ARTÍCULO
Recepción: 18 Julio 2021
Aprobación: 31 Mayo 2022
DOI: https://doi.org/10.4067/s0718-221x2022000100443
Resumen: El objetivo de este trabajo fue evaluar la variación radial de la morfología de las traqueidas de pinos híbridos. Se extrajeron tarugos radiales de árboles de pino híbrido “F1 INTA-PINDO”, en dos ensayos. Las muestras se tomaron a 1,30 m de altura, se disociaron y se midieron las características anatómicas de las traqueidas individuales obtenidas de la fracción de madera tardía de los anillos de crecimiento 4, 7, 10 y 11 mediante microscopía óptica con análisis de imagen. Para la obtención del ángulo microfibrilar se utilizó la técnica a través de las puntuaciones en traqueidas individuales. Para el ángulo microfibrilar dentro de cada edad, se encontraron diferencias estadísticas significativas entre sitios y familias para un nivel de confianza del 95 %, mientras que las variaciones no fueron significativas para las demás propiedades estudiadas. Se observaron cambios importantes en la morfología celular de las traqueidas para las edades consideradas. Los valores medios de ángulo microfibrilar, ancho del lumen y ancho total de las traqueidas disminuyeron con la edad, independientemente del sitio y las familias evaluadas. Los valores medios de espesor y la proporción de pared celular aumentaron con la edad. Estos resultados indican que las propiedades de la madera no serían uniformes en el sentido radial y deben ser tenidas en cuenta al momento de utilizarlas para determinados usos. Se identificaron familias con mejor performance en crecimiento y parámetros deseables para la calidad estructural de la madera, las mismas pertenecen a las mejores posiciones del ranking genético de crecimiento. Se concluye que, a los 11 años de edad este taxón produce madera madura, debido a las características de su morfología celular.
Palabras clave: Anatomía de la madera, ángulo microfibrilar, espesor de pared celular, madera tardía, variación radial.
Abstract: This work aimed to evaluate radial variation of tracheid morphology for hybrid pines. Radial increments cores were extracted from trees of hybrid pine “F1 INTA-PINDO” in two trials. The samples were taken at 1,30 m in height, they were dissociated, and the anatomical characteristics of individual tracheids obtained from the latewood fraction of growth rings 4, 7, 10, and 11 were measured by optical microscopy with image analysis. To obtain the microfibrillar angle, the technique "through individual tracheid pit apertures" was used. For microfibrillar angle within each age, significant statistical differences were found between sites and families for a confidence level of 95%, whereas the variations were not significant for others studied properties. Important changes with considered ages were observed in the cell morphology of the tracheids. The mean values of microfibrillar angle, lumen width, and the total width of tracheids decreased with age, regardless of sites and evaluated families. The mean values of cell wall-thickness and cell-wall ratio increased with age. These results indicate that wood properties would not be uniform in the radial direction and they must be taken into account for certain uses. Families with better growth performance and desirable parameters for structural wood quality were identified. The outstanding families belong to the best positions of growth genetic ranking. It is concluded that at 11 years old, this taxon produces mature wood, due to the characteristics of its cell morphology.
Keywords: Cell wall thickness, hybrid pine, latewood microfibrillar angle, tracheid width, tracheid lumen width, wood anatomy.
Introducción
De acuerdo a la Dirección Nacional de Desarrollo Foresto Industrial (DNDFI), Argentina cuenta con 1,37 millones de hectáreas de plantaciones forestales, de las cuales un 78% se encuentran en las provincias de las regiones Mesopotámica y Delta del Paraná. Dentro de las principales especies forestales cultivadas se encuentran los géneros Pinus (62%); Eucalyptus (26%); Salix (10%) y el resto agrupa a otras especies (Ministerio de Agricultura, Ganadería y Pesca 2019). Las coníferas de importancia económica en la región, han sido Pinus taeda y Pinus elliottii, demostrando buen crecimiento y adaptación. Además, se ha incorporado la F2 del híbrido interespecífico entre P. elliottii var. Elliottii y P. caribaea Morelet var. Hondurensis (PEE × PCH), más recientemente se han establecido plantaciones con material F1 de este híbrido generados en Argentina. La F2 australiana ha sido introducida en la región mesopotámica desde la década del ´90, demostrando excelente crecimiento y extraordinarias cualidades de rectitud de fuste, diámetro de ramas y estructura de copa (Cappa et al. 2013). En Argentina, las propiedades de la madera en la F2 australiana no han sido estudiada exhaustivamente, pero existen antecedentes donde se abordaron la calidad de la madera (Pereyra et al. 2007 y Pereyra et al. 2008).
En los últimos años se incrementó la demanda de material de propagación del híbrido PEE × PCH para la generación de nuevas plantaciones en la Mesopotamia Argentina (Gauchat et al. 2005). En consecuencia, en el año 2004 el Instituto Nacional de Tecnología Agropecuaria (INTA) inició junto a la empresa PINDO S.A. un programa de hibridaciones para la obtención de material F1 del híbrido PEE × PCH (F1 INTA-PINDO). En el marco de dicho programa, se estimaron parámetros genéticos para el crecimiento, la rectitud del fuste y calidad de ramas (Belaber et al. 2018). Y, por otro lado, se estudiaron algunas propiedades mecánicas de la madera de dicho material genético (Acevedo et al. 2016).
