Recepción: 08 Octubre 2015
Aprobación: 12 Mayo 2017
DOI: https://doi.org/10.21068/c2017.v18n01a16
Resumen: La creciente incertidumbre sobre el estado de conservación de los diferentes grupos biológicos resalta la importancia de trabajos enfocados en la recopilación de registros de diversidad. Para Colombia, la exploración biológica ha implicado el descubrimiento de zonas prioritarias de conservación, pero la falta de información sobre aspectos como la distribución de las especies, limita tanto el énfasis como la efectividad en los esfuerzos de conservación. En particular, el departamento del Quindío representa un área de interés faunístico importante por su posición geográfica y características climáticas. Sin embargo, carece de información base para una discusión inclusiva de la diversidad local. El presente trabajo recopila información bibliográfica sobre la riqueza, distribución y diversidad de anfibios en el departamento. La información publicada en diferentes fuentes bibliográficas indica que cerca de 105 especies de anfibios y reptiles (48 y 57 respectivamente) coexisten dentro de los límites políticos del departamento. Entre estas, los géneros Anolis y Pristimantis representan la mayor diversidad de especies. A pesar de su extensión política, las especies que allí han sido registradas presentan una importancia ecológica y actúan, además, como un reservorio de diversidad en relación a otras zonas.
Palabras clave: Anfibios, Cordillera Central, Conservación, Reptiles, Riqueza.
Abstract: The increasing uncertainty about the conservation status of the different biological groups emphasizes the importance of concentrating efforts on gathering diversity records. In Colombia, biological explorations have revealed priority areas for conservation, but the lack of information about topics as species distribution, restricts both the emphasis and efficiency of conservation efforts. In particular, Quindío department represents an important area of faunal interest due to its geographical position and climatic conditions. However, still lacks of basic information for an comprehensive discussion of local diversity. This works aims to summarize the information on richness, geographic distribution and diversity of the herpetofauna in the department. Published information from different bibliographic resources indicates that about 108 species of amphibians and reptiles (48 and 57 spp. respectively) coexist within the political boundaries. Among these, the genus Anolis and Pristimantis recorded the highest species diversity. Despite the reduced department’s size, the species that have been recorded exhibit remarkable ecological importance and also act as a reservoir of diversity regarding surrounding areas.
Keywords: Amphibians, Cordillera Central, Conservation, Reptiles, Richness.
Introducción
Se ha identificado a la cordillera de los Andes como el principal factor de diversificación en la región Neotropical (Antonelli et al. 2009, Rull 2011, Mendoza et al. 2015), pues al fragmentar poblaciones de tierras bajas y altas por medio de levantamientos asincrónicos, ha generado barreras al flujo génico que han promovido los eventos de especiación principalmente por alopatría (Lynch et al. 1997, Cadena et al. 2012, Madriñan et al. 2013). Además de las variaciones geológicas, la incapacidad inherente de los organismos (e.g. conservatismo de nicho) para adaptarse a condiciones ecológicas nuevas, acoplada a los cambios ambientales entre cordilleras (Antonelli et al. 2009, Hoorn et al. 2010), incrementan la verosimilitud y la preponderancia de este modo de especiación en los vertebrados neotropicales (e.g. especiación alopátrica; Wiens 2004, Wiens y Graham 2005). Entre las consecuencias más evidentes de la alopatría a nivel ecológico están la baja diversidad local y el alto recambio de especies (Cadena et al. 2012), siendo ambos descriptores típicos de las zonas altas andinas (Lynch 1999, Cadena et al. 2012, Muñoz-Ortiz et al. 2014). Por otro lado, esta región alberga más de la mitad de especies de reptiles (Sánchez-C. et al. 1995) y de anfibios (Ruiz–Carranza et al. 1996, Bernal y Lynch 2008), pero además presenta la riqueza y diversidad que hace distintiva al país (e.g. endemismos; Lynch et al. 1997, Sánchez–C. et al. 1995).
A pesar de la notable diversidad biológica que sustentan los Andes colombianos, esta área ha sido gravemente afectada por la intervención antrópica. En la actualidad, esta región sustenta cerca del 85 % de las actividades socioeconómicas y alberga más del 77 % de la población del país, convirtiéndose en el núcleo socioeconómico más importante del país (Corrales 2002, Rincón y Bernal 2007). La actual tendencia al incremento en la explotación pecuaria y la expansión acelerada de la frontera agrícola han hecho de la región Andina, el área con mayor índice de transformación ecológica mediada por efectos antrópicos (Etter y van Wyngaarden 2000, Murgueitio 2002, Etter et al. 2008). En general, esto ha implicado la disminución en extensión para bosques primarios en por lo menos 63 % (Corrales 2002, Etter et al. 2016), con notables implicaciones sobre diversidad biológica (Myers et al. 2000, Corrales 2002, Herzog et al. 2012).
Tanto anfibios como reptiles son considerados grupos sensibles ante la degradación ambiental. En general y como consecuencia de su fisiología, ambos grupos presentan diferentes respuestas en el contexto espacial de su riqueza, su comunidad y/o poblaciones al ser sometidos a estrés ambiental (Duellman y Trueb 1994, Di Tada et al. 1996, Driscoll 2004, Sanabria et al. 2007, Bionda et al. 2012, Hernández–Córdoba et al. 2013, Theisinger y Ratianarivo 2015). Un patrón recurrente en la región Andina relaciona una alta diversidad de anfibios en altitudes elevadas en comparación con reptiles (Navas 2002). Mientras que la relación funcional entre el número de especies y la elevación en reptiles está dada por la temperatura, el patrón es variable y dependiente principalmente de la localidad analizada (McCain 2010). En síntesis, la alta capacidad explicativa de los componentes microambientales soportan la estrecha dependencia de ambos linajes a las condiciones del entorno y ofrecen una explicación a la variación altitudinal de la riqueza especifica (Lomolino 2001, Cortés–Gomez et al. 2013). Aunque esta relación puede ser generalizada, el patrón altitudinal de diversidad puede variar de acuerdo con la región biogeográfica, grupo taxonómico, interacción trófica, producción, estabilidad o antigüedad de los ecosistemas (Patterson et al. 1989). Por lo que limita las conclusiones a espacios y tiempos particulares (Rickart et al. 1991). A grandes rasgos, esto puede reflejar la falta de comprensión sobre la generalidad del patrón o la realidad sobre la relación funcional entre elevación y riqueza (Kattan et al. 2004). Sin embargo, ha sido ampliamente soportada la mayor riqueza especifica en regiones con elevaciones intermedias (Graham et al. 2004, La Sorte y Jetz 2010, Santos et al. 2011).
Aunque se reconoce que la respuesta es diferencial entre anfibios y reptiles, los procesos de transformaciones globales como la pérdida de hábitat, el cambio climático, enfermedades, minería, uso de pesticidas y tráfico ilegal han ocasionado que muchas poblaciones de ambos grupos se encuentren en un estado de declive (Mendelson et al. 2006, Böhm et al. 2013, Valencia-Zuleta et al. 2014). Una de las principales problemáticas es el desconocimiento de la diversidad biológica, pero en particular en grupos focales. Hasta la fecha, únicamente el 25 % de anfibios y 18 % de reptiles evaluados se encuentran en Datos Insuficientes (DD sensu IUCN 2016). Si se reconoce que las regiones montañosas tropicales son “hotspots” de riqueza y endemismo para múltiples linajes de anfibios y reptiles (Myers et al. 2000, Orme et al. 2005), estas áreas deberían ser consideradas como prioridad en la evaluación, ejecución de criterios y planes de manejo que aporten en la conservación de la herpetofauna.
En el departamento del Quindío, pocos estudios han sido enfocados en describir la diversidad de herpetofauna. Hasta la fecha de redacción de este documento, Cadavid et al. (2005) es el único trabajo que recopila de manera sistemática información sobre la diversidad de anfibios y reptiles en el departamento, lo cual contrasta con otros trabajos enfocados en registrar observaciones de una o pocas especies (e.g. Vanegas-Guerrero et al. 2014, 2016a). La falta de una síntesis sobre la riqueza de estos grupos, genera la imposibilidad de discusiones subsecuentes en aspectos como delimitación de regiones con prioridades de conservación. El propósito del presente trabajo es recopilar la información contenida en diferentes fuentes bibliográficas (i.e revisiones taxonómicas, registros geográficos, inventarios taxonómicos reducidos), sobre la riqueza de herpetofauna en el departamento del Quindío. En el presente documento se analiza también su diversidad y distribución altitudinal.
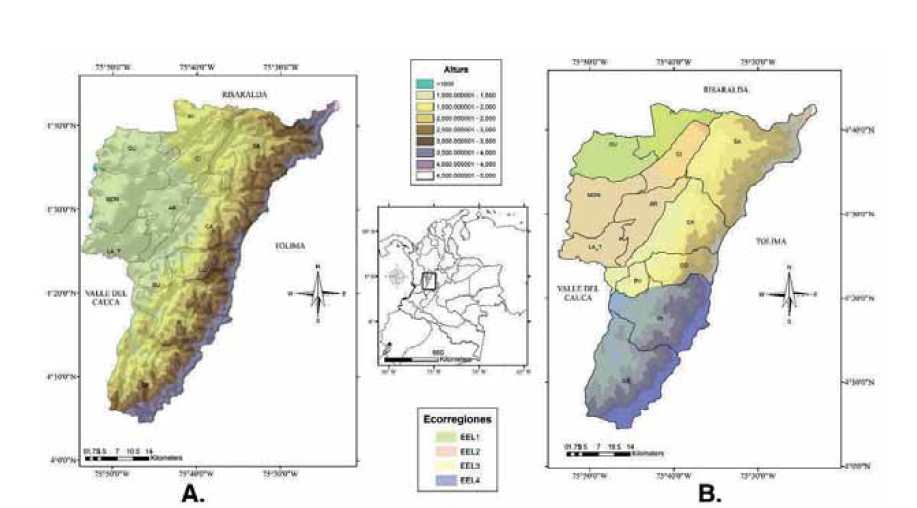
Figura 1
Departamento del Quindío, Colombia. A) Mapa político del departamento, con rangos altitudinales; B) segmentación por ecorregiones según la propuesta de Orozco–Serna (2012). Acrónimos de municipios: (Ar) Armenia, (SA) Salento, (LA_T) La Tebaida, (Fi) Filandia, (Qu) Quimbaya, (Mon) Montenegro, (Ci) Circasia, (Ca) Calarcá, (Bu) Buenavista, (Co) Córdoba, (Pi) Pijao y (Ge) Génova.
Material y métodos
Área de estudio
El departamento del Quindío se ubica en la zona centro-occidental de Colombia, entre los 4°04’41’’ N y 4°43’18’’ N y entre los 75°23’41’’W y 75°53’56’’ W, con una superficie aproximada de 1.845 km2 (Figura 1). El área consiste en un intrincado mosaico de ecosistemas y pisos térmicos dispuestos en un gradiente altitudinal ascendente desde el extremo occidental del departamento (800 m s.n.m.) hacia la zona oriental (>3500 m s.n.m.), donde aparecen las laderas propias de la vertiente occidental de la cordillera Central (Cadavid et al. 2005).
Obtención de datos
La información fue obtenida a partir de fuentes de bibliografía y registros disponibles en las bases de datos en línea del Instituto de Ciencias Naturales (ICN), Instituto de Investigación de Recursos Biológicos Alexander von Humboldt (IAvH), Museo herpetología de la Universidad de Antioquia (MHUA–A), Colección de anfibios y reptiles de la Universidad del Valle (UVC), Fundación Miguel Lillo (FML), Museo de La Salle – Universidad de La Salle (MLS), American Museum of Natural History, New York, USA (ANSP), Museo de Historia Natural, Colegio San José, Medellín, Colombia (CSJ), Museum of Comparative Zoology, Harvard University, Cambridge, Massachussetts, USA, (MCZ), United States National Museum, Smithsonian Institution, Washington, D.C., USA (USNM). La nomenclatura usada en este listado sigue las propuestas taxonómicas de Pyron y Wiens (2011) para anfibios y Uetz y Hošek (http://www.reptile–database.org, acceso: 24/07/2016) para reptiles.
