies are needed to understand the parasites associated with armadillos.
Phototrapping identification
Our study shows that species-level identification of long-nosed armadillos via camera trap records is possible in most cases; nevertheless, about one-third of the records could not be identified to species. The use of complementary diagnostic traits (see Fig. 3) is key to distinguish Dasypus pastasae from D. novemcinctus. It is noteworthy that the traits applied here could also be used to differentiate D. novemcinctus from the two other species of the Hyperoambon subgenus, Dasypus beniensis and Dasypus kappleri. We note that tail traits, rostral appearance and the number of mobile bands are the most easily recorded features in camera traps. The knee spurs, cephalic shield and penis are less often visible in camera traps; nevertheless, when recorded, they allow a clear distinction between the Dasypus species in Colombia. We do not include D. sabanicola in this comparison because of its low camera trap records obtained in this study. However, D. sabanicola can be easily differentiated from D. pastasae by its smaller size (head-body length 269-314 mm) (Feijó et al. 2018). Additionally, D. sabanicola is mostly associated with open areas of the Llanos and therefore has low overlap with the more forested areas where D. pastasae is usually found (Mondolfi 1968; Feijó et al. 2018; Chacón-Pacheco et al. 2020).
CONCLUSIONS
Our camera trap surveys demonstrated the importance of photo-trapping in facilitating the study of cryptic species, in this case the Pastaza armadillo. Our results suggest a strong association between D. pastasae and the riparian forest of the Colombian Altillanura and conclude that its detectability increases when cameras are placed in front of armadillo burrows. Our study provides important information about the ecology of this species that should help to promote conservation actions, for instance, by understanding the areas used more frequently by D. pastasae. Finally, by correctly identifying this species during wildlife monitoring it should be possible to obtain even more information about this rare armadillo.
Acknowledgments
This study was founded by a cooperation agreement between Fundación Omacha and Fundación Cunaguaro. We thank Keiler Tellez for their collaboration during the fieldwork in La Macarena, Meta. Cecilia Esquiaga for her help in the identification of endoparasites, and Andrea Ospina for the collaboration to compare images of D. novemcinctus with those of D. pastasae. William J. Loughry and Nina Attias provided valuable suggestions. Finally, we thank ODL-BICENTENARIO pipelines for economic support of the fieldwork in Puerto Gaitán and Puerto Lopez during 2019. CAC thanks to the Armadillo Conservation Program of the Eastern Plains liderated by Fundación Omacha for economic support of the fieldwork in Puerto Gaitán during 2014 and 2019. DAE thanks Coordenação de Aperfeiçoamento de Pessoal de Nível Superior – Brazil (CAPES) – Finance Code 88882.439397/2019-01 (Master Degree). AF is supported by the Second Tibetan Plateau Scientific Expedition and Research Program (No. 2019QZKK0402, 2019QZKK0501) and Chinese Academy of Sciences President’s International Fellowship Initiative (Grant 2021PB0021).
REFERENCES
Abella-Medrano, C. A., S. Ibáñez-Bernal, I. Macgregor-Fors, & D. Santiago-Alarcon. 2015. Spatiotemporal variation of mosquito diversity (Diptera: Culicidae) at places with different land-use types within a neotropical montane cloud forest matrix. Parasites Vectors 8:487. https://doi.org/10.1186/s13071-015-1086-9
Alfonso, C., & F. Sánchez. 2020. Mamíferos no voladores en un campus universitario de la Orinoquia colombiana. Mammalogy Notes 5:29-39. https://doi.org/10.47603/manovol5n2.29-39
Aya-Cuero, C. 2016. Transporte de material vegetal por el armadillo espuelón Dasypus kappleri Krauss, 1862 para la construcción de nido en un bosque de galería de los Llanos Orientales de Colombia. Edentata 17:57-60. https://doi.org/10.2305/iucn.ch.2016.edentata-17-1.10.en
Aya-Cuero, C., A. Rodríguez-Bolaños, & M. Superina. 2017. Population density, activity patterns, and ecological importance of giant armadillos (Priodontes maximus) in Colombia. Journal of Mammalogy 98:770-778. https://doi.org/10.1093/jmammal/gyx006
Aya-Cuero, C. et al. 2019a. Distribution of armadillos in Colombia, with an analysis of ecoregions and protected areas inhabited. Mammal Research 64:569-580. https://doi.org/10.1007/s13364-019-00435-4