Dada la importancia de contar con un leño lo más homogéneo posible en el proceso de industrialización de la madera, numerosos estudios relacionados a la aptitud dela misma se concentran en la variación radial de sus propiedades (Zobel y Van Buijtenen 1989). Entre las diferentes características estudiadas, el ángulo microfibrilar (AMF) es una de las propiedades anatómicas más influyentes en el comportamiento físico-mecánico de la madera (Donaldson 1996, Long et al. 2000, Deresse et al. 2003, Yin et al. 2011, Hein et al. 2013, Winck et al. 2013). Algunos estudios demostraron que el AMF es más sensible al clima que el ancho de los anillos de crecimiento y la densidad de la madera (Xu et al. 2012, Xu et al. 2013). Más recientemente, Xu et al. (2015) destacan que la variación anual en el AMF y el diámetro de las traqueidas están fuertemente ligados a factores climáticos, como temperaturas y precipitaciones. Estos autores, indicaron que los árboles podrían cambiar sus características internas para adaptarse al clima cambiante. Otros aspectos que contribuyen a la variación del AMF están relacionados al sitio (Larson et al. 2001, Gorman et al. 2018), a la densidad de plantación (Winck et al. 2013), al manejo silvícola (Winck et al. 2015) y a la edad del árbol (Donaldson 1996, Long et al. 2000, Harding et al. 2004, Wu et al. 2009). Estas variaciones del AMF, pueden tener una influencia trascendental en las características físicas-mecánicas de la madera limitando su uso estructural. La madera de coníferas producida en su etapa juvenil, apreciada en la construcción por su relación resistencia densidad, se caracteriza por presentar un cambio progresivo en la forma y dimensiones de las células del leño y también en sus propiedades (Panshin y De Zeeuw 1980). El AMF es una de las características más utilizada para la determinación del límite entre la madera juvenil y madura (Bhat et al. 2001, Kojima y Yamamoto 2004). Además, la variación del AMF influye en otras propiedades de la madera. Y, si bien existen algunos antecedentes referido al AMF para el pino híbrido (Wu et al. 2009, Bailleres et al. 2019), no se han publicado estudios para la F1 INTA-PINDO relacionados a esta temática. Consecuentemente, dada la relevancia del AMF, este trabajo propone conocer el grado de variación radial de las propiedades anatómicas de 15 familias híbridas “F1 INTA-PINDO”, obtenidas de 3 posiciones (superior, media e inferior) del ranking genético de crecimiento determinado al séptimo año, como así también el límite entre madera juvenil y madura.
Materiales y métodos
Caracterización de los ensayos y selección de los árboles
El material genético utilizado, pino híbrido PEE × PCH, en adelante “F1 INTA-PINDO”, proviene de 2 ensayos. El primero, implantado en el campo del INTA (26° 33' latitud S y 54° 40' longitud O, sitio 1) y el segundo establecido en propiedad de la empresa PINDO SA (26° 09' latitud S y 54° 26' longitud O, sitio 2). El diseño de plantación correspondió al de bloques completos, con tratamientos distribuidos al azar en parcelas lineales conformadas por 5 hermanos completos. Mayores detalles respecto al material genético y características de estos ensayos pueden verse en Belaber et al. (2018). El clima de la región de estudio se caracteriza por presentar una precipitación media anual de 1864,5 mm, con temperaturas medias en los meses más cálidos y fríos de 26,4 ºC y 14,9 ºC, en enero y julio, respectivamente, con valores extremos de temperaturas de 40,6 ºC y -5,6 ºC (Silva et al. 2019).
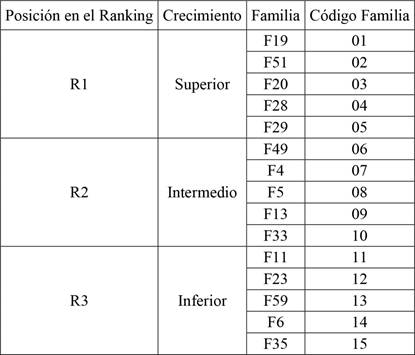
A los 11 años de edad se midió el diámetro a la altura de pecho (DAP) y altura total de todos los árboles de ambos ensayos. En base al ranking genético de crecimiento del 7mo año, 15 de las 66 familias fueron seleccionadas para este estudio, 5 de la posición superior del ranking (R1), 5 intermedias (R2) y 5 inferior (R3). Luego se tomó una muestra constituida por 150 árboles (Pérez-López 2005), 75 individuos de cada ensayo, abarcando la variabilidad en DAP. De cada árbol muestreado se extrajo un tarugo radial de 5 mm de diámetro tomado a la altura del DAP (Tabla 1).
Ranking genético de crecimiento para las 15 familias de ambos sitios.
Preparación de muestras y medición de caracteres anatómicos
Los tarugos radiales fueron secados en estufa a 70 °C por un período de 72 horas hasta alcanzar un contenido de humedad del 12 %. Posteriormente, se pulieron los tarugos, y se tomaron muestras del leño tardío de los anillos de crecimiento 4, 7, 10 y 11 se disociaron utilizando una solución de ácido acético y agua oxigenada en partes iguales (Franklin 1937), y luego se tiñeron con safranina al 1%. Los disociados fueron observados empleando un microscopio óptico trinocular marca Nikon modelo Eclipse LV100DA-U con iluminación por luz transmitida. El sistema cuenta con una cámara digital color modelo DS-FI2-U3 cuyo software de operación para procesamiento de imágenes es NIS-Elements D versión 4.13.00 64-bit. Para determinar el AMF se empleó la técnica denominada “a través de la apertura de las puntuaciones presentes en traqueidas” (Huang et al. 1997). El ángulo microfibrilar se midió sobre traqueidas individuales seleccionadas al azar y obtenidas del leño tardío, trazando una línea paralela al eje de la misma que pase por el centro de la puntuación (Figura 1). El ángulo formado (θ) entre la línea que pasa por el centro de la puntuación y la orientación axial (AO) de las traqueidas indica la orientación de las microfibrillas (AMF) en la capa S2 de la pared celular secundaria.
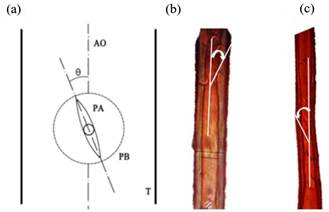
Figura 1:
(a) Diagrama del método utilizado para medir el ángulo microfibrilar de la capa S2 de la pared celular de las traqueidas, PA=apertura de la puntuación, PB=borde de la puntuación, T=traqueida (tomado de Yin et al. 2011), (b) y (c) fotos de traqueidas con indicación de AMF medido.
Sobre el material disociado, se realizaron 15000 mediciones para cada una de las tres características anatómicas evaluadas en las traqueidas (es decir: ancho total, ancho de lumen y ángulo microfibrilar) constituidas por 25 repeticiones evaluadas en 4 anillos de 5 árboles pertenecientes a 15 familias muestreadas en 2 sitios, siguiendo las recomendaciones de las normas IAWA (2004). El valor promedio para el ancho total, ancho de lumen y AMF para cada anillo se determinó a partir de mediciones sobre 25 traqueidas individuales. Mientras que, el espesor de pared celular se obtuvo mediante la diferencia entre el ancho total y el ancho del lumen dividido por dos.
