Benthic fish community structure in the Orinoco River Delta and Gulf of Paria (Venezuela), fifty years after the construction of a dike across Manamo Channel
Estructura comunitaria de la ictiofauna bentónica del delta del Orinoco y Golfo de Paria (Venezuela), 50 años después de la construcción del dique del caño Manamo
Benthic fish community structure in the Orinoco River Delta and Gulf of Paria (Venezuela), fifty years after the construction of a dike across Manamo Channel
Biota Colombiana, vol. 17, no. 1, pp. 64-89, 2016
Instituto de Investigación de Recursos Biológicos "Alexander von Humboldt"

This work is licensed under Creative Commons Attribution-NonCommercial-NoDerivs 4.0 International.
Received: 21 December 2015
Accepted: 06 June 2016
Abstract: To evaluate the impacts of water flow restriction and regulation caused by the construction of a dike across Manamo Channel, variations in the benthic fish communities were studied in the mouths of three streams or channels (Pedernales, Angostura and Manamo) of the Orinoco River Delta and one stream (Venado) that flows directly into the Gulf of Paria, during one (annual) hydrological cycle. Fishes were sampled with a shrimp trawl net. Of the 78 species collected, five were resident, 12 temporary and 62 occasional. Juvenile phases of all species were collected. Species with greater IVI values were Cathorops sp., Achirus achirus, Stellifer naso and Colomesus psittacus. No significant differences were obtained for the physical and chemical habitat parameters measured: depth, salinity and transparency, nor in the water levels of the streams even in different hydro-phases of the year. Salinity never dropped to zero and no changes in fish species composition were detected. These results lead us to hypothesize that the impacts of the dike constructed in Manamo Channel also extend to other streams of the Orinoco River Delta and even the one studied (Venado) that empties into the Gulf of Paria, where estuarine conditions prevailed during the entire study period of one year, with no observed changed in the hydrological regimen.
Keywords: Community structure, Estuary, Fishes, Manamo Channel, Shrimp trawl.
Resumen: Con el objetivo de evaluar el efecto de la regulación de las aguas del caño Manamo (por la construcción de un dique), sobre la ictiofauna bentónica de algunos caños del delta del río Orinoco y Golfo de Paria, se estudió la variación de la estructura comunitaria de peces bentónicos en la desembocadura de tres caños del delta del río Orinoco (Pedernales, Angostura y Manamo) y un caño del golfo de Paria (Venado) durante un ciclo hidrológico anual. De las 78 especies colectadas, cinco fueron residentes, 12 temporales y 61 ocasionales; todas registraron ejemplares en estado juvenil. Las especies con mayores valores de IVI fueron Cathorops sp., Achirus achirus, Stellifer naso y Colomesus psittacus. No se observaron diferencias significativas entre los parámetros físico-químicos analizados (profundidad, salinidad y transparencia), ni a nivel de caños, ni entre hidrofases. La salinidad nunca registró valores menores de 4 % y no se observaron cambios en la composición de especies de la comunidad. Con los resultados obtenidos se plantea la hipótesis de que la construcción del dique sobre el caño Manamo en los años 60, afectó el comportamiento natural de este sistema y de otros caños del delta e incluso del golfo de Paria (caño Venado), registrando en las desembocaduras de todos los caños características de un estuario continuo (sin cambios en el régimen hidrológico y salinidad) durante todo el año.
Palabras clave: Caño Manamo, Estuario, Estructura comunitaria, Peces, Red camaronera de arrastre.
Introduction
The Orinoco is one of the largest rivers in the world, occupying third place in water discharge (38.000 m3s-1), behind only the Amazon and the Congo (Rodríguez et al. 2007). Its drainage basin encompasses 1,080,000 km2, shared by Colombia and Venezuela. Traditionally the basin has been divided according to physiographic and limnological criteria into the upper, middle and lower sections, with the delta included in the last (Lasso et al. 2004 a). After flowing some 2000 km, at the city of Barrancas, which is about 50 km from its mouth, the Orinoco River divides into two major branches (Novoa y Cervigón 1986). From there, the principal Orinoco flow is carried west to east by the Río Grande and discharges into the Atlantic Ocean at Boca Grande. The remaining flow is transported mainly by Manamo and Macareo channels, two large distributary channels that carry water to the north and northeast respectively. Since 1966, the flow through Manamo Channel diminished considerably due to the construction of a flow control dike that restricted flow to just 200 m3s-1, and so impeded floodwaters from overflowing into surrounding floodplains (Novoa y Cervigón 1986).
As a consequence of the flow regulation, Manamo Channel has a situation that differs from the other channels in the delta; it does not behave as a positive estuary because having lost most of its freshwater flow, its upper reaches receive a greater saltwater penetration and as a result, greater dispersion of marine and brackish water fishes (Flores et al. 2004). The composition of the fish fauna does not have significant seasonal changes, and estuarine species prevail as temporary visitors or permanent residents in these habitats, as has been documented by Novoa and Cervigón (1986), Novoa (2000 a) and Lasso et al. (2004 b).
The Gulf of Paria drainage is situated between the Paria Peninsula and the Orinoco River Delta in the north-eastern part of Venezuela. The Guanipa River is part of this drainage, its headwaters originate in the state of Anzoátegui, its waters then flow some 340 km before emptying into the Gulf of Paria where it forms the northernmost border of the Orinoco Delta and is known as Venado Channel (Flores et al. 2004).
The streams studied were selected because of their accessible location, which permitted the collection of data about the benthic fish fauna throughout the year and to extend the study of Venado Channel, where only a few studies have been done (Lasso et al. 2004a).
The objective of this study was to compare the structure (composition, abundance, diversity and richness) of the benthic fish communities during one annual hydro-cycle at the mouths of Venado Channel (Gulf of Paria drainage) with those of the Angostura, Manamo and Pedernales channels (Orinoco River Basin), affected by the dam on Manamo Channel.
Materials and methods
Study area
From a hydrographic perspective, two major drainages are recognized in the deltaic region of coastal south-eastern Venezuela: the Gulf of Paria and the Orinoco River Delta. The first has an area of approximately 21,000 km2, which is a little more than 2 % of the country (Lasso y Meri 2003). The second has a surface of about 40.200 km2, of which the deltaic alluvial fan of the Orinoco occupies 18.810 km2 (PDVSA 1993).
Hydrographically, the delta of the Orinoco River can be divided into 12 drainages (Ponte et al. 1999), in this study we analyze the drainages of the Pedernales, Manamo and Angostura channels. The Guanipa River – Venado Channel belongs to the Gulf of Paria basin.
Freshwater flow is determined by the alteration of the wet and dry seasons. The greatest water discharges occur from June to September, and minimum flows from December to April, causing seasonal variation in salinity (Novoa 2000a). This is the normal behaviour of the majority of streams or channels in the Orinoco River delta, such as for example Macareo Channel (Figure 1). However, as mentioned above, on Manamo Channel a dike was installed to control water flow that reduces its flow to only 1 % of the total (Macareo and Boca Grande channels discharge 13 % and 86 % of the total, respectively) (Ponte et al. 1999), thus impeding the overflow and flooding of freshwaters onto its floodplain. With no seasonal variation in freshwater discharge (Figure 1), no significant variation is observed in salinity (Olivares y Colonnello 2000).
Figure 1. Annual hydrological cycle of Macareo and Manamo channels. Adapted fromNovoa (2000a).
It is important to keep in mind that this study was done in the estuarine delta and not the fluvial delta, as defined by Lasso and Sánchez-Duarte (2011). The upstream ecological boundary of the estuary would be the limit of the influence of brackish water in the main channels of the channels. The lower limit with the sea would be a narrow band of variable width, that forms below the sand and mud bars found in front of the channel’s mouths (Lasso y Sánchez-Duarte 2011).
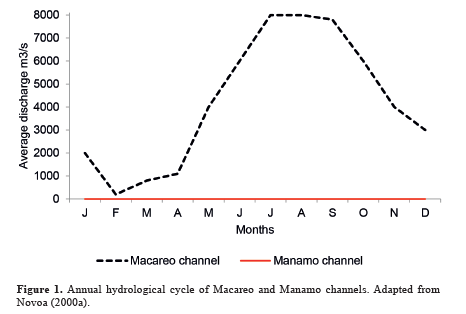
Figure 2 shows the geographic position of the study area, which is bounded by the Guanipa River - Venado Channel to the west, Manamo Channel to the south, Pedernales Channel in the southeast and the Gulf of Paria and Caribbean Sea to the north.
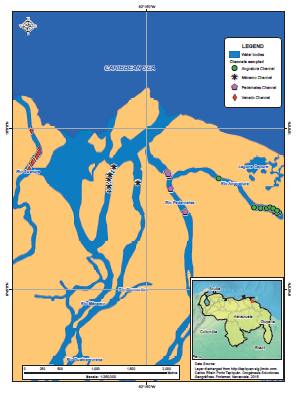
Figure 2.
