An updated biostratigraphy for the late Aragonian and Vallesian of the Vallès-Penedès Basin (Catalonia)
An updated biostratigraphy for the late Aragonian and Vallesian of the Vallès-Penedès Basin (Catalonia)
Geologica Acta: an international earth science journal, vol. 14, no. 3, pp. 195-217, 2016
Universitat de Barcelona

This work is licensed under Creative Commons Attribution-ShareAlike 4.0 International.
Received: 15 July 2015
Accepted: 15 March 2016
Funding
Funding source: Spanish Ministerio de Economía y Competitividad
Contract number: CGL2014-54373-P and RYC2013-12470 to ICV; CGL2014-55900-P
Award recipient: An updated biostratigraphy for the late Aragonian and Vallesian of the Vallès-Penedès Basin (Catalonia)
Funding
Funding source: Agència de Gestió d’Ajuts Universitaris i de Recerca
Contract number: 2014 SGR 416
Award recipient: An updated biostratigraphy for the late Aragonian and Vallesian of the Vallès-Penedès Basin (Catalonia)
Abstract: The Vallès-Penedès Basin (Catalonia, Spain) is a classical area for the study of the Miocene land mammal faunas and includes one of the densest and most continuous records in Eurasia. Furthermore, it is the type area for the Vallesian European land mammal age. After decades of study a huge amount of bio- and magnetostratigraphic data have been collected, allowing an unprecedented dating accuracy. Here we provide an updated local biostratigraphy for the late Aragonian, Vallesian and Turolian of the Vallès-Penedès Basin. This new biostratigraphic scheme is almost exclusively based on fossil rodents, which are the most abundant and one of the best known mammal orders in the area. Our proposal represents a significant refinement compared to previous attempts and provides a formal diagnosis and description of each zone, as well as clear definition of boundaries and a reference locality and section. The chronology of zone boundaries and main bioevents is based on detailed magnetostratigraphic data. The defined biozones allow for the correlation of the sites without associated magnetostratigraphical data. Finally, the correlation of the Vallès-Penedès local zones with other detailed local biostratigraphies, such as those of the Calatayud-Montalbán and Teruel basins (east-central Spain) is discussed. The sequence and chronology of the main bioevents is roughly comparable, although the rodent succession and the structure of the assemblage show important differences between these areas.
Keywords: Miocene, Biostratigraphy, Magnetostratigraphy, Rodents, Iberian Peninsula.
INTRODUCTION
The Vallès-Penedès Basin (Catalonia, Spain) is a reference area for the study of Miocene land mammal faunas of Europe. The earliest fossil finds date back to the late 19th century and the area has been sampled systematically ever since. Most of the older key contributions to the knowledge of the fossil faunas of this basin were by the team led by Miquel Crusafont Pairó between the 1940s and the 1970s. These comprised the discovery of numerous new sites, the description of many new taxa and the first attempts to place the Vallès-Penedès mammal succession in a detailed stratigraphical framework (for a more detailed review see Casanovas-Vilar et al., 2011a). Crusafont and coworkers soon noted that most of the late Miocene mammal assemblages from the Vallès-Penedès Basin differed from those of the purportedly coeval European sites known at the time, such as Pikermi in Greece. The Vallès-Penedès assemblages were characterized by the coexistence of the three-toed hipparionin horses with faunal elements that were characteristic of the middle Miocene, such as certain rhinos, cervids, suoids and chalicotheres. At that time, hipparionin horses were thought to be one of the defining elements of the so-called Pikermian faunas, which characteristically include a high diversity of open country herbivores, such as bovids and giraffids, whereas forest herbivores, such as cervids and certain rhinos, are very rare. The Pikermian faunas represent a rather uniform mammal community that later during the Turolian extended its range into Western Europe (Eronen et al., 2009). Then, typical Pikermian faunas were known from the sites of Piera in the Vallès-Penedès Basin (Villalta Comella and Crusafont Pairó, 1946; Crusafont Pairó, 1950), but apparently some ‘Pikermian’ taxa such as the hipparionin equids had dispersed earlier. Furthermore, Crusafont Pairó (1950) already noted that Hipotherium catalaunicum, the species present at the Vallès-Penedès sites, is more archaic than any of the hipparionin species present at Pikermi (see Bernor et al., 1996). On the other hand, Cremohipparion mediterraneum, one of the species described from Pikermi, might be present at the Piera sites (Pirlot, 1956; but see Bernor et al., 1996 for a different opionion). Therefore, the Vallès-Penedès assemblages clearly represented a transition between the forest faunas of the middle Miocene and the open-country, savannah-like faunas of the late Miocene.
At the time Crusafont and co-workers were surveying the basin, the Eastern Paratethys marine stages were often informally used for the correlation of the mammalian faunas. Pikermi was correlated to the late Pontian, whereas older European sites, such as La Grive (France), were correlated to the Sarmatian. The late Miocene Vallès-Penedès sites represented an intermediate stage, and were informally correlated to the Meotian, which is placed between the Sarmatian and the Pontian (Crusafont Pairó, 1950; Crusafont Pairó and Villalta Comella, 1947). The term “Spanish Meotian” was used for some time in the literature, until Crusafont formally proposed the Vallesian mammal stage for the late Miocene mammal assemblages of the Vallès-Penedès Basin (Crusafont Pairó, 1950). Now we know that the Meotian is indeed younger than the Vallesian (Hilgen et al., 2012). In later works (e.g.Crusafont Pairó and Truyols Santonja, 1960), the team better characterized the Vallesian succession in its type area and discussed further the correlation of the newly introduced mammal stage. The Vallesian was soon accepted in Eurasia because of the simple criterion used to define its lower boundary: the dispersal of hipparionin horses. These horses originated in North America and crossed the Bering Strait at a time of global sea-level lowering near to the middle/late Miocene transition (Bernor et al., 1996; Woodburne et al., 1981, 1995). A considerable effort has been devoted to firmly date the first appearance of these horses in the Old World, with ages ranging from 11.2 to 10.3Ma in different regions (Barry and Flynn, 1990; Barry et al., 1982, 2002; Bernor et al., 1988, 1996; Flynn et al., 2013; Garcés et al., 1996, 1997, 2003; Kappelman et al., 2003; Van Dam et al., 2014).
As far as the subdivision of the Vallesian is concerned, Crusafont Pairó and Truyols Santonja (1960) already noticed that this stage may be subdivided in its type area into the early and late Vallesian. The early Vallesian would include sites such as Can Poncic and Can Llobateres 1, whereas the late Vallesian would include the sites near the towns of Terrassa and Viladecavalls (Sant Miquel de Toudell, Can Purull, La Tarumba and others). The forest elements of middle Miocene origin are rarer during the late Vallesian, coinciding with the first record of new eastern immigrants. With the introduction of the MN (Mammal Neogene) zonation as a standard correlation scheme for the European Neogene faunas (Mein, 1975), the early and late Vallesian were equated to zones MN9 and MN10, respectively. This implied the extension to other European regions, not only of this mammal stage but also of its major subdivisions. Later studies of the Vallesian in its type area further subdivided it in a number of local zones mostly on the basis of the composition of the rodent faunas (Agustí, 1981, 1982; Agustí and Moyà-Solà, 1991). Finally, a magnetostratigraphic study of the Vallès sector of the basin undertaken during the 1990s allowed the correlation of several sites and fossiliferous sections to the Geomagnetic Polarity Time Scale (GPTS), thus enabling a well-supported chronology for the local zones and main bioevents (Agustí et al., 1997; Garcés et al., 1996). Later work in the Penedès sector of the basin provided further extension of the bio-magnetostratigraphic framework down into the late Aragonian and its transition with the Vallesian (Casanovas-Vilar et al., 2011b; Moyà-Solà et al., 2009). A preliminary new zonation ranging from the late Aragonian to the Vallesian was given in Casanovas Vilar (2007), later updated in Casanovas-Vilar et al. (2011b). These zonations, however, lacked formal definitions and precise zone boundaries. Furthermore, at that time only test samples were available for most of the late Aragonian sites of the basin. Here we provide an updated biostratigraphy of the late Aragonian and the Vallesian in the Vallès-Penedès Basin that is based on rodents, arguably the most abundant and one of the best studied mammal orders. Additionally, we also propose a preliminary biostratigraphy of the Turolian based on the limited information currently available in the basin. We have reviewed the microfauna of all the Vallesian sites and included all the newly-discovered late Aragonian sites from the area of els Hostalets de Pierola (Alba et al., 2006, 2011), in the Penedès sector of the basin. Unambiguous zone definitions and dated boundaries are provided. Finally, the chronology of the main bioevents and correlation of the proposed zonation with other Spanish and French records are discussed.
