EASTERN CUBAʹS RAINFOREST. II. FIERA I, HOMEOSTASIS I AND FIERA II COMMUNITIES IN WESTERN SIERRA MAESTRA
EASTERN CUBAʹS RAINFOREST. II. FIERA I, HOMEOSTASIS I AND FIERA II COMMUNITIES IN WESTERN SIERRA MAESTRA
Foresta Veracruzana, vol. 24, núm. 1, pp. 7-14, 2022
Recursos Genéticos Forestales
Resumen: En la Sierra Maestra se desarrolla la pluvisilva montana entre los 800 y los 1 500 msnm con una abundante humedad ambiental. Se estudiaron mediante la metodología de la Escuela Zurich Montpellier dos asociaciones con dos subasociaciones que se corresponden con las comunidades secundarias Fiera I, Homeostasis I y Fiera II.
Palabras clave: Bosques húmedos, comunidades vegetales, fitocenología, Sierra Maestra.
Abstract: In the Sierra Maestra the rainforests occupied mountains territories between 800 and 1 500 meters over sea level. By using the Zurich Montpellier school methodology successional phytocoenoses Fiera I, Homeostasis I and Fiera II are described. They were two associations with two subassociations too.
Keywords: Humid forests, community, sintaxonomy, Sierra Maestra.
Introduction
Reyes (2006) makes a classification of the vegetation of the Sierra Maestra, including the montane rainforest and its successional stages which is followed in this work (Reyes and Acosta 2005, 2006).
According to the great fragility Sierra Maestra’s ecosystems to badly changes of it use, the system used of "clean-tomb-itch-burn-inhabits" in so montainous areas, produced an extraordinary degradation, mainly in biomass and soils, breaking its delicate ecosystemic stability. According to Suárez et al. (2014) the current production of this degraded soils reaches around 50% of the potential minimum yield. Therefore, the objective of this work is to analyze the phytocoenoses found in the Sierra Maestra’s Early successional communities (Fiera I and Homeostasis I) and the first stage of the Late successional communities (Fiera II).
Material and methods
The natural conditions of the studied area and the sampling methodology is found in the former work of this series, Sierra Maestra’s mountain rainforest. I. Immediate secondary communities, although the last were exposed summarily.
Sampling methodology. According to the large Cuban experience, the methodology of the Zurich-Montpellier School (Braun-Blanquet, 1951) is considered the most coherent with our vegetation; therefore, phytocoenological inventories (lists, stands, samples, releves), with a minimum area of 625 m. (Reyes, 2005), were made by this method. The abundance-dominance of each species was estimated in the usual way in this type of work (Reyes and Acosta, 2012). In addition, observations of the oecotope (slope, exposition, altitude, general, nano and micro relief) were made in the place of the samples and their surroundings.
The ordering of vegetation inventories and the separation of phytocoenoses (syntaxa) was carried out by phytosociological methods (Scamoni, 1960). For the characteristic combination of the associations, the species with degrees of presence IV and V (Scamoni, op. cit.) were used, and for the subassociations and variants the differential combinations.
The Phytosociological Nomenclature International Code (Weber et al., 2000) was followed for the categorization and nomenclature of phytocoenoses ranks. The name of said syntaxa was made according to the exposed code.
Completed scientific names (genus, species and author) can be observed in tables and Acevedo-Rodríguez and Strong (2012), sometime amended by Greuter and Rankin (2016, 2017), Borhidi et al. (2017) and Sánchez (2017). Collected specimens are in Herbarium BSC.
Results
According to this application, the study of Fiera I, Homeostasis I and Fiera II successional stages the subsequent phytosociological arrangement was made in Western Sierra Maestra:
Clethro cubensis-Cyrilletea silvae Reyes 2019.
Clethro cubensis-Magnolietalia cubensis Reyes.
Cyatheo furfuraceae-Magnolion cubensis Reyes.