Study area. Angostura, Manamo, Pedernales channels (Orinoco Basin) and Venado Channel (Gulf of Paria drainage).

Figure 3.
a) Panoramic view of Pedernales Channel, b) fishing with shrimp trawl, c - d) samples collected by shrimp trawl, note puffers (Colomesus psittacus) and seabass (Epinephelus itajara).
Photos: P. Sánchez-Duarte (a), J. Hernández (b – d).Sampling
Samples were collected during each of the four annual hydro-phases (high water, falling water, low water and rising water). Samples were collected with a shrimp trawl, locally known as “chica”, that is used exclusively to fish for shrimp in the mouths of some of the channels of the delta. In our case the net was 11 m long and 8 m wide, with mesh size of 2 cm. It was pulled by a small boat powered by an outboard motor (Novoa 2000 a). The benthic fish fauna of the Orinoco River has been studied by various authors that have used shrimp trawls as experimental sampling gear (Ramos et al. 1982, Cervigón 1982, 1985, Novoa and Cervigón 1986, Novoa 1982, Novoa 2000 a – b, Lasso et al. 2004 b, 2008). Each trawl pull lasted ten minutes at constant velocity and for each operation, initial and final geographic coordinates, date, hour, depth, Secchi transparency, salinity and bottom type were recorded (Figure 3). Sampling effort was the same as used in previous studies of this area (Lasso et al. 2004b, 2008).
Some samples were immediately field processed and released. Others were fixed in formalin and taken to the Museo de Historia Natural La Salle (MHNLS) in Caracas for identification and processing.
Data analysis
Relative abundance (%) and relative biomass (%) were estimated for each trawl sample, based on capture per unit effort. Biomass and fish density were expressed as kg/ha and ind/ha, respectively (Lasso et al. 2004 b). Alpha diversity was calculated using the Shannon-Wienner index (H) (1963), equity index of Pielou (J) and richness index of Margalef (R1) (1969). The following indices were also calculated:
Importance Value Index (IVI) (Lasso et al. 2008). This index permits the evaluation of the importance of each species in the aquatic ecosystem, globally integrating the abundance, biomass and relative frequency of each in just one index using the following formula : IVI = FR + AR + BR. Since the result is the sum of three index percentage values, this IVI index varies from 1 – 300 %.
Community Dominance Index (IDC) (McNaughton 1968). Where IDC = (Y1 + Y2 / Y) • 100 Y1 + Y2= sum of the abundance of the two dominant species Y= total abundance of all species. Values above 40 % abundance are indicative of true community dominance (Goulding et al. 1988).
Keeping in mind that in samples collected using shrimp trawls, a large percentage of the capture corresponds to juveniles of benthic species, the specimens captured were measured, recording the disc width (DW) or standard length (SL) for species of special ecological or commercial interest.
Similarity dendrograms were made using SAS ® and PAST software version 1.34 (Hammer et al. 2001).
Physical and chemical habitat parameters and ecological community parameters were compared among hydro-phases having previously evaluated the homoscedasticity among the samples. Data with homogeneous variance among groups were evaluated using analysis of variance (ANOVA) at significance level p< 0.05 and not inferior to p< 0.01. When significant differences were found, the differing hydro-phases, environmental o ecological parameters causing the differences were identified using the Tukey test (Montgomery 1984). When the homogeneity of variance was less than p< 0.01 data were analyzed using a non-parametric Kruskal-Wallis test (Barletta et al. 2003).
Results
Physical and chemical habitat characteristics
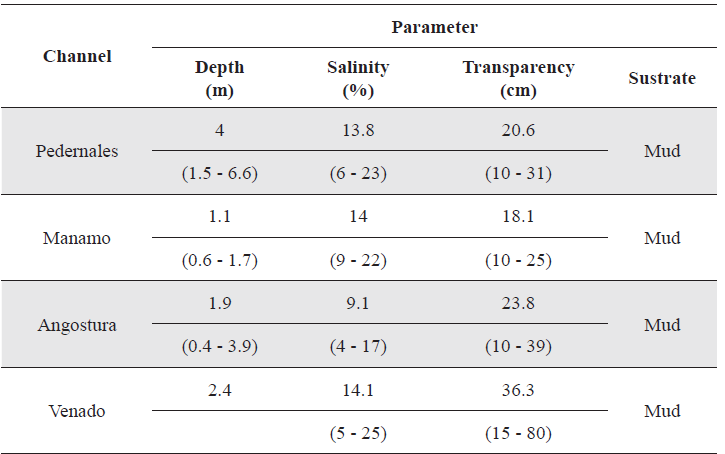
The variations of basic physical and chemical characteristics (depth, salinity and transparency) in each of the four channels studied during the annual hydrological cycle are given in Table 1.
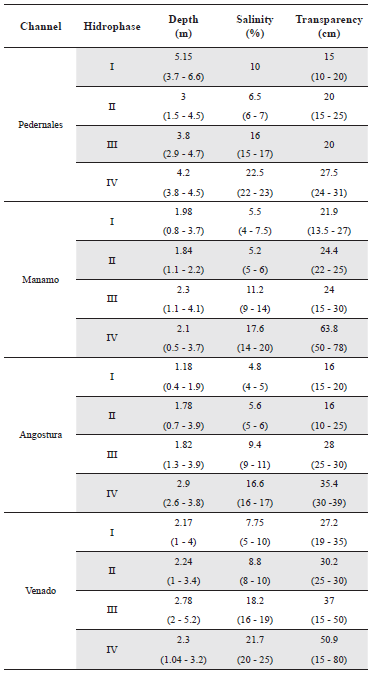
Characterization of the physical and chemical parameters measured in each of the channels studied during the four annual hydro-phases. Average values are given with ranges in parentheses.
In Pedernales Channel, an analysis of variance comparing physical and chemical parameters measured detected no significant differences for any of them during any of the four hydrophases depth (α = 0.05; p = 0.79); salinity (α = 0.05; p = 0.72) and transparency (α = 0.05; p = 0.84).
For Manamo, Angostura and Venado channels (due to the non normality of the data), a Kruskal-Wallis analysis was used to compare the values obtained for the physical and chemical parameters measured during the four hydrophases. In no case were any significant differences detected (Table 2).
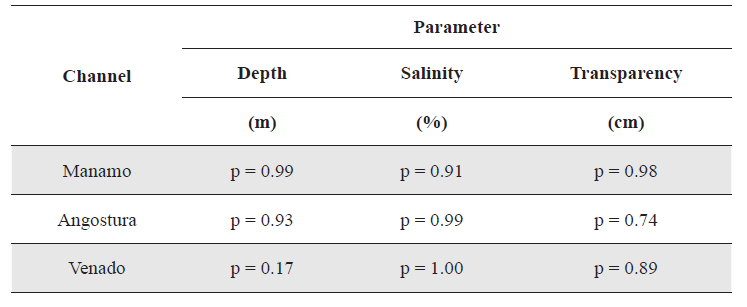
Results of p-value in the Kruskal-Wallis analyses applied to the physical and chemical parameters measured during the four hydrophases, in three of the channels studied α = 0.05.
The variations of basic physical and chemical characteristics (depth, salinity and transparency) of the four channels studied during the annual hydrological cycle are given in Table 3.
Because data were not normally distributed, Kurskal-Wallis analyses were used to compare among hydro-phases the values obtained for physical and chemical parameters recorded. No significant differences were detected for any parameter: depth (α = 0.05; p = 0.95); salinity (α = 0.05; p = 0.72) and transparency (α = 0.05; p = 0.96).
A dendrogram of similarity, drawn based on physical and chemical habitat parameters recorded for the four annual hydro-phases shows greater similarity between Manamo and Angostura channels, and those two grouped together with Pedernales Channel. The most dissimilar channel was Venado (Figure 4).
Characterization of the physical and chemical parameters measured in the channels studied during the four annual hydro-phases. Average values are given with ranges in parentheses.
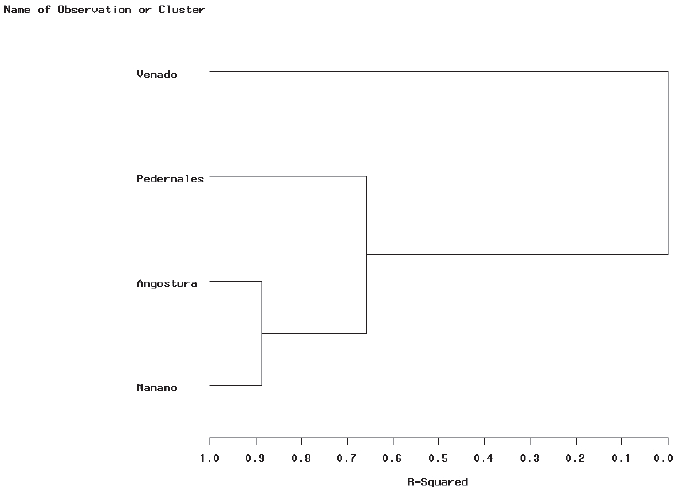
Figure 4.