GEOLOGICAL SETTING
The Vallès-Penedès Basin (Figs. 1;2) is part of a widespread system of NE-SW and NNE-SSW oriented horst and half-grabens that were formed as a result of the opening of the northwestern Mediterranean during the latest Oligocene and the Miocene (Bartrina et al., 1992; Cabrera and Calvet, 1996; Cabrera et al., 2004; Roca and Guimerà, 1992; Roca et al., 1999). It is an elongated half-graben (100km long by 12-14km wide) parallel to the Catalan coastline and bounded by the prelittoral range to the NW and the littoral range to the SE (Garraf-Montnegre horst). The Vallès-Penedès master fault defines its northwestern margin, and its basement dips gently towards this margin, where it attains up to 4,000m in depth (Bartrina et al., 1992; Roca et al., 1999). The sedimentary filling of the Vallès-Penedès Basin started in the early Miocene (Ramblian/early Burdigalian; Cabrera, 1981a, b; Cabrera et al., 1991, 2004; de Gibert and Casanovas-Vilar, 2011) and ended in the late Miocene (Turolian/Messinian). Major features of the stratigraphic record in the Vallès-Penedès half-graben were closely controlled by the tectonic activity of its main bounding faults and the sea level changes in the western Mediterranean (Bartrina et al., 1992; Cabrera and Calvet, 1996; Cabrera et al., 2004; Roca et al., 1999).
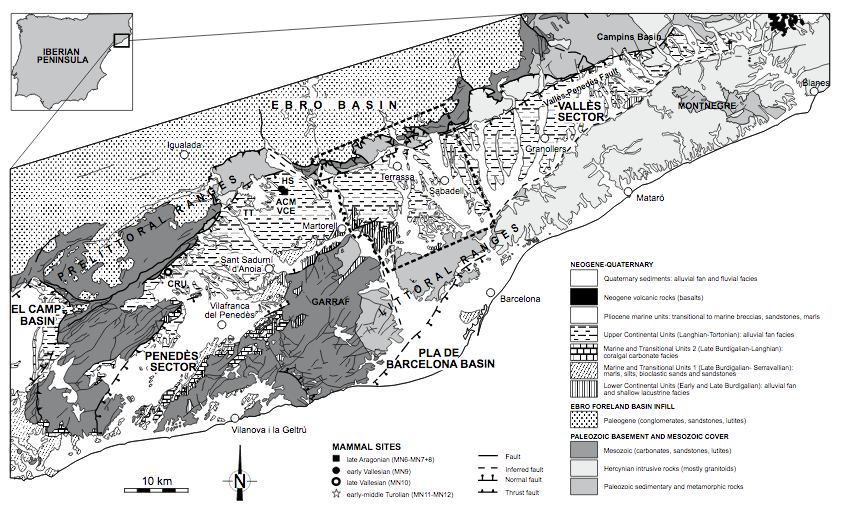
FIGURE 1
Simplified geological map of the Vallès-Penedès Basin with major Miocene mammal sites (modified from Institut Cartogràfic i Geològic de Catalunya, 2015). The area enclosed by the dashed perimeter is shown in detail in Figure 2. ACM: Abocador de Can Mata; CRU: Can Cruset; HS: Hostalets Superior; TT: Torrentet de Traginers; VCE: Variant de la Carretera de l’Ecoparc.
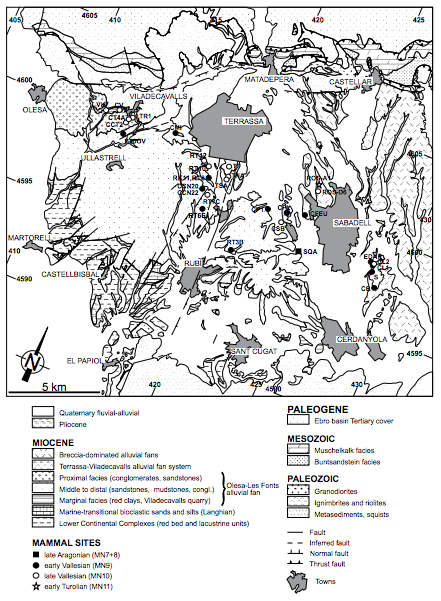
FIGURE 2
Detailed geological map of the western Vallès sector (see Fig. 1) showing the position of the main mammal sites (modified after Garcés et al., 1996). For locality acronyms see Electronic Appendix I, available at www.geologica-acta.com.
Most of the Miocene record consists of continental sediments, predominantly alluvial red mudstones, sandstones and conglomerates, sourced from the surrounding reliefs. During the early Miocene (Ramblianearly Aragonian; MN3-MN4), alluvial fans were generally small and their catchments were mostly located at the littoral range (Cabrera, 1981a, b; Cabrera et al., 1991, 2004; de Gibert and Casanovas-Vilar, 2011). Small lacustrine systems, which have yielded the richest mammal-bearing sites of this interval, developed in areas near the southeastern margin (Cabrera, 1981a, b; Cabrera et al., 1991; Casanovas-Vilar et al., 2011a, 2015). Subsequent deformation led to the partitioning of these older units into different faultbounded blocks (Cabrera, 1981a, b; Cabrera and Calvet, 1996). Prominent sea-level highstands during the MidMiocene Climatic Optimum (Zachos et al., 2001) favored a series of marine transgressions that affected the Vallès Penedès. Three different marine transgressions are known: a late Burdigalian, a Langhian and an early Serravallian one (Cabrera and Calvet, 1996; Cabrera et al., 1991; de Gibert and Casanovas-Vilar, 2011; Roca et al., 1999). The Langhian transgression was the most widespread, with shallow marine environments extending as far as the Vallès sector of the basin. Continental sedimentation was fully reestablished after the early Serravallian (late Aragonian), lasting until the Tortonian (middle Turolian). The Vallès Penedès master fault remained active until the Tortonian, controlling the development of large alluvial fan systems, some attaining a radius of 10–15km, sourced from the reliefs of the pre-littoral range (Cabrera and Calvet, 1996; Garcés et al., 1996; Roca et al., 1999; Casanovas-Vilar et al., 2008; de Gibert and Casanovas-Vilar, 2011). Most of the vertebrate fossil sites are located in distal to terminal fan, mudstone-dominated facies (Fig. 2). At the end of the Miocene, the dramatic sea level fall of the late Messinian interrupted the sedimentation in the basin (Cabrera and Calvet, 1996). Miocene deposits are unconformably overlain by Pliocene alluvial-fluvial and shallow marine sediments and Pleistocene-Holocene alluvial-colluvial units.
MATERIAL AND METHODS
Magnetostratigraphy
Magnetostratigraphic studies in the Vallès-Penedès basin first aimed to provide an independent chronostratigraphic frame for the rich paleobiological record. Early works focused on the Vallesian successions (Garcés et al., 1996; Agustí et al., 1997), and later on extended down into the late Aragonian (Moyà-Solà et al., 2009; Casanovas-Vilar et al., 2011b, 2015). The sequences studied for magnetostratigraphy include four main composite sections: Abocador de Can Mata, Les Fonts, Montagut and Viladecavalls, plus a few shorter ones (Fig. 3). Despite the diversity of lithologies and grain-size, all the sequences contain abundant mudstone beds, which were sampled with a density of 1–2m/site. The time resolution is approximately 10kyr, which is sufficient to rely on the completeness of the magnetostratigraphic record at the scale of chrons and subchrons of the GPTS.
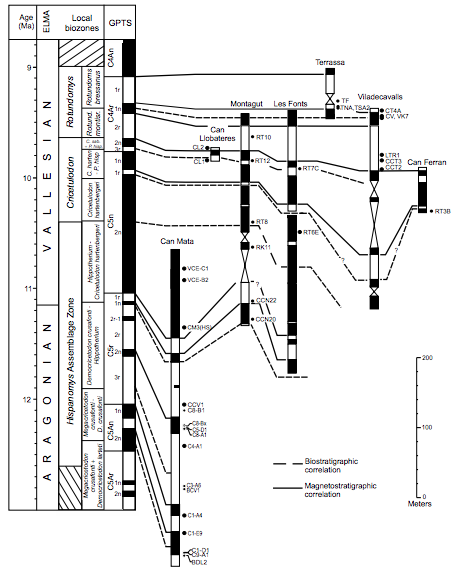
FIGURE 3
Correlation of the key local magnetostratigraphical sections of the Vallès-Penedès Basin to the Geomagnetic Polarity Time Scale (GPTS). The European Land Mammal Ages (ELMA) and local biozones of the Vallès-Penedès are also indicated. We have indicated some of the major sites in the Can Mata composite section. For locality acronyms see Electronic Appendix I. Neogene time scale and boundaries after Hilgen et al. (2012). GPTS after Ogg (2012).
A correlation of the local magnetostratigraphy with the GPTS (Ogg, 2012) was feasible after the unambiguous identification of chron C5n, a distinctly long normal chron that characterizes the early Tortonian, in both the Vallès sector (Garcés et al., 1996) and in the Abocador de Can Mata (ACM) sequence of els Hostalets de Pierola (Moyà-Solà et al., 2009). The overall magnetostratigraphic sequence represents approximately 3.6Myr, from 12.6Ma to 9.0Ma (Fig. 3). Here we summarize the magnetostratigraphic results of previous works (Garcés et al., 1996; Agustí et al., 1997; Moyà-Solà et al., 2009;Casanovas-Vilar et al., 2011b, 2015) and further update the results of the Abocador de Can Mata section with new magnetostratigraphical data from Ecoparc de Can Mata (Alba et al., 2012; CasanovasVilar et al., 2015), which extend the upper part of the section (Fig. 3)
Biostratigraphy
The Vallès-Penedès record comprises slightly more than 200 micromammal sites ranging from the late Aragonian to the Turolian. However, the distribution of the sites is clearly uneven, with most sites (>130) correlated to the latest Aragonian and early Vallesian (Fig. 4). By contrast, the Turolian is represented by only two sites. 70% of the sites within the studied time interval have associated magnetostratigraphic data, with the proportion increasing to 95% for the late Aragonian, the best sampled part of the record (Fig. 4). Regarding the Vallesian part of the record, magnetostratigraphic data are available for the most of the sites. However, the correlation of a significant number of sites, including some important ones such as Can Poncic 1 or Castell de Barberà, is based on biostratigraphy. Finally, the two Turolian sites known to date have no associated magnetostratigraphic data and have delivered a relatively poor rodent fauna that only allows a gross biostratigraphical correlation.