Studied associations corresponding to the stages of Fiera I, Homeostasis I and Fiera II:
· Piperi adunci-Myrsinietum coriaceae.
· Cyatheo-Cyrilletum silvae.
Piperi adunci-Myrsinietum coriaceae Reyes & Acosta ass. nov. Table 1, holotypus: rel. 1.
It constitutes an early successional community (Fiera I), this inventories in spite to have common elements, may present some differences according to the origin of the community. These samples could be around 25 years old. Studied 17.03.2003 (20. 04ʹ N 76. 34ʹ W).
One of the inventories seems to have been a pasture, near the west branch the La Plata River. The tree layer (E.), with 40% coverage, is between 6 and 8 m high with very isolated 10 m specimens. It is also observed a shrub-herbaceous layer with around 2 m (with some specimens of 2.5 to 3 m), very dense, with 100% coverage.
Near this one there is another edaphotope (edapho-climatic conditions of the site), which is an open landslide, colonized by pines; 15 years ago, it only had some scattered pine trees around 5 m high, so the age of around 25 years is estimated. Currently it has closed areas, the sampled part has an arboreal coverage of scattered pines between 30 and 40% and around 10 to 15 m high, open and with very humid zones. In the shrub layer, with 60% coverage, the most important species are Pinus maestrensis Bisse, Piper aduncum L. and Cyrilla silvaeBerazaín. The herbaceous layer is very dense, with 100%, the dominant species are: Schizachyrium gracile (Spreng.) Nash, Lycopodiella cernua L., Sticherus bifidus (Willd.) Ching, Andropogon bicornis L. and Pteridium arachnoideum (Kaulf.) Maxon. None of the arboreal species of the mountain rainforest is found, and there are several heliophilous (sun-loving) and pioneer species. At the same time the arborescent ferns that dominate successional stages more advanced are said (table 1). The substrate is constituted by ferralitic weathering crust with important rocky areas.
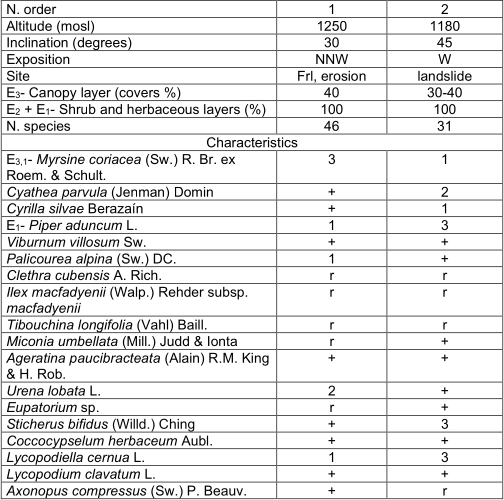
In addition. Inv. 1. Miconia dodecandra (Desr.) Cogn. 2, Brunellia comocladifolia subsp. cubensis Cuatrec. 3, Cecropia peltata L. r, Zanthoxylum martinicense (Lam.) DC. r, Cyathea furfuracea Baker 2, Urochloa dictyoneura (Fig. & De Not.) Veldkamp 3, Pteridium arachnoideum (Kaulf.) Maxon 3, Ocotea leucoxylon (Sw.) Laness. r, Magnolia cubensis Urb. subsp. cubensis r, Ixora ferrea (J. F. Gmel.) Benth. r, Psychotria grandis Sw. +, Callicarpa ferruginea Sw. +, Hypericum hypericoides (L.) Crantz 1, Alsophylla major Caluff & Shelton r, Eugenia laeteviridis Urb. r, Lobelia assurgens L. 1, Meriania albiflora Carmenate & Michelang. r, Chromolaena odorata (L.) R.M. King & H. Rob. r, Spermacoce laevis Lam. +, Scleria lithosperma (L.) Sw. +, Begonia cubensis Hassk. 1, Ichnanthus sp. r, Zeugites americanus Willd. +, Smilax domingensis Willd. r, Vitis tiliifolia Humb. & Bonpl. ex Roem. & Schult. r, Passiflora sexflora Juss. r, Cissus grisebachii Planch. r, Hypolepis nigrescens Hook. +; Inv. 2. Pinus maestrensis Bisse 3, Schizachyrium gracile (Spreng.) Nash 3, Andropogon bicornis L. 2, Guzmania sp. 2, Chaptalia sp. 2, Pluchea odorata (L.) Cass. +, Psychotria greeneana Urb. +, Bletia purpurea (Lam.) DC. r, Phaius tankervilliae (Banks) Blume 1, Desmodium canum (J.F. Gmel.) Schinz & Thell. +, D. triflorum (L.) DC. r, Bidens pilosa L. +, Epidendrum umbelliferum J.F. Gmel. r, E.nocturnum Jacq. r.