Los datos se analizaron con modelos lineales mixtos. Se incluyó al árbol, como fuente de variación de efecto aleatorio, y para cada sitio y edad se tomó la familia como fuente de efecto fijo. Los análisis estadísticos se realizaron utilizando el software Infostat (Di Rienzo et al. 2020), fijando un nivel de confianza del 95%. Se utilizó la técnica de análisis multivariado denominado “Conglomerados o Análisis de Cluster” para agrupar familias por características anatómicas similares y para diferenciar el límite entre madera juvenil/madura.
Resultados y discusión
Los valores de diámetros a la altura de pecho de los árboles muestreados estuvieron entre 11,5 cm y 40,1 cm. Mientras que la altura total tomó valores entre 12,5 m y 26,8 m. En la Tabla 2 se presenta las medidas resúmenes para las características morfológicas de las traqueidas para cada sitio. El sitio 2 presentó menores valores medios de AMF, ancho y lumen de traqueidas, y mayor valor medio de espesor y proporción de pared celular; con mayor homogeneidad, representadas mediante un menor coeficiente de variación para las distintas variables.
Medidas resúmenes de las características morfológicas de las traqueidas.
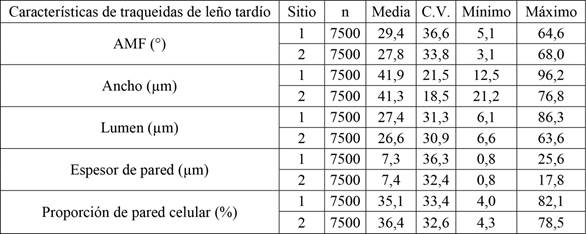
AMF= ángulo microfibrilar; n=número de mediciones; C.V.=coeficiente de variación.
Variación de ángulo microfibrilar entre familias para cada sitio y edad
A los 4 años de edad (anillo 4), se detectaron diferencias estadísticamente significativas entre familias para el AMF en los sitios estudiados (valor-p≤0,022). Los valores medios de AMF registrados para el sitio 2 fueron menores en un 7,1% respecto al sitio 1. Los mayores valores de AMF encontrados para el sitio 1 podrían ser atribuidos a una alta velocidad de crecimiento inicial propiciado por una disponibilidad privilegiada de recursos minerales asociados al uso anterior de la tierra. Para el sitio 1 las familias con menores valores de AMF fueron la 02, 03, 04, 05, 06 y 07, arrojando un valor medio de 36,5° (Figura 2a). Por otra parte, para el sitio 2, presentaron menores valores de AMF las familias 01, 02, 03, 04, 05, 07, con una media de 32,9° (Figura 2b). Estas familias pertenecen a la posición 1 y 2 del ranking genético de crecimiento, indicando que, al seleccionar por crecimiento, indirectamente se estaría eligiendo los ejemplares con mejor desempeño en cuanto al AMF.
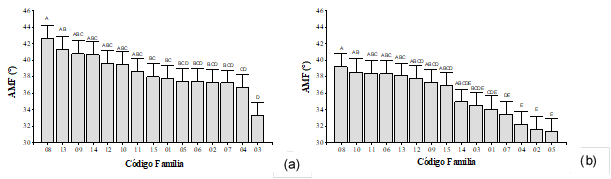
Figura 2:
AMF a los 4 años para (a) el sitio 1 y (b) el sitio 2.
A los 7 años de edad (anillo 7), no se encontraron diferencias estadísticamente significativas para el AMF entre las familias del sitio 1 (valor-p=0,111), mientras que sí fueron detectadas para el sitio 2 (valor-p= 0,012). Los valores medios de AMF para el sitio 1 y 2 fueron de 32,8° y 29,5°, respectivamente. Para el sitio 2 los menores valores de AMF ocurrieron para las familias 01, 03, 04, 05, 07, 08 y 14, alcanzando en estas un promedio de 28,1°. Se identificaron 4 familias (03, 04, 05 y 07), con menores valores medios de AMF a los 4 y 7 años de edad. A los 10 años de edad (anillo 10), no se detectaron diferencias estadísticamente significativas para el AMF entre familias dentro del sitio 1 y 2, (valores-p ≥0,179). Sin embargo, se mantuvo la tendencia de un menor valor de AMF para el sitio 2 (25,5°) respecto del sitio 1 (27,1°). A los 11 años de edad (anillo 11) se evidenciaron diferencias estadísticamente significativas para el AMF entre familias en ambos sitios (valores-p ≤ 0,046). Los valores medios para el sitio 1 y 2 fueron de 18,9° y 20,4°, respectivamente. Los menores valores medios de AMF, en el sitio 1 incluyó a las familias 01, 02, 03, 04, 05, 07 y 09 (Figura 3a). Mientras que para el sitio 2, estuvieron las familias 02, 03, 04, 07, 08, 09 y 14 (Figura 3b). Se identificaron 5 familias con menores valores AMF que son comunes a ambos sitios: 02, 03, 04, 07 y 09 (Figura 3a y Figura 3b).
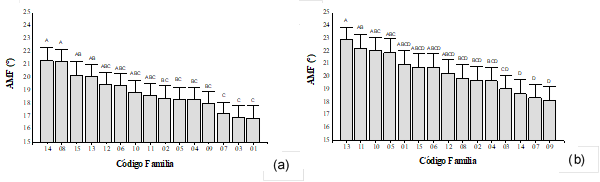
Figura 3:
AMF a los 11 años para (a) el sitio 1 y (b) el sitio 2.
En resumen, los valores promedios de AMF del sitio 2 fueron menores a los del sitio 1, la excepción se dio a la edad de 11 años (Figura 3a y Figura 3b). Esto puede deberse al uso anterior de la tierra previo a la instalación del ensayo, el cual correspondió a monte nativo para el sitio 1 y plantación de pino para el sitio 2. Las mejores condiciones físicas y de fertilidad del sitio 1, por provenir de monte nativo, pudieron haber contribuido al desarrollo inicial en los primeros años de crecimiento, resultando en un mayor incremento radial de los árboles y repercutiendo en esta característica anatómica que está muy ligada a la velocidad de crecimiento y por lo tanto al tamaño del individuo. En el caso del sitio 2, el contenido de nutrientes del suelo pudo verse disminuido debido al uso anterior de la tierra, la exportación de nutrientes producto de la cosecha y preparación del terreno para el establecimiento de la nueva plantación (Martiarena et al. 2007). Siendo, el contenido de fósforo el más afectado por estas actividades, limitante del crecimiento por su baja disponibilidad en el suelo de la provincia Misiones (Goya et al. 2003).
