Licófitos y helechos (Lycophyta-monilophyta) de el cañón, sierra de boniato, Santiago de Cuba, Cuba
Licófitos y helechos (Lycophyta-monilophyta) de el cañón, sierra de boniato, Santiago de Cuba, Cuba
Foresta Veracruzana, vol. 19, núm. 2, pp. 9-18, 2017
Recursos Genéticos Forestales
Resumen: Se presenta una lista actualizada de los licófitos y los helechos de la localidad de El Cañón, Sierra de Boniato, Santiago de Cuba, a partir de la revisión de materiales de herbario y de literatura, así como de muestreos en la localidad. Se identificaron 62 taxones infragenéricos, agrupados en 23 géneros para un total de 13 familias; estas representan el 10% de las reportadas para la región Oriental; de las cuales una es endémica. Las familias más representativas dentro del área de estudio resultaron ser: Lomariopsidaceae, Polypodiaceae, Pteridaceae y Thelypteridaceae. No se reportaron especies con categorías de amenaza. Con respecto a la diversidad de especies por formaciones vegetales, las de mayor riqueza estuvieron representadas en los bosques de galería y los bosques semidecíduos mesófilos, con 50 especies compartidas entre ambas, complementando lo reportado para la Sierra Maestra Oriental.
Palabras clave: Sierra de Boniato, Cuba oriental, licófitos, helechos.
Abstract: An inventory of the Lycophytes and ferns of locality El Cañón is presented. This area is located in Boniato Mountain, Santiago de Cuba province. A review of herbarium materials and specialized literature was necessary, as well as of samplings recollected in the field. 62 infrageneric taxa were identified containing 23 genera and 13 families; it represents the 10% of the flora reported for the Western region of Cuba, one of them is endemic. The most representative families were Lomariopsidaceae, Polypodiaceae, Pteridaceae and Thelypteridaceae. None species is threatened. The plant formations with more richness of species were gallery and semi-deciduous forests, with 50 species that shared both; it is reported for the Sierra Maestra Mountains.
Keywords: Sierra de Boniato, East Cuba, lycophytes, ferns.
Introducción
Los licófitos y los helechos forman parte importante de la diversidad vegetal del mundo tropical, caracterizando a veces los tipos de vegetación a los que se asocian. Éstos desempeñan un papel importante en la conservación de los ecosistemas, ya que constituyen un indicador biológico de la salud de los mismos.
El archipiélago cubano posee una riqueza pteridológica de 715 táxones infragenéricos con un endemismo de 15.1%. Estos se distribuyen principalmente en las tres grandes regiones montañosas del territorio (Borhidi, 1991), siendo la Sierra Maestra, la que reporta la mayor diversidad de táxones infragenéricos (611), debido a la gran variedad de ecótopos, los que proporcionan las variadas condiciones geológicas, climáticas y altitudinales (Caluff y Shelton, 2000a;Caluff y Fuentes, 2010).
La Sierra de Boniato forma parte de la Sierra Maestra, e incluida en el distrito fitogeográfico Turquinense (Borhidi y Muñiz, 1986), se extiende unos 24 kms, cerrando la cuenca de Santiago de Cuba por el norte y siendo su mayor altitud de 593 msm (Núñez, 1972). Esta zona ha estado sometida durante mucho tiempo a fuertes presiones antrópicas ocasionadas por la expansión de la población, las actividades agrícolas, la explotación de especies para cultivo como ornamentales y el ecoturismo; lo cual ha provocado cambios en la composición de la flora de licófitos y helechos que se establecen en sus diferentes ecosistemas.
Para este estudio se seleccionó la localidad de El Cañón, a pesar de ser un área antropizada, donde predomina la vegetación secundaria, porque aún conserva valores nativos de la flora fanerogámica y pteridológica especialmente en sus pequeños relictos de bosques de galería y bosques semideciduos mesófilos.