Una vez generada la matriz con las especies y sus localidades se realizaron dos tipos de análisis: 1) Distribución altitudinal de las especies: se consideran siete rangos altitudinales de aproximadamente 500 m s.n.m.: R1: <1000 m s.n.m., R2: 1000 - 1500 m s.n.m., R3: 1501 - 2000 m s.n.m., R4: 2001 - 2500 m s.n.m., R5: 2501 - 3000 m s.n.m., R6: 3001 - 3500 m s.n.m. y R7: > 3501 m s.n.m. Este tipo de análisis por rangos de 500 m ha sido realizado previamente en diferentes localidades, aportando suficiente resolución para la descripción de la distribución altitudinal en diferentes grupos (e.g. Wathen et al. 2014). 2) Segmentación por ecorregiones: siguiendo la propuesta de Orozco–Serna (2012), se tomaron cuatro zonas delimitadas por corrientes hídricas y apoyada en las caracterís-ticas biológicas de cada municipio denominadas de la siguiente manera: ecorregión I (EEL I): Filandia y Quimbaya; ecorregión II (EEL II): Circasia, Armenia, Montenegro y La Tebaida; ecorregión III (EEL III): Salento, Calarcá, Córdoba y Buena Vista y ecorregión IV (EEL IV): Pijao y Génova (Figura 1B).
Análisis de datos
Se interpretó la diversidad alfa tanto en el departamento, municipios y ecorregión, como el número de especies reportadas en cada uno de esos sitios. Se calculó además el índice de Whittaker entre pares de ecorregiones y global, tanto para anfibios como reptiles. Se evaluaron por lo tanto los patrones espaciales tanto políticos (e.g. municipios) como altitudinales. La diversidad altitudinal (beta diversidad) fue evaluada de acuerdo a las matrices de riqueza entre rangos altitudinales a partir de un análisis de agrupamiento basado en el índice de Jaccard. Este análisis complementó funcionalmente con los valores de diversidad del índice de Whittaker. Por último, para encontrar las especies responsables de las diferencias entre rangos altitudinales se realizó un análisis de SIMPER que indica el aporte en la disimilitud de cada taxón sobre la totalidad de grupos (e. g. rangos altitudinales) en la matriz de riqueza. No se incluyeron en los análisis algunas especies consideradas como introducidas en el departamento por no presentar en algunos casos poblaciones naturales dentro de la zona de estudio. Los análisis fueron realizados en usando la librería Vegan (Oksanen et al. 2007) en R (R Core Team 2016, http://www. R-project. org).
Resultados
La herpetofauna del Quindío se compone de 105 especies: 48 anfibios y 57 reptiles (Anexo 1, Tabla 1). Entre estas, los anuros representan el 41 % de las especies, seguido de serpientes con 32 % (30 sp.) y saurios con un 20 % (22 sp.). Otros grupos como Caudata (1 %), Gymnophiona (3 %) y Testudinata (4 %) fueron los menos representativos de la riqueza del departamento. Se registró un total de 12 familias de los tres órdenes de anfibios. Ocho familias de anuros, tres de gimnofiones y una de caudados (Figura 2A). Craugastoridae fue la familia más diversa (19 sp.; 39,58 %), seguida por Centrolenidae (8 sp., 16,66 %) y Bufonidae (3 sp., 6,25 %). En contraste, las familias con menor diversidad fueron Hemiphractidae y Leptodactylidae (2 sp., 4,16 % cada una) y Ranidae, Caeciliidae, Siphonopidae, Plethodontidae y Typhlonectidae (1 sp. cada una).
Los reptiles reportados en el departamento se distribuyen en 16 familias representando los órdenes Squamata (13 familias), con los subórdenes Sauria (8 familias) y Serpentes (5 familias), y el orden Testudinata (3 familias) (Figura 2B). Dipsadidae fue la familia con mayor número de especies registradas (19 sp.; 33,33 %), seguido de Dactyloidae (7 sp.; 12,28 %), Gekkonidae y Gymnopthalmidae (4 sp.; 7 % cada una). Iguanidae, Chelydridae, Scincidae, Corytophanidae, Tropiduridae, Geoemydidae, Emydidae, Kinosternidae y Viperidae se encontraron representadas por una sola especie.
La mayor parte de los registros estuvieron localizados sobre la zona norte del departamento (Figura 3), en los municipios de Filandia (44 sp.), Calarcá (33 sp.), Armenia (29 sp.) y Salento (27 sp.). Mientras que La Tebaida (7 sp.), Montenegro (7 sp.), Pijao (5 sp.), Quimbaya (4. sp.), Circasia (2 sp.) y Génova (1 sp.), presentaron los registros más bajos. En contrastante la relación entre riqueza anfibios/reptiles en diferentes municipios como Salento que presenta registros de 34 especies de anfibios y solo dos registros de reptiles.
Diversidad por ecorregiones
El mayor número de especies tanto de anfibios como reptiles se encontró en la ecorregión I, seguida de la II y III (52, 47 y 46 sp. respectivamente; Figura 4). La EEL IV, que incluye los municipios Pijao y Génova, presentó solamente 12 registros (11,11 %). Para anfibios, el número de especies fue contrastante entre la EEL III (32 sp.) y EEL IV (9 sp.), resaltando esta última por la presencia de especies únicas para el departamento como: Bolitoglossa vallecula, Nymphargus ruizi, N. grandisonae y Parvicaecilia pricei. Las ecorregiones ubicadas al noroccidente del departamento (EEL I y EEL II) presentaron un número de especies intermedio con relación a las demás (25 y 15 sp., respectivamente). Por otro lado, Pristimantis achatinus, Rhinella horribilis, Dendropsophus columbianus y Colostethus fraterdanieli presentaron registros en las cuatro ecorregiones, mientras que Centrolene savagei, Pristimantis erythropleura, P. palmeri, P. thectopternus, B. vallecula y Nymphargus griffithsi estuvieron presentes en el 75 % de las ecorregiones.
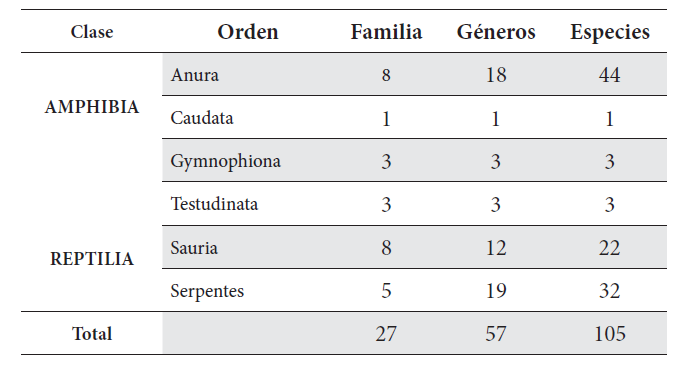
Tabla 1.
Resumen de la composición de la herpetofauna del departamento del Quindío, Colombia.
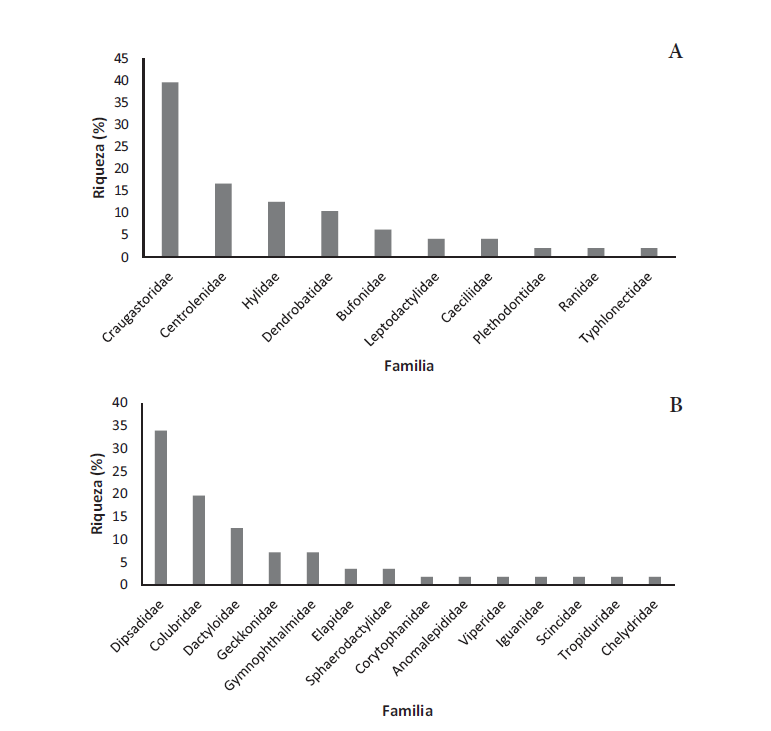
Figura 2.
Riqueza (%) de las familias en el departamento del Quindío, Colombia. A) Anfibios, B) Reptiles.
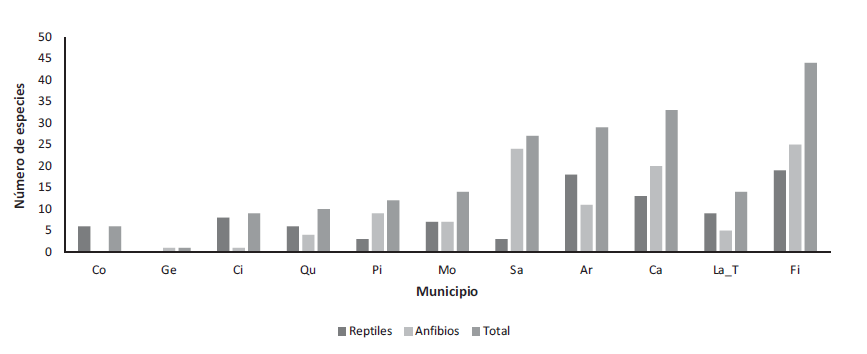
Figura 3.
Distribución de la riqueza en los municipios con registros de herpetofauna en el departamento del Quindío. Fi: Filandia, Sa: Salento, Ca: Calarcá, Ar: Armenia, La_T: La Tebaida, Mo: Montenegro, Ge: Génova, Qu: Quimbaya, Pi: Pijao, Ci: Circasia, Co: Córdoba.
Los reptiles presentaron la mayor diversidad en la EEL II (31 sp., 29,48 %) zona enmarcada en los municipios de La Tebaida, Montenegro y Armenia. En esta ecorregión se destacan los únicos registros de Chelydra acutirostris, Leptodeira septentrionalis, Tantilla melanocephala, Mastigodryas danieli y Basiliscus galeritus. La EEL IV presentó el menor número de especies para reptiles (3 sp.), correspondientes a Chironius montícola, Bothriechis schlegelii y Micrurus mipartitus. Las EEL I y EEL III recogen cerca del 63 % de riqueza (40 sp.) de reptiles en el departamento.
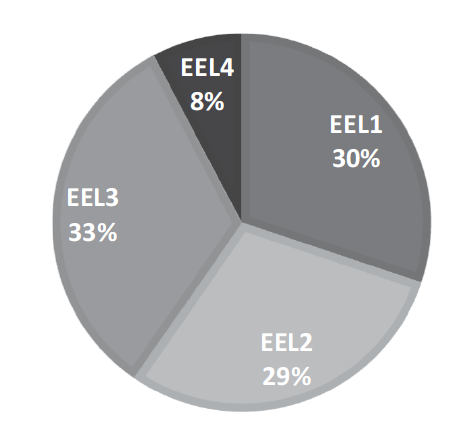
Figura 4.
Riqueza porcentual de la herpe-tofauna por ecorregión en el departamento del Quindío, Colombia.