Dendrogram of similarity amog the four channels based on physical and chemical habitat parameters (depth, salinity and transparency) recorded during the four annual hydro-phases. Cluster analysis performed using minimum variance of Ward (SAS).
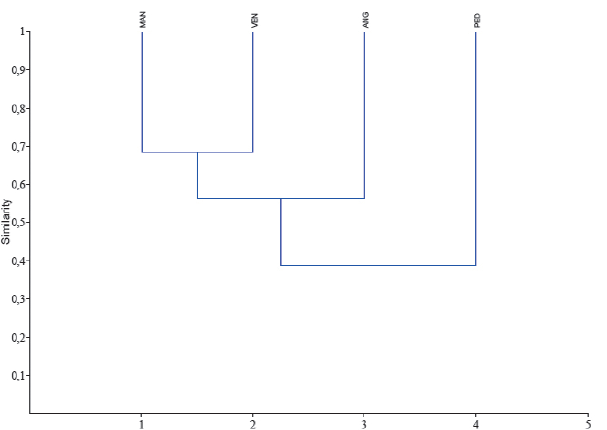
Figure 5.
Similarity dendrogram of species presence - absence, for the channels studied, using the Jaccard Index (PAST version 1.34, Hammer et al. 2001).
Composition and species richness
During the four hydro-phases sampled, 78 benthic fishes were collected (Appendix I). The total corresponds to 29 families in eight orders; of these, Perciformes were the most diverse group with 34 species, followed by Siluriformes (16 species) and Clupeiformes (14 species); the other orders contained one to four species. The four families with the greatest species richness were Sciaenidae (12 species), Engraulidae (10 species), Carangidae (9 species) and Ariidae (7 species); the remaining families had three or fewer species.
Following the proposal of Lasso et al. (2009) species were classified based on salinity preference (Appendix I), and five habitat groups were distinguished. These are: primary freshwater families (F) that are physiologically intolerant of salinity; occasional freshwater (OF) families that are found in freshwater habitats but that can tolerate ample intervals in salty water (euryhaline) or that need estuarine areas for reproduction or feeding; estuarine (E) species that live in the estuary throughout the year; estuarine occasional (EO) species are marine species that enter the estuary for reproduction or feeding; and finally the strictly marine species (M). It was found that the majority of the species were estuarine-marine (41 sp. - 52 % of the total), followed by freshwater occasional (18 sp. – 23 %); in third place the strictly freshwater species (7 sp. – 9 %) followed by the estuarine occasional species (5 sp. – 6 %). The four groups comprised 90 % of all species identified.
Using presence-absence data for each species collected from the four channels during the sampling period and similarity dendrogram was made using the Jaccard Index, that grouped the channels in accordance with the species present (Figure 5).
Based on the abundance of each species collected from the four channels during the sampling period, a similarity dendrogram was made using the Morisita-Horn Index, that grouped the streams according the presence and abundance fo the species during the four annual hydrophases (Figure 6).
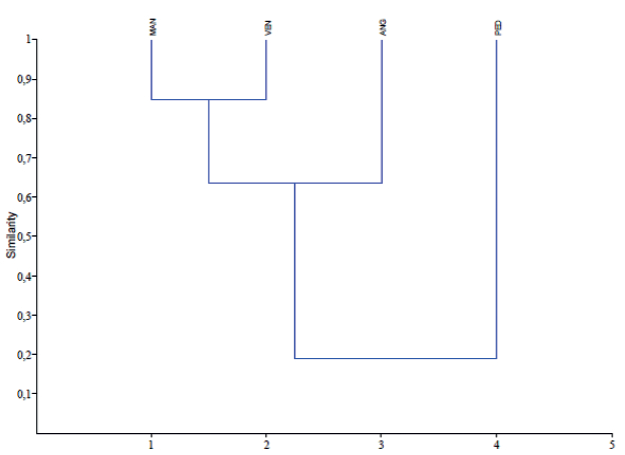
Figure 6.
Similarity dendrogram for the channels according to the presence and bundance of fish species sampled during the study period, using the Morisita-Horn Index (PAST version 1.34, Hammer et al. 2001).
The two analyses gave the same results, indicating that the two most similar channels are Manamo and Venado, with Pedernales being the most dissimilar.
Abundance and total biomass recorded during the annual hydrocycle
During the annual hydrocycle, of the 78 species collected, ten comprised 82 % of the total capture, and the 68 remaining, just 18 % (Figure 7).
Analysis of total biomass collected during the study for the 78 species collected showed that 15 species contributed 7.7 % of the total capture, and the remaining 64 species just 12.3 % (Figure 8).
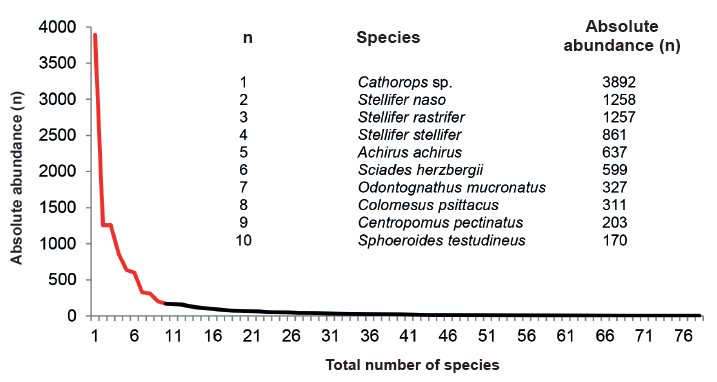
Figure 7.
Distribution of the abundance of the 78 species collected from Pedernales, Manamo, Angostura and Venado channels during the hydrocycle studied.
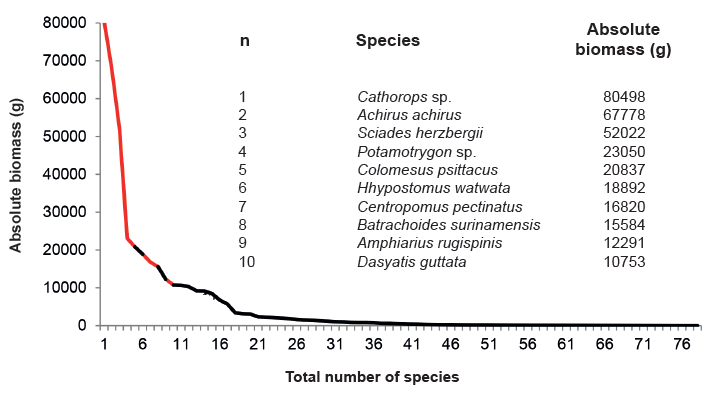
Figure 8.
Distribution of biomass for the 78 fishes collected during the four hydrophases studied in Pedernales, Manamo, Angostura and Venado channels.
Ecological Indices
The variation in the Shannon diversity (H´), equity (J) and richness of Margalef (R1) during the four hydrophases studied, for each of the four channels studied is given below.
In Pedernales Channel the ecological indices had similar behaviour during the entire hydrocycle. An analysis of variance indicates that there are no significant differences in among any of the indices calculated, Shannon diversity (α = 0.05; p = 0.59), richness of Margalef (α = 0.05; p = 0.94) and equity (α = 0.05; p = 0.29) (Figure 9).
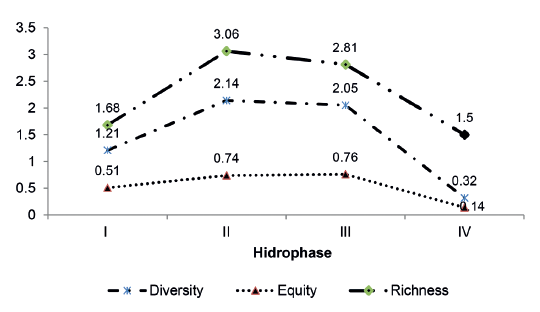
Figure 9.
Variation in ecological indices (non significant) for Shannon diversity (H´), equity (J) and richness of Margalef (R1) in Pedernales Channel, during the annual hydrocycle
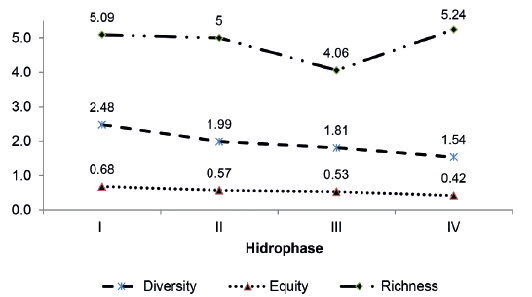
Figure 10.
Variation in the Shannon diversity index (H´), equity (J) and Margalef richness (R1) in Manamo Channel, during the annual hydrocycle studied.
The variation in diversity, equity and richness for the four hydrophases in Manamo Channel are shown in Figure 10. Analysis of variance showed that there were no significant differences for the Shannon diversity index (α = 0.05; p = 0.15), or for the Margalef richness index (α = 0.05; p = 0.76). Because data were not normally distributed, a Kruskal-Wallis analysis was calculated that there were no significant differences in the equity index either (α = 0.05; p = 0.37).