Variación de ángulo microfibrilar del material híbrido para cada sitio y edad
Para esta sección del análisis se consideró a todas las familias como un único material genético. Para las edades evaluadas, los valores medios de AMF variaron aproximadamente en 20° y 15° para el sitio 1 y 2, respectivamente. Entre los 4 y 11 años de edad los valores de AMF variaron entre 38,5° y 18,9° para el sitio 1 y entre 35,8° y 20,4° para el sitio 2, siendo estas diferencias significativas (valores-p ≤ 0,0001). En ambos sitios existió una disminución del AMF con la edad, observándose menores variaciones para el sitio 2. Para los períodos comprendidos entre los 4 y 7 años, y 7 a 10 años el descenso anual en el AMF fue de aproximadamente 2 °/año. Mientras que, entre los 10 y 11 años el AMF disminuyó en promedio aproximadamente 7 °/año. Considerando ambos sitios, el AMF presentó una reducción porcentual de 38,2% en su valor promedio.
Valores similares a los encontrados en este estudio fueron reportados por Wu et al. (2009). Estos autores observaron que el AMF disminuyó entre 34° y 17° entre la médula y la corteza en árboles de 11 años de edad de PEE, PCH y en la F1PEE× PCH australiana. El menor valor de AMF medio fue para el híbrido F1PEE × PCH. También observaron una estabilización en el valor del mismo a partir de los 8 años de edad. Asimismo, Harding et al. (2004), reportaron para los 11 mejores clones australianos F1 de pino híbrido PEE × PCH a los 5 años de edad, valores ponderados de AMF obtenidos con Silviscan de 19,8°, 25,4° y 29,1° mínimos, medios y máximos, respectivamente. Por su parte, Long et al. (2000) indicaron que, a partir de los 10 años el valor de AMF se aproxima a los 10°, (esta variación depende de la especie), proporcionando madera más estable y homogénea en sus propiedades mecánicas. Sugiriendo, que a partir de los 11 años se obtendría un leño más estable dimensionalmente y con propiedades estructurales sustancialmente mejoradas. Asimismo, Long et al. (2000), mencionan que en varios trabajos teóricos y experimentales se ha demostrado que el módulo de elasticidad de la madera y la resistencia de las fibras se mantienen constantes para valores bajos de AMF (5°-10°), y que en maderas con AMF de 40°, su resistencia en la dirección longitudinal es reducida a un tercio (1/3) y su rigidez a un quinto (1/5) de su valor, respecto a las maderas que presenten valores entre 0° a 10° de AMF.
Para el crecimiento en diámetro a la altura de pecho y la altura total de los árboles no se detectaron diferencias importantes entre los ensayos 1 y 2. Mientras que fue notable el efecto del sitio sobre el AMF. Contrariamente, Donaldson (1996) no detectó efecto del sitio sobre el AMF en el material examinado de Pinus radiata, pero sí de la edad fisiológica; remarcando que el envejecimiento fisiológico produjo una disminución significativa en los valores de AMF en la madera juvenil. Por su parte, para Pinus taeda, Bendtsen y Senft (1986) determinaron valores de 31,1° en madera juvenil y 18,9° en madera madura. También para Pinus taeda,Groom et al. (2002) establecieron un valor promedio de AMF de 15,4° grados, con valores de 26,8° y 6,1° en los anillos de crecimiento 5 y 48, respectivamente. En este trabajo, se observó que a los 11 años los valores de AMF fueron más uniformes que a edades más tempranas (Figura 4a y Figura 4b), verificado por los menores desvíos respecto al valor medio de cada familia, lo que estaría indicando una estabilización en las características del leño y posiblemente con presencia de madera con mejores rasgos para usos sólidos, con un mayor valor de densidad, rigidez y resistencia.
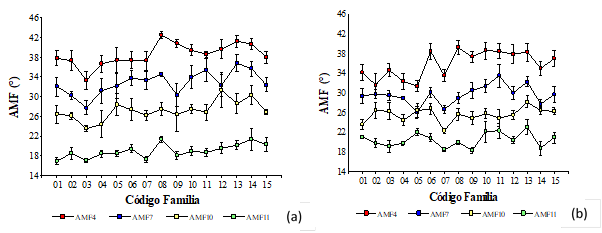
Figura 4:
Variación del ángulo microfibrilar por familias a los 4 (AMF4), 7 (AMF 7), 10 (AMF10) y 11 (AMF11) años de edad para (a) sitio 1 y (b) sitio 2.
Finalmente, a través del análisis de conglomerados se reconocieron 4 familias (01, 03, 04 y 07), que independientemente de la edad evaluada y del sitio en el que se encontraban, arrojaron menores valores medios de AMF. Esta es una de las características deseable de selección, que suele ser utilizada como indicador de la calidad estructural de la madera, a edades tempranas.
Variación del ancho y lumen de traqueidas entre familias para cada sitio y edad
No se evidenciaron diferencias estadísticamente significativas para el ancho de traqueidas entre las familias a las edades 4, 7, 10 y 11 años en los sitios evaluados (valores-p≥ 0,063). El valor medio de ancho de traqueidas del leño tardío fue de 41,6 µm, con mínimos de 30,3 µm y máximos de 65,3 µm. Además, no hubo variaciones significativas entre familias para el lumen de traqueidas para las edades y sitios considerados (valores-p ≥ 0,107). Los valores promedios para el lumen de traqueidas para ambos sitios oscilaron entre 38,8 µm a los 4 años y 19,7 µm a los 11 años de edad.