Análisis altitudinal
La riqueza de anfibios presentó una distribución gaussiana de acuerdo al gradiente altitudinal, con un pico máximo de riqueza en altitudes intermedias en el rango comprendido entre los 2000-2500 m s.n.m. (R4; 32 sp.) y disminuyendo gradualmente hacia los extremos (Figura 5). Con respecto a R1 disminuye un 75 % y 81 % cuando se compara con R7. El 71 % de las especies de anfibios en el departamento se encontraron distribuidas entre los 1500-2500 m s.n.m. (R3-R4; 34 sp). Los reptiles presentan una tendencia inversa entre la riqueza y altitud, con un valor máximo entre los 1500-2000 m s.n.m. (R2; 47 sp.), es decir un mayor número de registros (68 %) a alturas menores de 1500 m s.n.m. (R1- R2; 40 sp.).
Amplitud altitudinal de los anfibios dentro de la gradiente
Se encontraron registros de 12 familias en el rango de los 500 – 3501 m. R1 se caracterizó por presentar el único reporte de Typhlonectes natans (Typhlonectidae) para el departamento. Tanto R1 como R2 tuvieron mayor dominancia de hylidos (3 sp.) y leptodactylidos (2 sp.). En R3 y R4 se evidenció alta dominancia de craugastoridos (9 y 12 sp., respectivamente), centrolenidos (6 y 7 sp., respectivamente) y dendrobatidos (4 y 5 sp., respectivamente). R5 y R6 presentaron un patrón similar en riqueza de Craugastoridae (9 sp. cada una). Sin embargo, en R5 se presentó un número mayor de especies de las familias Bufonidae y Centrolenidae (2 sp. cada uno), en comparación con R6 en el cual solo se registró una especie para ambas familias. En R7 se encontraron únicamente registros de Craugastoridae (5 sp.) y Bufonidae (1sp.). Craugastoridae estuvo presente en todos los rangos, excepto en R1, dominando en el número de especies desde los 1.500 m s.n.m.
Amplitud altitudinal de las especies de reptiles
Se encontraron registros de 16 familias para el departamento, registrando para Dipsadidae la mayor riqueza en alturas menores de 3000 m s.n.m. (R1-R5). R1 y R2 estuvieron más representados por especies de Dipsadidae (13 y 14 spp., respectivamente), Colubridae (10 y 9 sp., respectivamente), Gekkonidae (4 sp. en cada una) y Dactyloidae (2 y 6 sp., respectivamente). C. acutirrostris (Chelydridae) solo se registró en R1. Para R3, las familias mejor representadas fueron Dipsadidae (18 sp.), Dactyloidae (7 sp.) y Gymnophthalmidae (5 sp.). A partir de los 2.000 m s.n.m., la riqueza de estos grupos disminuye, pero aún es notoria la dominancia en R4 y R5 por las familias: Dipsadidae (15 y 13 sp., respectivamente), Dactyloidae (5 sp. y 2 sp., respectivamente), Colubridae (5 sp. y 3 sp., respectivamente) y Gymnophthalmidae (4 sp. en R4). En R6, solo se encontraron registros de Atractus crassicaudatus (Dipsadidae), Riama striata (Gymnophthalmidae) y Anolis heterodermus (Dactyloidae), siendo ésta última la única representante a 3.500 m de altura (R7).
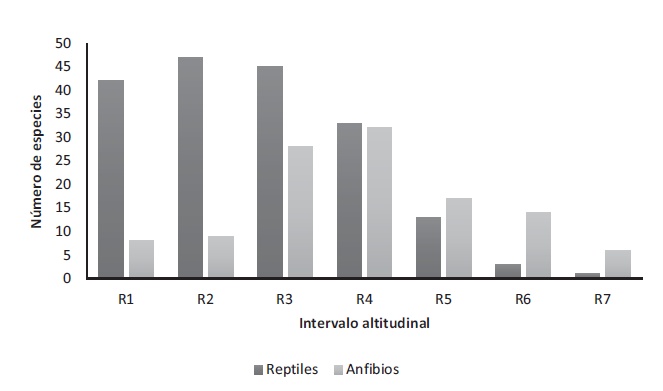
Figura 5.
Distribución altitudinal de la herpetofauna en el departamento del Quindío.
Patrones de Beta diversidad en anfibios
Se encontró una similitud del 76 % (Bootstrap = 100 %) entre la composición de los rangos altitudinales R3 y R4, que sugiere la continuidad en el gradiente altitudinal de la fauna anfibia entre los 1500 y 2500 m s.n.m. (Figura 6A). Dos grupos son discriminados en el análisis de agrupamiento compuesto por R3 a R7 y R1 con R2. El grupo que presentó la composición más disímil con respecto a los demás rangos altitudinales fue R7 (Bootstrap = 87 %), este rango comparte solo entre 15 a 30 % de similaridad con R5 y R6. R1 con R2 evidencia la similaridad de las zonas bajas (55 %, Bootstrap = 100), compartiendo un porcentaje de la fauna anfibia que genera una tendencia continua en la identidad del ensamble.
El índice de disimilitud de Whittaker indicó una notable identidad de los rangos extremos en cuanto a sus características composicionales (Whittaker= 1,00, Tabla 2). Los rangos extremos forman por lo tanto estructuras independientes respecto a los rangos altitudinales intermedios. Por último, los rangos R3 y R4 presentan el mayor número de especies y por lo tanto la mayor similaridad entre todos los 1500–2500 m s.n.m. (Whittaker= 0,15). En general, el patrón de agrupamiento entre rangos altitudinales responde a la presencia de especies como Pristimantis alalocophus (2,84 %), Osornophryne percrassa (2,84 %), Dendropsophus columbianus (2,84 %), Pristimantis permixtus (2,83 %) y Rhinella horribilis (2,83 %). Las especies Colostethus ucumari (0,84 %) y Centrolene geckoideum (0,84 %) implican la mayor disimilitud entre rangos y por lo tanto son exclusivas de determinadas alturas.
Patrones de Beta diversidad en reptiles
La mayor similitud entre rangos altitudinales se encontró en el clúster formado por R1 y R2 (80 %, Bootstrap= 99 %; Figura 6B). Al igual que este, los rangos R3 y R4 fueron identificados como con una estructura similar. Estos últimos fueron anidados con R5 entre un 30-40 % de similaridad (Bootstrap=41 %), donde comienza a cambiar la identidad del gradiente. La formación de estos grupos implica que para reptiles se encuentran tendencias a ensambles de tierras bajas-medias y altas separados por los 3000 m s.n.m. Esto fue indicado en el Cluster de R6-R7 (45 %, Bootstrap=65 %). El índice de Whittaker que presentó un recambio general de 1.18 que soporta los Clusters formados a partir del índice de Jaccard (Tabla 2). El análisis de SIMPER indicó que la diferencia entre alturas fue impuesta principalmente por A. crassicaudatus (4,328 %).
Tabla 2.
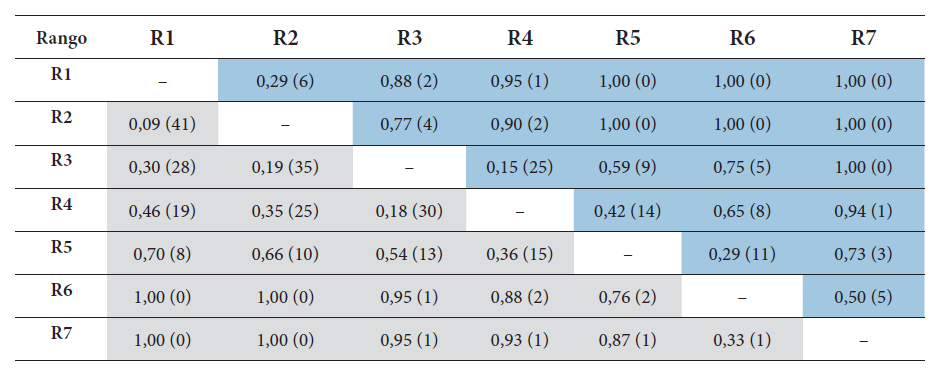
Recambio de especies de anfibios (sobre la diagonal) y reptiles (bajo la diagonal) entre rangos altitudinales según el índice de Whittaker. Entre paréntesis se encuentra el número de especies compartidas entre rangos altitudinales.
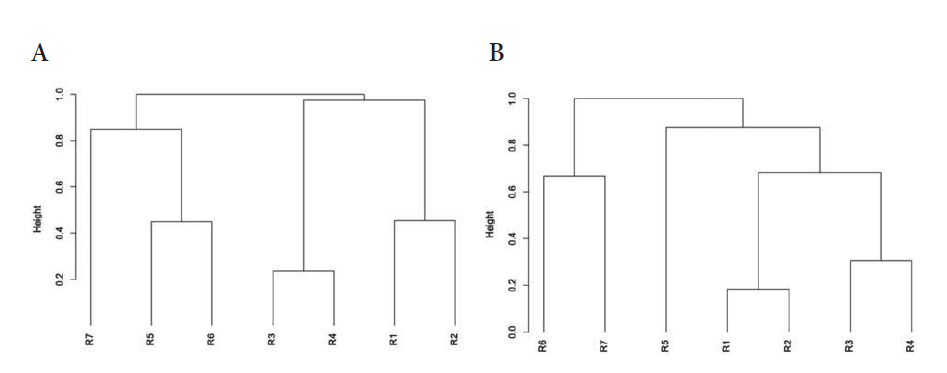
Figura 6.
Análisis de agrupamiento basado en distancias de Jaccard para los intervalos altitudinales en el departamento el Quindío, Colombia. A) Anfibios; B) Reptiles.
Discusión
Generalidades de la herpetofauna en el departamento
Se ha determinado que la región Andina de Colombia alberga cerca de 396 especies de anuros (Bernal y Lynch 2008) y 277 de reptiles (Romero et al. 2008). Entre los departamentos del área, el Quindío recoge un 6 % de la riqueza anfibia (Acosta y Cuentas 2016, http://www.batrachia.com) y 10,2 % de los reptiles conocidos para el país (http://www.reptile–database.org, acceso: 24/07/2016). Además, aunque la región presenta especies típicas de zonas bajas (cerca de 1500 m s.n.m. en el departamento), es un área con principal representación por parte de la fauna Andina, respondiendo a la extensión de la cordillera Central en sus límites políticos (73 %).
Este documento es el primer listado de herpetofauna que se realiza para el departamento. Estudios previos realizados por Cadavid et al. (2005) únicamente registran las especies en una serie de transectos, sin recopilar información exhaustiva sobre la riqueza en el departamento (Vanegas-Guerrero et al. 2014, 2016a). El número de especies de anfibios y reptiles registrados en el Quindío (105 sp.) es congruente con el patrón de riqueza de la vertiente Oriental en la cordillera Central, donde han sido reportadas hasta 145 especies (Llano–M. et al. 2010). A pesar de esto, es evidente una baja riqueza de herpetofauna en comparación con otros departamentos cercanos (e.g. Valle del Cauca, Tolima). Sin embargo, si se considera la relación riqueza/área en las comparaciones con otras entidades administrativas cercanas, se encuentra una mayor riqueza específica para el Quindío. Tolima presenta cerca de siete veces más extensión de la cordillera Central que el Quindío y flanco occidental de la cordillera Central en el Valle del Cauca acoge únicamente 62 sp. (Cardona-B. et al. 2013). En este sentido, la relación riqueza/área resalta la elevada densidad de especies de anfibios y reptiles en el departamento. Sin embargo, factores socioeconómicos (e.g. siembra por monocultivos, aplicación de agentes químicos, Guo et al. 1999) o relacionados con la heterogeneidad espacial (e.g. número de biomas presentes en el área), podrían ser explicaciones plausibles a la baja diversidad alfa.