El Cañón se sitúa en el valle intramontano de la vertiente norte de la Sierra de Boniato, entre los 75° 48' 20.13" y 75° 47' 39.07" Oeste y 20° 6' 7.01" y 20° 6'29.35" Norte; se halla a unos 20 kms de la ciudad de Santiago de Cuba y posee una altitud entre 260 y 350 msnm. La región se encuentra surcada por el río Boniatico y varios afluentes (Renda et al., 1980).
Este trabajo tiene como objetivo determinar el estado actual de la flora pteridológica, en cuatro formaciones vegetales de la localidad de El Cañón, así como algunos aspectos de su ecología. Se destacan las especies endémicas nativas, las naturalizadas y las introducidas. Este estudio completará otros trabajos realizados en la Sierra Maestra y de Cuba Oriental a fin de desarrollar proyectos comunitarios con vista a difundir los valores de la pteridoflora de la localidad y contribuir a su protección y conservación. Servirá para promover el ecoturismo sostenible en el área como un valor agregado del centro turístico Puerto de Boniato.
Material y métodos
Revisión bibliográfica y de materiales de herbario. Se realizó la revisión bibliográfica sobre el área de estudio, y de materiales de herbario con datos de reportes o localización de especies recolectadas en años anteriores por otros especialistas.
La determinación hasta el nivel de especie se efectuó con el uso de claves dicotómicas (Sánchez y Morejón, 2011) y para la clasificación taxonómica se siguió el sistema de clasificación de Smith et al. (2006).
Para determinar el origen biogeográfico de las especies se consultaron los criterios de Caluff y Shelton (2002); Monterrosa et al. (2009); Palacios-Ríos et al. (2006);Sánchez (2000, 2007), Sánchez y Regalado (2003), Sánchez et al. (2006) y las bases de datos de www.Itis.Gov/selvlet/SingleRpt/; www.mobot.org/Manual.Plantas.
Para evaluar el grado de amenaza de las especies presentes en el área de estudio se consultó los criterios de Regalado et al. (2015).
Con el objetivo de evaluar la similitud biológica entre las formaciones vegetales de la localidad estudiada se utilizó el Índice de Sörensen (1948), teniendo en cuenta las especies compartidas en las áreas de estudio.
Trabajo de campo. Se efectuó entre los meses de septiembre 2013 a julio 2014, realizándose muestreos en cuatro formaciones vegetales de la localidad: bosque de galería, bosque semidecíduo mesófilo ambas formaciones constituyen pequeños relictos antropizados, la vegetación segetal y la vegetación ruderal, siguiendo la clasificación de Capote y Berazaín (1984).
El muestreo en la localidad se realizó por el conteo total de las especies (Ferro-Díaz, 2015) realizándose recorridos en forma de transectos lineales y menos frecuentemente sin rumbo fijo o aleatorio (Martínez y Alverson, 2006). En cada sitio de muestreo se efectuó la recolección del material botánico de interés, dudoso o desconocido, para lo cual se empleó el método tradicional de herborización. Las muestras de fácil identificación se realizaron en el campo. Para el muestreo se tuvieron en cuenta los siguientes datos:
Nombre científico
Nombre vernáculo
Localidad de recolecta
Coordenadas geográficas
Fecha
Altitud
Formación vegetal
Iluminación circundante
Tipo de suelo
Nombre de los recolectores
Observaciones
Se elaboró una lista de especies derivada de los muestreos en la localidad y la revisión de materiales de herbario. Para cada especie se consideró el tipo de formación vegetal en que vive, algunos aspectos de su ecología, modo de vida y la abundancia local por formaciones vegetales. El total de ejemplares colectados fueron depositados en la Sección Pteridophyta del Herbario BSC del Centro Oriental de Biodiversidad y ecosistemas.
Resultados
Riqueza de especies de la localidad. La pteridoflora de la localidad de El Cañón se compone de 62 táxones infragenéricos, agrupados en 23 géneros y 13 familias (tabla 1). El total de táxones infragenéricos reportadas constituye el 13% del total de la pteridoflora conocida para el distrito Turquinense, el 13% de las del sector Maestricum y el 10% de las presentes en la subprovincia Cuba Oriental. Del total de especies reportadas para la localidad una es endémica, 48 son nativas de la región caribeña y de América, ocho naturalizadas y dos introducidas. Se reportaron 53 especies (85%) con Preocupación menor (LC), a las restantes nueve no se le halló algún grado de amenaza.