Considerando a todas las familias como un único material genético, se evidenció una disminución estadísticamente significativa del ancho y del lumen de traqueidas con la edad del árbol (valor-p≤0,000). La reducción fue más marcada entre los 10 y 11 años, representando un 46,8% de la variación total en el ancho, lo que podría estar indicando presencia de madera madura a los 11 años. Los valores expresados en este estudio se encuentran dentro del rango determinado para este taxón (Nuñez 2018) y menores a los determinados por Boschiero (2009) para madera adulta de PCH. Mientras que, para P. taedaWinck et al. (2019) encontraron patrones contradictorios a este, reportaron que el ancho de traqueidas no varió significativamente con la posición social del árbol ni con los tratamientos de poda. Por otro lado, de los 4 a los 11 años, el descenso en el valor promedio de lumen celular fue de 10,3 µm, este comportamiento fue más pronunciado entre los 10 y 11 años de edad, donde la merma fue 36,9% (3,8 µm). Los valores expresados en este estudio se encuentran dentro del rango determinados por Nuñez (2018) para el híbrido “F1 INTA-PINDO” quien determinó un valor promedio de 27,7 µm para el ancho del lumen del leño de verano, siendo muy similar a los alcanzados en este trabajo (25,7 µm). Por otro lado, fueron menores a los determinados por Boschiero (2009) para madera adulta de PCH.
Variación del espesor y proporción de pared celular entre familias para cada sitio y edad
Para la mayoría de las edades, no se evidenciaron diferencias estadísticamente significativas entre familias para el espesor y proporción de pared celular en los sitios para las edades consideradas (valores-p ≥ 0,103), salvo para el espesor de pared en sitio 2 a los 10 años (valor-p = 0,036) y para la proporción de pared celular en sitio 1 a los 7 (valor- p = 0,041) y 10 (valor-p = 0,005) años de edad. De todas maneras, el sitio 2 presentó mayores valores medios de espesor de pared en las edades 4, 7 y 10, excepto a los 11 años.
Considerando a todas las familias de ambos sitios como un único material genético, el espesor y la proporción de pared presentaron diferencias estadísticamente significativas entre edades (valor-p = 0,000), aumentaron con el transcurso del tiempo. Este comportamiento fue más notorio entre los 10 y 11 años para ambas variables, donde el incremento porcentual representó un 31,2 % y 37,0 % de la variación total, para el espesor y proporción de pared celular, respectivamente. Los valores medios de espesor de pared del leño tardío estuvieron entre 5,5 µm a los 4 años y 8,8 µm a los 11 años de edad, con valor mínimo y máximo de 3,4 µm y 12,4 µm, respectivamente. No obstante, estos valores podrían variar, si las mediciones se realizan sobre cortes obtenidos con micrótomo, debido a que en las fibras disociadas pueden sufrir un aplastamiento como consecuencia de la técnica utilizada. Por otra parte, los valores medios de proporción de pared celular permanecieron entre el 26,1% y 45,0 %, a los 4 y 11 años, respectivamente, registrándose valores extremos de 13,7% y 62,1% para el rango de edades evaluadas. La proporción de pared a los 11 años fue similar en ambos sitios. La tendencia encontrada para estas variables es inversa ala del AMF, esto podría indicar la posibilidad de contar con madera de calidad estructural más homogénea, ya que también en el sitio 2 se registraron menores valores medios de AMF y mayores valores medios de espesor y proporción de pared de celular. No se encontraron antecedentes para el espesor y proporción de pared celular para el pino híbrido “F1 INTA PINDO”. Sin embargo, para leño adulto del parental masculino (PCH) Boschiero (2009) reportó valores entre 14,2 µm y 26,5 µm para espesor de pared celular. Por otro lado, para Pinus taeda de 16 años de edad, Winck et al. (2019) reportaron valores similares de espesor de pared celular (8,1 µm).
Transición de madera juvenil a madera madura
La evaluación de las características anatómicas en su conjunto es considerada de importancia, para definir un uso más adecuado de la materia prima. Sin embargo, para especificar parámetros indicadores de la calidad estructural de la madera se aconseja tener en cuenta al AMF y el espesor de pared celular como valor de diagnóstico (Donaldson 1996, Long et al. 2000, Hein et al. 2013), dado que están íntimamente relacionados con su comportamiento físico-mecánico, y por lo tanto, con su procesamiento tecnológico (Winck et al. 2015). A través del análisis de conglomerado se pudieron observar 2 regiones bien demarcadas por los valores de AMF y ancho de traqueidas, indicando la formación de madera madura a partir de los 10 años de edad (Figura 5a y Figura 5b). Sin embargo, cuando se realiza el dendrograma para el lumen, espesor y proporción de pared celular, el límite ocurre a los 7 años de edad.
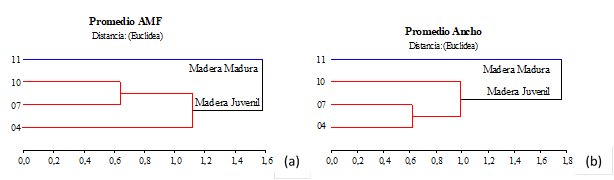
Figura 5:
Dendrograma del límite entre madera juvenil y madura en base a (a) AMF y (b) ancho de traqueidas.
La edad de demarcación entre madera juvenil y madura no es la misma para cada propiedad, sino que varía según la característica que se considere, coincidiendo con Larson et al. (2001), Nawrot et al. (2014) y Wang et al. (2021). Esto indica que el control fisiológico del proceso de maduración de las variables anatómicas está determinado por el desarrollo del fuste y de la copa durante el crecimiento del árbol (Zobel y Sprague 1998). Para las familias de pino híbrido “F1 INTA-PINDO” utilizadas este estudio, la edad de transición varía entre los 7 y 10 años, según la variable considerada. No obstante, este rango de edades para la transición entre madera juvenil y madura es muy similar al registrado para uno de los parentales de este taxón. Por ejemplo, Sackser et al. (2018) en base a la densidad básica de la madera, determinaron que para Pinus elliottii, se encuentra entre los 7 y 9 años de edad, según Zobel y Sprague (1998) entre los 5-8 años, para Clark y Saucier (1989) la producción de madera juvenil se genera entre los 10 y 14 años. Mientras que, para Pinus taeda, se encuentra entre los 7 y 11 años (Clark y Saucier 1989 y Tasissa y Burkhart 1998). Además, la edad de transición varía entre especies y entre árboles de una misma especie. Bailleres et al. (2019) determinaron a través del módulo de elasticidad dinámico, que la edad de transición para el híbrido PEE × PCH ocurre entre los 5 y 10 años de edad. También Meza y Romero (1999) para PCH, indicaron que la madera juvenil se forma hasta los 11 años, coincidiendo con las deducciones del presente estudio para el híbrido “F1 INTA-PINDO”.