Distribución altitudinal de la herpetofauna
El patrón general de la diversidad con relación al gradiente altitudinal del Quindío se observa como una disminución del número de especies con respecto al incremento en la altura. Esta tendencia ha sido ampliamente registrada en múltiples ensambles y es explicada a través de las respuestas eco-fisiológicas de ambos grupos ante las variaciones ambientales en los gradientes altitudinales (Kattan 1987, Duellman 1988, Fauth et al. 1989, Navas 1999, Cadavid et al. 2005, Bernal y Lynch 2008, García y Cárdenas 2010). Entre los factores que determinan la abundancia y riqueza de especies han sido propuesta (i) la estrategia reproductiva en anfibios (Kattan 1987, Peñuela et al. 2011), (ii) la capacidad de regulación térmica por efectos comportamentales en reptiles (Watling et al. 2011) además de (iii) la especialización motora, neural y metabólica en ambos grupos (Navas 1999).
alta
Los reptiles estuvieron representados en todos los rangos altitudinales por dipsadidos, gimnophthalmidos y especialmente dactyloides. Contrario a los anfibios, este grupo presenta un patrón continuo de pérdida de riqueza con el incremento de la altura. Es decir, una tendencia de zona baja-media y otra de zona alta. La zona baja-media, acotada entre los 800 y 3000 m s.n.m. (R1-R5), presenta el 98 % de las especies de este grupo. Las serpientes presentan riqueza similar en estos intervalos contrario a los saurios que incrementan con respecto altitud (Cardona–B. et al. 2013). La zona alta (superior a 3000 m s.n.m., R6–R7) presenta pocos representantes (tres especies en tres familias), consecuencia de las limitaciones térmicas para el mantenimiento de las temperaturas corporales para sus actividades (Navas 1999). Algunas especies de lagartos han podido establecerse en estas alturas debido a su capacidad de termorregulación y la relación superficie/volumen (Navas 1999, Zug et al. 2001). Las formas fosoriales de serpientes, como Atractus crassicaudatus, y algunos gimnopthalmidos pueden encontrarse en estas alturas, facultados por las condiciones climáticas (Wiens y Slingluff 2001). Además, la tolerancia térmica y la tendencia euritermica de especies como A. heterodermus pueden influenciar la presencia en estas zonas (presentando al menos un rango de 12°C, Miyata 1983), lo que podría ser una adaptación a estos ambientes junto con la termorregulación comportamental.
Conservación de la herpetofauna
El 40 % de la herpetofauna (29 especies de anfibios y 12 reptiles) del departamento del Quindío se encuentra en la categoría de Preocupación Menor (Least Concern (LC); IUCN 2016). El 37 % aún no ha sido evaluada (39 reptiles) y el 13 % están amenazadas (13 anfibios y un reptil). Las especies amenazadas se encuentran en las categorías: En Peligro Crítico (una especie), En Peligro (5 especies) y Vulnerable (9 especies). En particular estas especies se concentran en la EEL III (11 especies) y I (7 especies). Cabe resaltar que, dentro de los límites políticos del departamento no se registró ninguna especie endémica.
En el listado se incluyen registros de especies introducidas como Hemidactylus sp. y Lepidodactylus lugubris, dado a su amplia distribución y adaptabi-lidad a sitios urbanos (Urbina y Castro 2010, Caicedo–Portilla y Dulcey–Cala 2011, Daza et al. 2012). Existen también algunas especies que, a pesar de ser nativas para Colombia, no presentan una distribución natural en el Quindío (e.g. Kinosternon leucostomum y Trachemys callirostris; Páez et al. 2012) y son por lo tanto consideradas como introducidas (Arroyave–Bermudez et al. 2014).
Conclusiones
Es claro que, aunque la extensión del departamento no es comparable con otras entidades administrativas cercanas, la riqueza de especies e identidad ecológica de los taxones encontrados en el Quindío resalta la importancia de este departamento como reservorio de diversidad. Además se resalta la importancia de las ecorregiones EEL III y I para la inclusión de planes de manejo que ayuden a conservar las especies con algún grado de amenaza. Entre estas, una buena cantidad de registros se encuentran en la EEL I, un área que según Orozco-Serna, corresponde a zonas de cultivos de café. Aunque este artículo pretende solo servir como recopilación de la información publicada, futuros trabajos podrían ser dirigidos a discutir las características de los ecosistemas presentes en éstas ecorregiones que pueden estar determinando la presencia o abundancia de registros en algunas zonas. La información del estado de conservación en el departamento para estos grupos vertebrados aún no es bien conocida, lo que recalca la importancia de promover la publicación y recopilación de conocimiento para su posterior evaluación, y así abordar aspectos relevantes para su conservación. Por último, se espera que este trabajo sea base para futuros estudios que enriquezcan la información de las especies presentes en esta área y funcione además como guía para la demarcación de áreas con importancia crítica de conservación para el Quindío.
Agradecimientos
Los autores agradecen a las iniciativas de acceso libre a las diferentes fuentes de consulta sobre biodiversidad por permitir la recopilación de registros. Agradecen también la lectura crítica de tres revisores anónimos.
Referencias
Acosta-Galvis, A. R. 2000. Ranas, salamandras y caecilias (Tetrapoda: Amphibia) de Colombia. Biota Colombiana 1 (3): 289-319.
Acosta-Galvis, A. R. 2007. Taxonomía y evaluación de la homología de los caracteres para las salamandras del género Bolitoglossa (Caudata: Plethodontidae) de Colombia (Msc Dissertation). Instituto de Ciencias Naturales, Universidad Nacional de Colombia, Bogotá, Colombia. 334 pp.
Amézquita, A., R. Márquez, R. Medina, D. Mejía-Vargas, T. R. Kahn, G. Suarez y L. Mazariegos. 2013. A New species of andean poison frog, Andinobates (Anura: Dendrobatidae) from the Northwestern Andes of Colombia. Zootaxa 1620: 163-178.
Antonelli, A., J. A. Nylander, C. Persson e I. Sanmartín. 2009. Tracing the impact of the Andean uplift on Neotropical plant evolution. Proceedings of the National Academy of Sciences 106 (24): 9749-9754.
Ardila, M. C. y A. R. Acosta. 2000. Anfibios. Pp: 617–628. En: Rangel-Ch, J. O. (Ed.). La Región de vida paramuna. Colombia diversidad biótica III. Universidad Nacional De Colombia, Instituto De Ciencias Naturales, Bogotá.
Arias–Monsalve, H. F. y J. A. Rojas-Morales. 2013. Short comunication Urotheca decipiens (Serpentes: Colubridae): First Record for the department of Risaralda, Colombia. Boletín Científico, Centro de Museos. Museo de Historia Natural 17(1): 144 -146.
Arroyave-Bermúdez, F. J., O. Y. Romero–Goyeneche, M. A. Bonilla–Gómez y R. G. Hurtado–Heredia. 2014. Tráfico ilegal de tortugas continentales (Testudinata) en Colombia: una aproximación desde el análisis de redes. Acta Biológica Colombiana 19 (3): 381-392.
Ayala, S. 1986. Saurios de Colombia: lista actualizada y distribución de ejemplares colombianos en los museos. Caldasia 15 (71-75): 555-576.
Bernal, M. H. y J. D. Lynch. 2008. Review and analysis of altitudinal distribution of the andean anurans in Colombia. Zootaxa 1826: 1-25
Bionda, C., N. Gari, E. Luque, N. Salas, R. Lajmanovich y A. Martino. 2012. Ecología Trófica en larvas de Rhinella Arenarum (Anura: Bufonidae) en agroecosistemas y sus posibles implicaciones para la conservación. Revista de biología tropical 60 (2): 771-779.
Böhm M, B. Collen, J. E. M. Baillie, P. Bowles, J. Chanson, N. Cox, G. Hammerson, M. Hoffmann, et al. 2013. The conservation status of the world’s reptiles. Biological conservation 157: 372-385. http://www.sciencedirect.com/science/article/pii/S0006320712003357
Bolívar-García, W., A. Giraldo y J. Méndez-Narváez. 2011. Amphibia, Anura, Strabomantidae, Pristimantis Palmeri Boulenger 1912: Distribution extension for the Central Cordillera, Colombia. Check List: Journal of Species Lists and Distribution 7: 9-10.
Brame, A. H. y D. B. Wake. 1963. The salamanders of South America. Los Angeles County Museum Contributions in Science 69: 1-73.
Brown, J. L., E. Twomey, A. Amézquita, M. B. D. Souza, J. P. Caldwell, S. Lötters, R. Von May, P. R. Melo–Sampaio, D. Mejía-Vargas, P. E. Perez-Peña, M. Pepper, E. H. Poelman, M. Sanchez-Rodriguez y K. Summers. 2011. A taxonomic revision of the neotropical poison frog genus Ranitomeya (Amphibia: Dendrobatidae). Zootaxa 3083: 1-120.
Bustos-Gómez, F. y A. Lopera 2003. Preferencia por cebo de los escarabajos coprófagos (Coleoptera: Scarabaeidae: Scarabaeinae) de un remanente de bosque seco tropical al norte del Tolima (Colombia). Pp: 59-65. En: Onore, G., P. Reyes-Castillo y M. Zunino (Eds). Escarabeidos de Latinoamérica: estado del conocimiento. Monografías Tercer Milenio Zaragoza, España.
Cadavid, J., C. Román-Valencia y A. Gómez. 2005. Composición y estructura de anfibios anuros en un transecto altitudinal de los andes centrales de Colombia. Revista del Museo Argentino de Ciencias Naturales 7 (2): 103-118.
Cadena, C. D., K. H. Kozak, J. P. Gómez, J. L. Parra, C. M. McCain, R. C. Bowie, A. C. Carnaval, C. Moritz, C. Rahbek, T. E. Roberts, N. J. Sanders, C. J. Schneider, J. VanDerWal, K. Zamudio y C. H. Graham. 2012. Latitude, elevational climatic zonation and speciation in New World vertebrates. Proceedings of the Royal Society of London B: Biological Sciences 279 (1726): 194-201.
Caicedo-Portilla, R. y C. L. Dulcey-Cala. 2011. Distribución del gecko introducido Hemidactylus frenatus (Dumeril y Bribon 1836) (Squamata: Gekkonidae) en Colombia. Biota Colombiana 12 (2): 45-56.
Calderón-Espinosa, M. L., A. M. Ortega-León y J. G. Zamora-Abrego. 2013. Intraspecific variation in body size and shape in an andean highland anole species, Anolis ventrimaculatus (Squamata: Dactyloidae). Revista de Biología Tropical 61 (1): 255-262.
Cardona-B., V. E., R. A. Viáfara-Vega, A. Valencia-Zuleta, A. Echeverry-Bocanegra, O. D. Hernández-Córdoba, A. F. Jaramillo-Martínez, R. Galvis-Cruz, J. A. Gutiérrez y F. Castro-Herrera. 2013. Diversidad de la herpetofauna en el valle del cauca (Colombia): un enfoque basado en la distribución por ecorregiones, altura y zonas de vida. Biota Colombiana 14 (2): 156-233.
Cisneros-Heredia, D. F. y R. W. Mcdiarmid. 2007. Revision of the characters of Centrolenidae (Amphibia: Anura: Athesphatanura), with comments on its taxonomy and the description of new taxa of glassfrogs. Zootaxa 1572: 1 – 82.
Corrales, E. 2002. Transformaciones socioeconómicas y situación de la biodiversidad en los andes colombianos desde el periodo prehispánico. Cuaderno de desarrollo rural 49: 85–163.
Cortés-Gómez, A. M., F. Castro-Herrera y N. J. Urbina-Cardona. 2013. Small changes in vegetation structure create great changes in amphibian ensembles in the Colombian Pacific rainforest. Tropical Conservation Science 6 (6): 749-769.
Daza, J. D, S. L. Travers y A.M. Bauer. 2012. New records of the mourning gecko Lepidodactylus lugubris (Duméril y Bibron 1836) (Squamata: Gekkonidae) from Colombia. Check List 8 (1): 164-167
Di Tada, I. E., M. V. Zabattieri, M. E. Bridarolli, N. E. Salas y A. L. Martino. 1996. Anfibios anuros de la provincia de Córdoba. Pp: 191-215. En: Di Tada, I. E. y E. H. Bucher (Eds). Biodiversidad de la provincia de Córdoba. Universidad Nacional de Río Cuarto. Río Cuarto, Argentina.
Driscoll, D. A. 2004. Extinction and outbreaks accompany fragmentation of a reptile community. Ecological Applications 14 (1): 220-240.
Duellman, W. E. 1979. The herpetofauna of the Andes: Patterns of distribution, origin, differentiation and present communities. Pp: 371-459. En: Duellman, W. E. (Ed.). The South American herpetofauna: its origin, evolution and dispersal. Museum of natural history monograph 7. The University Of Kansas.