En el área de estudio se halló el híbrido natural Adiantum melanoleucum × Adiantum pyramidale(Caluff, 2014 com. pers.), el cual muestra caracteres morfológicos combinados de ambas especies progenitoras. Este taxón fue localizado exclusivamente en el bosque de galería y se encuentra aún en estudio.
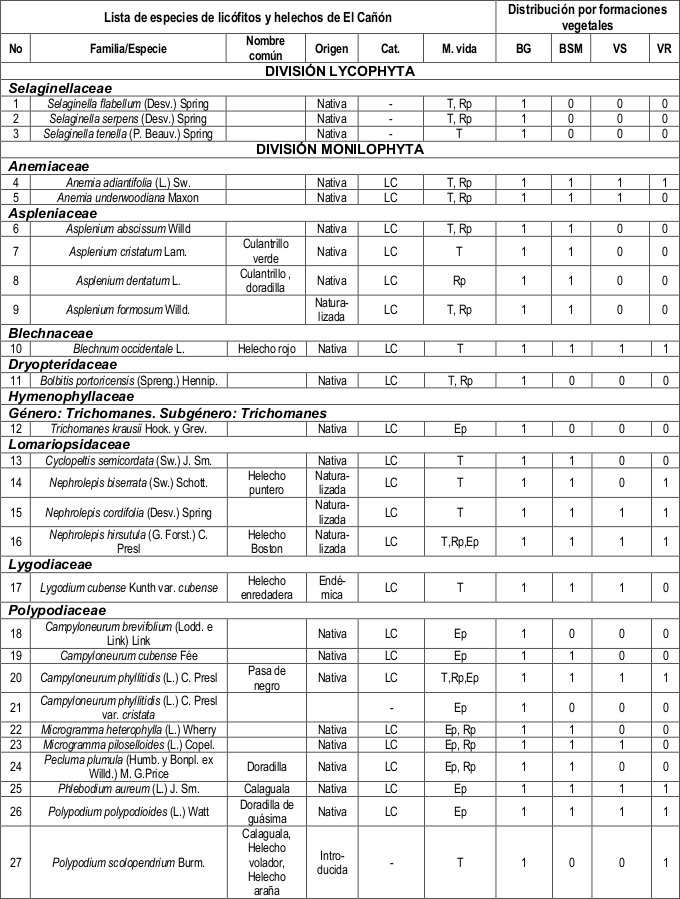
Dónde: Cat.: Categoría de amenaza, M. vida: Modo de vida, BG: Bosque de galería, BSM: Bosque semidecíduo mesófilo, VS: Vegetación segetal, VR: Vegetación ruderal, T: Terrestre, Rp: Epipétrico o rupícola, Ep: Epífito, 1: Presencia de la especie en la formación vegetal y 0: Ausencia de la especie en la formación vegetal.