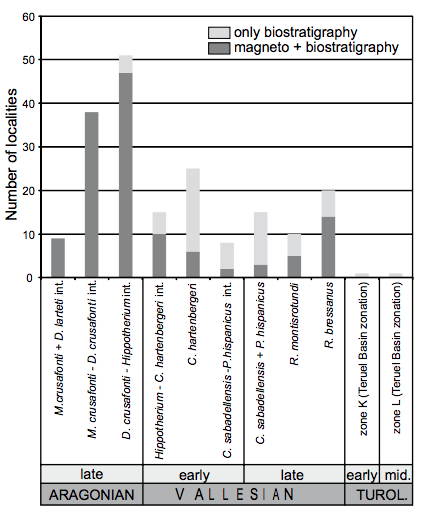
FIGURE 4
Number of rodent localities by local biozone. The number of localities with available bio- and magnetostratigraphical data is indicated with dark gray, localities dated only by biostratigraphy are indicated in light gray. Note the higher abundance of localities and greater dating accuracy for the late Aragonian (see text for details).
Rodents define the basis for the proposed local biozonation. Most of the material was recovered using systematic screen-washing (see Daams and Freudenthal, 1988a), except for a small part of the old collections of a few sites (Can Llobateres 1, Sant Quirze, Hostalets Superior) that were surface collected. Similarly, a few of the remains of certain large-sized rodents, such as castorids and some sciurids, were recovered during the systematic excavation of a number of sites (particularly Sant Quirze, Castell de Barberà and Can Llobateres 1). In Electronic Appendix I (available at https://www.geologica-acta.com) we provide a list of the sites included in this work, indicating their bio-and magnetostratigraphic correlation, and an updated faunal list of the rodent fauna. In most cases, the rodent fauna has not been studied in detail yet. In case systematic descriptions are available, the references are provided.
Abbreviations
ELMA: European Land Mammal Age; FCO: First Common Occurrence; FHA: First Historical Appearance (see Walsh, 1998); FLO: First Local Occurrence; FO: First Occurrence; GPTS: Geomagnetic Polarity Time Scale; LLO: Last Local Occurrence; LO: Last Occurrence; MN: Mammal Neogene units.
Locality acronyms
Locality names are mostly provided in full throughout this work, with a few exceptions: the composite stratigraphic section of Can Mata (Alba et al., 2006, 2011), including both the ACM and the ECM; B40OV (Alba et al., 2010); ROS; RT (Garcés et al., 1996; Agustí et al., 1997); and EDAR (Checa Soler and Rius Font, 2003). The abbreviations used are the following, ACM: local stratigraphic section of Abocador de Can Mata [Can Mata Landfill]; B40OV: Autovia Orbital de Barcelona [Orbital Highway of Barcelona], section Olesa de MontserratViladecavalls; BCV: Barranc de Can Vila [Can Vila Ravine] (ACM sector); BDL: Bassa de Lixiviats [Settling Pond of Leachates] (ACM sector); C1–C9, Cells 1 to 9 (ACM sectors); ECM: local stratigraphic section of Ecoparc de Can Mata [Waste Treatment Plant of Can Mata]; EDAR: Estació Depuradora d’Aigües Residuals Sabadell-Riu Ripoll [Wastewater Treatment Plant Sabadell-Ripoll River]; S5, sector 5 (B40OV sector); ROS: Ronda Oest de Sabadell [West Ring Road of Sabadell]; RT: Autopista de Rubí-Terrassa [Rubí-Terrassa Highway]; VCE: Variant de la Carretera de l’Ecoparc [Bypass of the Waste Treatment Plant Road].
BIOSTRATIGRAPHY: LOCAL BIOZONES OF THE LATE ARAGONIAN TO THE TUROLIAN
Here we propose a new local biostratigraphy ranging from the late Aragonian to the middle Turolian. A new local biozonation is introduced for the late Aragonian and Vallesian. Although our proposal clearly follows those of Agustí et al. (1997) and Casanovas-Vilar et al. (2011b), more clear-cut definitions are provided here, together with additional information on the small and large mammal fauna. Furthermore, the correlation of these biozones with other local zonations of Iberian basins is discussed. Concerning the Turolian, it is represented by too few sites to attempt a local biozonation. Therefore, the Turolian localities from the Vallès-Penedès are correlated to other biostratigraphical schemes.
The biozonation is mainly based on the composition of the rodent fauna. Rodents stand out as one of the best known mammal groups in the study area. Their dense and continuous record comprises almost 200 localities and more than 18,000 remains. In addition, certain species (e.g. the members of the Cricetulodon hartenbergeri – Rotundomys bressanus lineage) show very short ranges, allowing for high resolution during particular time intervals. The fact that most of the known sites have only delivered small mammals argues against the definition of a biostratigraphical scheme based on large mammals. Also, certain macromammal groups, such as perissodactyls and artiodactyls, are in need of taxonomic revision. Therefore, we will just briefly mention the main bioevents (such as FLO and LLO) in the large mammal faunas, but, with the exception of the FLO of the equid Hippotherium, they will not be used for the definition of the biozones.
Three different zones are recognized, which are further divided into several subzones. This is a major difference with previous works (Agustí et al., 1997; Casanovas-Vilar et al., 2011b), which did not consider such a hierarchy. This scheme is introduced for practical purposes, since the main zones allow a rough local correlation even if sample size is very limited (less than 15–20 specimens). On the other hand, the subzones provide a finer local correlation, although a greater sample size is usually required to identify the species that characterize each subzone. FLO and LLO data are used to characterize the different zones. Following Murphy and Salvador (1999), the zones are named after their diagnostic taxa. Their full name (including the kind of biostratigraphical unit) is given the first time they are introduced, whereas an abridged version is used afterwards. For a graphical summary of the local zonation and the main bioevents, see Figure 5. A comparison with previously proposed local zonations is povided in Figure 6.
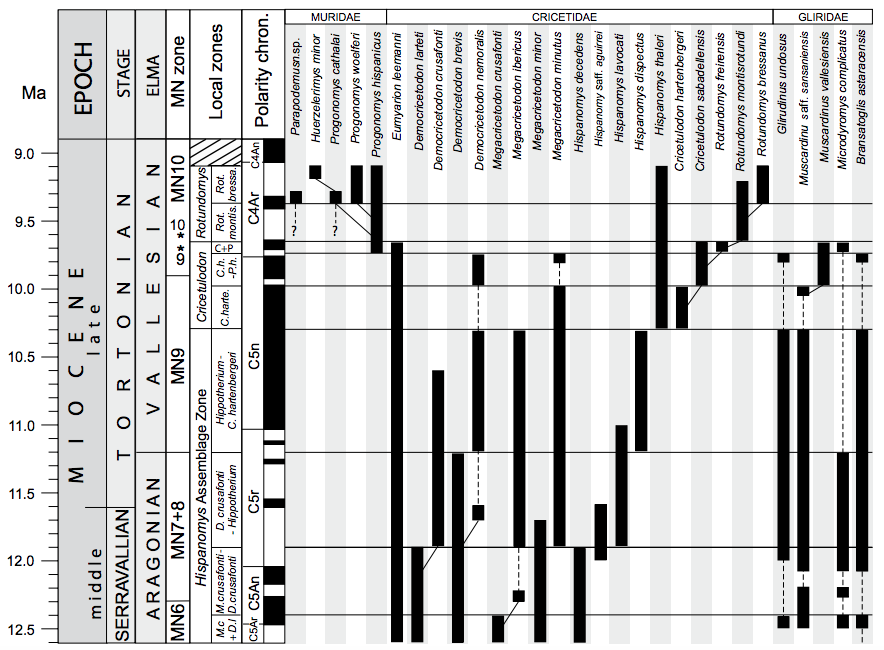
FIGURE 5
Local biozonation of the Vallès-Penedès Basin for the late Aragonian to Vallesian. The ranges of biostratigraphically important rodent taxa are indicated as well as the putative phylogenetic relationships of some lineages. Dashed lines indicate discontinuities in the ranges of those taxa. See main text for details. In the case of the MN (Mammal Neogene) zones, we indicated their boundaries as defined on the basis of selected appearance events (see Hilgen et al., 2012) as well as the chronological position of their reference sites (“faunal concept” of MN zones after De Bruijn et al., 1992; see text for details). Reference sites are: Sansan (France) for MN6, La Grive M (France) for MN7+8, Can Llobateres 1 (Spain) for MN9 and Masía del Barbo (Spain) for MN10. The estimated age of Sansan and La Grive is older than their respective MN zones so that they do not appear in the figure. These reference sites are indicated by an *. The age of the MN zone boundaries are after Hilgen et al. (2012) except for the MN6/MN7+8 boundary which is re-defined in this work (see text for details). Miocene time scale and boundaries after Hilgen et al. (2012). GPTS after Ogg (2012).