For Angostura Channel the variation in the diversity, equity and richness indices for the hydrophases studied are given in Figure 11. An analysis of variance detected no significant differences for the ecological indices calculated which were the Shannon diversity index (α = 0.05; p = 0.87), equity (α = 0.05; p = 0.87) and richness of Margalef (α = 0.05; p = 0.90).
Variation in the ecological indices calculated for Venado Channel, Shannon diversity (H´), equity (J), and Margalef richness (R1) are given in Figure 12. An analysis of variance detected no significant differences for the Margalef richness index (α = 0.05; p = 0.77). Because data were not normally distributed, a Kruskal-Wallis analysis was used to show that there were no significant differences in the Shannon diversity index (α = 0.05; p = 0.78) or equity (α = 0.05; p = 0.39), during the hydrocycle studied.
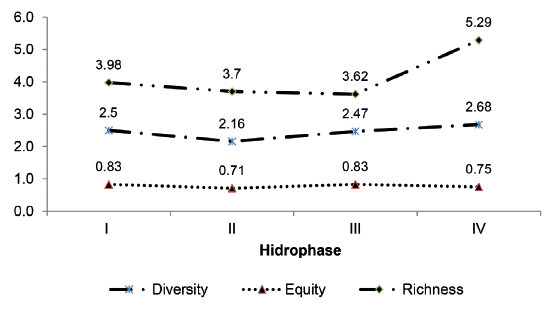
Figure 11.
Variation in the Shannon diversity index (H´), equity (J) and Margalef richness (R1) in Angostura channel, during the hydrological cycle.
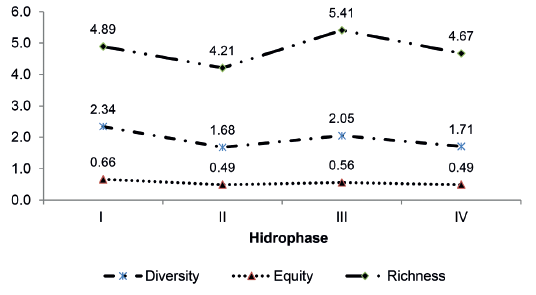
Figure 12.
Variation in the Shannon diversity index (H´), equity (J) and Margalef richness (R1) in Venado channel during the hydrocycle studied.
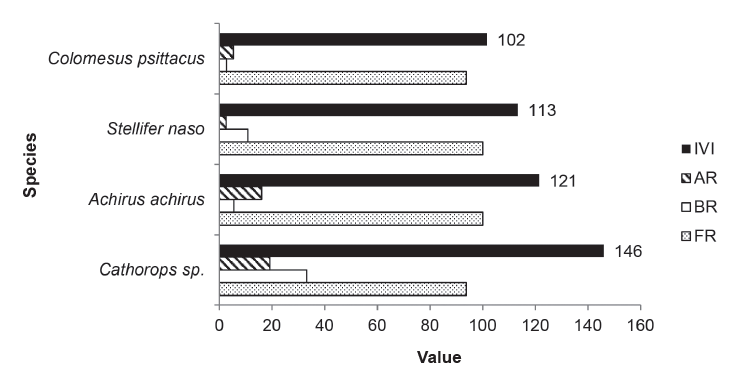
Figure 13.
Importance value index (IVI) for the four dominant species in the Orinoco River Delta and Gulf of Paria channels studied during the hydrocycle. AR = relative abundance, BR = relative biomass, FR = relative frequency.
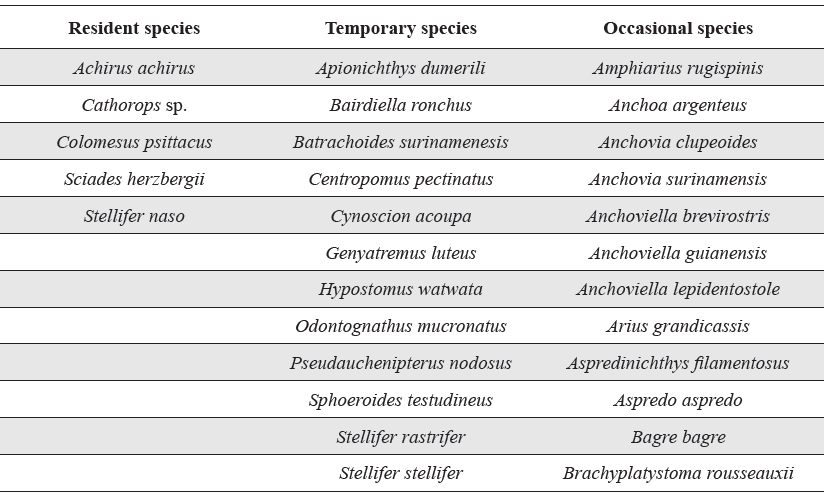
Classification by constancy for the benthic fish species collected from Pedernales, Manamo, Angostura and Venado channels during the hydrocycle. Species are presented in alphabetical order.
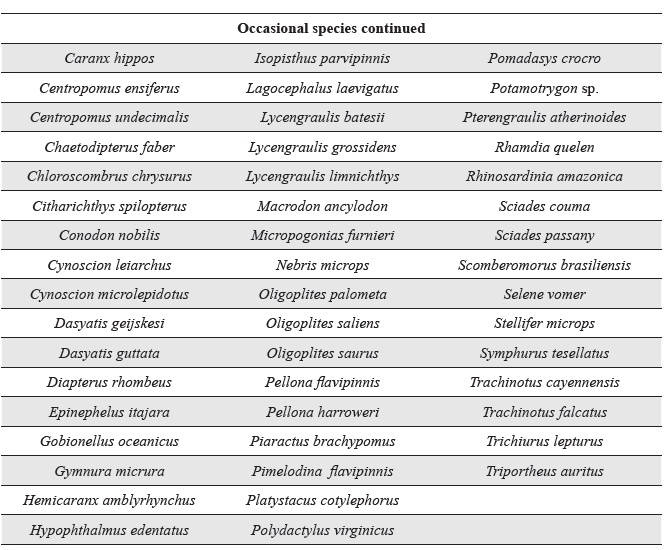
Classification by constancy for the benthic fish species collected from Pedernales, Manamo, Angostura and Venado channels during the hydrocycle. Species are presented in alphabetical order.
Importance Value Index (IVI), Constancy Index (C) and Community Dominance Index (IDC)
To determine the dominant fish species present during the hydrocycle in the four channels studied, the IVI values were calculated for the 78 species collected. The species with IVI values greater than 100 % are shown in Figure 13.
To determine the permanence of fish species in the channels studied, the Constancy index was calculated for the 78 species collected. Five were found to be residents (6 %), 12 temporary (15 %), and the remainder, occasional (78 %) (Table 4).
The community dominance index recorded for each of the hydrophases in the four channels studied and the total are shown in Figure 14. A strong dominance was detected during three of the hydrophases (high water, low water and rising water) as indicated by abundance values above 40 % (Goulding et al. 1988), and for falling water phase the value was close to the cutoff point.
During the high water phase, the marine catfish (Cathorops sp.) and a croaker (Stellifer naso) dominated, but during the falling water phase S. naso and S. stellifer were dominant. During the low water and rising water phases, the most important species were again Cathorops sp. and S. rastrifer.
In Table 5 are given the data of disc width (DW) or standard length (SL) for species of special ecological or commercial interest collected using shrimp trawls.
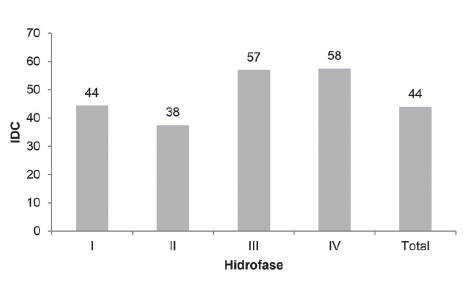
Community dominance index values (IDC) for the four channels studied during each of the four hydrophases and the total.
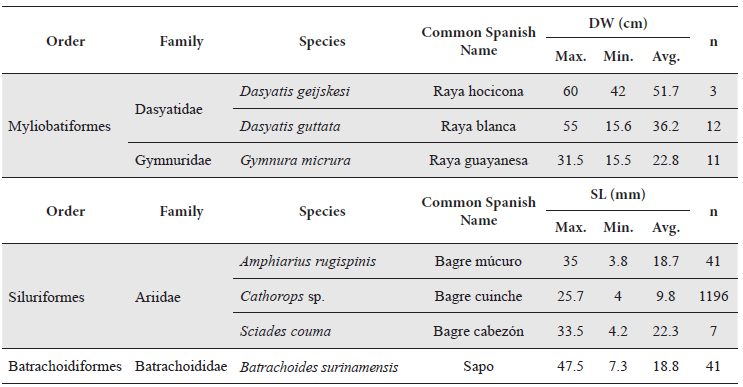
Lengths (DW or SL) of species of special ecological or commercial interest for Pedernales, Manamo, Angostura and Venado channels during the hydrological cycle.