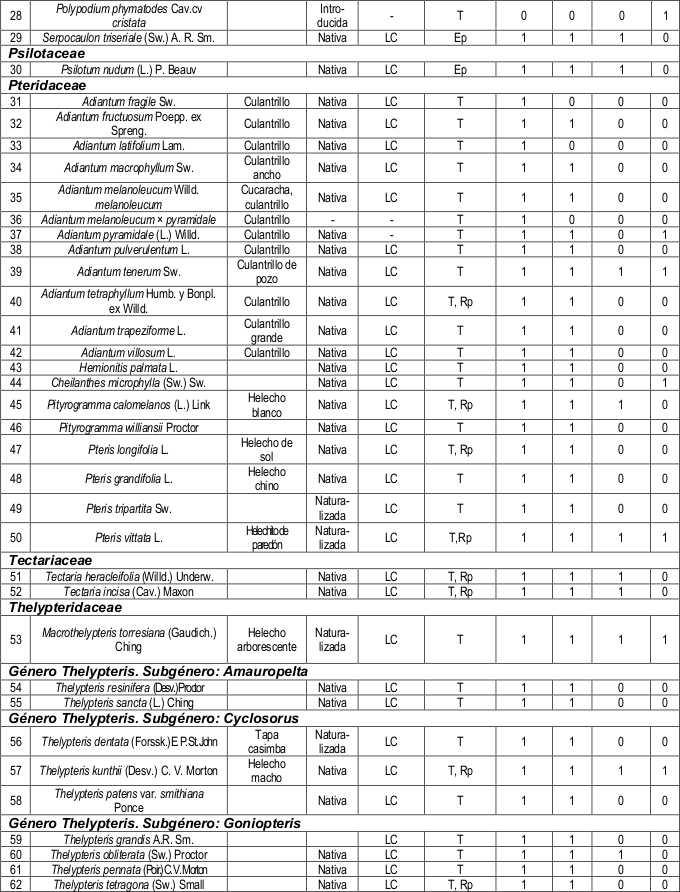
Dónde: Cat.: Categoría de amenaza, M. vida: Modo de vida, BG: Bosque de galería, BSM: Bosque semidecíduo mesófilo, VS: Vegetación segetal, VR: Vegetación ruderal, T: Terrestre, Rp: Epipétrico o rupícola, Ep: Epífito, 1: Presencia de la especie en la formación vegetal y 0: Ausencia de la especie en la formación vegetal.
Riqueza de especies por formaciones vegetales. La formación vegetal con la mayor riqueza de táxones infragenéricos resultó el bosque de galería, con un máximo 61 de táxones infragenéricos. También los pequeños relictos de bosques semideciduos mesófilos, con 50 especies. La vegetación segetal (con 20 de especies) y la ruderal (con 16 de especies) resultaron ser las de menor riqueza (figura 1).
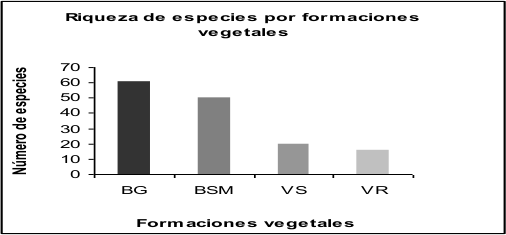
Figura 1.
Riqueza de especies por formaciones vegetales
Donde: BG: Bosque de galería; BMS: Bosque semidecíduo mesófilo; VS: Vegetación segetal; VR: Vegetación ruderal.
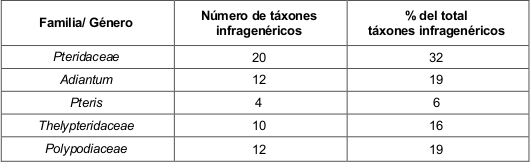
Familias representativas por formaciones vegetales. Las familias mejor representadas en las cuatro formaciones vegetales resultaron ser: Polypodiaceae, con el género Polypodium; Pteridaceae y Thelypteridaceae (tabla 2).
Las familias Aspleniaceae, Hymenophyllaceae y Selaginellaceae, resultaron ser las menos representativas y solamente se hallaron restringidas al bosque de galería, ubicándose en las zonas más próximas al curso de agua.
Análisis del modo de vida. Un análisis del modo de vida de los licófitos y los helechos en las formaciones vegetales estudiadas, revela que estos son mayormente terrestres (55%). Un 65% posee un modo de vida estricto y el 21% de las especies poseen más de un modo de vida, siendo más abundantes las que poseen el modo de vida terrestre y rupícola. Se registraron dos especies: Campyloneurum phyllitidis y Nephrolepis hirsutula en los tres modos de vida descritos (figura 2).