En este sentido, se reconoce que la madera juvenil representa las células del xilema formadas por un cambium inmaduro durante los primeros años de vida del árbol. Este tipo de leño exhibe elementos longitudinales más cortos y paredes celulares más delgadas, con una mayor variabilidad. Por lo tanto, las propiedades de la madera juvenil difieren de las del resto del árbol, con cualidades mecánicas inferiores respecto a la madera madura (Winck et al. 2013, Gorman et al. 2018). Por su parte, Zobel (1984) sostiene que el leño juvenil en las especies de coníferas no es de mala calidad, sino simplemente diferente. Por lo tanto, se debe adaptar al destino final para que pueda cumplir con los requisitos relacionados al uso y la satisfacción de las necesidades de los consumidores Zobel (1984). Este autor también hace referencia a que la madera juvenil en algunas ramas de la industria (particularmente en la industria de la pulpa y el papel) alcanza propiedades de rendimiento igual o superior al de la madera madura. Consecuentemente, no puede tratarse como un producto de bajo valor, sino más bien es necesario buscar nuevas alternativas para su procesamiento y utilización más racional. También Nawrot et al. (2014) indica que la presencia de madera juvenil no puede ser eliminada, debido a que es producto del proceso de crecimiento y desarrollo de los árboles, y que solo se puede reducir su proporción dentro de ciertos límites, utilizando medidas silvícolas apropiadas. En concordancia con esto, Larson et al. (2001) y Gorman et al. (2018) encontraron diferencias significativas en el período de transición de la madera juvenil a madura entre sitios, y, atribuyeron períodos de transición más largos en los rodales en el que los árboles exhibieron ramas inferiores persistentes. Por ello indicaron que las condiciones dela masa forestal pueden tener una influencia sustancial en las características físicas y mecánicas del material cuando el mismo es utilizado como madera estructural y/o productos aserrados sólidos. En el material estudiado la poda fue realizada a los 3 años, si se hubieran efectuado una segunda poda, la madera podría haber alcanzado una mayor uniformidad en el sentido radial.
Conclusiones
Los valores medios de ángulo microfibrilar del sitio 2 fueron menores a los del sitio 1, la excepción se dio para los 11 años. Dado que los sitios son cercanos y de características similares, esto puede deberse a un efecto relacionado al uso anterior de la tierra, previo a la instalación del ensayo.
Ocurrió una disminución del ángulo microfibrilar con la edad, independientemente de la familia evaluada y del sitio considerado. Este decrecimiento anual fue más marcado en el último período evaluado, indicando posiblemente la presencia de madera madura. Se recomienda constatar este comportamiento, tomando barrenas de los árboles en pie de ambos ensayos y para evaluar anualmente dichas características en los años consecutivos.
El ancho total de traqueidas, el ancho del lumen, el espesor y la proporción de pared celular no variaron significativamente según sitios y familias consideradas. Sin embargo, para las distintas variables analizadas existió una diferenciación, con un patrón más marcado entre los 10 y 11 años de edad.
Este estudio indica que las familias pertenecientes a la posición 1 del ranking de crecimiento son más prometedoras, tanto por su crecimiento como por sus características en cuanto al valor de ángulo microfibrilar, además porque mantuvieron su superioridad a lo largo del tiempo. Estas fueron las familias 01, 03, 04 y 07. Esto sugiere la factibilidad de realizar la propagación del material genético en base a su crecimiento volumétrico, sin que sean afectadas negativamente las características relacionadas a la calidad estructural. Sin embargo, con los análisis de los perfiles de micro densidad de todos anillos de crecimiento y la caracterización de las propiedades físico-mecánicas de los materiales genéticos y sus relaciones con la morfometría celular se podrán obtener conclusiones concretas.
Las familias pertenecientes a la posición 3 del ranking genético, de menor crecimiento, presentaron un mayor valor de ángulo microfibrilar. Por estas dos razones no se recomienda invertir recursos en su reproducción.
Agradecimientos
Los autores agradecen a la Estación Experimental Agropecuaria de Montecarlo - Instituto Nacional de Tecnología Agropecuaria por el financiamiento brindado para desarrollar la investigación. A la empresa Pindó S.A. quien nos facilitó el sitio donde oportunamente se instaló uno de los ensayos de progenies, además por su colaboración en el mantenimiento y el cuidado del mismo. Y a Elsa Figueredo por la cooperación en las tareas de laboratorio.
Referencias:
Acevedo, P.; Winck, R.A.; Suirezs, M.T.; Gauchat, M.E.; Belaber, E. 2016. Compresión paralela y trac- ción perpendicular de la madera de Pinus elliottii var. elliottii x P. caribaea var. hondurensis. En XXX Jornadas Forestales de Entre Ríos, Concordia, Argentina.5p. https://www.jornadasforestales.com.ar/edic_antes.php
Bailleres, H.; Lee, D.J.; Kumar, C.; Psaltis, S.; Hopewell, G.; Brancheriau, L. 2019. Improving returns from southern pine plantations through innovative resource characterisation. Project number PNC361-1415. Forest & Wood Products Australia: Melbourne, Victoria, Australia. 170p. https://era.daf.qld.gov.au/id/eprint/7939/
Belaber, E.C.; Gauchat, M.E.; Reis, H.D.; Borralho, N.M.; Cappa, E.P. 2018. Genetic Parameters for Growth, Stem Straightness, and Branch Quality for Pinus elliottii var. elliottii × Pinus caribaea var. hondurensis F1 hybrid in Argentina. Forest Science 64(6): 595-608. https://doi.org/10.1093/forsci/fxy021
Bendtsen, B.A.; Senft, J. 1986. Mechanical and anatomical properties in individual growth rings of plantation grown eastern cottonwood and loblolly pine. Wood and Fiber Science 18(1): 23-38. https://wfs.swst.org/ index.php/wfs/article/view/990/990
Bhat, K.M.; Priya, P.B.; Rugmini, P. 2001. Characterization of juvenile wood in teak. Wood Science and Technology 34(6) 517 - 532. https://doi.org/10.1007/s002260000067
Boschiero, A.T. 2009. Caraterização da estrutura anatómica do lenho, dos anéis de crescimento e dos canais de resina de árvores de Pinus caribaea var. hondurensis Barr, et Golf. Teses de Mestrado. Universidade de São Paulo. São Paulo, Brazil. https://www.teses.usp.br/teses/disponiveis/11/11150/tde18052009151531/pt-br.php
Cappa, P.; Marcó, M.A.; Nikles, D.G.; Last, I.A. 2013. Perfomance of Pinus elliottii, Pinus caribaea, their F1, F2 and backcross hybrids and Pinus taeda to 10 years in the Mesopotamia región, Argentina. New Forests 44(4): 197-218. https://doi.org/10.1007/s11056-012-9311-2
Clark, A.; Saucier, J.R. 1989. Influence of initial planting density, geographic location, and species on juvenile Wood formation in southern pines. Forest Products Journal 39: 42-48. https://agris.fao.org/agris- search/search.do?recordID=US19900017416
Deresse, T.; Shepard, R.K.; Shaler, S.M. 2003. Microfibril angle variation in red pine (Pinus resinosa Ait.) and its relation to the strength and stiffness of early juvenile wood. Forest Products Journal 53: 34-40. http://kb.forestprod.org/Main/ind/?id=67004
Di Rienzo, J.A.; Casanoves, F.; Balzarini, M.G.; Gonzalez, L.; Tablada, M.; Robledo, C.W. 2020. InfoStat versión 2020. Grupo InfoStat, FCA, Universidad Nacional de Córdoba: Argentina. https://www.infostat. com.ar/index.php?mod=page&id=46
Donaldson, L.A. 1996. Effect of physiological age and site on microfibril angle in Pinus radiate. IAWA Journal 17(4): 421-429. https://doi.org/10.1163/22941932-90000639
Franklin, G.L. 1937. Permanent preparations of macerated wood fibers. Tropical Woods 49: 21-22.