Cyatheo-Cyrilletum sylvae Reyes & Acosta ass. nov. Table 2, holotypus rel. 3.
This association constitutes the successional stages Homeostasis I and Fiera II, which is separated into two subassociations (studied near Barrio Adentro-El Manguito 15-22.03.2004; 20° 04ʹ N 76° 42ʹ W).
· Cyatheo-Cyrilletum sylvae typicum (Homeostasis I).
· Cyatheo-Cyrilletum sylvae arthrostylidietosum multispicati (Fiera II).
Typicum Reyes subass.nov. table 2, typus rel. 3.
In this mountain rainforest, during the phase of Homeostasis I, Cyrilla sylvae and the arborescent ferns Cyathea furfuracea Baker and C. parvula dominate the shrub-arboreal layer with a height between 5 and 8 m, occasionally reaching up to 12 m; they generally close the canopy, with 100% coverage. This creates umbrophilous conditions, entering the original forest species, they reach this layer or are already in the lower, in turn, begin to disappear the pioneer species such as Pteridium arachnoideum, Gleichenella pectinata (Willd.) Ching, Urena lobata L. and Chromolaena odorata (L.) R.M. King & H. Rob., among others. Several species are constant in this stage (table 2). The diversification of the mulch is clearly observed; there is an L layer of 1.5 to 3 cm and F between 0.5 and 1.5 cm; in layer H are large differences, from vestiges, to a well-developed root mat of 10 cm in thickness.
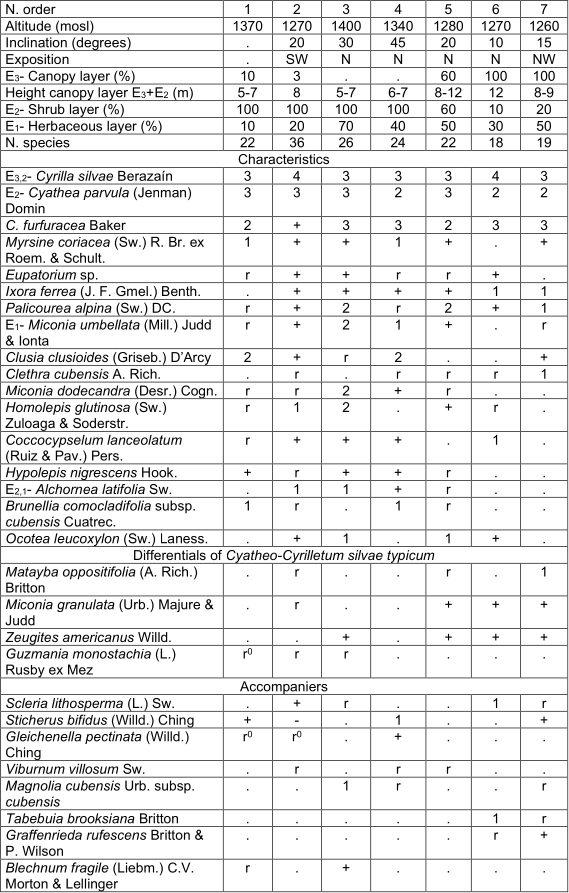
In addition. Inv. 1. Lycopodium clavatum L. r, Arthrostylidium multispicatum Pilg. r, Bidens cynapiifolius Kunth r; Inv. 2. Sideroxylon jubilla (Urb.) T.D. Penn. r, Hypericum hypericoides (L.) Crantz (r), Pteridium arachnoideum (Kaulf.) Maxon 1, Lobelia oxyphylla Urb. (r), Passiflora sexflora Juss. r, Cissus trifoliata (L.) L. r; Inv. 3. Rubus turquinensis Rydb. r, Ageratina paucibracteata (Alain) R.M. King & H. Rob. +, Isochilus linearis (Jacq.) R. Br. r; Inv. 4. Lycopodiella cernua L. r, Psychotria greeneana Urb. +; Inv. 5. Meriania albiflora Carmenate & Michelang. +; Inv. 6. Antirhea sp. 1.1, Lobelia assurgens L. r, Lobelia sp. 1; Inv. 7. Politrichum sp. 2.