Duellman, W. E. 1988. Patterns of species diversity in anuran amphibians in the american tropics. Annual of Missouri Botanical Garden 75: 79-104.
Duellman, W. E. y L. Trueb. 1994. Biology of Amphibians. The John Hopkins University Press Ltd. London. 670 pp.
Etter, A. y W. van Wyngaarden. 2000. Patterns of landscape transformation in Colombia, with emphasis in the Andean region. AMBIO: A Journal of the Human Environment 29 (7): 432-439.
Etter, A., C. McAlpine y H. Possingham. 2008. Historical patterns and drivers of landscape change in Colombia since 1500: a regionalized spatial approach. Annals of the Association of American Geographers 98 (1): 2-23.
Etter, A., P. Amaya y P. A. Arévalo. 2016. Bosques, sabanas y páramos. Pp: 27. En: Gómez, M. F., L. A. Moreno, G. Andrade y C. Rueda (Eds). Biodiversidad 2015. Estado y tendencias de la biodiversidad continental de Colombia. Instituto de Investigación de Recursos Biológicos Alexander von Humboldt. Bogotá, D. C.
Fauth, J. E., B. I. Crother y J. B. Slowinski. 1989. Elevational patterns of species richness, evenness, and abundance of the Costa Rican leaf litter herpetofauna. Biotropica 21 (2): 178-185.
García, J. C. y H. Cárdenas. 2010. Efecto de Rapoport en las ranas Terrarana de América continental: gradientes de riqueza latitudinal y altitudinal. Revista Novedades Colombianas 10 (1): 1-16.
Gómez-Hoyos, D. A., O. H. Marin-Gomez y J. Vanegas-Guerrero. 2012. Unusual amplexus in Dendropsophus columbianus (Anura: Hylidae). Herpetology notes 5: 497-498
Graham, C. H., S. R. Ron, J. C. Santos, C. J. Schneider y C. Moritz. 2004. Integrating phylogenetics and environmental niche models to explore speciation mechanisms in dendrobatids frogs. Evolution 58 (8): 1781-1793.
Grant, T. 2007. A new, toxic species of Colostethus (Anura: Dendrobatidae: Colostethinae) from the cordillera Central of Colombia. Zootaxa 1555: 39-51.
Guo, H., H. Li y Z. Dao. 1999. Dynamism of socio-economy and biodiversity interaction-a case from gaoligong mountains. Acta Botanica Yunnanica 22 (S1): 42-51.
Hernández-Córdoba, O. D., F. Castro-Herrera y M. Paez-Melo. 2013. Bioacumulación de mercurio en larvas de anuros en la zona afectada por la minería de oro en el río Dagua, Buenaventura, Valle Del Cauca, Colombia. Acta Biológica Colombiana 18 (2): 341-348.
Herzog, S. K., R. Martínez, P. M. Jorgensen y H. Tiessen. 2012. Cambio climático y biodiversidad en los andes tropicales. Instituto interamericano para la investigación del cambio global (Iai), Sao José Dos Campos y comité científico sobre problemas del medio ambiente (Scope), Paris. 426 pp.
Heyer, W. R. 1994. Variation within the Leptodactylus Podicipinus-Wagneri Complex of Frogs (Amphibia: Leptodactylidae). Smithsonian Contributions to Zoology: 1–124.
Holdridge, L. R. 1967. Life Zone Ecology. Tropical Science Center, San Jose, Costa Rica. 206 pp.
Hoorn, C., F. P. Wesselingh, H. Ter Steege, M. A. Bermudez, A. Mora, J. Sevink, I. Sanmartín, A. Sanchez-Meseguer, C- L. Anderson, J. P. Figueiredo, C. Jaramillo, D. Riff, F. R. Negri, H. Hooghiemstra, J. Lundberg, T. Stadler, T. Särkinen y A. Antonelli. 2010. Amazonia through time: Andean uplift, climate change, landscape evolution, and biodiversity. Science 330 (6006): 927-931.
IUCN. 2016. The IUCN red list of threatened species. Version 2015.2. . Downloaded On 27 July 2016.
Kattan, G. H. 1987. Patrones de composición taxonómica y de modos reproductivos en comunidades de ranas en el Valle Del Cauca. Cespedesia 15-16 (53, 54, 55, 56): 75-83.
Kattan, G. 2005. Arlequín quimbaya, Atelopus Quimbaya. Pp: 105. En: Rueda Almonacid J.V., J.V. Rodríguez Mahecha, E. La Marca, S.Lötters, T. Kahn y A. Angulo (Eds.) Ranas arlequines. Conservación internacional series libretas de campo. Conservación Internacional Colombia, Bogotá, Colombia.
Kattan, G. H., P. Franco, V. Rojas y G. Morales. 2004. Biological Diversification in a complex region: a spatial analysis of faunistic diversity and biogeography of the Andes of Colombia. Journal of Biogeography 31: 1829-1839.
La Sorte, F. A. y W. Jetz. 2010. Avian distributions under climate change: Towards improved projections. Journal of Experimental Biology 213: 862-869.
Llano-M., J., A. M. Cortés-G y F. Castro-H. 2010. Lista de anfibios y reptiles del departamento del Tolima. Biota Colombiana 11 (1–2): 89–106.
Lomolino, M. V. 2001. Elevation gradients of species-density: Historical and prospective views. Global ecology and biogeography 10 (1): 3-13.
Lynch, J. D. 1986, New species of Eleutherodactylus of Colombia (Amphibia: Leptodactylidae) Ii: four species from the cloud forest of the western cordilleras. Caldasia 15: 629 – 647.
Lynch, J. D. 1997. Intrageneric relationships of mainland Eleutherodactylus II. Review of the Eleutherodactylus sulcatus group. Revista de la Academia Colombiana de Ciencias Exactas, Físicas y Naturales 2: 353-372.
Lynch, J. D. 1991. New diminutive Eleutherodactylus from the cordillera central of Colombia (Amphibia: Leptodactylidae). Journal of Herpetology 25 (3): 344-352.
Lynch, J. D. 1992. Distribution and variation in a colombian frog Eleutherodactyluserythropleura (Amphibia: Leptodactylidae). Studies on neotropical fauna and environment 27 (4): 211-226.
Lynch, J. D. 1999. Ranas pequeñas, la geometría de evolución y la especiación en los andes colombianos. Revista de la Academia Colombiana de Ciencias Exactas, Físicas y Naturales 23 (86): 143-159.
Lynch, J. D. 2000 “1999”. Una aproximación a las culebras ciegas de Colombia (Amphibia: Gymnophiona). Revista de la Academia Colombiana de Ciencias Exactas, Físicas y Naturales 23: 331.
Lynch, J. D. 2009. Snakes of the genus Oxyrhopus (Colubridae: Squamata) in Colombia: taxonomy and geographic variation. Papeis Avulsos de Zoologia 49 (25): 319-337.
Lynch, J. D., P. M. Ruiz-Carranza y J. V. Rueda-Almonacid. 1983. Notes on the distribution and reproductive biology of Centrolene geckoideum Jimenez de la Espada In Colombia And Ecuador (Amphibia: Centrolenidae). Studies on neotropical fauna and environment 18: 239-243.
Lynch, J. D., P. M. Ruiz-Carranza y M. C. Ardila-Robayo. 1994. The identities of the Colombian frogs confused with Eleutherodactylus latidiscus (Boulenger) (Amphibia: Anura: Leptodactylidae). Occasional papers of the Museum of Natural History, University of Kansas 170: 1–42.
Lynch, J. D., P. M. Ruiz-Carranza y M. C. Ardila-Robayo. 1996. Three new species of Eleutherodactylus (Amphibia: Leptodactylidae) from high elevations of the Cordillera Central of Colombia. Caldasia 18: 329-342.
Lynch, J. D. y P. M. Ruíz-Carranza y M. C. Ardila-Robayo. 1997. Biogeographic patterns of Colombian frogs and toads. Revista de la Academia Colombiana de Ciencias Exactas, Físicas y Naturales 21 (80): 237-248.
Lynch, J. D. y A. M. Suárez-Mayorga. 2002. Análisis biogeográfico de los anfibios paramunos. Caldasia 24(2): 471-480.
Madriñán, S., A. J. Cortés y J. E. Richardson. 2013. Páramo is the world’s fastest evolving and coolest biodiversity hotspot. Frontiers in genetics 4: 192.
Marin, O. H y D. G. Hoyos. 2011. Estado actual de Ranitomeya bombetes (Anura: Dendrobatidae): plan de manejo y conservación para las poblaciones de Ranitomeya bombetes (Myers y Daly 1980) en Quindío, Colombia. Editorial Académica Española. 108 pp.
McCain, C. M. 2010. Global analysis of reptile elevational diversity. Global Ecology and Biogeography 19 (4): 541-553.
Medem, F. 1977. Contribución al conocimiento sobre la taxonomía, distribución geográfica y ecología de la tortuga “bache” (Chelydra serpentina acutirostris). Caldasia 12 (56): 41-101.
Mendelson J. R, K. R. Lips, R. W. Gagliardo, G. B. Rabb, J. P. Collins, J. E. Diffendorfer, P. Daszak, R. Ibáñez, K. C. Zippel, D. P. Lawson, K. M. Wright, S. N. Stuart, C. Gascon, H. R. da Silva, P. A. Burrowes, R. L. Joglar, E. La Marca, S. Lötters, L. H. du Preez, C. Weldon, A. Hyatt, J. V. Rodriguez-Mahecha, S. Hunt, H. Robertson, B. Lock, C. J. Raxworthy, D. R. Frost, R. C. Lacy, R. A. Alford, J. A. Campbell, G. Parra-Olea, F. Bolaños, J. J. Calvo-Domingo, T. Halliday, J. B. Murphy, M. H. Wake, L. A. Coloma, S. L. Kuzmin, M. S. Price, K. M. Howell, M. Lau, R. Pethiyagoda, M. Boone, M. J. Lannoo, A. R. Blaustein, A. Dobson, R. A. Griffiths, M. L. Crump, D. B. Wake y E. D. Brodie Jr. 2006. Confronting amphibian declines and extinctions. Science 313: 48.
Mendoza, Á. M., O. E. Ospina, H. Cárdenas-Henao y J. C. García-R. 2015. A likelihood inference of historical biogeography in the world’s most diverse terrestrial vertebrate genus: Diversification of direct-developing frogs (Craugastoridae: Pristimantis) across the Neotropics. Molecular phylogenetics and evolution 85: 50-58.
Miyata, K. 1983. Notes on Phenacosaurus heterodermus in the sabana de Bogotá, Colombia. Journal of Herpetology 17 (1): 102-105.
Mueses-Cisneros, J. J. y M. A. Anganoy-Criollo. 2008. Una nueva especie del grupo Hyloscirtus larinopygion (Amphibia: Anura, Hylidae) del suroccidente de Colombia. Papéis Avulsos de Zoologia 48: 129-138.
Murgueitio, E. 2002. Sistemas de producción ganadera y sus impactos en la transformación de los ecosistemas andinos de Colombia. Pp: 124. En: Memorias del seminario internacional sobre transformación de ecosistemas. Universidad Javeriana, Bogotá, Colombia.
Muñoz‐Ortiz, A., Á. A. Velásquez‐Álvarez, C. E. Guarnizo y A. J. Crawford. 2014. Of peaks and valleys: testing the roles of orogeny and habitat heterogeneity in driving allopatry in mid‐elevation frogs (Aromobatidae: Rheobates) of the northern Andes. Journal of Biogeography 42 (1): 193-205.
Murphy, P. G. y A. E. Lugo. 1986. Ecology of tropical dry forest. Annual review of ecology and systematics 17: 67-88.
Myers, N., R. A. Mittermeier, C. G. Mittermeier, G. A. B. Da Fonseca y J. Kent. 2000. Biodiversity hotspots for conservation priorities. Nature 403: 853-858.
Nadkarni, N. M. y N. T. Wheelwrught. 2000. Monteverde: Ecology and conservation of a tropical cloud forest. Oxford University Press. 608 pp.