Gauchat, M.E.; Rodríguez, G.H.; Belaber, E.; Bischoff, D. 2005. Híbridos de alta productividad combinando crecimiento y forma: Pinus elliottii var. elliottii×Pinus caribaea var. honduresnsis. Idia XXI 5(8): 162-170.
Gorman, T.M.; Kretschmann, D.E.; Green, D.W.; Wiemann, M.C. 2018. Effect of site characteristics on juvenile wood transition in lodge pole pine in the inland northwest. Wood and Fiber Science 50(2): 180-192. https://wfs.swst.org/index.php/wfs/article/view/2686
Goya, J.; Pérez, C.; Frangi, J.; Fernández, R. 2003. Impacto de la cosecha y destino de los residuos sobre la estabilidad del capital de nutrientes en plantaciones de Pinus taeda L. Ecología Austral 13(2):139-150. http://sedici.unlp.edu.ar/handle/10915/42428
Groom, L.; Shaler, S.; Mott, L. 2002. Mechanical Properties of Individual Southern Pine Fibers. Part III: Global Relationships Between Fiber Properties and Fiber Location Within an Individual Tree. Wood and Fiber Science 34(2): 238-250. https://wfs.swst.org/index.php/wfs/article/view/1884
Harding, K.J.; Copely, T.R.; Toon, P.G.; Dieters, M.J. 2004. Improving wood quality in a hybrid pine clonal forestry program in Queensland, Australia. IUFRO Forest Genetics Meeting Proceedings. pp. 59-371. https://espace.library.uq.edu.au/view/UQ:101610
Hein, P.R.G.; Silva, J.R.M.; Brancheriau, L. 2013. Correlations among microfibril angle, density, modulus of elasticity, modulus of rupture and shrinkage in 6-year-old Eucalyptus urophylla × Eucalyptus grandis. Maderas. Ciencia y Tecnología 15(2): 171-182. http://dx.doi.org/10.4067/S0718-221X2013005000014
Huang, C.L.; Kutscha, N.P.; Leaf, G.J.; Megraw, R.A. 1997. Comparison of microfibril angle measure- ment techniques. In: Butterfield, B.G (ed.). Microfibril angle in wood. IAWA and IUFRO: Christchurch, New Zealand. pp. 177-205
IAWA. 2004. List of microscopic features for softwood identification. IAWA Journal 25(1): 1-70. https://www.iawa-website.org/uploads/soft/Abstracts/01_IAWA-Softwood_List.pdf
Kojima, Y.; Yamamoto, H. 2004. Effect of microfibril angle to the longitudinal tensile creep behavior of wood. Journal of Wood Science 50 (3): 301-306. https://doi.org/10.1007/s10086-003-0565-3
Larson, P.R.; Kretschmann, D.E.; Clark, A. III.; Isebrands, J.G. 2001. Formation and properties of juvenile wood in southern pines. General technical report FPL-GTR-129. USDA Forest Service, Forest Products Laboratory: Madison, WI. 42p. https://www.fpl.fs.fed.us/documnts/fplgtr/fplgtr129.pdf
Long, J.; Conn, A.B.; Batchelor, W.B.; Evans, R. 2000. Comparison of methods to measure fibril angle in wood fibres. Appita Journal 53(3): 206-209. https://users.monash.edu.au/~batchelo/Downloads/Comparison%20or%20methods%20to%20measure%20wood%20fibres.pdf
Martiarena, R.; Von Wallis, A.; Fernández, R.; Knebel, O. 2007. Efecto de prácticas de preparación de terreno sobre el contenido de nutrientes y el crecimiento en plantaciones forestales. En: Avances en Ingeniería agrícola 2005-2007. ISBN 978-987-1253-29-6: 303-310
Meza, D.J.; Romero, A. 1999. Estimaron una edad de transición de 11 años en Pinus caribaea var. hon- durensis Barr et Golf. de una plantación en la Sabana Oaxaca. Tesis profesional. Universidad Nacional de Chapingo. Mexico. 76p.
Ministerio de Agricultura, Ganadería y Pesca. 2019. Plan estratégico forestal y foresto industrial Argentina 2030. Secretaría de Ambiente y Desarrollo Sustentable. Ministerio de Agricultura, Ganaderia y Pesca: Buenos Aires, Argentina. 178p. https://www.argentina.gob.ar/sites/default/files/2021/04/plan_estrategico_foresto_industrial_2030.pdf
Nawrot, M.; Pazdrowski, W.; Walkowiak, R.; Szymański, M.; Kaźmierczak, K. 2014. Analysis of coniferous species to identify and distinguish juvenile and mature wood. Journal of Forest Science 60(4): 143-153. https://www.agriculturejournals.cz/publicFiles/78_2013-JFS.pdf
Nuñez, V.S. 2018. Dinámica de la actividad cambial en pino híbrido y sus especies parentales. Estudio de caso. Tesina de grado. Facultad de Ciencias Forestales.Universidad Nacional de Misiones, Eldorado, Misiones, Argentina. 30p.