In addition. Inv. 1. Lycopodium clavatum L. r, Arthrostylidium multispicatum Pilg. r, Bidens cynapiifolius Kunth r; Inv. 2. Sideroxylon jubilla(Urb.) T.D. Penn. r, Hypericum hypericoides (L.) Crantz (r), Pteridium arachnoideum (Kaulf.) Maxon 1, Lobelia oxyphylla Urb. (r), Passiflora sexflora Juss. r, Cissus trifoliata (L.) L. r; Inv. 3. Rubus turquinensis Rydb. r, Ageratina paucibracteata (Alain) R.M. King & H. Rob. +, Isochilus linearis (Jacq.) R. Br. r; Inv. 4. Lycopodiella cernua L. r, Psychotria greeneana Urb. +; Inv. 5. Meriania albiflora Carmenate & Michelang. +; Inv. 6. Antirhea sp. 1.1, Lobelia assurgens L. r, Lobelia sp. 1; Inv. 7. Politrichum sp. 2.
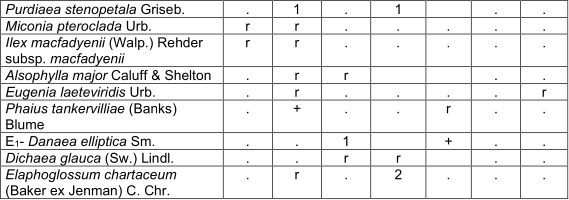
arthrostylidietosum multispicati Reyes subass. nov. Table 3, holotypus rel. 7.
They form an arboreal layer of 8 to 12 m, with isolated individuals of 15 m and coverage between 40 and 100%. The most abundant species in this layer are Miconia dodecandra, Myrsine coriacea and Cyrilla silvae, sometimes also Brunelliacomocladifolia subsp. cubensis, Alchornea latifolia and Clusia clusioides; are also presented Ixora ferrea, Viburnum villosum Sw., Cyathea furfuracea, Ocotea leucoxylon,Clethra cubensis, Matayba oppositifolia, Miconia pteroclada Urb. and occasionally Aiouea montana, Schefflera morototoni(Aubl.) Maguire and Magnolia cubensis subsp. cubensis. The shrub layer is still dense and generally covers between 60 and 90%. The most abundant species are Cyathea parvula, C. furfuracea, Cyrilla sylvae and Palicourea alpina. Hypolepis nigrescens, Alchornea latifolia and Graffenrieda rufescens Britton & P. Wilson, are sometimes abundant; Ixora ferrea, Miconia umbellata (Mill.) Judd & Ionta, Myrsine coriacea and Alsophyllamajor, among others are also observed. The herbaceous layer covers mostly between 50 and 70%; it is very diverse and the most abundant species are Cyathea parvula, Hypolepis nigrescens, Elaphoglossum chartaceum (Baker ex Jenman) C. Chr. and Miconia umbellata; sometimes Psychotria greeneana Urb., Palicourea alpina, Coccocypselum lanceolatum (Ruiz & Pav.) Pers. and Homolepis glutinosa. They are constant although with little coverage Callicarpa ferruginea Sw., Blechnum fragile (Liebm.) C.V. Morton & Lellinger, Arthrostylidium multispicatum, Eupatorium sp. and Oplismenus hirtellus subsp. setarius (Lam.) Mez ex Ekman. Two variants are observed with small differences, Schefflera morotottoni presents a lower height of the arboreal layer and develops on the north slope; in turn, Alsophylla major has a higher height and coverage of the trees, mainly the sub-variant Matayba oppositifolia (table 3). It occurs in leached red ferralitic soils, on steep slopes and between 1 100 and 1 400 mosl.