Navas, C. A. 1999. Biodiversidad de anfibios y reptiles en el páramo: una visión eco-fisiológica. Revista de la Academia Colombiana de Ciencias Exactas, Físicas y Naturales 23 (Suplemento Especial): 465-474.
Navas, C. A. 2002. Herpetological diversity along Andean elevational gradients: links with physiological ecology and evolutionary physiology. Comparative biochemistry and physiology Part A: Molecular y Integrative Physiology 133 (3): 469-485.
Navas, C. A., J. M. Carvajalino-Fernández, L. P. Saboya-Acosta, L. A. Rueda-Solano y M. A. Carvajalino-Fernández. 2013. The body temperatura of active amphibians along tropical elevation gradient: patterns of mean and variance and inference from environmental data. Funcional Ecology 2013: 1-7.
Oksanen, J., R. Kindt, P. Legendre, B. O’Hara, M. H. H. Stevens, M. J. Oksanen y M. A. S. S. Suggests. 2007. The vegan package. Vegan: Community Ecology Package. R package version 1.18-4/r1180. http://RForge.R-project.org/projects/vegan/.
Orme, C. D. L, R. Davies, M. Burgess, F. Eigenbrod, N. Pickup, V. A. Olson, A. J. Webster, T. Ding, P. Rasmussen, R. Eidgley, A. Stattersfield, P. Bennett, T. Blackburn, K. Gaston y I. Owens. 2005. Global hotspots of species richness are not congruent with endemism or threat. Nature 436: 1016-1019.
Orozco-Serna, J. C. 2012. Zonificación territorial por ecorregiones estratégicas locales en el departamento del Quindío. Scientia Et Technica 1 (52): 219-224.
Osorio, D. y A. Quintero. 2012. Espadarana prosoblepon Boettger 1892 (Amphibia: Anura): Distribution extension on the western slopes of the Cordillera Central, Colombia. Check List 8 (5): 898-899.
Páez, V. P., M. A. Morales-Betancourt, C. A. Lasso, O. V. Castaño-Mora y B. C. Bock. 2012. V. Biología y conservación de las tortugas continentales de Colombia. Serie editorial recursos hidrobiológicos y pesqueros continentales de Colombia. Instituto de Investigación de Recursos Biológicos Alexander Von Humboldt (Iavh). Bogotá, D. C., Colombia, 528 pp.
Passos, P. y J. C. Arredondo. 2009. Rediscovery and redescription of the andean earth–snake Atractus Wagleri (Reptilia: Serpentes: Colubridae). Zootaxa 1969: 59-68
Passos, P. y J. D. Lynch. 2011. Revision of Atractus (Serpentes: Dipsadidae) from middle and upper magdalena drainage of Colombia. Herpetological Monographs 24: 149-173.
Passos, P., J. C. Arredondo, R. Fernandes y J. D. Lynch. 2009. Three new Atractus (Serpentes: Dipsadidae) from the andes of Colombia. Copeia 2009 (3): 425-436.
Patterson, B. D., P. L. Meserve y B. K. Lang. 1989. Distribution and abundance of small mammals along an elevational transect in temperate rainforests of Chile. Journal Of Mammalogy 70: 67-78.
Peñuela, M., Hernández, O. y F. Castro. 2011. Modos reproductivos de la anurofauna Vallecaucana. Momentos De Ciencia 8 (1): 55-63.
Peters, J. A. y B. Orejas-Miranda. 1970. Catalogue of the neotropical squamata: Part I. Snakes. United States National Museum Bulletin 297: 1 -347.
Pyron, R. A. y J. J. Wiens. 2011. A large-scale phylogeny of Amphibia including over 2800 species, and a revised classification of extant frogs, salamanders, and caecilians. Molecular Phylogenetics and Evolution 61 (2): 543-583.
Quintero-Ángel, A., D. Osorio-Domínguez, F. Vargas-Salinas y C. A. Saavedra-Rodríguez. 2012. Roadkill rate of snakes in a disturbed landscape of central andes of Colombia. Herpetology Notes 5: 99-105.
Restrepo, C. A. y A. Botero-Botero. 2012. Ecología trófica de la nutria neotropical Lontra longicaudis (carnívora, mustelidae) en el río La Vieja, Alto Cauca, Colombia. Boletín Científico Museo de Historia Natural 16 (1): 207-214.
Rickart, E. A., L. R. Héaney y C. B. Utzurrum. 1991. Distribution and ecology of small mammals along an elevational transect in southeastern Luzun, Philippines. Journal of Mammalogy 72: 458-469.
Rincón, A. y N. R. Bernal. 2007. Factores antrópicos asociados e interrelaciones con el estado de los ecosistemas andinos. Pp: 174-179. En: Armenteras, D. y N. Rodríguez (Eds.). 2007. Monitoreo de los ecosistemas andinos 1985–2005: síntesis y perspectivas. Instituto de Investigaciones Alexander Von Humbolt. Bogotá, D.C. Colombia.
Roa-Trujillo, S. H. y P. M. Ruíz-Carranza. 1991. Una nueva especie de Eleutherodactylus (Amphibia: Leptodactylidae) de la cordillera central de Colombia. Caldasia 16 (78): 343-348.
Rojas-Morales, J. A. y S. Escobar-Lasso. 2010. Defensive behavior of Dipsas sanctijoannis (Serpentes: Dipsadidae). Phyllomedusa 9(2): 147-150.
Rojas-Morales, J. A., S. Escobar-Lasso y P. D. A. Gutiérrez-Cárdenas. 2011. Contribución al conocimiento de los anfibios de la región centro–sur de caldas: primeros registros de ranas de cristal (Anura: Centrolenidae) para el municipio de Manizales, Colombia. Boletín Científico Museo De Historia Natural Universidad De Caldas 15: 75-83.
Romero, J. H., C. C. Vidal y J. D. Lynch. 2008. Estudio preliminar de la fauna amphibia en el cerro murrucucú, parque natural nacional paramillo y zona amortiguadora, Tierralta, Córdoba, Colombia. Caldasia 30 (1): 209-229.
Ruiz-C, P. M. y M. Osorno-M. 1994. Tres nuevas especies de Atelopus Dumeril y Bibron 1841 (Amphibia: Bufonidae) de la cordillera central de Colombia. Revista de la Academia Colombiana de Ciencias Exactas, Físicas y Naturales 19 (72): 165-179.
Ruiz-Carranza, P. M. y J. D. Lynch. 1995. Ranas centrolenidae de Colombia VIII. Cuatro nuevas especies de Centrolene de la cordillera central. Lozania 65: 1-16.
Ruiz-Carranza, P. M., J. I. Hernández-Camacho y M. C. Ardila-Robayo. 1986. Una nueva especie colombiana del género Centrolene Jimenez de la Espada 1872 (Amphibia: Anura) y redefinición del género. Caldasia 15: 431-444.
Ruiz-Carranza, P. M. y J. D. Lynch. 1991. Ranas centrolenidae de Colombia III Nuevas Especies de Cochranella del grupo granulosa. Lozania 59: 1-18.
Ruiz-Carranza, P. M., M. C. Ardila-Robayo y J. D. Lynch. 1986. Lista actualizada de la fauna de amphibia de Colombia. Revista de la Academia Colombiana de Ciencias Exactas, Físicas Y Naturales 20 (77): 365–415.
Rull, V. 2011. Neotropical biodiversity: timing and potential drivers. Trends in ecology & evolution 26 (10): 508-513.
Sanabria, E., L. Quiroga y J. C. Acosta. 2007. Hábitos alimentarios de infantiles de Pleurodema nebulosum (Anura: Leptodactylidae), en Matagusanos, San Juan, Argentina. Revista Peruana de Biología 14 (2): 295-296.
Sánchez-C, H., O. Castaño-M y G. Cárdenas-A. 1995. Diversidad de los reptiles en Colombia. P. 277–326. En: Rangel-Ch, J. O. (Ed.), Colombia Diversidad Biótica I. Instituto De Ciencias Naturales–Universidad Nacional De Colombia–Inderena, Bogotá.
Sanchez-Pacheco, S. J. 2010. Lectotype designation and redescription of the gymnophthalmid lizard Riama Columbiana (Andersson 1914) with notes on the type locality. Papéis Avulsos De Zoologia 50(2): 31-41
Sánchez-Pacheco, S. J., D. A. Kizirian y P. M. Sales-Nunes. 2011. A new species of Riama from Ecuador previously referred to as Riama hyposticta (Boulenger 1902) (Squamata: Gymnophthalmidae). American Museum Novitates 3719: 1–15.
Santos, N. D., D. P. Costa, L. S. Kinoshita y G. L. Shepherd. 2011. Aspectos brioflorísticos e fitogeográficos de duas formações costeiras de floresta atlântica Da Serra do Mar, Ubatuba/Sp, Brasil. Biota Neotropica 11 (2): 425-438.
Stuart, S. N., M. Hoffmann, J. Chanson, N. Cox, R. Berridge, P. Ramani y B. Young. 2008. Threatened amphibians of the world [Barcelona, Spain; International Union for the Conservation Of Nature, Gland. Switzerland. Conservation International, Arlington, Virginia, U.S.A.]: Lynx Editions. 758 pp.
Theisinger, O. y M. C. Ratianarivo. 2015. Patterns of reptile diversity loss in response to degradation in the spiny forest of southern Madagascar. Herpetological Conservation and Biology 10 (1): 273-283.
Urbina-Cardona, J. N. y F. Castro. 2010. Distribución actual y futura de anfibios y reptiles con potencial invasor en Colombia: Una aproximación usando modelos de nicho ecológico. Pp: 65-71. En: Varela-Ramírez, A. (Ed.) Biodiversidad y Cambio Climático. Ideam-Proyecto inap componente b alta montaña. Pontificia Universidad Javeriana, Bogota, Colombia.
Valencia-Zuleta, A., A. F. Jaramillo-Martinez, A. Echeverry-Bocanegra, R. Viáfara-Vega, O. Hernández-Córdoba, V. E. Cardona-Botero, J. Gutiérrez-Zúñiga y F. Castro-Herrera. 2014. Conservation Status of the herpetofauna, protected areas, and current problems in Valle del Cauca, Colombia. Amphibian & Reptile Conservation 8 (2) [Special Section]: 1-18.
Vanegas-Guerrero, J. 2015. Reptilia, Sauria, Gymnophthalmidae, Anadia rhombifera (Günther 1859): Distribution extension and first records from Quindío department, Colombia. Check List 11 (1): 1512.
Vanegas-Guerrero, J., J. C. Mantilla-Castaño y P. Passos. 2014. Atractus Titanicus Passos, Arredondo, Fernandes y Lynch, 2009 (Serpentes: Dipsadidae): Filling Gaps in Its geographical distribution. Check List 10 (3): 672-673.
Vanegas-Guerrero, J., G. A. Gonzales-Durán y S. Escobar-Lasso. 2016a. Distribution, diet, and vocalizations of the endangered colombian toad Osornophryne percrassa (Anura, Bufonidae). Herpetological Conservation and Biology 11 (1): 90-100.
Vanegas-Guerrero, J., C. Fernández, W. Buitrago-González y F. Vargas-Salinas. 2016b. Urban Remnant Forests: Are They Important for Herpetofaunal Conservation in the Central Andes of Colombia?. Herpetological Review 47 (2): 180-185.
Velasco, J. A., P. D. Gutiérrez-Cárdenas y A. Quintero-Ángel. 2010. A New Species of Anolis of the Aequatorialis Group (Squamata: Iguania) from the Central Andes of Colombia. Herpetological Journal 20: 231–236
Wathen, S., J. H. Thorne, A. Holguin y M. W. Schwartz. 2014. Estimating the Spatial and temporal distribution of species richness within Sequoia and Kings Canyon National Parks. PloS one 9 (12): e112465.
Watling, J. I., C. R. Hickman y J. L. Orrock. 2011. Invasive shrub alters native forest amphibian communities. Biological Conservation 144 (11): 2597-2601.
Wiens, J. J. 2004. Speciation and ecology revisited: phylogenetic niche conservatism and the origin of species. Evolution 58 (1): 193-197.