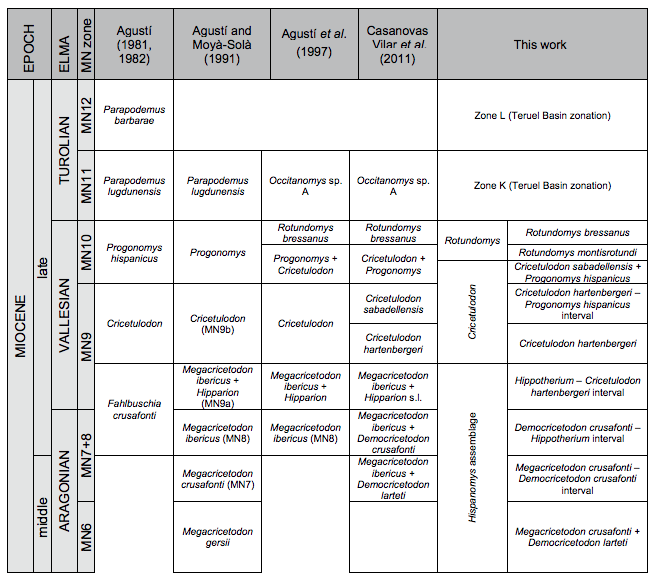
FIGURE 6
Comparison of the local biozones defined in this work with previous biozonations for the Vallès-Penedès Basin.
In this section we also propose correlations with the records of the Calatayud-Montalbán and Teruel basins (Daams et al., 1999; Álvarez-Sierra et al., 2003; Van Dam et al., 2001, 2014; García-Paredes et al., in press). These correlations are only introduced here and they are more thoroughly discussed in the next section. Finally, a correlation with the MN zones is also provided. Currently, two different concepts of the MN-zonation coexist (Van Dam et al., 2001; Van Dam, 2003; Hilgen et al., 2012). On the one hand MN zones can be defined using a strict biostratigraphic approach based on the FO of one or a few selected taxa (e.g. Steininger, 1999; Agustí et al., 2001; Hilgen et al., 2012). Alternatively, a more “faunal” approach can be used, defining the zones after the complete faunas of selected localities (De Bruijn et al., 1992). In this latter concept, the MN zones are viewed as an ordered sequence of reference localities. Note that this has important implications, since the reference localities for two MN zones may be very close in age (as for MN9 and MN10; see Figs. 5; 7), which may not be desirable. Even worse, the use of different concepts can lead to puzzling situations in which the reference locality for a given MN zone is correlated to a different MN zone as biostratigraphically defined. For example, Can Llobateres 1 (see Figs. 5; 7), the reference locality for MN9, has an estimated age of 9.76Ma (see Electronic Appendix I) which would imply a correlation with the MN10 as defined by the FCO of the murid Progonomys at 9.98Ma (Hilgen et al., 2012; Van Dam et al., 2014). Here we apply the biostratigraphical concept of the MN-zonation based on FHA events (in Europe; Hilgen et al., 2012), the ages of which may or may not correspond with the ages of first appearance events of Agustí et al. (2001) as defined in selected ‘boundary sections’. We acknowledge that first occurrences may be regionally diachronic (see Van der Meulen et al., 2011, 2012). Therefore, a MN zone boundary age as defined by a FHA event in one region may not automatically apply to other regions across Europe. However, in Figures 5; 7 we have also indicated the position of the reference sites for the different MN zones.
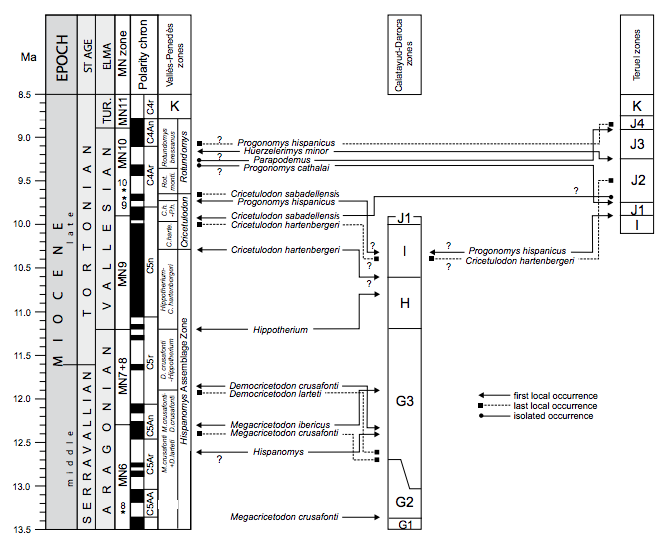
FIGURE 7
Comparison of the Vallès-Penedès local zonation with the local biostratigraphical schemes of the Calatayud-Montalbán and Teruel basins (east-central Spain). Hatching indicates hiatuses or insufficiently known faunas. The local zonation of the Calatayud-Daroca basin is taken from Daams et al. (1999), Álvarez-Sierra et al. (2003), Van Dam et al. (2014) and García-Paredes et al. (2016) whereas that of the Teruel basin from Van Dam et al. (2001, 2006). The first and last local occurrences are indicated for biostratigraphically important taxa, note important diachronies. Isolated occurrences indicate that the taxon has only been recovered from one site in that area. Question marks next to occurrence data indicate poorly constrained ages. For further details see text and Figure 5.
Hispanomys assemblage zone
Definition: The Cricetodontinae Democricetodon, Megacricetodon and Hispanomys are always present and stand out as the most common rodents in most of the sites. Democricetodon and Megacricetodon are usually represented in the sites by two coexisting species which differ markedly in size and morphology. Hispanomys is represented by different species with rather short ranges. The lower boundary would correspond to the FLO of the genus Hispanomys, whereas the upper boundary corresponds to the decrease in diversity and abundance of the Cricetodontinae that coincided with the FLO of the Cricetinae Cricetulodon.
Boundaries: The lower boundary is not recorded, but the genus Hispanomys is already present in the lowermost part of the ACM composite section, within chron C5Ar.1r. This would yield an estimated age of at least 12.6Ma for the lower boundary. The upper boundary would be placed at half-way within chron C5n.2n, with an estimated age of 10.3Ma.
Correlations: MN6 (late part) or MN7+8 – MN9 (early part).
Subzones: Four different subzones can be recognized on the basis of the coexistence of the different species of the cricetodontines Megacricetodon and Democricetodon as well as on the occurrence of the first hipparionin equids of the genus Hippotherium.
Megacricetodon crusafonti + Democricetodon larteti concurrent range subzone
Definition: The basis is defined by the FLO of Megacricetodon crusafonti. The top is defined by the LLO of Megacricetodon crusafonti.
Synonyms: Megacricetodon crusafonti + Megacricetodon gersii zone in Alba et al. (2006) and Casanovas-Vilar (2007).
Boundaries: The age for the lower boundary is not known, the upper boundary is placed within chron C5An.2n, with an estimated age of 12.39Ma.
Reference locality: ACM/C9-A1.
Reference section: ACM composite section (Moyà Solà et al., 2009).
Additional mammal data: Among the cricetids, FLO of Hispanomys decedens, which becomes a major component of the rodent assemblages. Eumyarion is relatively rare, whereas other cricetids, such as Megacricetodon minor and Democricetodon brevis, may be common in certain sites. Among the eomyids, FLO of Keramidomys. The glirids are relatively diverse, although they are scarce and their record is sparse. FLO of Paraglirulus werenfelsi and Myomimus (cf.) dehmi.
Other remarkable localities: ACM/BDL2, ACM/C1- D1.
Correlations: Because of the presence of Megacricetodon crusafonti, this unit can be correlated to MN6 sensu Mein (1999) and Agustí et al. (2001) or to MN7+8 sensu Hilgen et al. (2012). It is time-equivalent to the lowermost part of zone G3 of the Calatayud-Montalbán Basin (Daams et al., 1999; Álvarez-Sierra et al., 2003; Van Dam et al., 2014; García-Paredes et al., in press), although the defining criterion (presence of Megacricetodon crusafonti) fits better with the diagnosis of zone G2 (Daams et al., 1999; see discussion below).
Megacricetodon crusafonti – Democricetodon crusafonti interval subzone
Definition: The base is defined by the LLO of Megacricetodon crusafonti, the top is marked by the FLO of Democricetodon crusafonti.
Synonyms: Megacricetodon ibericus + Democricetodon larteti concurrent range zone in Alba et al. (2006), Casanovas-Vilar (2007) and Casanovas-Vilar et al. (2011b).
Boundaries: The lower boundary is placed within chron C5An.2n, with an estimated age of 12.39Ma. The upper boundary is placed within chron C5r.3r, with an estimated age of 11.88Ma.
Reference locality: ACM/BCV1.
Reference section: ACM composite section (Moyà Solà et al., 2009).
Additional mammal data: Amongst the cricetids, FLO of Megacricetodon ibericus, LLO of Hispanomys decedens, and FLO of Hispanomys aff. aguirrei. Democricetodon larteti and Eumyarion leemanni are the most common elements of rodent assemblages. Hispanomys may also be a common component. Amongst the eomyids, FLO of Eomyops. The glirid faunas are diverse, although they never constitute a major part of the rodent assemblage. FLO of castorids in the Vallès-Penedès (Chalicomys batalleri).
Other remarkable localities: ACM/C1-A4, ACM/C1- E9, ACM/C3-A6.