Aya-Cuero, C., J. Chacón-Pacheco, T. C. S. Anacleto. 2019b. Dasypus kappleri (Cingulata: Dasypodidae). Mammalian Species 51:51-60. https://doi.org/10.1093/mspecies/sez009
Aya-Cuero, C., F. Mosquera-Guerra, D. A. Esquivel, F. Trujillo, & D. Brooks. 2019c. Medium and large mammals of the mid Planas River basin, Colombia. Biota Colombiana 20:76-92. https://doi.org/10.21068/c2019.v20n02a06
Aya-Cuero, C., C. Díaz-B, & D. A. Esquivel. 2020. Mamíferos medianos y grandes del Espacio Territorial de Capacitación y Reincorporación (ETCR) Playa Rica, La Macarena, Meta, Colombia. Mammalogy Notes 6:1-7. https://doi.org/10.47603/mano.v6n2.188
Barreto, M., P. Barreto, & A. D’alessandro. 1985. Colombian armadillos: stomach contents and infection with Trypanosoma cruzi. Journal of Mammalogy 66:188-193. https://doi.org/10.2307/1380985
Botero, P., & H. Serrano. 2019. Caracterización Biofísica del área de ampliación, geología, geomorfología y suelos. Expediciones científicas en las áreas del PNN Serranía del Chiribiquete (A. Vásquez, ed.). Fundación para la Conservación y Desarrollo Sostenible. Bogotá, D.C., Colombia.
Carter, T. S., M. Superina, & D. M. Leslie. 2015. priodontes Maximus (cingulata: Chlamyphoridae). Mammalian Species https://doi.org/10.1093/mspecies/sew002
Castillo-Figueroa, D., D. Martínez-Medina, M. E. Rodríguez-Posada, & S. Bernal. 2019. Structural differences in mammal assemblages between savanna ecosystems of the Colombian Llanos. Papeis Avulsos de Zoologia 59:1-11. https://doi.org/10.11606/1807-0205/2019.59.14
Chacón-Pacheco, J., C. Aya-Cuero, T. C. S. Anacleto. 2020. Dasypus sabanicola (Cingulata: Dasypodidae). Mammalian Species 52:49-56. https://doi.org/10.1093/mspecies/seaa004
Cicery, E., L. Gualtero, & R. E. Cicery. 2005. Monografía del municipio de Puerto Gaitán, visión geohistórica y cultural. http://www.puertogaitan-meta.gov.co
Constantino, R. 2002. An illustrated key to Neotropical termite genera (Insecta: Isoptera) based primarily on soldiers. Zootaxa 67:1-40. https://doi.org/10.11646/zootaxa.67.1.1
D’alessandro, A., P. Barreto, N. Saravia, & M. Barreto. 1984. Epidemiology of Trypanosoma cruzi in the oriental plains of Colombia. The American Journal of Tropical Medicine and Hygiene 33:1084-1095. https://doi.org/10.4269/ajtmh.1984.33.1084
Desbiez, A. L. J., & D. Kluyber. 2013. The role of giant armadillos (Priodontes maximus) as physical ecosystem engineers. Biotropica 45:537-540. https://doi.org/10.1111/btp.12052
Díaz-Pulido, A., & E. Payán-Garrido (eds.). 2012. Manual de foto-trampeo: una herramienta de investigación para la conservación de la biodiversidad en Colombia. Instituto de Investigaciones de Recursos Biológicos Alexander von Humboldt y Panthera Colombia. Bogotá D.C., Colombia. https://doi.org/10.21068/a2020rrhhxix
Eckert, K., D. Keiter, & J. Beasley. 2019. Animal visitation to Wild Pig (Sus scrofa) wallows and implications for disease transmission. Journal of Wildlife Diseases 55:488-493. https://doi.org/10.7589/2018-05-143
Feijó, A., & P. Cordeiro-Estrela. 2016. Taxonomic revision of the Dasypus kappleri complex, with revalidations of Dasypus pastasae (Thomas, 1901) and Dasypus beniensis Lönnberg, 1942 (Cingulata, Dasypodidae). Zootaxa 4170:271-297. https://doi.org/10.11646/zootaxa.4170.2.3
Feijó, A., B. Patterson, & P. Cordeiro-Estrela. 2018. Taxonomic revision of the long-nosed armadillos, Genus Dasypus Linnaeus, 1758 (Mammalia, Cingulata). PLOS ONE:13:e0195084. https://doi.org/10.1371/journal.pone.0195084
Feijó, A. et al. 2019. Phylogeny and molecular species delimitation of long-nosed armadillos (Dasypus: Cingulata) supports morphology-based taxonomy. Zoological Journal of the Linnean Society 186:813-825. https://doi.org/10.1093/zoolinnean/zly091
Fernández, F. 2006. Familia Formicidae. Introducción a los Hymenoptera de La Región Neotropical (J. L. Nieves-Aldrey, F. Fontal-Cazallaa & F. Fernández, eds.). Sociedad Colombiana de Entomologia y Universidad Nacional de Colombia, Bogotá D.C., Colombia. https://doi.org/10.25100/cm.v48i4.3662
Fleck, D. W., & R. S. Voss. 2016. Indigenous knowledge about the greater long-nosed armadillo, Dasypus kappleri (Xenarthra: Dasypodidae), in northeastern Peru. Edentata 17:1-7. https://doi.org/10.2305/iucn.ch.2016.edentata-17-1.2.en
Hoffman, R. L. 2005. A new genus of Spirostreptidae from northern Andes. Myriapodologica 8:83-91.