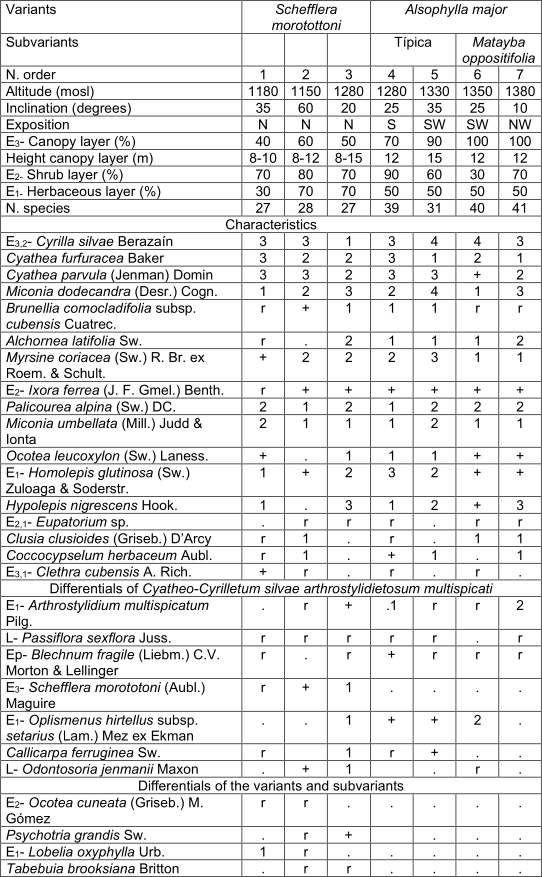
In addition. Inv. 1. Alsophila balanocarpa (D.C. Eaton) D.S. Conant +, Zeugites americanus Willd. r; Inv. 3. Miconia granulata (Urb.) Majure & Judd r; Inv. 4. Melastomataceae +, Dendropanax arboreus (L.) Decne. & Planch. r, Serpocaulon loriceum (L.) A.R. Sm. r, Odontosoria scandens (Desv.) C. Chr. r, Cissus trifoliata (L.) L. r; Inv. 5. Dendrophthora sp. r, Hedyosmum grisebachii Solms r, Cheiroglossa palmata (L.) C. Presl r, Asplenium serra Langsd. & Fisch. r; Inv. 6. Isochilus linearis (Jacq.) R. Br. r, Hymenophyllaceae r, Lyonia maestrensis Acuña & Roig r, Trichomanes robustum E. Fourn. +, Elaphoglossum glabellum J. Sm. r, Pleopeltis squamata (L.) J. Sm. r, Grammitis sp. r, Trichomanes scandens L. r; Inv. 7. Rubus turquinensis Rydb. r, Blechnum occidentale L. r, Phaius tankervilliae (Banks) Blume +, Leucobrium giganteum Mull. Hal. 1, Danaea elliptica Sm. r, Antirhea sp. r, Hymenophyllum polyanthos (Sw.) Sw. r.