Correlations: This unit may covers the lower part of the MN7+8 (Mein, 1999; Agustí et al., 2001; Hilgen et al., 2012). It could be equivalent to the MN7 sensu Mein and Ginsburg (2002). It would also be equivalent to the middle part of zone G3 of the Calatayud-Montalbán Basin (Daams et al., 1999; Álvarez Sierra et al., 2003; Van Dam et al., 2014; García-Paredes et al. in press).
Democricetodon crusafonti – Hippotherium interval subzone
Definition: The base is defined by the FLO of Democricetodon crusafonti, the top is defined by the FLO of the equid Hippotherium.
Synonyms: Megacricetodon ibericus + Democricetodon crusafonti concurrent range zone in Alba et al. (2006), Casanovas-Vilar (2007) and Casanovas-Vilar et al. (2011b).
Boundaries: The lower boundary is placed within chron C5r.3r, with an estimated age of 11.88Ma. The upper boundary is placed within chron C5r.1n, with an estimated age of 11.18Ma.
Reference locality: ACM/C5-D1.
Reference section: ACM composite section (Moyà Solà et al., 2009).
Additional mammal data: Among the cricetids, FLO of Hispanomys lavocati, Megacricetodon minutus and Democricetodon nemoralis. Hispanomys is a major component of the rodent faunas together with Democricetodon crusafonti. Eumyarion leemanni may be very common. On the contrary, Megacricetodon ibericus is generally rare. FLO of Anomalomys gaudryi. A diverse glirid fauna is present, although the overall abundance of the family is low. FLO of Gliruluslissiensis. Similarly, the sciurids are diverse and include an important number of genera belonging to the Xerinae (Spermophilinus, Heteroxerus, Tamias) and the Sciurinae (Miopetaurista, Albanensia, Neopetes). FLO of the castorid Euroxenomys.
Other remarkable localities: ACM/C4-A1, ACM/C8- A1, ACM/C8-Bx, ACM/C8-B1, Sant Quirze A.
Correlations: This unit covers the upper part of the MN7+8 (Mein, 1999; Agustí et al., 2001; Hilgen et al., 2012) and may be equivalent to the MN8 sensu Mein and Ginsburg (2002). It can also be correlated to the upper part of zone G3 of the Calatayud-Montalbán Basin (Daams et al., 1999; Álvarez-Sierra et al., 2003; Van Dam et al., 2014; García-Paredes et al., in press).
Hippotherium – Cricetulodon hartenbergeri interval subzone
Definition: The base is defined by the FLO of the equid Hippotherium, the top is defined by the FLO of Cricetulodon hartenbergeri.
Synonymies: Megacricetodon ibericus + Hipparion zone in Agustí et al. (1997). Megacricetodon ibericus + Hipparion s.l. concurrent range zone in Casanovas-Vilar (2007) and Casanovas-Vilar et al. (2011b).
Boundaries: The lower boundary is placed within chron C5r.1n, with an estimated age of 11.18Ma. The upper boundary is located at half-way within chron C5n.2n, with an estimated age of 10.3Ma.
Reference locality: Creu Conill 20.
Reference section: Montagut composite section (Garcés et al., 1996).
Additional mammal data: The rodent fauna contains the same components as the previous biozone except for a few new occurrences. Megacricetodon ibericus is extremely common in certain sites. Eumyarion leemanni and different Hispanomys species may also be locally common. The cricetodontines record an important number of LLOs: Megacricetodon ibericus, Democricetodon crusafonti, Hispanomys lavocati and Hispanomys dispectus. Amongst the insectivores, FLO of the anourosoricine Crusafontina endemica. Regarding the macromammals, the FLO of Hippotherium at Creu Conill 20 (11.18Ma) is the most outstanding event. FLO of giraffids.
Other remarkable localities: Creu Conill 22, Hostalets de Pierola Superior (upper, Vallesian levels of els Hostalets de Pierola), ECM/VCE-C1 and, most likely, Can Missert and Castell de Barberà.
Correlations: This unit correlates to the lower part of the MN9 (Mein, 1999; Agustí et al., 2001; Hilgen et al., 2012). It is also time-equivalent to zone H of the Calatayud Montalbán Basin (Daams et al. 1999; Álvarez-Sierra et al., 2003; Van Dam et al., 2014; García-Paredes et al., in press). Initially, Daams and Freudenthal (1988a) defined zone H by the presence of Megacricetodon ibericus in the Calatayud-Montalbán Basin, which occurs much later there than in the Vallès-Penedès Basin (see discussion). Currently the FLO of Hispanomys nombrevillae, which has an estimated age of 11.20 (López-Guerrero et al., 2014), is used to define the lower boundary of zone H (Van Dam et al., 2014; García-Paredes et al., in press).
Cricetulodon range zone
Definition: Cricetulodon is the dominant rodent in all sites. The lower boundary is defined by the FLO of this genus and the upper boundary by its LLO.
Boundaries: The lower boundary is placed at half-way within chron C5n.2n, with an estimated age of 10.3Ma. The upper boundary is placed near the base of chron C4Ar.2r, with an estimated age of 9.65Ma.
Correlations: MN9 (late part) – MN10 (early part).
Subzones: Three different subzones can be recognized on the basis of the species of the genus Cricetulodon present and the occurrence of the earliest murids of the genus Progonomys.
Cricetulodon hartenbergeri range subzone
Definition: The base is defined by the FLO of Cricetulodon hartenbergeri and the top by its LLO.
Boundaries: The lower boundary is located at half-way within chron C5n.2n, with an estimated age of 10.3Ma. The upper boundary is placed near the base of chron C5n.1r (9.98Ma).
Reference locality: Can Poncic 1.
Reference section: Montagut composite section (Garcés et al., 1996).
Additional mammal data: The cricetid fauna is markedly less diverse than in the previous biozone. Cricetulodon hartenbergeri is the most abundant rodent. FLO of Hispanomys thaleri and Anomalomys gaillardi. Regarding the macromammals, LLO of Anchitherium at the locality of Can Poncic 1, where Hippotherium is also recorded (a co-occurrence of both equids is also observed in other sites from central Europe).
Other remarkable localities: B40OV/S5C, Santiga, EDAR-2, EDAR-6.
Correlations: This unit correlates to the upper part of the MN9 (Mein, 1999; Agustí et al., 2001; Hilgen et al., 2012). It would also cover all or part of zone I of the Calatayud-Montalbán and Teruel basins (Daams et al., 1999; Álvarez Sierra et al., 2003; Van Dam et al., 2001, 2006, 2014).
Cricetulodon hartenbergeri – Progonomys hispanicus interval subzone
Definition: The base is defined by the LLO of Cricetulodon hartenbergeri, while the top is defined by the FLO of Progonomys hispanicus.
Synonymies: Cricetulodon sabadellensis range zone in Casanovas-Vilar et al. (2011b).
Boundaries: The lower boundary is placed near the base of chron C5n.1r (9.98Ma) and the upper boundary is placed at half-way of chron C4Ar.3r, with an estimated age of 9.73Ma.
Reference locality: Can Llobateres 1.
Reference section: Montagutcomposite section (Garcés et al., 1996).
Additional mammal data: The cricetine Cricetulodon sabadellensis overwhelmingly dominates the rodent assemblages. Hispanomys thaleri may be relatively common in certain sites, although all other cricetodontines are very rare. Furthermore, this biozone records the LLO of the genera Megacricetodon (M. minutus) and Democricetodon (D. nemoralis), which had characterized the middle Miocene rodent faunas. Eumyarion leemanni may be common in certain sites. The glirid fauna is very diverse, although this family represents a small percentage of the rodent assemblages. Certain glirid genera show their LLO, notably Glirudinus and Bransatoglis. FLO of Muscardinus vallesiensis. Amongst the eomyids, FLO of Keramidomys pertesunatoi, which is restricted to this biozone.
Other remarkable localities: Can Feu 2, Can Pallars de Llobateres.
Correlations: Because of the absence of Progonomys, this unit should be correlated to the upper-most part of the MN9 (Mein, 1999; Agustí et al., 2001; Hilgen et al., 2012). However, it is time-equivalent to the earliest MN10 (Hilgen et al., 2012) because Progonomys is already recorded as a common element in the Calatayud-Montalbán Basin at 10.0–9.9Ma, while in the Vallès-Penedès Basin it does not occur until the following biozone (see discussion below). The C. hartenbergeri – P. hispanicus interval subzone would be approximately time-equivalent to subzone J1 in the Teruel (Van Dam et al., 2001, 2006) and CalatayudMontalbán basins (Van Dam et al., 2006, 2014; García Paredes et al., in press).
Cricetulodon sabadellensis + Progonomys hispanicus concurrent range subzone
Definition: The base is defined by the FLO of Progonomys hispanicus, while the top is defined by the LLO of Cricetulodon sabadellensis.
Boundaries: The lower boundary is placed within chron C4Ar.3r, with an estimated age of 9.73Ma. The upper boundary is placed near the base of chron C4Ar.2r, with an estimated age of 9.65Ma.
Reference locality: Trinxera de Can Llobateres 1.
Reference section: Montagut composite section (Garcés et al., 1996).