Humphreys, R. K., & G. D. Ruxton. 2018. A review of thanatosis (death feigning) as an anti-predator behaviour. Behavioral ecology and sociobiology 72:1-16. https://doi.org/10.1007/s00265-017-2436-8
Instituto Geográfico Agustín Codazzi 2021. Región Orinoquia Temperatura. http://www.siac.gov.co/iw/web/tiempo-y-clima/region-orinoquia-temperatura
Johansson, Ö. et al. 2020. Identification errors in camera-trap studies result in systematic population overestimation. Scientific Reports 10:1-10. https://doi.org/10.1038/s41598-020-63367-z
Johnson, N. F., & C. A Triplehorn (eds.). 2005. Borror and DeLong’s Introduction to the Study of Insects. Thompson Brooks/Cole, Belmont.
Kennedy, M. J. 1979 (ed.). Basic methods of specimen preparation in parasitology. Manuscript reports/IDRC, Ottawa.
Lavelle, P. et al. 2014. Soil ecosystem services and land use in the rapidly changing Orinoco River Basin of Colombia. Agriculture, Ecosystems & Environment 185:106-117. https://doi.org/10.1016/j.agee.2013.12.020
Linnaeus, C. (ed.). 1758. Systema naturæ per regna tria naturæ, secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymis, locis. Tomus I. Editio decima, reformata. Holmiæ. (Salvius), Vindobonae [Vienna]. https://doi.org/10.5962/bhl.title.156783
Lira-Torres, I., & M. Briones-Salas. 2012. Abundancia relativa y patrones de actividad de los mamíferos de los Chimalapas, Oaxaca, México. Acta Zoológica Mexicana 28:566-585. https://doi.org/10.21829/azm.2012.283859
Loughry, W. J., & C. M. Mcdonough (eds.). 2013. The nine-banded armadillo: a natural history. University of Oklahoma Press, Oklahoma. https://doi.org/10.1644/13-mamm-r-228
Mondolfi, E. 1968. Descripción de un nuevo armadillo del género Dasypus de Venezuela (Mammalia-Edentata). Memoria de la Fundación La Salle de Ciencias Naturales 27:149-67.