Panshin, A.J.; de Zeeuw, C. 1980. Textbook of wood technology: Structure, identification, properties, and uses of the commercial woods of the United States and Canada. 4th ed. ISBN 0070484414. 722p. McGraw-Hill: New York. https://searchworks.stanford.edu/view/801946.
Pereyra, O.; Suirezs, T.M.; Bobadilla, E.A.; Weber, E.M.; Gamarra, N.G. 2007. Estudio de los pesos específicos aparentes, contracciones, hinchamiento, anisotropía y punto de saturación de fibras de la madera del híbrido Pinus elliottiivar. elliottii× Pinus caribaea var. hondurensis. En III Congreso Iberoamericano de Productos Forestales. Buenos Aires, Argentina.
Pereyra, O.; Suirezs, T.M.; Bobadilla, E.A.; Weber, E.M.; Gamarra, N.G. 2008. Propiedades mecáni- cas de la madera del híbrido Pinus elliottii var. elliottii× Pinus caribaea var. hondurensis, Misiones, Argentina. En 13ª Jornadas Técnicas Forestales y Ambientales. El Dorado. Misiones, Argentina.
Pérez-López, C. 2005. Muestreo estadístico: conceptos y problemas resueltos. 1ª ed. ISBN 9788420544113. 392p. Editorial Alhambra: Argentina. https://libinter.com.ar/libro.php?libro_id=5392
Sackser, M.R.; Valdés, P.A.; Grance, L.A.; Bohren, A.V.; Aguilera, M.A.; Andrade, N.Y.; Grance, J.R.F. 2018. Densidad de la madera de Pinus elliottii var. Elliottii Engelm de 21 años de edad, a distintas alturas desde la medula a la corteza; estimación del límite madera juvenil-madura. Revista Forestal Yvyrareta 26: 29-37. https://rid.unam.edu.ar/handle/20.500.12219/2588
Silva, F.; Haberle, T.; Müller, H.; Münz, R.; Visentini, J. 2019. Normales del clima de la región de Montecarlo. Actas XVIII Jornadas Técnicas Forestales y Ambientales. UNaM: Eldorado, Misiones, Argentina. 526-528p. https://inta.gob.ar/sites/default/files/normales_del_clima_de_la_region_de_montecarlo_2019.pdf
Tasissa, G.; Burkhart, H.E. 1998. Juvenile-mature wood demarcation in loblolly pine trees. Wood and Fiber Science 30 (2): 119-127. https://wfs.swst.org/index.php/wfs/article/view/64
Wang, Y.; Zhang, R.; Zhou, Z. 2021. Radial variation of wood anatomical properties determines the de- marcation of juvenile-mature wood in Schima superba. Forests 12(4): e512. https://doi.org/10.3390/f12040512.
Winck, R.A.; Fassola, H.E.; Area, M.C. 2015. Efecto del raleo sobre las propiedades anatómicas de la madera de Pinus taeda. Maderas. Ciencia y Tecnología 17(2): 391-406. http://dx.doi.org/10.4067/S0718-221X2015005000037
Winck, R.A.; Fassola, H.E.; Tomazello, M.F.; Area, M.C. 2013. Case Study: Microfibril angle and its relationship with basic density in Pinus taeda L. wood from silvopastoral systems. Revista O Papel 745: 55- 61. http://hdl.handle.net/20.500.12123/4090
Winck, R.A.; Martínez, M.I.; Romberg, E.I.; Fassola, H.E.; Bohren, A.V.; Aquino, D.R. 2019. Morfometría de traqueidas de Pinus taeda L. provenientes de un ensayo de poda en NE de Argentina. Floresta 49(3): 503-512. http://dx.doi.org/10.5380/rf.v49i3.59546
Wu, H.; McRae, T.; Southerton, S.; Gapare, W.; Baltunis, B.; Li, X.; Dillon, S.; Ivkovic, M.; Powell, M.; Dieters, M.; Harding, K.; Matherson, C.; Ilic, J. 2009. Quantitative and molecular genetics of juvenile wood traits in radiate and slash/Caribbean pines. Forest and Wood Products Australia: Melbourne, Victoria. 309p. https://www.fwpa.com.au/resources/resources/85-quantitative-and-molecular-ge-netics-of-juvenile-wood-traits-in-radiata-and-slash-caribbean-pines.html
Xu, J.M.; Lu, J.X.; Bao, F.C.; Evans, R.; Downes, M.G.; Huang, R.F.; Zhao, Y.K. 2012. Cellulose microfibril angle variation in Picea crassifolia tree rings improves climate signals on the Tibetan plateau. Trees 26:1007-1016. https://doi.org/10.1007/s00468-012-0678-7
Xu, J.M.; Lu, J.X.; Bao, F.C.; Evans, R.; Downes, M.G. 2013. Climate response of cell characteristics in tree rings of Picea crassifolia. Holzforschung 67(2): 217-225. https://doi.org/10.1515/hf-2011-0144
Xu, J.M.; Lu, J.X.; Evans, R.; Downes, G.M. 2015. Climatic signal in cellulose microfibril angle and tracheid radial diameter of Picea crassifolia at different altitudes of the Tibetan plateau, northwest China. Wood Science and Technology 49(6) 1307-1318. https://doi.org/10.1007/s00226-015-0753-5
Yin, Y.; Song, K.; Liu, B.; Jiang, X. 2011. Variation of microfibril angle in plantation trees of Cunninghamia lanceolata determined by pit apertures and x-ray diffraction. IAWA Journal 32(1): 77-87. https://doi.org/10.1163/22941932-90000044
Zobel, B.J. 1984. The changing quality of the world wood supply. Wood Science and Technology 18(1): 1-17. https://doi.org/10.1007/BF00632127.
Zobel, B.J.; Sprague, J.R. 1998. Juvenile wood in forest trees. 1st ed. ISBN 9783642721281. 311p. Springer-Verlag: Berlin, Heidelberg. https://link.springer.com/book/10.1007/978-3-642-72126-7.
Zobel, B.J.; Van Buijtenen, J.P. 1989. Wood variation: its causes and control. ISBN 9783642740718. 378p. Springer-Verlag: Berlin, Germany.
Notas de autor
(Corresponding author: winck.rosa@inta.gob.ar