Additional mammal data: Cricetulodon sabadellensis overwhelmingly dominates the rodent faunas in certain sites. However, in a few localities this species is replaced by its putative descendant, Rotundomys freirensis, which also becomes the most abundant element in the rodent assemblages. Rotundomys freirensis is confined to this biozone. Eumyarion leemanni is very rare and its highest record is at the base of this biozone. The murids first appear at the base of this biozone and are represented by just one species, Progonomys hispanicus, which is moderately common in all sites. The glirid fauna is still diverse, although Microdyromys complicatus, Muscardinus hispanicus and Muscardinus vallesiensis have their last records within this biozone. The castorids and sciurids are rare, the latter family being represented solely by Spermophilinus bredai in most sites.
Other remarkable localities: Can Llobateres 2, Trinxera de Can Llobateres 0, RT- 11, RT-7C.
Correlations: This unit correlates to the early part of the MN10 (Mein, 1999; Agustí et al., 2001; Hilgen et al., 2012). It would also be time-equivalent to the lowermost part of subzone J2 of the Teruel Basin (Van Dam et al., 2001, 2006, in preparation). However, Progonomys hispanicus is the only taxon in common between both areas (see discussion).
Rotundomys abundance zone
Definition: Rotundomys is the dominant rodent in all sites. The lower boundary is defined by the FLO of Rotundomys montisrotundi, the upper boundary is defined by the LLO of Rotundomys bressanus.
Boundaries: The lower boundary is placed near the base of chron C4Ar.2r, with an estimated age of 9.65Ma. The upper boundary is poorly constrained at the Vallès-Penedès, but the uppermost record of Rotundomys bressanus is placed within chron C4Ar.1r, with an estimated age of 9.1Ma.
Correlations: MN10 (late part).
Subzones: Two different subzones can be recognized on the basis of the species of the genus Rotundomys present.
Rotundomys montisrotundi lineage subzone
Definition: The base is defined by the FLO of Rotundomys montisrotundi, the top is defined by the FLO of its putative descendant Rotundomys bressanus.
Boundaries: The lower boundary is placed near the base of chron C4Ar.2r (9.65Ma), the upper boundary is located half-way within chron C4Ar.1n, with an estimated age of 9.36Ma.
Reference locality: Camí de Can Tarumbot 2.
Reference section: Viladecavalls composite section (Garcés et al., 1996).
Additional mammal data: The rodent faunas are not well known. The cricetine Rotundomys montisrotundi is present at most sites, being one of the most common rodents. Progonomys hispanicus is the only murid species present.
Other remarkable localities: Can Casablanca, La Tarumba 1, Ceràmiques Viladecavalls, Sant Miquel de Toudell.
Correlations: This unit correlates to the early part of the MN10 (Mein, 1999; Agustí et al., 2001; Hilgen et al., 2012). It is also approximately time-equivalent to a large part of subzone J2 of the Teruel Basin (Van Dam et al., 2001, 2006. Nevertheless, Progonomys hispanicus is the only species in common between both areas, the genus Rotundomys not being recorded in the Teruel Basin (see discussion).
Rotundomys bressanus lineage subzone
Definition: The base is defined by the FLO of Rotundomys bressanus, and the top by the LLO of this taxon.
Boundaries: The lower boundary half-way within chron C4Ar.1n, with an estimated age of 9.36Ma. The upper boundary of this subzone is poorly constrained, but the uppermost occurrence of Rotundomys bressanus is within chron C4Ar.1r, which yields an estimated age of at least 9.1Ma. However, the upper boundary of this biozone is extended to the Vallesian/Turolian boundary for convenience. The age of such boundary is estimated at 8.9Ma (Hilgen et al., 2012).
Reference locality: Trinxera Nord Autopista.
Reference section: Terrassa composite section (Garcés et al., 1996).
Additional mammal data: Rotundomys bressanus is usually the major component of the rodent assemblages.
Rotundomys montisrotundi may be present at some sites, but it is very rare. Among the murids, FLO of Progonomys cathalai, Progonomys woelferi, Huerzelerimys minor and Parapodemus sp. nov. The eomyids, glirids, sciurids and castorids are rare. FLO of the glirid Muscardinus heintzi. A number of extinctions affecting the latter families appear to have taken place within this biozone, but this may reflect the lack of an adequate record in successive zones rather than a real pattern. FLO of the equid Cremohipparion mediterraneum.
Other remarkable localities: ROS-D6? (it may belong to the previous biozone), Trinxera Sud Autopista 2, Torrent de Febulines 3, Torrent de Febulines [classical site], Can Cruset. The fauna of the sites around Terrassa (Trinxera Nord Autopista, Trinxera Sud Autopista, Torrent de Febulines) is sometimes considered altogether in the literature and referred to as ‘Terrassa’ (Agustí et al., 1984; Morales et al., 1999).
Correlations: This unit correlates to the late part of the MN10 (Mein, 1999; Agustí et al., 2001; Hilgen et al., 2012). It is approximately time-equivalent to subzone J3 of the Teruel Basin (Van Dam et al., 2001, 2006). A biostratigraphical correlation with the Teruel zonation is not feasible because the genus Rotundomys is not recorded in this basin (see discussion).
Huerzelerimys vireti range zone (zone K of Teruel Basin)
The site ROS-A1 is correlated to the Huerzelerimys vireti range zone (zone K), defined in the Teruel Basin as the interval between the FLO and the LLO of the murid H. vireti (Van Dam et al., 2006). Zone K combined with zone J4 (Van Dam et al., 2006) correlate to the MN11 as stratigraphically defined (Mein, 1999; Agustí et al., 2001; Hilgen et al., 2012).
The faunas of this zone are insufficiently known and just include one site (ROS-A1) in the Vallès-Penedès Basin. The estimated age of ROS-A1 would then be between 8.8 and 7.6Ma (Agustí et al., 2001; Van Dam et al., 2006; Hilgen et al., 2012). The murid Huerzelerimys vireti is common. It records the FLO of the cricetine Kowalskia fahlbuschi, which is a major component of the rodent assemblages. The large mammal fauna is insufficiently known. There is a significant impoverishment of many groups, and the bovids and hipparionin equids dominate the herbivore assemblage.
Parapodemus barbarae – Stephanomys ramblensis interval zone (zone L of Teruel Basin)
The site Torrentet de Traginers, in the town of Piera, is tentatively correlated to Parapodemus barbarae – Stephanomys ramblensis interval zone (zone L) of the Teruel Basin (Van Dam et al., 2001, 2006), which would yield an estimated age between 7.6Ma and 7.4–6.8Ma (Agustí et al., 2001, 2006; Van Dam et al., 2001, 2006; Hilgen et al., 2012). This zone is considered to be equivalent to MN12 as stratigraphically defined (Mein, 1999; Agustí et al., 2001; Hilgen et al., 2012). Zone L is defined in the Teruel Basin as the interval between the first appearance of P. barbarae and that of S. ramblensis, but also covers the total range of the murids Occitanomys adroveri and Huerzelerimys turoliensis (Van Dam et al., 2001). The only known microfauna from the VallèsPenedès Basin (Torrentet dels Traginers) includes just one taxon, Occitanomys adroveri, which justifies the proposed correlation. However, this micromammal sample is scarce, comprising just 20 specimens, and clearly more data are needed.
The classical macromammal sites of Piera (Plana del Castell, Torrent del Gall Mullat, Finca Llopart, Teuleria de les Flandes) would also belong to this zone. The fauna of all these sites is usually considered altogether with that of Torrentet de Traginers and referred to in the literature as ‘Piera’ (see for example Agustí et al., 1985, 1997; Morales et al., 1999). As far as macromammals are concerned, this zone would record the FLO of the boselaphin bovid Tragoportax aff. gaudryi. The giraffid Birgerbohlinia, the suid Microstonyx and the equid Cremohipparion are very abundant. On the contrary, rhinocerotids and cervids are scarce and markedly less diverse than in preceding zones.
DISCUSSION AND CORRELATION WITH OTHER RECORDS
The proposed local biozonation differs from earlier attempts in providing a formal diagnosis and description of each zone, as well as clear boundaries and a reference locality and section. However, slightly modified concepts of previous biostratigraphic schemes are retained (see Fig. 6). In this section we compare our zonation with earlier works and further discuss its correlation with other records.