Mosquera-Guerra, F., F. Trujillo, A. P. Díaz-Pulido, & H. Mantilla-Meluk. 2018. Diversidad, abundancia relativa y patrones de actividad de los mamíferos medianos y grandes, asociados a los bosques riparius del río Bita, Vichada, Colombia. Biota Colombiana 19:202-218. https://doi.org/10.21068/c2018v19n01a13
Pardo-Vargas, L. E., & E. Payán-Garrido. 2015. Mamíferos de un agropaisaje de palma de aceite en las sabanas inundables de Orocué, Casanare, Colombia. Biota Colombiana 16:54-66. https://doi.org/10.21068/c2018.v19n02a10
Pardo-Vargas, L. E., M. J. Campbell, M. V. Cove, W. Edwards, G. Reuben, & W. F. Laurance. 2019. Land management strategies can increase oil palm plantation use by some terrestrial mammals in Colombia. Scientific Reports 9:1-12. https://doi.org/10.1038/s41598-019-44288-y
Ramos, D. G. S., et al. 2016. Endoparasites of wild animals from three biomes in the State of Mato Grosso, Brazil. Arquivo Brasileiro de Medicina Veterinária e Zootecnia 68:571-578. https://doi.org/10.1590/1678-4162-8157
Rocha, E. C., E. Silva, J. C. Dalponte, & G. M. Giúdice. 2012. Effect of ecotourism activities on richness and abundance of species of medium and large mammals in the cristalino region, Matto Grosso, Brazil. Revista Árvore 36:1061-1072. https://doi.org/10.1590/s0100-67622012000600007
Rodrigues, T. F., A. M. Mantellatto, M. Superina, & A. G. Chiarello. 2020. Ecosystem services provided by armadillos. Biological Reviews 95:1-21. https://doi.org/10.1111/brv.12551
Rodríguez-Bolaños, A., L. Carvajal-R, & W. Ariza-C (eds.). 2014. Fauna del Bosque de Los Guayupes, Cuenca del Río Guayuriba, Acacías-Meta. Cormacarena – Universidad Distrital Francisco José de Caldas, Bogotá D.C. https://doi.org/10.33304/revinv.v09n1-2017009
Rodríguez-Bolaños, A., L. Carvajal-R, & W. Ariza-C (eds.). 2015. Fauna de los bosques de las cuencas de los ríos Planas y Tillavá, Puerto Gaitán, Meta, Colombia. Cormacarena – Universidad Distrital Francisco José de Caldas, Bogotá D.C. https://doi.org/10.48082/espacios-a20v41n42p18
Rodríguez-Durán, A., K. Valencia, M. Superina, & R. Peæa. 2018. Distribución y usos de los armadillos en sabanas inundables de Arauca, Colombia. Biota Colombiana 19:117-127. https://doi.org/10.21068/c2018.v19n02a10
Rodríguez-Mahecha, J. V., M. Alberico, F. Trujillo, & J. Jorgenson (eds.). 2006. Libro rojo de los mamíferos de Colombia. Conservación Internacional Colombia y Ministerio de Medio Ambiente, Vivienda y Desarrollo Territorial, Bogotá D.C.
Rowcliffe, M. 2021. Activity: Animal Activity Statistics. R package version 1.3.1. https://CRAN.R-project.org/package=activity
Serrato, P (ed.). 2018. La Altillanura colombiana: Aspectos biofísicos. Instituto Geográfico Agustín Codazzi, Bogotá D.C.
Superina, M., F. Trujillo, F. Mosquera, R. Combariza, & C. A. Parra (eds.). 2014. Plan de acción para la conservación de los armadillos de los Llanos Orientales, Cormacarena, Corporinoquia, ODL y Fundación Omacha. Bogotá D.C.
Superina, M., A. C. Duarte, & F. Trujillo. 2019. Connecting research, management, education and policy for the conservation of armadillos in the Orinoco Llanos of Colombia. Oryx 53:17-26. https://doi.org/10.1017/s0030605318000790
Szeplaki, O. E., J. G. Ochoa, & J. A. Clavijo. 1988. Stomach contents of the greater long-nosed armadillo (Dasypus kappleri) in Venezuela. Mammalia 52:422-425. https://doi.org/10.1515/mamm-1988-0314
Thomas, O. 1901. New species of Saccopteryx, Sciurus Rhipidomys, and Tatu from South America. The Annals and magazine of natural history, zoology, botany, and geology 7:366-71. https://doi.org/10.1080/00222930108678484
Tompkins, D. M. et al. 2002. Parasites and host population dynamics. The ecology of wildlife diseases (P. J. Hudson, A. Rizzoli, B. T. Grenfell, H. Heesterbeek & A. P. Dobson, eds.). Oxford University Press, Oxford.
Trujillo, F., & M. Superina. 2013. Armadillos de los Llanos Orientales. ODL, Fundación Omacha, Cormacarena, Corporinoquia, and Bioparque Los Ocarros, Bogotá D.C.
Wetzel, R. M., & E. Mondolfi. 1979. The subgenera and species of long-nosed armadillos, genus Dasypus L. Vertebrate ecology in the northern neotropics, 43-63.
Información adicional
Editor asociado:: Dr. Agustín Manuel Abba
Cite as: Aya-Cuero, C. A., F. Mosquera-Guerra, C. Castillo, D. A. Esquivel, C. Rojano & A. Feijó. 2021. Ecological aspects of Dasypus pastasae in the colombian altillanura, with comments on activity patterns and its identification via camera traps. Mastozoología Neotropical, 28(2):e0594. https://doi.org/10.31687/saremMN.21.28.2.0.07.e0594