In the case of the marine rays, Dasyatis geijskesi can reach more than 1 m disc width, Dasyatis guttata can reach 2 m (Cervigón 1994) and Gymnura micrura is commonly found at 90 cm but reaches more than 1 m disc width (Cervigón et al. 1992). All individuals of these species collected during this study were juveniles.
Of the marine catfishes, Amphiarius rugispinis, is the most abundant in the lower interior delta of the Orinoco River (Cervigón 1991), in this study it was present in samples from all four of the annual hydrophases but was much more abundant during high water.
It reaches 42 cm SL, is frequently collected around 30 cm (Cervigón et al. 1992), but in this study many individuals (22 %) fell into the 5-10 cm range, followed by those of the 25-30 cm (20 %) interval. Sciades couma can reach 1 m SL and 30 kg weight (Cervigón 1991), in this study the smallest individual measured 4.2 cm and the largest 33.5 cm with an average size of 22.3 cm SL; so all could be considered juveniles.
The toadfish, Batrachoides surinamensis, is known to have been affected by shrimp trawling (Novoa 2000b; Lasso et al. 2004b). A total of 41 individuals were measured, with a minimum size of 7.3 cm SL and a maximum of 47.5, with an average of 18.8. The literature reports that this species reaches 50 cm SL, but that it is commonly collected around 35 cm (Cervigón et al. 1992). Most individuals captured during this study were smaller than 20 cm SL; 63.4 % of the total number of individuals were measured.
The most numerically abundant species was the marine catfish Cathorops sp. which also contributed the largest percentage of biomass captured (Figures 7 and 8). Of the 1196 individuals measured, the smallest was 4 cm and the largest 25.7 cm SL, with an average of 9.8 cm, with most (40.6 %) falling between 8-10 cm. Cervigón et al. (1992) reported a maximum size of 30 cm SL, with most specimens observed around 20 cm. Although the sizes found in this study agree with those reported, the maximum size found was much smaller, and only 1.6 % of the specimens had reached more than 20 cm SL, which was the most commonly observed size in Cervignón’s study. Juveniles are commonly found around sand and mud bars near the mouths of channels where they form immense, dense aggregations; larger adults are typically found further out to sea (Cervigón 1991).
In contrast, just a few individuals of the stingray (Dasyatis guttata) significantly contributed to total biomass captured. Twelve specimens were measured of this species, with SL between 15-55 cm disc width, five of which in the 25- 5 cm size interval. This species can reach 2 m disc width, so the specimens captured in this study were all juveniles.
Discussion
One of the major difficulties encountered when studying tropical estuarine fishes is the determination and quantification of the effects of abiotic parameters on the observed spatial and temporal variations in fish community composition (Andrade-Tubino 2008, Barreiros et al. 2009). The physical and chemical factors that affect the distribution of tropical and subtropical estuarine fishes are rainfall, salinity, turbidity, temperature and depth. With the exception of temperature, in tropical regions all of these are subject to greater fluctuations than those encountered in cold temperate estuaries (Blaber 1997). Temperature is the factor of greatest importance in temperate estuaries, and salinity is the most important in the tropics, where the seasonal cycles of abundance are related to salinity variations (Andrade-Tubino 2008, Barreiros et al. 2009).
The physical and chemical characteristics analyzed in this study did not show seasonal fluctuations during the hydrological cycle, contrary to what has been reported for other tropical and subtropical estuaries (Barletta et al. 2003,2008, Arceo-Carranza and Vega-Candejas 2009, Yáñez-Arancibia et al. 1985, Zubiria et al. 2008). This is a consequence of being cut off from the influence of seasonal freshwater flooding by the construction of the dam on Manamo Channel (Novoa 2000a, Monente & Colonnello 2004). A consequence of this flood regulation was the transformation of a fluvial system into and estuarine system. The tide regime is now the dominant influence in the study area throughout the year, and the only positive influence of freshwater comes from local surface rainfall drainage from the island interiors when annual precipitation exceed the losses to evapotranspiration (Monente & Colonnello 2004).
As Ortaz et al. (2007) mentioned in general terms, the human alterations of the Orinoco River Delta have been few in number and of low intensity, with the exception of what occurred as a result of the closing of Manamo Channel in the 60’s. The construction of that dike modified the hydrological regime of Manamo Channel and so eliminated the pattern of seasonal flooding (Novoa 2000 a, Monente and Colonnello 2004). Echezuría et al. (2002) mention that the flow reduction in Manamo Channel caused a series of hydrochemical changes in the water and in the area of influence, increasing estuarine characteristics in a large portion of the northern delta, including Manamo and Macareo channels. The mixture of the two water types (fresh and salt) has increased the concentrations of chlorine, sodium and magnesium sulphate. High values of those ions can be found many kilometers upstream, near the city of Tucupita.
Of the three physico-chemical parameters measured, the most important in a tropical estuary is salinity (Barletta et al. 2008), since it directly determines the presence of fish species in the different hydrophases, and favors changes in the composition of fish aggregations, leading to the alternating occurrence throughout the year of the species best adapted to the reigning conditions at any given time (Sánchez & Rueda 1999). In contrast to the usual situation, we never found 0 salinity values in any of the four channels studied. The lowest salinity value recorded was 4.8 % from Angostura Channel.
Based on the presence-absence data, a greater similarity was observed between Manamo (Orinoco River Basin) and Venado (Gulf of Paria Basin) channels. This was not the expected result since we expected to find a greater similarity among the Orinoco Basin channels studied rather than a highest correlation between one Orinoco Basin and one Gulf of Paria channels, as occurred with the physico-chemical parameters measured. The obtained result came about because of the 78 fishes collected, Manamo and Venado channels share 11, Manamo, Angostura and Venado share 16, and of the 27 species found in Pedernales Channel, 20 are widely distributed in all four channels.
Novoa (2000a) mentioned that in the mouths of the channels in the northern part of the Orinoco River Delta and southern part of the Gulf of Paria, a little more than half of the species are estuarine-marine species. Novoa and Cervigón (1986) indicated that the northernmost channels of the lower delta, during the rainy season, and with greater water flows, diverse freshwater species were frequently observed such as the juveniles of pimelodid catfishes, and some species of Sciaenidae and Characidae, although, even during the rainy season, the relative importance of these freshwater species is very low in these areas. This same result was obtained in our study of these four channels, where of the 78 species collected, few (7 species, 9 %), were strictly freshwater, and all had very low abundance (fewer than 10 individuals) throughout the entire hydrocycle. Juvenile “morocoto” (Piaractus brachypomus, Serrasalmidae) and catfishes of the family Pimelodidae: “bagre dorado” (Brachyplatystoma rousseauxii), “bagre paisano” (Hypophthalmus edentatus) and “mandi” (Pimelodina flavipinnis) were some of the species found.
In most tropical estuaries, seasonal changes in salinity determine the movement of fishes upstream and downstream in them. During the high water phase, the increase in fresh water causes a decrease in salinity that permits some freshwater species to move into estuarine zones, while marine species move towards the sea, looking for higher salinity waters (Neves et al. 2010). But in this study, no significant spatial or temporal changes in salinity were observed, and so, it is reasonable to assume, since there was no salinity fluctuation, there were no significant changes in the benthic fish community, which throughout the study period was dominate by estuarine-marine species. After the hydraulic regime was regulated in Manamo Channel a series of ecological changes began, among them was the elimination or substitution of the original freshwater fish with an estuarine one (Echezuría et al. 2002).
As for abundance, in all four channels, ten or fewer species comprised 80 % of the captures. The numerical dominance of just a few species is a characteristic observed in tropical and subtropical estuaries, as well as in coastal lagoons, that usually have low species diversity but high abundance of certain species (Whitfield 1999). Cervigón et al. (1992) observed this in different tropical and subtropical estuaries where usually fewer than six species comprised around 70 % of the fishes captured, a phenomenon also observed in other countries such as the estuary in Marañon state (Carvalho-Neta et al. 2011), and Laguna de los Patos in Brazil and the York River in Virginia, USA (Vieira 2006).
The same phenomenon as described above was also observed for biomass. In each of the four channels studied, just ten species contributed more than 80 % of the total catch weight, a characteristic also observed in other tropical and subtropical estuaries (Barletta et al. 2005,2008).
Estuaries are of great ecological importance to fish communities, since they provide protection and refuge for juveniles and adults during the reproductive season (Andrade-Tubino et al. 2008, Blaber et al. 2000, Barletta-Bergan et al. 2001, Barletta et al. 2003). A large percentage of the captures obtained with shrimp trawls are juveniles of benthic fishes that may include more than 20 commercially valuable species, which after they mature may be captured as part of the oceanic fish catch (Ecology & Environment 2003, Lasso et al. 2004b). Lasso et al. (2004b) listed 64 commercially valuable benthic fish species that live in channel mouths and are captured as juveniles by shrimp trawlers. In this study, juvenile phases of all of the 78 fishes were collected from the mouths of the four channels sampled.