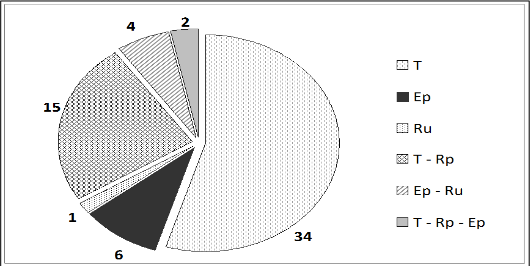
Figura 2.
Proporción de especies en cada modo de vida por formaciones vegetales. Dónde: T: Terrestre, Rp: Epipétrico, Ru: Rupícola, Ep: Epífito.
Análisis de similitud biológica por formaciones vegetales. Al analizar la similitud biológica entre las formaciones vegetales estudiadas, se halló mayor porcentaje de similitud entre el bosque de galería y el bosque semideciduo mesófilo con un 90% de similitud (figura 3), compartiendo entre ambas un total de 50 táxones infragenéricos.
También se halló similitud entre la vegetación segetal con vegetación ruderal (figuras 3 y 4). Del análisis realizado se pudo verificar que 11 especies son compartidas por las cuatro formaciones vegetales, las que coinciden con especies de amplia distribución.
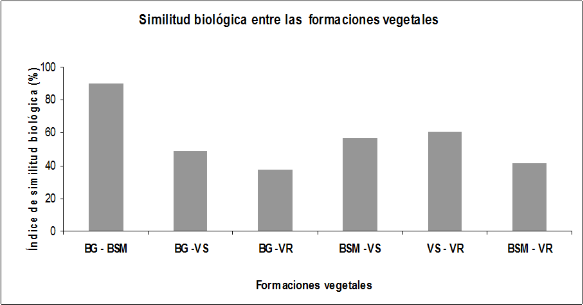
Figura 3.
Análisis de similitud biológica por formaciones vegetales.
Donde: BG: Bosque de galería; BMS: Bosque semidecíduo mesófilo; VS: Vegetación segetal; VR: Vegetación ruderal.
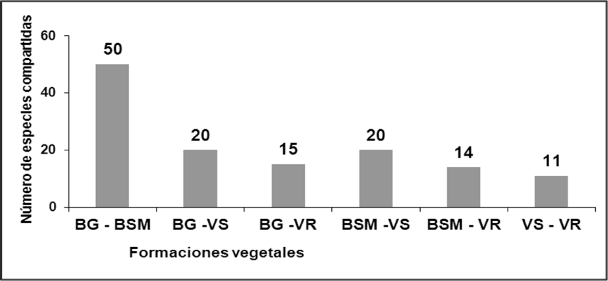
Figura 4.
Especies compartidas entre las formaciones vegetales.
Donde: BG: Bosque de galería; BMS: Bosque semidecíduo mesófilo; VS: Vegetación segetal; VR: Vegetación ruderal.
Discusión
La diversidad pteridológica de El Cañón está condicionada, por encontrarse en un valle intramontano, en la vertiente norte de la serranía, protegida del influjo de los vientos, con una pluviosidad (300-500 mm), una humedad relativa superior al 85%, y una mayor cobertura de vegetación (Chávez y Hernández, 1957; Montenegro, 1991 a, b, c; Viña-Bayés y Viña-Dávila, 1991). Asimismo, se halla más conservada de la actividad antropogénica, puesto que posee fuertes pendientes poco propicias para el desarrollo de las actividades agrícolas propias de la localidad. Resultados similares fueron obtenidos por Caluff y Shelton (1980), en su lista de especies de la Sierra de Boniato, en la cual se registran las especies recolectadas y su abundancia relativa donde se aprecia esta zona, como la de mayor diversidad.
La insuficiencia de especies endémicas podría estar determinada por las características edafoclimáticas del área estudiada, las cuales no poseen condiciones extremas y no propician el desarrollo de endémicos, unido al grado de antropización y el predominio de la vegetación secundaria. La antropización la evidencia, además, la presencia de cuatro especies con categoría de invasoras; éstas son: Macrothelypteris torresiana, Nephrolepis hirsutula, Pteris tripartita y Thelypteris dentata, las cuales son nativas de los trópicos de Asia Oriental alcanzando amplia distribución en toda el área estudiada (Caluff y Fuentes, 2010).