Regarding the late Aragonian, the proposed zonation is similar to that of Alba et al. (2006), Casanovas-Vilar (2007) and Casanovas-Vilar et al. (2011b). Alba et al. (2006) were the first to recognize that the sites of the lowermost part of the ACM series belonged to a distinct biozone and tentatively correlated it to part of the MN6. These authors introduced the Megacricetodon crusafonti + Megacricetodon gersii zone for this part of the record. However, this conclusion was precipitate because the micromammal sample at that time was very limited and even the attribution of the Megacricetodon remains to a particular species could be questioned. Subsequent studies concluded that those sites belonged to a distinct biozone that might be correlated to part of the MN6, but admitted that the material recovered was too scarce to adequately characterize it (Casanovas-Vilar, 2007; Casanovas-Vilar et al., 2011b). Additional sampling of the lower part of the ACM series has provided good samples for some sites (ACM/C9-A1, ACM/BDL2), allowing an unambiguous identification of the characteristic species. As a result, we propose the Megacricetodon crusafonti + Democricetodon larteti concurrent range subzone, which would be equivalent to the M. crusafonti + M. gersii zone of Alba et al. (2006). Megacricetodon gersii does not occur in the studied assemblages, since earlier citations correspond to small-sized specimens of M. crusafonti. The cricetid M. crusafonti was considered to be the more characteristic element of the MN6, but Aguilar (1980) showed that the material from the reference locality (Sansan, France) belonged to a different species, M. gersii, whereas M. crusafonti was apparently restricted to the Spanish sites. Daams and Freudenthal (1988b) and Daams et al. (1999) showed that, in the Calatayud-Montalbán Basin (northcentral Spain), M. gersii occurred in older levels than M. crusafonti. Therefore, Agustí et al. (2001) proposed a subdivision of the MN6 into MN6a and MN6b in Western Europe, each subunit being characterized by a different Megacricetodon species. This subdivision roughly corresponds to zones G1 and G2 of the Calatayud Montalbán Basin (Daams et al., 1999). However, Hilgen et al. (2012) recently evaluated the boundaries of the MN zones and proposed to define the lower boundary of MN6 by the FHA of M. gersii and the lower boundary of MN7+8 by the FHA of M. crusafonti. In accordance, zone G1, which solely includes M. gersii, would be correlated to the MN6, whereas zone G2, characterized by the presence of M. crusafonti, would be correlated to lower part of MN7+8. Therefore, the M. crusafonti + D. larteti subzone could either be correlated to the MN7+8 sensu Hilgen et al. (2012) or to the MN6b sensu Agustí et al. (2001). In any case, our magnetostratigraphic correlations (Figs. 3; 5; 7) indicate that the M. crusafonti + D. larteti zone of the Vallès-Penedès is time-equivalent to the lower-most part zone G3 of the Calatayud-Montalbán Basin. Zone G3 of the Calatayud-Montalbán Basin is defined by the presence of transitional assemblages between M. crusafonti and M. ibericus (Daams and Freudenthal, 1988b; Daams et al., 1999), with the latter unambiguously present in the youngest sites of this zone (Álvarez-Sierra et al., 2003; Van Dam et al., 2014). Therefore, zone G3 covers most of the MN7+8 in that area.
These M. crusafonti – M. ibericus transitional assemblages are not recognized in the Vallès-Penedès Basin. The medium-sized Megacricetodon, which is quite abundant in the lowermost part of the ACM series, perfectly fits the diagnosis of M. crusafonti. Megacricetodon ibericus is present (although sporadically) in the next biozone (M. crusafonti – D. crusafonti interval subzone; see below), whereas no transitional forms are recorded between the LLO of M. crusafonti and the FLO of M. ibericus. The definition of the Vallès-Penedès M. crusafonti + D. larteti subzone would fit better with that of zone G2 of the Calatayud-Montalbán Basin, even though it is timeequivalent to the lower part of zone G3. Therefore, we conclude that M. crusafonti persisted for a slightly longer time in the Vallès-Penedès.
Interestingly, the genus Hispanomys is already present in the lowermost levels of the M. crusafonti + D. larteti subzone, with an estimated age of ca. 12.6Ma. This age is near to that of the first record of this genus in the Calatayud-Montalbán Basin which is at 12.4Ma (López-Guerrero et al., 2014). Nevertheless, Hispanomys is represented by different species in these areas. In Calatayud-Montalbán, the oldest Hispanomys species present is Hispanomys cf. aguirrei (López-Guerrero et al., 2014) whereas in the Vallès-Penedès it is Hispanomys decedens, the same species that occurs in La Grive M, L5 and L7, France (López-Antoñanzas and Mein, 2011). An as yet undescribed form closely related to H. aguirrei occurs in younger late Aragonian sites in the Vallès-Penedès. The various Hispanomys species have proven their biostratigraphic value in the Calatayud-Montalbán Basin, where they could well serve as the basis for the subdivision of the long local zone G3 into shorter intervals (see López-Guerrero et al., 2014). In the Vallès-Penedès Hispanomys species could also be of great use in refining the diagnoses for some zones and their boundaries. However, this would first require a detailed taxonomic revision of the genus in this area. The rest of the late Aragonian is covered by the Megacricetodon crusafonti – Democricetodon crusafonti and the Democricetodon crusafonti – Hippotherium interval subzones. These are equivalent to previous zonations that prominently considered the occurrence of Megacricetodon ibericus as a defining criterion (Alba et al., 2006; Casanovas Vilar, 2007; Casanovas-Vilar et al., 2011b). Thus, the late Aragonian of the ACM series was divided into two zones taking into account the coexistence of M. ibericus with two different large-sized Democricetodon species, D. larteti in the older sites and D. crusafonti in the younger ones (Fig. 6). The choice of M. ibericus as a diagnostic criterion was influenced by previous zonations (Agustí and Moyà-Solà, 1991; Agustí et al., 1997) and by the local biostratigraphy of the relatively closely situated Calatayud-Montalbán Basin, where medium-sized species of Megacricetodon are the major components of the late Aragonian and earliest Vallesian rodent assemblages (Daams and Freudenthal, 1988a, b; Daams et al., 1999). This is not the case in most of the Vallès-Penedès sites, where these cricetids may be rare or even absent, particularly in the well-sampled ACM series. Therefore, the zones proposed here are based on the FLO and LLO of abundant taxa (D. larteti, D. crusafonti, Hippotherium). The M. crusafonti - D. crusafonti and D. crusafonti - Hippotherium interval subzones are timeequivalent to zone G3 of the Calatayud-Montalbán Basin (Daams et al., 1999). Nevertheless, in zone G3 D. larteti only occurs in the oldest sites of zone G3, its last record being around 12.6Ma (Van Dam et al., 2014), much earlier than in the Vallès-Penedès Basin (11.9). For most of zone G3, D. crusafonti is the only large-sized Democricetodon species present in the Calatayud-Montalbán Basin. On the other hand, during the first half of zone G3 transitional assemblages between M. crusafonti and M. ibericus occur in the Calatayud-Montalbán Basin, M. ibericus not being recorded until 11.9Ma (Van Dam et al., 2014), i.e. 0.4Myr later than in the Vallès-Penedès. Both local zonations do not match because of this diachrony in the FLOs and LLOs of the defining taxa, even though gaps in the record of the Calatayud-Montalbán Basin would allow for somewhat older ages for the FLO of M. ibericus. On the other hand, the proposed zonation for the late Aragonian can probably be extended to France, particularly to the fissure fillings of La Grive (for an updated review of the La Grive fauna, see Mein and Ginsburg, 2002). Some fissures, such as La Grive M, would correlate to the M. crusafonti – D. crusafonti interval subzone or even to the earlier M. crusafonti + D. larteti subzone, being characterized by the presence of D. larteti and Hispanomys decedens, as well as by the absence of some faunal elements typical of the subsequent zone such as D. crusafonti and M. ibericus. Other fissure fillings, such as La Grive L3 and L5 would correlate to the D. crusafonti – Hippotherium interval subzone (Casanovas-Vilar, 2007; Casanovas-Vilar et al., 2008, 2011b).
Concerning the correlation of the late Aragonian zones of the Vallès-Penedès with the MN system, we follow the taxonomic choices of Agustí et al. (2001) for the definition of the boundaries of MN6 and MN7+8, because it allows for higher-resolution correlations for this time interval. We therefore propose to define the lower boundary of MN7+8 by the FHA of M. ibericus rather than based on the FHA of M. crusafonti (as proposed by Hilgen et al., 2012). As a consequence, our M. crusafonti + D. larteti subzone would correlate to MN6. The first record of M. ibericus occurs within chron C5An.2n in the ACM series, yielding an estimated age of around 12.3Ma. This would imply a slightly younger age for the MN6/MN7+8 boundary than previous estimates (between 13.1 and 12.5Ma, Agustí et al., 2001; Hilgen et al., 2012). The proposed age for the MN6/MN7+8 boundary is only slightly younger than the lower boundary for our M. crusafonti – D. crusafonti interval subzone (12.39Ma). Note that this definition of the MN7+8 can only be applied to southwestern Europe, where M. ibericus is present. The boundaries in other regions, such as the North Alpine Foreland Basin, are based on other taxa and place the MN6/MN7+8 boundary at considerably older ages, around 13.9Ma (Kälin and Kempf, 2009).
The beginning of the Vallesian is marked by the dispersal of the equid Hippotherium, but as far as the small mammal faunas are concerned, the earliest Vallesian assemblages do not differ from the latest Aragonian ones. In other words, the small mammal assemblages of the D. crusafonti – Hippotherium subzone cannot be distinguished from those of the earliest Vallesian. This had been already noted by previous authors, who introduced the M. ibericus + Hipparion(sensu lato) zone for the first half of the early Vallesian (Agustí and Moyà-Solà, 1991; Agustí et al., 1997; Casanovas-Vilar et al., 2011b). Later early Vallesian faunas are clearly distinguished by the presence of the cricetine genus Cricetulodon, which dominates the small mammal assemblages. In this work we define the Hippotherium – Cricetulodon hartenbergeri interval subzone for the earliest Vallesian, which would be equivalent to the M. ibericus + Hipparion zone. While Megacricetodon ibericus may be very common at certain early Vallesian sites (such as Creu Conill 22) it is extremely rare in others (such as in the ECM series), to the point that there is not enough material to confidently identify the species. Therefore, we decided to base the new local zonation on two taxa that are very common after their FLO: Hippotherium and Cricetulodon. This new definition implies that when only small mammals have been recovered we cannot distinguish the D. crusafonti – Hippotherium subzone from the Hippotherium – C. hartenbergeri subzone. Nevertheless, if the locality has associated high-quality magnetostratigraphical data, such as in the case of the ECM series, the sites can be confidently correlated to one of the subzones on the basis of the characteristic pattern of polarity reversals (Fig. 3). Although Hippotherium may be rare or even absent (as in the ECM sites), it is present in most of the sites that have delivered macromammals. In the Vallès-Penedès, Hippotherium is first recorded within chron C5r.1n (Creu Conill 20), with an estimated age of 11.18Ma (Garcés et al., 1996, 1997; dates recalculated taking into account the latest version of the GPTS after Ogg, 2012). This age is consistent with the record of the Calatayud-Montalbán Basin, where its first appearance could be placed between 11.2 and 10.8Ma (Garcés et al., 2003; Van Dam et al., 2014). It is also congruent with the data from Central Europe (Bernor et al., 1988; Woodburne, 2009), but it is somewhat older than age estimates for this event in other areas (Sinap Formation, Siwaliks), which range from 10.8 to 10.3Ma (Barry and Flynn, 1990; Barry et al., 1982, 1985, 2002; Bernor et al., 1996; Flynn et al., 2013; Kappelman et al., 2003). These age differences may reflect a slight diachrony in the dispersal of this taxon or may be due to a sampling effect; Hippotherium may have been initially very rare or linked to particular paleoenvironments that became more widespread after its first dispersal.