The Shannon-Wiener diversity index incorporates equity in its calculation and so is a better evaluation of diversity since it indicates whether a community is dominated by just a few species. Margalef (1974) stated that for fish communities normal values fall between 1.0 and 3.5. However, in estuaries, where marked differences in environmental parameters are the norm, with great fluctuations in physical, chemical and biological parameters, lower diversities are found when compared with other systems (Barros et al. 2011).
In the four channels studied during the annual hydrological cycle no significant differences were detected in any of the indices of diversity, equity or richness. Diversity values (H´) ranged from 0.32 (Pedernales Channel) and 2.68 (Angostura Channel). Magurran (2004) classified diversity as follows: H´1< very low, 1-2 low, 2-3 medium, 3-4 high y > 4 very high. Following this system the values obtained in this study are very low to medium. Lasso et al. (2004b) obtained values from a study done in the same study area between 0.63 a 2.68, that are also considered to be very low to medium, and indicates that these systems are dominated by just a few species.
The lowest equity value (0.14) was found in Pedernales Channel, during rising water phase and the largest (0.83) for Angostura Channel, during high and low water phases. The low equity registered for Pedernales Channel during rising water phase was caused by a small croaker (Stellifer rastrifer) that contributed 95 % of all specimens collected during that phase. The greatest values for Angostura Channel, during high and low water phases indicate that the 20 species collected during those phases had similar abundance, although another small croaker (S. naso) comprised 21 % of the captures during low water, and a flatfish (Achirus achirus) contributed 17 %. Lasso et al. (2004b) reported equity values between 0.29 and the maximum of 1.0.
The lowest richness value (1.5) was observed in Pedernales Channel during rising water phase with 10 species, and the largest were for Manamo Channel (5.24) with 34 species and Angostura Channel (5.29) with 39 species. Lasso et al. (2004b) reported from 1 to 24 species per trawl pull.
In the most recent list of fish species present in the Orinoco River Delta, Lasso et al. (2009) reported a total of 438 species, of which around 150 are benthic species found in the lower delta. The 78 species identified in this study are thus about 50 % of the species reported from the area.
To determine which species are of greatest importance in the community during the complete hydrological cycle, the ecological criteria of abundance, biomass and frequency of occurrence were considered. There is a direct relationship between these criteria and the species with higher IVI values (Figure 13), among which are included the species that are permanent residents in the community (Table 4) and at the same time those that are dominant in the community (Figure 14).
The importance value index (IVI) for the annual hydrological cycle studied indicates that the small marine catfish, Cathorops sp. (IVI = 146), an estuarine-marine species, alone contributed 33 % of the biomass captured, and 20 % of the individuals (abundance) (Figure 15a). In a study in the same area Lasso et al. (2004b) reported that Cathorops spixii, was the only species collected that had an IVI value above 100 %. It was also the species with the highest IVI value (160 %) reported in a similar study of Macareo, Cocuina and Mariusa channels (Lasso et al. 2008). This species predominates in saline waters of the delta, where large quantities of both juveniles and adults form large schools (Cervigón 1985) and it has been reported as the most abundant species in other studies in the region (Novoa 2000b, Lasso et al. 2004b). Species of marine catfishes (Ariidae) can be considered the most important in terms of number of species, density and biomass in tropical and subtropical estuaries (Araújo 1988, Barletta et al. 2003, 2005, 2008, Dantas et al. 2010, Dantas et al. 2011), as has been shown in studies of estuaries throughout the Americas such as Términos Lagoon in México (Lara-Domínguez et al. 1981), Bahía Sepetiba, Río de Janeiro, Brasil (Azevedo et al. 1998) and the Goiana estuary of north-eastern Brazil (Dantas et al. 2010), to name just a few. They are followed by the flatfish, Achirus achirus (IVI = 121), a freshwater, estuarine and marine species responsible for 16 % of the abundance collected and 5.4 % of the biomass captured (Figure 15b). It is one of the most characteristic and abundant species found on muddy substrates of the lower Orinoco River Delta, where both juveniles and adults are captured (Cervigón 1985). Lasso et al. (2004b) in a study done in the same area, that it was fourth in abundance, with an IVI value of around 80 %.
In third place for dominance is the estuarine-marine croaker species Stellifer naso (IVI = 113), with a relative abundance of 3% and biomass of 11 % (Figure 15c). The group of Stellifer species is dominant in terms of biomass of the fish community of the lower delta, with both juveniles and adults common (Cervigón 1985). Lasso et al. (2004 b, 2008) reported Stellifer juveniles as the second most important in dominance (IVI near 100 %) and adults in third place (IVI = 150). Finally, the pufferfish, Colomesus psittacus (IVI = 102), a characteristic species of the lower delta according to Cervigón (1985), placed fourth in dominance. It is an estuarine-marine species, occasionally entering freshwater responsible for 6 % of relative abundance and 3 % of biomass (Figure 15d).
Constancy of the 78 species collected in the channels studied varied. Only five species (6 %) were considered year long residents, 12 (15 %) temporary, and 61 occasional. The fish community is thus composed of many rare species and just a few dominants, which is typical of estuaries (Yañez-Arancibia et al. 1985, Barletta-Bergan et al. 2001) and consistent with the findings of Longhurst and Pauly (2007) who studied tropical systems. According to Day et al. (1989), estuaries are highly dynamic environments, where rapid physical and chemical changes cause high energy costs to the fishes that live there. Because of this, few species are residents in any given locality, and few remain in the estuary for their entire life cycle, with most being just temporary visitors (Santos et al. 2002). This behaviour has been observed in other estuaries, where about 64 % of the fishes captures were occasional visitors, and only 17 % residents (Freitas Jr. 2005).
Although the above situation is the norm for most estuarine fishes, there are a few that dominate in the channels studied throughout the year and all four hydrophases. During high, low and rising waters, the marine catfish Cathorops sp is the dominant species with a croaker species (Stellifer spp) in second place. The community dominance index value for all four channels studied was 44 %, with Cathorops sp and Stellifer naso dominant.

Figure 15.
Most important fish in terms of abundance, biomass and relative frequency (IVI). a) Cathorops sp., b) Achirus achirus, c) Stellifer naso and d) Colomesus psittacus.
Photos: J. Hernández (a), A. Giraldo (b – d).Conclusion
During the hydrological cycle studied, in the mouths of three Orinoco channels (Pedernales, Manamo and Angostura) and one channel in the Gulf of Paria basin (Venado) no fluctuations in the abiotic parameters recorded (depth, turbidity and salinity) were found. Given that salinity is the determinant factor regulating tropical estuarine fish species distribution, no alteration of species present was detected in any of the four channels in any time of the four annual hydrophases. This supports the conclusion that the construction of a dike in Manamo Channel 50 years ago, considerably affected its normal hydrological cycle, and that the impacts not only directly affect Manamo Channel, but extend to the entire Orinoco River subdrainge studied, which includes Pedernales and Angostura channels, and even one channel in the Gulf of Paria basin (Venado), making evident the importance of interconnections among Orinoco River Delta channels.
Acknowledgements
This study was carried out within the framework of the “Aquatic biodiversity monitoring program of the Gulf of Paria and Orinoco River Delta” financed by ConocoPhillips – Venezuela and carried out by the Fundación La Salle de Ciencias Naturales (Venezuela). From the Humboldt Institute we thank Claudia María Villa, for revision of the manuscript and Edwin Fabian Tamayo by the mapping. Special thanks to Donald Taphorn for the translation and the evaluators for their comments.
References
Andrade-Tubino, M., A. L. Reis & M. Vianna. 2008. Organização espaço-temporal das ictiocenoses demersais nos ecossistemas estuarinos Brasileiros: uma síntese. Oecologia Brasiliensis 4: 640-661.
Araújo, F. J. 1988. Distribuição, abundância relativa e movimentos sazonais de bagres marinhos (Siluriformes, Ariidae) no estuário da Lagoa dos Patos (RS), Brasil. Revista Brasileira de Zoologia 5 (4): 509–543.
Arceo-Carranza, D. & M. E. Vega-Candejas. 2009. Spatial and temporal characterization of fish assemblages in a tropical coastal system influenced by freshwater inputs: northwestern Yucatan península. Revista de Biología Tropical 57 (1-2): 89-103.
Azevedo, M. C. C., F. G. Araújo, A. G. da Cruz Filho e A. C. Santos. 1998. Distribuição e abundância relativa de bagres marinhos (Siluriformes, Ariidae) na baía de Sepetiba, Rio de Janeiro. Revista Brasileira de Zoologia 15 (4): 509-543.
Barletta, M., A. Barletta-Bergan, U. Saint-Paul & G. Hubold. 2003. Seasonal changes in density, biomass, and diversity of estuarine fishes in tidal mangrove creeks of the lower Caeté Estuary (northern Brazilian coast, east Amazon). Marine Ecology Progress Series 256: 217-228.