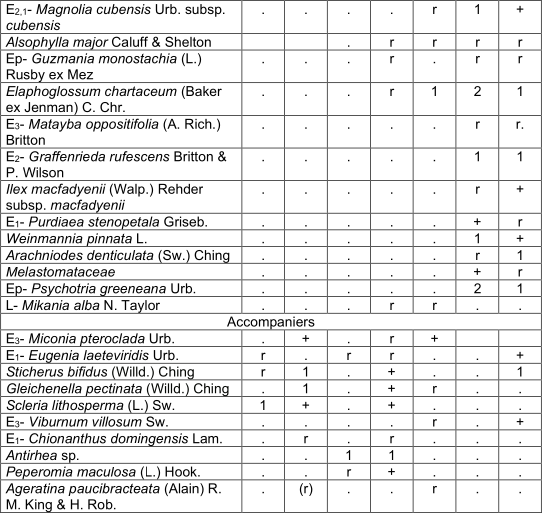
In addition. Inv. 1. Alsophila balanocarpa (D.C. Eaton) D.S. Conant +, Zeugites americanus Willd. r; Inv. 3. Miconia granulata (Urb.) Majure & Judd r; Inv. 4. Melastomataceae +, Dendropanax arboreus (L.) Decne. & Planch. r, Serpocaulon loriceum (L.) A.R. Sm. r, Odontosoria scandens (Desv.) C. Chr. r, Cissus trifoliata (L.) L. r; Inv. 5. Dendrophthora sp. r, Hedyosmum grisebachii Solms r, Cheiroglossa palmata (L.) C. Presl r, Asplenium serra Langsd. & Fisch. r; Inv. 6. Isochilus linearis (Jacq.) R. Br. r, Hymenophyllaceae r, Lyonia maestrensis Acuña & Roig r, Trichomanes robustum E. Fourn. +, Elaphoglossum glabellum J. Sm. r, Pleopeltis squamata (L.) J. Sm. r, Grammitis sp. r, Trichomanes scandens L. r; Inv. 7. Rubus turquinensis Rydb. r, Blechnum occidentale L. r, Phaius tankervilliae (Banks) Blume +, Leucobrium giganteum Mull. Hal. 1, Danaea elliptica Sm. r, Antirhea sp. r, Hymenophyllum polyanthos (Sw.) Sw. r.
Discussion
This work ratified all of the exposed in Reyes and Acostaʹs (2005) about the successional stages in Sierra Maestra: here was Fiera I, Homeostasis I and Fiera II stages developed.
The Fiera I (Capote et al., 1988) is the stage where there is an strong competition between the pioneer species and other heliophylous elements, including some elements of the mountain rainforest as seedlings or bush size; their physiognomy and composition, although they have common elements, may present some differences according to the origin of the community. At the end in this stage begins some taxa to increase in size and cover with some advantage about other species. The Homeostasis I stage is where some species dominate the shrub - arboreal layer and make a cover (shade) that reduce light entrance, help umbrophilous rainforest species and influence negatively the pioneer and heliophilous ones. The late successional stage (Fiera II) begins when the tree species surpass the layer of Homeostasis I, interacting competitively among them.
It is of interest the great floristic dissimilarities that are observed among the Fiera I, mainly with a type of "r" selection (Gliessman, 2002), and Homeostasis I and Fiera II, where already in the late communities dominate elements that are fundamentally of the type of selection "c" (Reyes and Acosta, 2005).
Conclusión
In general, in these stages important vegetation’s composition and structural changes were implemented, where pioneer species were negatively influenced and begin rainforest species eintrance and/or dominate because the shade reduce light entrance in ecosystem’s inner.
Cited literature
ACEVEDO-RODRÍGUEZ, P. and STRONG, M.T. 2012. Catalogue of Seed Plants of the West Indies. Smithsonian Contributions to Botany 98. 1192 p. http://repository.si.edu/xmlui/handle/ 10088/17551.