Wiens, J. J. y C. H. Graham. 2005. Niche conservatism: integrating evolution, ecology, and conservation biology. Annual review of ecology, evolution, and systematics: 519-539.
Wiens, J. y L. Slingluff. 2001. How Lizards Turn Into Snake: A phylogenetic analysis of body–form Evolution in anguid lizards. Evolution 55 (11): 2303-2318.
Zug, G. R., L. J. Vitt y J. P. Caldwell. 2001. Herpetology. Second Edition. Academic Press. 630 pp.
Anexos
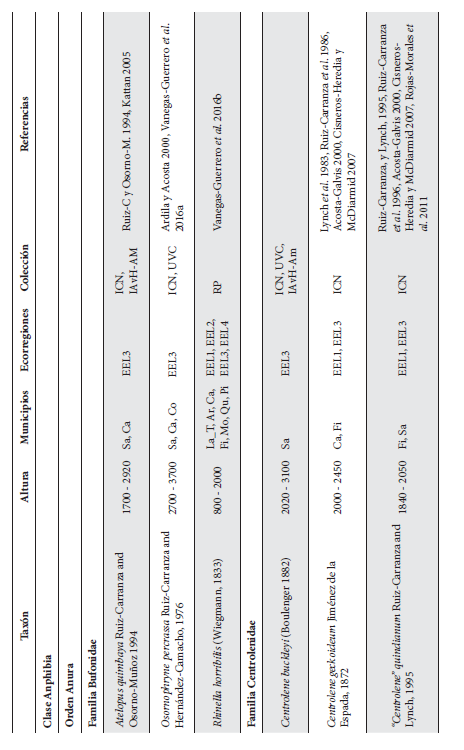
Anexo 1.
Listado taxonómico de la herpetofauna del departamento del Quindío (Colombia). Lista de las especies reportadas con base en los museos (ICN, IAvH-Am, IAvH-R, MHUA-A, UVC, FML, ANSP, CSJ, MCZ, USNM) y referencias bibliográficas que soportan su presencia en área. RP indica registros propios. Las especies con posibles registros se indican como OP. La distribución por áreas naturales fue establecida según Orozco-Serna (2012) separado en cuatro zonas estratégicas a partir de características socio-culturales y biológicas (EEL I - IV). El símbolo (†) denota la especies introducidas para el departamento y el (††) denota introducida y no incluida en los análisis por carencia de información sobre establecimiento de poblaciones.
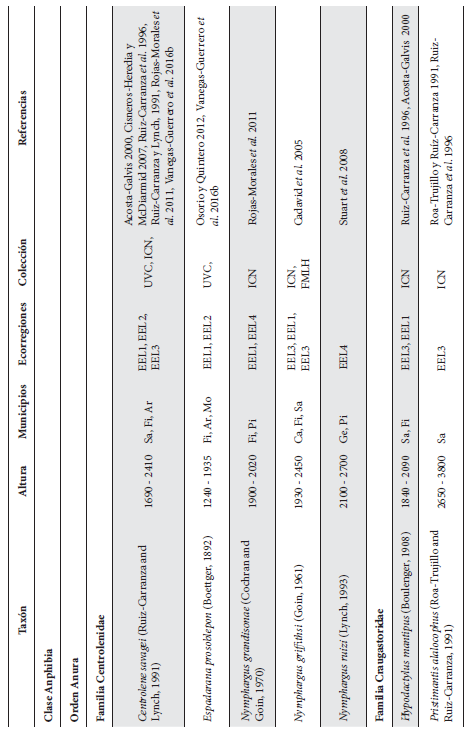
Cont. Anexo 1.
Listado taxonómico de la herpetofauna del departamento del Quindío (Colombia). Lista de las especies reportadas con base en los museos (ICN, IAvH-Am, IAvH-R, MHUA-A, UVC, FML, ANSP, CSJ, MCZ, USNM) y referencias bibliográficas que soportan su presencia en área. RP indica registros propios. Las especies con posibles registros se indican como OP. La distribución por áreas naturales fue establecida según Orozco-Serna (2012) separado en cuatro zonas estratégicas a partir de características socio-culturales y biológicas (EEL I - IV). El símbolo (†) denota la especies introducidas para el departamento y el (††) denota introducida y no incluida en los análisis por carencia de información sobre establecimiento de poblaciones.
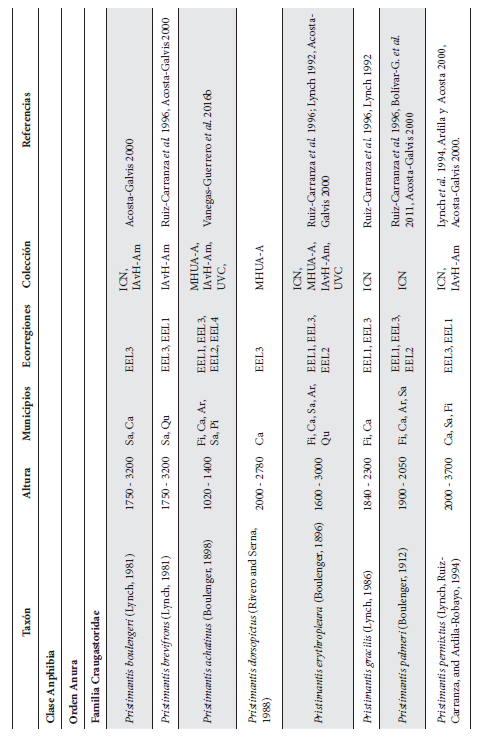
Cont. Anexo 1.
Listado taxonómico de la herpetofauna del departamento del Quindío (Colombia). Lista de las especies reportadas con base en los museos (ICN, IAvH-Am, IAvH-R, MHUA-A, UVC, FML, ANSP, CSJ, MCZ, USNM) y referencias bibliográficas que soportan su presencia en área. RP indica registros propios. Las especies con posibles registros se indican como OP. La distribución por áreas naturales fue establecida según Orozco-Serna (2012) separado en cuatro zonas estratégicas a partir de características socio-culturales y biológicas (EEL I - IV). El símbolo (†) denota la especies introducidas para el departamento y el (††) denota introducida y no incluida en los análisis por carencia de información sobre establecimiento de poblaciones.
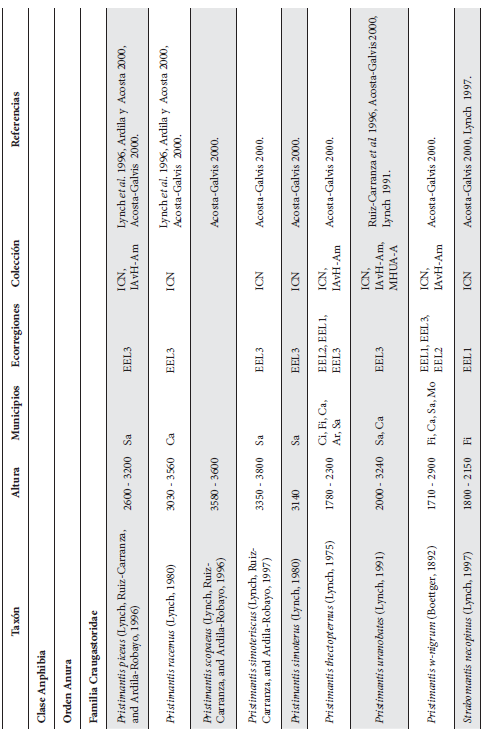
Cont. Anexo 1.
Listado taxonómico de la herpetofauna del departamento del Quindío (Colombia). Lista de las especies reportadas con base en los museos (ICN, IAvH-Am, IAvH-R, MHUA-A, UVC, FML, ANSP, CSJ, MCZ, USNM) y referencias bibliográficas que soportan su presencia en área. RP indica registros propios. Las especies con posibles registros se indican como OP. La distribución por áreas naturales fue establecida según Orozco-Serna (2012) separado en cuatro zonas estratégicas a partir de características socio-culturales y biológicas (EEL I - IV). El símbolo (†) denota la especies introducidas para el departamento y el (††) denota introducida y no incluida en los análisis por carencia de información sobre establecimiento de poblaciones.
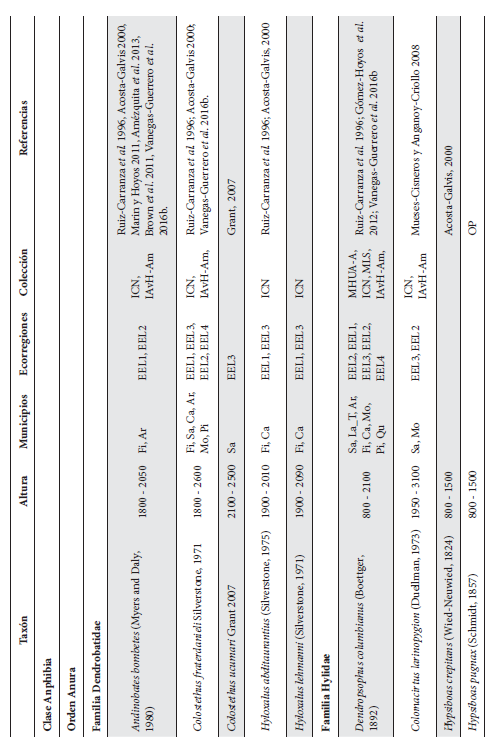
Cont. Anexo 1.
Listado taxonómico de la herpetofauna del departamento del Quindío (Colombia). Lista de las especies reportadas con base en los museos (ICN, IAvH-Am, IAvH-R, MHUA-A, UVC, FML, ANSP, CSJ, MCZ, USNM) y referencias bibliográficas que soportan su presencia en área. RP indica registros propios. Las especies con posibles registros se indican como OP. La distribución por áreas naturales fue establecida según Orozco-Serna (2012) separado en cuatro zonas estratégicas a partir de características socio-culturales y biológicas (EEL I - IV). El símbolo (†) denota la especies introducidas para el departamento y el (††) denota introducida y no incluida en los análisis por carencia de información sobre establecimiento de poblaciones.
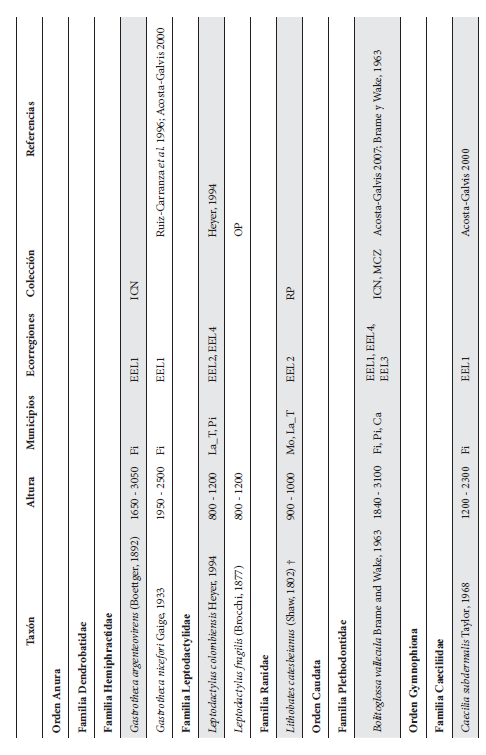
Cont. Anexo 1.
Listado taxonómico de la herpetofauna del departamento del Quindío (Colombia). Lista de las especies reportadas con base en los museos (ICN, IAvH-Am, IAvH-R, MHUA-A, UVC, FML, ANSP, CSJ, MCZ, USNM) y referencias bibliográficas que soportan su presencia en área. RP indica registros propios. Las especies con posibles registros se indican como OP. La distribución por áreas naturales fue establecida según Orozco-Serna (2012) separado en cuatro zonas estratégicas a partir de características socio-culturales y biológicas (EEL I - IV). El símbolo (†) denota la especies introducidas para el departamento y el (††) denota introducida y no incluida en los análisis por carencia de información sobre establecimiento de poblaciones.