Barletta, M., A. Barletta-Bergan, U. Saint-Paul & G. Hubold. 2005. The role of salinity in structuring the fish assemblages in a tropical estuary. Journal of Fish Biology 66: 45–72.
Barletta, M., C. S. Amaral, M. F. M. Corre, F. Guebert, D. V. Dantas, L. Lorenzi & U. Saint-Paul. 2008. Factors affecting seasonal variations in demersal fish assemblages at an ecocline in a tropical–subtropical estuary. Journal of Fish Biology 73: 1314-1336.
Barletta-Bergan, A., M. Barletta & U. Saint-Paul. 2001. Structure and seasonal dynamics of larval fish in the Caeté River Estuary in North Brazil. Estuarine, Coastal and Shelf Science 56: 193-206.
Barreiros, J. P., J. O. Branco, F. Freitas, L. Machado, M. Hostim-Silva & J. R. Verani. 2009. Space–time distribution of the ichthyofauna from Saco da Fazenda dstuary, Itajaı´, Santa Catarina, Brazil. Journal of Coastal Research 25 (5): 1114-1121.
Barros, D. F., M. F. Torres & F. L. Frédou. 2011. Ichthyofauna of the estuary of São Caetano de Odivelas and Vigia (Pará, Amazon Estuary). Biota Neotropica 11 (2): 367-373.
Blaber, S. J. 1997. Fish and fisheries in tropical estuaries. Chapman & Hall. London, England. 367 pp.
Blaber, S. J. M., D. P. Cyrus, J.-J. Albaret, C. V. Ching, J. W. Day, M. Elliott, M. S. Fonseca, D. E. Hoss, J. Orensanz, I. C. Potter & W. Silvert. 2000. Effects of fishing on the structure and functioning of estuarine and nearshore ecosystems. ICES Journal of Marine Science 57: 590-602.
Carvalho-Neta, R. N. F., J. L. S. Nunes e N. M. Piorski. 2011. Peixes estuarinos do Maranhão. Pp: 95-104. En: Nunes, J. L. S. e N. M. Piorski (Eds.). Peixes marinhos e estuarinos do Maranhão. Café y Lápis, Fapema, São Luís, Brasil.
Cervigón, F. 1982. La íctiofauna estuarina del caño Mánamo y áreas adyacentes. Pp: 162-192. En: Novoa, D. (Ed.). Los recursos pesqueros del río Orinoco y su explotación. Corporación Venezolana de Guyana. Editorial Arte, Caracas, Venezuela.
Cervigón, F. 1985. La ictiofauna de las aguas estuarinas del delta del río Orinoco en la costa atlántica occidental, Caribe. Pp: 57-78. En: Yáñez-Arancibia, A. (Ed.). Ecología de comunidades de peces en estuarios y lagunas costeras: hacia una integración de ecosistemas. UNAM, Ciudad de México, México.
Cervigón, F. 1991. Los peces marinos de Venezuela. Fundación Científica Los Roques, Volumen 1, Segunda edición, Caracas, Venezuela. 425 pp.
Cervigón, F. 1994. Los peces marinos de Venezuela. Fundación Científica Los Roques, Volumen 3, Segunda edición, Caracas, Venezuela. 295 pp.
Cervigón, F., R. Cipriano, W. Fischer, L. Garibaldi, M. Hendrickx, A. J. Lemus, R. Márquez, J. M. Poutiers, G. Robaina y B. Rodríquez. 1992. Guía de campo de las especies comerciales marinas de aguas salobres de la costa septentrional de Sur América. Fichas FAO de identificación de especies para los fines de pesca. FAO, Roma.
Dantas, D. V., M. Barletta, M. F. Costa, S. C. T. Barbosa-Cintra, F. E. Possatto, J. A. A. Ramos, A. R. A. Lima & U. Saint-Paul. 2010. Movement patterns of catfishes (Ariidae) in a tropical semi-arid estuary. Journal of Fish Biology 76: 2540-2557.
Dantas, D. V., M. Barletta, A. R. A. Lima, J. A. A. Ramos, M. F. Costa & U. Saint-Paul. 2011. Nursery habitat shifts in an estuarine ecosystem: Patterns of use by sympatric catfish species. Estuaries and Coasts 35(2): 587 - 602. DOI: 10.1007/s12237-011-9452-0.
Day, J. W., C. A. Hall, W. M. Kemp & A. Yáñez-Arancibia. 1989. Estuarine ecology. John Wiley & Sons, New York. 558 pp.
Ecology & Environment. 2003. La actividad pesquera en el golfo de Paria: actualización del estudio de línea base, CONOCO-Phillips. http://www.conocophillipsparia.com/ files/ LB_Pesquera.pdf. [Consulta: 11 de agosto de 2015].
Flores, A. L., A. Ochoa y G. Love. 2004. Evaluación de amenazas y oportunidades. Pp: 24-28. En: Lasso, C. A., L. E. Alonso, A. L. Flores y G. Love. (Eds.). Evaluación rápida de la biodiversidad y aspectos sociales de los ecosistemas acuáticos del delta del río Orinoco y golfo de Paria, Venezuela. Boletín RAP de Evaluación Biológica 37. Conservation Internacional, Washington DC, USA.
Freitas Junior, F. 2005. Ictiofauna do Estuário do Saco da Fazenda, Itajaí, SC. Itajaí, Monografia. Universidade do Vale do Itajaí, Brasil. 77 pp.
Goulding, M., M. Carvalho Leal & E. Ferreira. 1988. Rio Negro. Rich life in poor water: Amazonian diversity and foodchain ecology as seen trough fishes communities. SPB Academic Publishing, The Hague, Holanda. 200 pp.
Hammer, Ø., D. A. T. Harper & P. D. Ryan. 2001. Palaeontological Statistics software package for education and data analysis. Palaeontologia Electronica 4 (1): 9 pp.
Krebs, Ch. J. 1972. Ecology. Harper & Row, New York, USA. 694 pp.
Lara-Domínguez, A. L., A. Yáñez-Arancíbia & F. Ameuzcua Linares. 1981. Biology and ecology of sea catfish Arius melanopus Gunther in Términos Lagoon, Southern Gulf of Mexico (Pisces: Ariidae). Anales del Instituto de Ciencias del Mar y Limnología 8 (1): 267–304.
Lasso, C. A. y J. Meri. 2003. Estudio de las comunidades de peces en herbazales y bosques inundables del bajo río Guanipa, cuenca del golfo de Paria, Venezuela. Memoria Fundación La Salle de Ciencias Naturales 155: 75–90.
Lasso, C. A., O. M. Lasso-Alcalá, C. Pombo y M. Smith. 2004 a. Íctiofauna de las aguas estuarinas del delta del río Orinoco (caños Pedernales, Mánamo, Manamito) y golfo de Paria (río Guanipa): diversidad, distribución, amenazas y criterios para su conservación. Pp: 70-84. En: Lasso, C. A., L. E. Alonso, A. L. Flores y G. Love. (Eds.), Evaluación rápida de la biodiversidad y aspectos sociales de los ecosistemas acuáticos del delta del río Orinoco y golfo de Paria, Venezuela. Boletín RAP de Evaluación Biológica 37. Conservation Internacional, Washington DC, USA.
Lasso, C. A., O. M. Lasso-Alcalá, C. Pombo y M. Smith. 2004 b. Composición, abundancia y biomasa de la ictiofauna béntica del golfo de Paria y delta del Orinoco. Pp: 85-102. En: Lasso, C. A., L. E. Alonso, A. L. Flores y G. Love. (Eds.), Evaluación rápida de la biodiversidad y aspectos sociales de los ecosistemas acuáticos del delta del río Orinoco y golfo de Paria, Venezuela. Boletín RAP de Evaluación Biológica 37. Conservation Internacional, Washington DC, USA.
Lasso, C. A., O. M. Lasso-Alcalá, A. Giraldo, P. Sánchez, K. González-Oropeza, J. Hernández-Acevedo y J. C. Rodríguez. 2008. Peces. Pp: 73-126. En: Lasso, C. A. y J. C. Señaris (Eds.). Biodiversidad animal del caño Macareo, Punta Pescador y áreas adyacentes, Delta del Orinoco. StatoilHydro Venezuela AS – Fundación La Salle de Ciencias Naturales, Caracas, Venezuela.
Lasso, C. A., P. Sánchez-Duarte, O. Lasso-Alcalá, J. Hernández-Acevedo, R. Martín, H. Samudio, K. González-Oropeza, J. Hernández-Acevedo y L. Mesa. 2009. Lista actualizada de los peces del delta del río Orinoco, Venezuela. Biota Colombiana 10 (1-2): 123-148.
Lasso, C. A. y P. Sánchez-Duarte. 2011. Los peces del delta del Orinoco. Diversidad, bioecología, uso y conservación. Fundación La Salle de Ciencias Naturales y Chevron C. A., Caracas, Venezuela. 498 pp.
Longhurst, A. R. e D. Pauly. 2007. Ecologia dos oceanos tropicais. Edusp, São Paulo, Brasil. 424 pp.