BORHIDI, A.; FERNÁNDEZ-ZEQUEIRA. M. and OVIEDO, P.R. 2017. Rubiáceas de Cuba. Akademiai Kiadó. 494 p. Disponible en: https://go.gale.com/ps/anonymous?id=GALE%7CA581941449&sid=googleScholar&v=2.1&it=r&linkaccess=abs&issn=02366495&p=AONE&sw=w. BRAUN-BLANQUET, J. 1951. Pflanzensoziologie; Grundzüge der Vegetationskunde. 2 Aufl. Wien.
BRAUN-BLANQUET, J. 1951. Pflanzensoziologie; Grundzüge der Vegetationskunde. 2 Aufl. Wien.
CAPOTE, R.P.; MENÉNDEZ, L.; GARCÍA, E.E. and HERRERA, R.A. 1988. Sucesión Vegetal. En: Herrera, R.A., Menéndez, L., Rodríguez, M.A., García, E.E, Eds. Ecología de los bosques siempreverdes de la Sierra del Rosario, Cuba. Montevideo. ROSTLAC. p. 272-295.
GLIESSMAN, S.R. 2002. Agroecología: Procesos ecológicos en agricultura sostenible. CATIE, Turrialba.
GREUTER, W. and RANKIN, R.R. 2016. The Spermatophyta of Cuba. A Preliminary Checklist. Part II: Checklist. Botanischer Garten & Botanisches Museum Berlin-Dahlem Jardín Botánico Nacional, Universidad de La Habana. 398 p. ISBN 978-3-946292-06-7. Disponible en: https://www.researchgate.net/publication/301326599.
GREUTER, W. and RANKIN, R.R. 2017. Vascular plants of Cuba. A preliminary checklist. Second, updated Edition of The Spermatophyte of Cuba, with Pteridophyte added. Botanischer Garten - Botanisches Museum Berlin-Dahlem & Jardín Botánico Nacional, Universidad de la Habana. 444 p. Disponible en: https://www.bgbm.org/sites/default/files/plantas_vasculares_de_cuba_2017-12-18.pdf.
REYES, O.J. 2005. Estudio sinecológico de las pluvisilvas submontanas sobre rocas del complejo metamórfico. Foresta Veracruzana 7(2): 15-22.
REYES, O.J. 2006. Clasificación de la vegetación de la Sierra Maestra. Editorial Academia. Biodiversidad de Cuba Oriental, Vol. VIII. p. 23-41.
REYES, O.J. and ACOSTA, F.C. 2005. Successional Stages in Mountain Rainforest of La Bayamesa Nacional Park, Cuba. In Rapid Biological Inventories, Report/Informe Nr. 13. Appendice/Apéndice I. p. 150-161.
REYES, O.J. and ACOSTA, F.C. 2006. Vegetation. Pico Mogote. In Rapid Biological Inventories. Report 09. Maceira, D., A. Fong & W.S. Alverson (Eds.). p. 40-46. The Field Museum, Chicago.
REYES, O.J. and ACOSTA, F.C. 2012. Sintáxones de los pinares de Pinus cubensis de la zona nororiental de Cuba. Lazaroa 33: 111-169.
SÁNCHEZ, C. 2017. Lista de los helechos y licófitos de Cuba. Brittonia, DOI 10.1007/s12228 017-9485-1. ISSN: 0007-196X (print) ISSN: 1938-436X (electronic, published online 23 June 2017). 24 p.
SCAMONI, A. 1960. Waldgesellschaften und Waldstandorte. Akademie-Verlag. Berlin. 326 p.
SUÁREZ, C.; MARTÍNEZ, Y.; CORRÍA, N.; MARTÍNEZ, L.; RAMÓN, A.M. and HERNÁNDEZ, A. 2014. Situación Ambiental de la Región Especial de Desarrollo Sostenible (REDS) Sierra Maestra (inédito). Informe Órgano de Montaña Sierra Maestra. 48 p.
WEBER, H.E.; MORAVEC, J. and. THEURILLAT, J.P. 2000. Internacional Code of Phytosociological Nomenclature. 3rd Edition. Journal of Vegetation Science 11: 739-768.