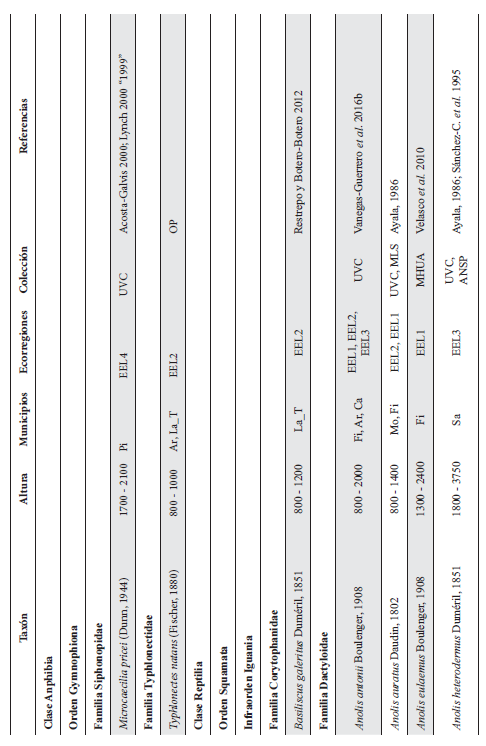
Cont. Anexo 1.
Listado taxonómico de la herpetofauna del departamento del Quindío (Colombia). Lista de las especies reportadas con base en los museos (ICN, IAvH-Am, IAvH-R, MHUA-A, UVC, FML, ANSP, CSJ, MCZ, USNM) y referencias bibliográficas que soportan su presencia en área. RP indica registros propios. Las especies con posibles registros se indican como OP. La distribución por áreas naturales fue establecida según Orozco-Serna (2012) separado en cuatro zonas estratégicas a partir de características socio-culturales y biológicas (EEL I - IV). El símbolo (†) denota la especies introducidas para el departamento y el (††) denota introducida y no incluida en los análisis por carencia de información sobre establecimiento de poblaciones.
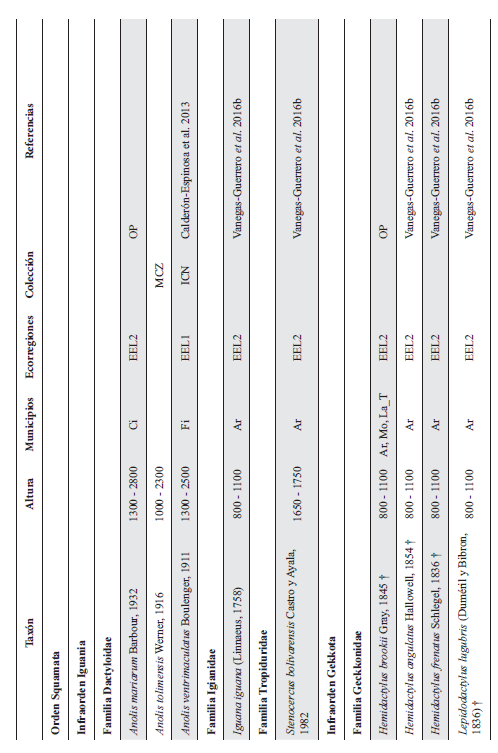
Cont. Anexo 1.
Listado taxonómico de la herpetofauna del departamento del Quindío (Colombia). Lista de las especies reportadas con base en los museos (ICN, IAvH-Am, IAvH-R, MHUA-A, UVC, FML, ANSP, CSJ, MCZ, USNM) y referencias bibliográficas que soportan su presencia en área. RP indica registros propios. Las especies con posibles registros se indican como OP. La distribución por áreas naturales fue establecida según Orozco-Serna (2012) separado en cuatro zonas estratégicas a partir de características socio-culturales y biológicas (EEL I - IV). El símbolo (†) denota la especies introducidas para el departamento y el (††) denota introducida y no incluida en los análisis por carencia de información sobre establecimiento de poblaciones.
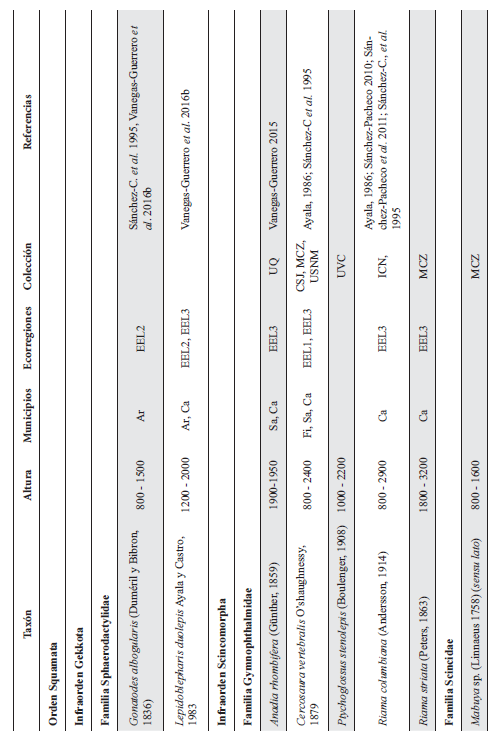
Cont. Anexo 1.
Listado taxonómico de la herpetofauna del departamento del Quindío (Colombia). Lista de las especies reportadas con base en los museos (ICN, IAvH-Am, IAvH-R, MHUA-A, UVC, FML, ANSP, CSJ, MCZ, USNM) y referencias bibliográficas que soportan su presencia en área. RP indica registros propios. Las especies con posibles registros se indican como OP. La distribución por áreas naturales fue establecida según Orozco-Serna (2012) separado en cuatro zonas estratégicas a partir de características socio-culturales y biológicas (EEL I - IV). El símbolo (†) denota la especies introducidas para el departamento y el (††) denota introducida y no incluida en los análisis por carencia de información sobre establecimiento de poblaciones.
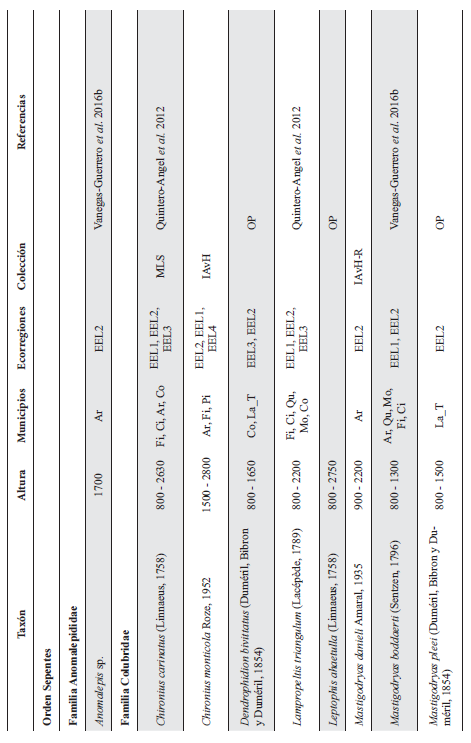
Cont. Anexo 1.
Listado taxonómico de la herpetofauna del departamento del Quindío (Colombia). Lista de las especies reportadas con base en los museos (ICN, IAvH-Am, IAvH-R, MHUA-A, UVC, FML, ANSP, CSJ, MCZ, USNM) y referencias bibliográficas que soportan su presencia en área. RP indica registros propios. Las especies con posibles registros se indican como OP. La distribución por áreas naturales fue establecida según Orozco-Serna (2012) separado en cuatro zonas estratégicas a partir de características socio-culturales y biológicas (EEL I - IV). El símbolo (†) denota la especies introducidas para el departamento y el (††) denota introducida y no incluida en los análisis por carencia de información sobre establecimiento de poblaciones.
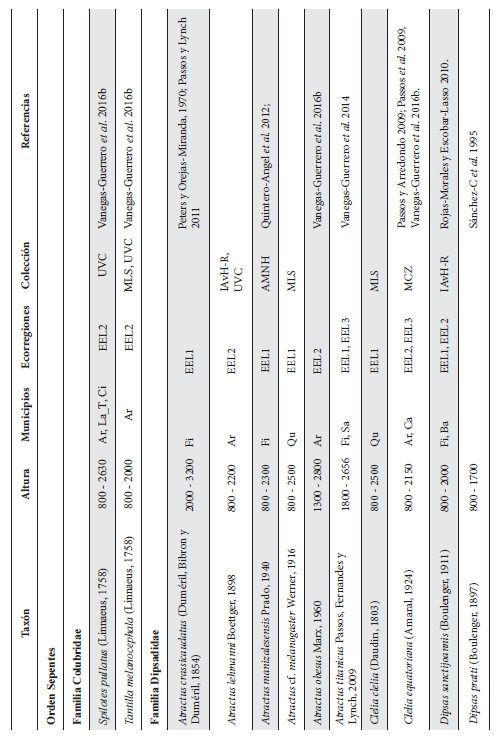
Cont. Anexo 1.
Listado taxonómico de la herpetofauna del departamento del Quindío (Colombia). Lista de las especies reportadas con base en los museos (ICN, IAvH-Am, IAvH-R, MHUA-A, UVC, FML, ANSP, CSJ, MCZ, USNM) y referencias bibliográficas que soportan su presencia en área. RP indica registros propios. Las especies con posibles registros se indican como OP. La distribución por áreas naturales fue establecida según Orozco-Serna (2012) separado en cuatro zonas estratégicas a partir de características socio-culturales y biológicas (EEL I - IV). El símbolo (†) denota la especies introducidas para el departamento y el (††) denota introducida y no incluida en los análisis por carencia de información sobre establecimiento de poblaciones.
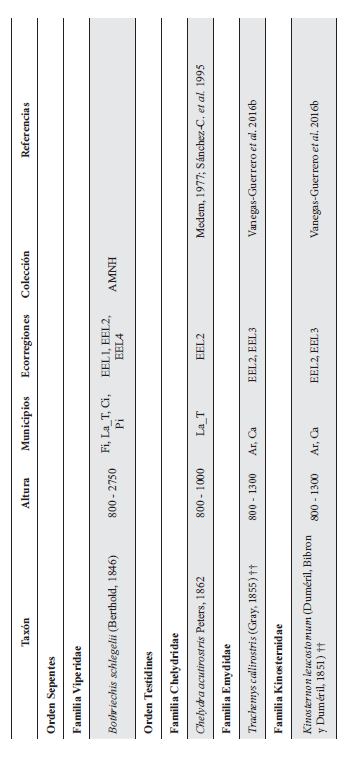
Cont. Anexo 1.
Listado taxonómico de la herpetofauna del departamento del Quindío (Colombia). Lista de las especies reportadas con base en los museos (ICN, IAvH-Am, IAvH-R, MHUA-A, UVC, FML, ANSP, CSJ, MCZ, USNM) y referencias bibliográficas que soportan su presencia en área. RP indica registros propios. Las especies con posibles registros se indican como OP. La distribución por áreas naturales fue establecida según Orozco-Serna (2012) separado en cuatro zonas estratégicas a partir de características socio-culturales y biológicas (EEL I - IV). El símbolo (†) denota la especies introducidas para el departamento y el (††) denota introducida y no incluida en los análisis por carencia de información sobre establecimiento de poblaciones.
Cont. Anexo 1.
Listado taxonómico de la herpetofauna del departamento del Quindío (Colombia). Lista de las especies reportadas con base en los museos (ICN, IAvH-Am, IAvH-R, MHUA-A, UVC, FML, ANSP, CSJ, MCZ, USNM) y referencias bibliográficas que soportan su presencia en área. RP indica registros propios. Las especies con posibles registros se indican como OP. La distribución por áreas naturales fue establecida según Orozco-Serna (2012) separado en cuatro zonas estratégicas a partir de características socio-culturales y biológicas (EEL I - IV). El símbolo (†) denota la especies introducidas para el departamento y el (††) denota introducida y no incluida en los análisis por carencia de información sobre establecimiento de poblaciones.
Notas de autor
Universitat de Barcelona
Universitat de Barcelona
Información adicional
Citación del artículo: Román-Palacios, C., S. Fernández -Garzón, A. Valencia-Zuleta, A. F. Jaramillo-Martínez y R. A. Viáfara-Vega. 2017. Lista anotada de la herpetofauna del departamento del Quindío, Colombia. Biota Colombiana 18 (1): 251-281