La riqueza de táxones infragenéricos del bosque de galería, así como en los bosques semideciduos mesófilos, se ve favorecida por hallarse en la vertiente norte de la Sierra de Boniato donde existen condiciones edáficas y climáticas favorables para su desarrollo, además de ser los mejores conservados de la actividad antropogénica. Resultados similares fueron apreciados por Caluff y Shelton (2000 a, b). Por el contrario la vegetación segetal y la ruderal, resultaron ser las de menor riqueza de táxones infragenéricos, ya que, en ambos casos, la cobertura arbórea es mayormente dispersa y no constituye barrera para el desarrollo de algunos helechos de amplia plasticidad ecológica, capaces de tolerar fuertes variaciones de humedad e iluminación.
La vegetación segetal es una zona de cultivo de plantas comestibles principalmente: plátano (Musa sp.), la yuca (Manihot esculenta Crantz), el quimbombó (Abelmoschus esculentus (L.) Moench), y están sometidas continuamente al deshierbe del terreno.
La vegetación ruderal se compone de pocas especies, mayormente sinantrópicas y naturalizadas, que a menudo colonizan los patios, las orillas de caminos y construcciones abandonadas o deterioradas. Entre estas especies se encuentran Adiantum tenerum, Cheilanthes microphylla, Nephrolepis hirsutula, Pteris vittata, Polypodium polypodioides y Thelypteris kunthii, estos resultados son similares a los obtenidos por los autores Caluff y Shelton (2000 b). Estos autores refieren estas y otras especies como comunes en sitos perturbados.
La representatividad en el área de las familias: Lomariopsidaceae, Polypodiaceae, Pteridaceae y Thelypteridaceae se debe a que éstas, se componen de especies en su mayoría de gran porte y que poseen adaptaciones morfofisiológicas que facilitan su amplia distribución, por ejemplo, la presencia de reproducción vegetativa, así como la producción de esporas de latencia prolongada poco exigentes para su germinación. En el caso de helechos epífitos, como Phlebodium aureum, la presencia de un rizoma suculento cubierto de escamas protectoras le permite tolerar prolongados períodos de sequía.
A diferencia de las anteriores, las familias Aspleniaceae, Hymenophyllaceae y Selaginellaceae, suelen tener pequeño tamaño y hojas diminutas, de textura delicada. Especialmente la familia Hymenophyllaceae, se compone de especies que poseen una sola capa de células en el tejido de lámina, por lo que los requerimientos ecológicos en estos helechos son más específicos y no son tolerantes a la sequía, ni a la iluminación intensa.
En un análisis más detallado del modo de vida epífito de las especies presentes, se observó que, para los bosques de galería, la altura sobre el forófito, podía ser variable y localizarse, fundamentalmente, en el lado del curso de agua. Sin embargo, no ocurre así para las restantes formaciones vegetales donde por lo general, estos se hallaron viviendo por encima de los seis metros, asociados fundamentalmente a bromeliáceas aprovechando los residuos de agua que éstas le proporcionan. Las especies de fanerógamas más usadas por los epífitos como forófitos fueron Mangifera indica L., Samanea saman Merr., Spondias mombin L. y Roystonea regia (Kunth) O. F. Cook, fundamentalmente árboles adultos de corteza rugosa, ya que estos le ofrecen mejor fijación al rizoma.
La similitud biológica entre el bosque de galería y el bosque semideciduo mesófilo, está condicionada por ser sitos más conservados, donde predomina mayor humedad relativa, propicia para el desarrollo de estas especies. La similitud biológica entre la vegetación segetal con vegetación ruderal, se debe a que ambas formaciones se componen de pocas especies principalmente sinantrópicas y de gran plasticidad ecológica, y se hallan perturbadas por la actividad antropogénica.
Conclusión
La pteridoflora de la localidad de El Cañón se compone de 62 taxones infragenéricos agrupados en 23 géneros y 13 familias.