Magurran, A. E. 2004. Measuring biological diversity. Blackwell Publishing: Oxford, United Kingdom. 215 pp.
Margalef, R. 1969. Perspectives in ecological theory. TheUniversity of Chicago, Chicago. 111 pp.
Margalef, R. 1974. Ecología. Ediciones Omega S. A. Barcelona, España. 951 pp.
Mc Naughton, S. J. 1968. Ecology Structure and function in California grassland. Ecology 49: 962-972.
Monente, J. A. y G. Colonnello. 2004. Consecuencias ambientales de la intervención del delta del Orinoco. Pp. 114-125. En: C. A. Lasso, L. E. Alonso, A. L. Flores y G. Love (Eds.), Evaluación rápida de la biodiversidad y aspectos sociales de los ecosistemas acuáticos del delta del río Orinoco y golfo de Paria, Venezuela. Boletín RAP de Evaluación Biológica 37. Conservation Internacional, Washington DC, USA.
Montgomery, D. 1984. Design and analysis of experiments. Second edition, John Wiley & Sons, Nueva York, USA. 250 pp.
Neves, L., T. Teixeira & F. Araújo. 2010. Structure and dynamics of distinct fish assemblages in three reaches (upper, middle and lower) of an open tropical estuary in Brazil. Marine Ecology 32: 115-131.
Novoa, D. (comp.). 1982. Los recursos pesqueros del Río Orinoco y su explotación. Corporación Venezolana de Guayana, Editorial Arte, Caracas, Venezuela. 386 pp.
Novoa, D. 2000 a. La pesca en el golfo de Paria y delta del Orinoco costero. CONOCO Venezuela, Editorial Arte, Caracas, Venezuela. 120 pp.
Novoa, D. 2000 b. Evaluación del efecto causado por el efecto de la pesca de arrastre costera sobre la fauna íctica en la desembocadura del caño Mánamo (Delta del Orinoco, Venezuela). Acta Ecologica del Museo Marino de Margarita 2: 43-62.
Novoa, D. y F. Cervigón. 1986. Resultados de los muestreos de fondo en el área estuarina del Delta del río Orinoco, Venezuela. En: IOC/FAO Workshop on Recruitment in Tropical Coastal Demersal Communities. Intergovernmental Oceanographic Comission, Workshop Report 44 (Supplemento), Roma, Italia.
Olivares, E. y G. Colonnello. 2000. Salinity gradient in the Mánamo river, a dammed distributary of the Orinoco delta, and its influence on the presence of Eichhornia crassipes and Paspalum repens. Interciencia 25 (5): 242-248.
Ortaz, M., A. Machado-Allison y V. Carrillo. 2007. Evaluación ecológica rápida de la ictiofauna en cinco localidades del delta del río Orinoco, Venezuela. Interciencia 32(9): 601-609.
Petróleos de Venezuela (PDVSA). 1993. Imagen Atlas de Venezuela. Una visión espacial. Editorial Arte, Caracas, Venezuela. 271 pp.
Ponte, V., A. Machado-Allison y C. A. Lasso. 1999. La ictiofauna del delta del río Orinoco, Venezuela: una aproximación a su diversidad. Acta Biológica Venezuélica 19 (3): 25-46.
Ramos, F., D. Novoa e I. Itriago. 1982. Resultados de los programas de pesca exploratoria realizados en el Delta del Orinoco. Pp: 162-192. En: Novoa, D. (Ed.), Los recursos pesqueros del río Orinoco y su explotación. Corporación Venezolana de Guyana, Editorial Arte, Caracas, Venezuela.
Rodríguez, M. A., K. O. Winemiller, W. M. Lewis & D. C. Taphorn. 2007. The fresh water habitats, fishes and fisheries of the Orinoco River basin. Aquatic Ecosystem Health 10 (2): 140-152.
Sánchez, C. y M. Rueda. 1999. Variación de la diversidad y abundancia de especies ícticas dominantes en el Delta del Río Magdalena, Colombia. Revista de Biología Tropical 47 (4): 1067-1079.
Santos, C., R. Schwarz-Junior, J. F. de. Oliveira-Neto e H. L. A. Spach. 2002. Ictiofauna em duas planícies de maré do setor euhalino da Baía de Paranaguá, Pr, Brasil. Boletim do Instituto de Pesca 28: 49-60.
Shannon, C. E. & W. Weaver. 1963. The mathematical theory of communication. University of Illinois, Urbana. 119 pp.
Vieira, J. P. 2006. Ecological analogies between estuarine bottom trawl fish assemblages from Patos Lagoon, Rio Grande du Sul, Brazil and York River, Virginia, USA. Revista Brasileira de Zoologia 23 (1): 234 – 247.
Whitfield, K. A. 1999. Ichthyofaunal assemblages in estuaries: A South African case study. Reviews in Fish Biology and Fisheries 9:151-186.
Yáñez-Arancibia, A., A. L. Lara-Domínguez & H. Álvarez Guillén. 1985. Fish community ecology and dynamic in estuarine inlets. Pp. 127-168. En: Yáñez-Arancibia, A. (Ed.), Ecología de comunidades de peces en estuarios y lagunas costeras: hacia una integración de ecosistemas. UNAM, Ciudad de México, México.
Zubiria, W., A. Lacayo, A. Acero y J. C. Narváez. 2008. Diversidad y abundancia de la ictiofauna de un complejo de lagunas costeras en una reserva natural del Caribe colombiano. Memoria Fundación La Salle de Ciencias Naturales 68 (170): 125-139.
Appendix 1.
List of fish species collected during one year of sampling in Pedernales, Manamo, Angostura and Venado channels, of the Orinoco River Delta and the Gulf of Paría. Freshwater (F), Occasional Freshwater (OF), Estuarine (E), Estuarine Occasional (EO) and Marine (M).
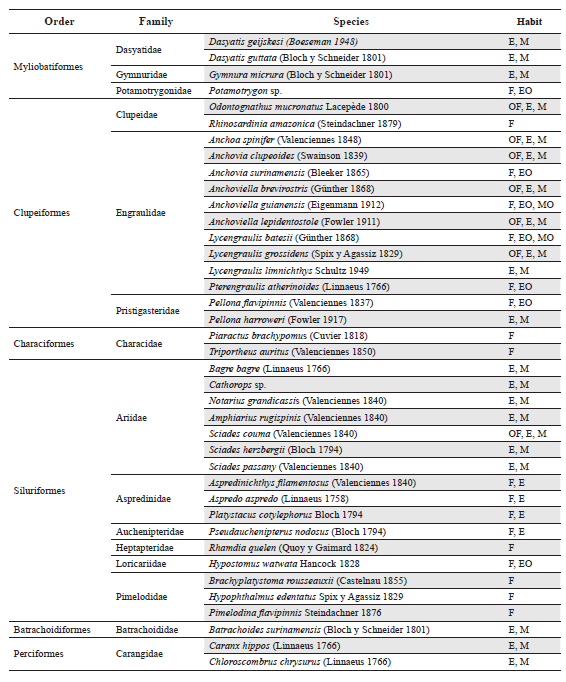
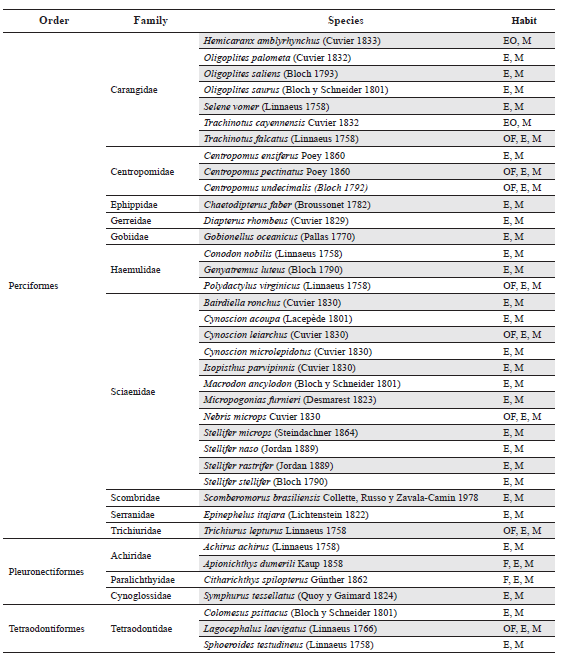
List of fish species collected during one year of sampling in Pedernales, Manamo, Angostura and Venado channels, of the Orinoco River Delta and the Gulf of Paría. Freshwater (F), Occasional Freshwater (OF), Estuarine (E), Estuarine Occasional (EO) and Marine (M).
Additional information
Citación del artículo. : Sánchez-Duarte, P. y C. A. Lasso. 2016. Benthic fish community structure in the Orinoco River Delta and Gulf of Paria (Venezuela), fifty years after the construction of a dike across Manamo Channel. Biota Colombiana 17 (1) 64-89. DOI: 10.21068/C2016v17r01a06