De las cuatro formaciones vegetales estudiadas los bosques de galería y los pequeños relictos de bosques semideciduos mésófilos, son los que conservan la mayor diversidad de especies, lo que está determinado entre otros factores por ser los más conservados de la actividad antropogénica.
No se registraron especies amenazadas en el área estudiada.
Las familias de mayor representatividad en las cuatro formaciones vegetales resultaron ser: Lomariopsidaceae, Polypodiaceae, Pteridaceae y Thelypteridaceae.
Las formaciones vegetales de mayor similitud biológica resultaron ser los bosques de galería y los bosques semidecíduos mesófilos.
Recomendaciones
Ampliar el estudio de la pteridoflora a otras localidades de la Sierra de Boniato.
Desarrollar proyectos comunitarios que permitan difundir los valores de la pteridoflora de la localidad de El Cañón y contribuir a su protección y conservación.
Agradecimientos
Santiago de Cuba por su colaboración, en la asesoría de ésta investigación; en especial a Manuel García Caluff y Maité Serguera Niño.
Al Departamento de Botánica del Centro Oriental de Ecosistemas y Biodiversidad por el apoyo brindado para la publicación de esta investigación.
Literatura citada
Borhidi, A. y MUÑÍZ, O. 1986. Phytogeographic survey of Cuba II. Floristic relationships and phytogeographic subdivisión. Acta Bot. Hungarica 32(1-4):3-48.
BORHIDI, A. 1991. Phytogeography and Vegetation Ecology of Cuba. Akadémiai Kiadó. Budapest. 858 p.
Caluff, M.G. 2014. Comunicación personal. Jardín de los Helechos, ¨La Caridad¨, No. 129, Carr. El Caney, Santiago de Cuba. Director del Jardín de los Helechos.
CALUFF, M.G. y SHELTON, G. 1980. Lista de pteridíofitos de la Sierra de Boniato. [Inédito] Centro Oriental de Ecosistemas y Biodiversidad.
CALUFF, M.G. y SHELTON, G. 2000 a. Helechos y plantas afines por áreas geográficas en los macizos montañosos orientales. En: Informe del proyecto Diversidad biológica del macizo montañoso Sierra Maestra. Programa Científico-Técnico Nacional “Desarrollo sostenible de la montaña”. Tomo I. Bioeco. Santiago de Cuba. 84-99 p.
CALUFF, M.G. y SHELTON, G. 2000 b. Helechos y plantas afines por formaciones vegetales en la Sierra Maestra. En: Informe del proyecto Diversidad biológica del macizo montañoso Sierra Maestra. Programa Científico-Técnico Nacional “Desarrollo sostenible de la montaña”. Tomo I. Bioeco. Santiago de Cuba. 196-204 p.
CALUFF, M.G. y SHELTON, S.G. 2002.Catálogo del Jardín de los Helechos de Santiago de Cuba. Revista del Jardín Botánico Nacional 23 (2): 163-199.
CALUFF, M.G. y FUENTES, V. 2010. Malezas pteridofíticas de Cuba. Rev. Jard. Bot. Nac. 29: 51-56 p.
CALUFF, M.G.; SÁNCHEZ, C. y SHELTON, G. 2010. Helechos y plantas afines (Pteridophyta) de Cuba. Fitogeografía. Rev. Jard. Bot. Nac. 29:21-49 p.
CAPOTE, R. y BERAZAÍN, R. 1984. Clasificación de las formaciones vegetales de Cuba. Rev. Jard. Bot. Nac. 5 (2): 27-74 p.
CHÁVEZ, A. y HERNÁNDEZ, E. 1957. Observatorio de la Universidad de Oriente. Informe climatológico 41(2): 1-73.
FERRO-DÍAZ, J. 2015. Manual revisado de métodos útiles en el muestreo y análisis de la vegetación. ECOVIDA 5(1):139-186.
MARTÍNEZ, E. y ALVERSON, W. 2006. Plantas con semillas (Spermatophyta). En: Maceira, D, Fong A, Alverson G y Wachter, T (eds). Cuba: Pico Mogote. Rapid Biological Inventories 9. The Field Museum, Chicago. 51-53 p.
MONTENEGRO, M.U. 1991a. Mapa de Temperatura máxima media anual. Escala 1: 500 000. En: Atlas de Santiago de Cuba. Academia de Ciencias de Cuba. Santiago de Cuba. Mapa 26. 51-52 p.
MONTENEGRO, M.U. 1991b. Precipitación media anual. Escala 1: 750 000. En: Atlas de Santiago de Cuba. Academia de Ciencias de Cuba. Santiago de Cuba. Mapa 33. 58 p.
MONTENEGRO, M.U. 1991c. Precipitaciones y Humedad relativa, escala 1: 500 000. En: Atlas de Santiago de Cuba. Academia de Ciencias de Cuba. Mapa 28. 55-56 p.
MONTERROSA, S.J.A.; PEÑA-CHOCARRO, M.C.; KNAPP, S. y ESCOBAR, L.R. 2009. Guía de identificación de helechos de El Salvador. El Salvador 558 p.
NÚÑEZ, A. 1972. Geografía de Cuba, vol II. Instituto Cubano del Libro, La Habana, 283 p.
PALACIOS-RÍOS, M.; CALUFF, G.M. y SHELTON, G. 2006. Psilotaceae. En: Greuter, W y Rankin, R (eds). Flora de la República de Cuba. Serie A. Plantas vasculares, 11(11). Königstein: Koeltz Scientific Books. 6 p.
REGALADO, L.; SÁNCHEZ, C. y GONZÁLEZ-OLIVA, L. (Eds.) 2015. Categorización de helechos y licófitos de la flora de Cuba - 2015. Bissea 9 (número especial 3):1-146.
RENDA, A.; CALZADILLA, E.; BOUZA, J.A. y VALLE, M. 1980. Estudio sobre las condiciones edafológicas, fisiográficas y agrisilviculturales de la Sierra Maestra, Provincia Santiago de Cuba. Ministerio de la Agricultura, Centro de Investigación Forestal. 95 p.
SÁNCHEZ, C. 2000. Hymenophyllaceae. En: Greuter, W, Manitz, H y Rankin, R (eds). Flora de la República de Cuba. Serie A. Plantas vasculares, 4. Königstein: Koeltz Scientific Books. 70 p.
Sánchez, C. 2007. Los Helechos y Licófitos de Cuba. Ed. Científico Técnica. 226 p.
SÁNCHEZ, C. y REGALADO, L. 2003. Aspleniaceae. En: Greuter, W y Rankin, R (eds). Flora de la República de Cuba. Serie A. Plantas vasculares, 8(1). Königstein: Koeltz Scientific Books. 65 p.
SÁNCHEZ, C. y MOREJÓN, R. 2011. Clave de identificación para los géneros de helechos y licófitos de Cuba. Revista del Jardín Botánico Nacional Vol. 32/33 (2011-2012): 31-45.
SÁNCHEZ, C.; CALUFF, M. y REGALADO, L. 2006. Thelypteridaceae. En: Greuter, W. y Rankin, R. (eds). Flora de la República de Cuba. Fascículo 11(13). Koeltz Scientific Books Liechtenstein 4- 118. 123 p.
SMITH, A.R.; PRYER, K.M.; SCHUETTPELZ, E.; KORALL, P.; SCHNEIDER, H. and WOLF, P.G. 2006. A Classification for extant ferns. Taxon 55 (3):705-731.
SÖRENSEN, T.1948. Methods of establishing groups of equal amplitude in plant sociology based on similarity of species content. Det Kong. Danske Vidensk. Selsk. Biol. Skr. 5(4):1-34.
VIÑA-BAYÉS, N. y VIÑA-DÁVILA, N. 1991. Mapa de Carso. Escala 1: 250 000 En: Atlas de Santiago de Cuba. Santiago de Cuba. 22 p.
Bases de datos www.Itis.Gov/selvlet/SingleRpt/ www.mobot.org/Manual.Plantas