The correlation of Castell de Barberà and Can Missert sites with the early Vallesian is more controversial. The first accounts on these sites (Crusafont-Pairó and Golpe-Posse, 1974a, b) reported the presence of Hippotherium. Nevertheless, in subsequent publications (see Agustí et al., 1997, 2005; Casanovas-Vilar, 2007; Casanovas-Vilar et al., 2011b), these sites were correlated to the late Aragonian, because (at least in the case of Castell de Barberà), the scarce hipparionin material (see Rotgers and Alba, 2011) would have been collected from a younger level (although very close, less than ten meters higher stratigraphically). On the other hand, the stratigraphical proximity of this site to Can Llobateres 1 (see Crusafont Pairó and Truyols Santonja, 1951) certainly would argue against a late Aragonian age for Castell de Barberà. Here we assume that Castell de Barberà may well belong to the earliest Vallesian. Unfortunately, the small mammal faunas are of no help to solve this question. In the case of Can Missert, Crusafont-Pairó and Golpe-Posse (1974b) reported Hippotherium, although the presence of such material was not confirmed afterwards, resulting in a correlation to the latest Aragonian (see Agustí et al., 1997; 2005). However, Robles et al. (2011) reported a Hippotherium incisor from Can Poal, a nearby site that would be stratigraphically positioned below Can Missert. Therefore, we also favor a correlation of Can Missert to the early Vallesian.
The appearance of the cricetine Cricetulodon is also a remarkable Vallesian bioevent. This genus is first recorded in Turkey by the end of the middle Miocene (MN7+8; Ünay and de Bruijn, 1984; Koufos, 2003; de Bruijn et al., 2013) and dispersed into the Iberian Peninsula during the early Vallesian. The oldest European species known, C. hartenbergeri, is recorded within chron C5n.2n in the Vallès-Penedès Basin (RT8 site), yielding an estimated age of 10.3Ma. In the Calatayud-Montalbán Basin its first occurrence is also in chron C5n.2n in the Pedregueras section (Van Dam et al., 2014), with a proposed age of 10.6Ma (Pedregueras 2A site). In the Vallès-Penedès Basin, Cricetulodon is not recorded in the well-sampled sites of the ECM series, which range between 10.9 and 10.6Ma (Alba et al., 2012). On the other hand, the absolute chronology of the Pedregueras section is not direct, but largely depends on its lithostratigraphic correlation with the Nombrevilla section, for which magnetostratigraphic dating is available (Garcés et al., 2003; Van Dam et al., 2014). Thus, although Cricetulodon may occur earlier in the Calatayud-Montalbán Basin than in the Vallès-Penedès Basin, the uncertainty associated to this date allows for slightly younger ages than reported in the CalatayudMontalbán record.
The different subzones of the Cricetulodon and Rotundomys zones are slightly modified versions of the ones introduced by Casanovas-Vilar et al. (2011b) and provide a high resolution for the dating of the Vallesian sites. These zones consider the evolution of the C. hartenbergeri - R. bressanus lineage together with the appearance of certain murid taxa. Progonomys hispanicus is the earliest murid in the Vallès-Penedès, being recorded in RT-11 and RT-7C, which are correlated to the uppermost part of chron C4Ar.3r with an estimated age of 9.7Ma. The early evolution of the Muridae is well documented in the middle Miocene of Pakistan (Jacobs, 1977), where the genus Progonomys first appeared at around 12Ma (Barry and Flynn, 1990; Barry et al., 1982, 2002; Flynn et al., 1995, 2013; Wessels, 2009). Updated stratigraphic correlations attained in the Calatayud-Montalbán and Teruel basins (Van Dam et al., 2014) show a very rare presence of Progonomys during the early Vallesian, at 10.4Ma and a common occurrence of this form after 10.0-9.9Ma. In Turkey, its first record is at 10.1 Ma (Kappelman et al., 2003). In the Vallès-Penedès, Hartenberger and Thaler (1963) and Michaux (1971) reported a couple of murid molars from Can Llobateres 1 (kept at the University of Montpellier) which resemble the first Progonomys morphotypes in the Teruel Basin (assigned to Muridae incertae sedis by Mein et al., 1993). This would indicate a slightly older age for this bioevent in the VallèsPenedès, closer to 9.8Ma. Similarly, Agustí (1981) reported a single molar of a small-sized Rotundomys (which we assign to R. montisrotundi) from that site. The FLO of the genus Rotundomys (R. freirensis from RT-11 and RT-7B) coincides with that of Progonomys, whereas R. montisrotundi occurs slightly later (at around 9.65Ma). After the collection of thousands of small mammal remains in the successive campaigns undertaken at Can Llobateres 1 during the 1990s and 2010–2013, no further Rotundomys or murid remains have been recovered. This may indicate that the old collections include material from the upper levels of the Can Llobateres section, which are correlated to the late Vallesian, or from other sites of the basin.
Previous zonations for the late Vallesian (Agustí and Moyà-Solà, 1991; Agustí et al., 1997; Casanovas-Vilar et al., 2011b) were constructed in relation to the concept of the ‘Vallesian Crisis’. Therefore, the oldest zone of the late Vallesian was not only characterized by the concurrent range of Progonomys and Cricetulodon, but also by the abrupt extinction of a number of mammal taxa that presumably preferred humid and forested environments. Regarding the macromammals, the suids Listriodon and Parachleuastochoerus, the bovid Miotragocerus, amphicyonid and barbourofelid carnivorans and finally, hominids disappeared. Certainly, a number of extinctions seem to have affected the rodents, particularly the cricetids (Democricetodon, Megacricetodon, Eumyarion) and glirids (Glirudinus, Bransatoglis, Microdyromys). However, other alleged extinctions may in fact reflect a smaller sample size as compared to the early Vallesian, which is insufficient to record certain rare taxa (Casanovas-Vilar et al., 2014). Indeed, the above-mentioned taxa would ultimately disappear, but probably during a longer time interval comprising the whole late Vallesian and part of the Turolian (Casanovas-Vilar et al., 2014).
The biostratigraphic correlation of the Vallesian zones with other local biostratigraphic schemes is even less straightforward than in the case of the late Aragonian, given the little faunal overlap between the Vallès-Penedès and the Calatayud-Montalbán and Teruel basins during this time interval (Fig. 7). The Hippotherium – C. hartenbergeri zone is time-equivalent to zone H of the Calatayud-Montalbán Basin, which is defined by the FLO of Hispanomys nombrevillae in that area (Van Dam et al., 2014; García-Paredes et al., in press). On the other hand, the C. hartenbergeri subzone would cover zone I from the same area, even though in the Calatayud-Montalbán Basin, C. hartenbergeri is probably recorded in somewhat older sites (ranging from 10.6 to 10.4Ma) than in the Vallès-Penedès Basin (Van Dam et al., 2014). Cricetulodon sabadellensis characterizes the rest of the early Vallesian and the beginning of the late Vallesian (from 9.98 to 9.65Ma). On the other hand, successive species of Rotundomys are the basis for the definition of the late Vallesian local biozones of the Vallès-Penedès. In the Teruel Basin, C. hartenbergeri has a longer record than in the Calatayud-Montalbán Basin, extending into the late Vallesian to around 9.5Ma (subzone J2; Van Dam et al., 2001, 2006). Interestingly, one specimen resembling C. sabadellensis was recently identified in Puente Minero 2 (estimated age 9.73Ma; Van Dam et al., unpublished data), suggesting the presence of this taxon in the late Vallesian of the Teruel Basin as well. The Vallès-Penedès zonation for the early Vallesian can also be recognized in the Seu d’Urgell Basin in the Catalan Pyrenees (Casanovas-Vilar et al., 2011b).
Concerning the late Vallesian, the local zones of the Teruel Basin are based on the presence of different murid taxa that allow a subdivision of this time interval into subzones J1, J2 and J3 (Van Dam et al., 2001, 2006). This family overwhelmingly dominates the rodent faunas of the Teruel Basin, but, in the Vallès-Penedès the most common taxa are the cricetines Cricetulodon and Rotundomys. This has been interpreted as an evidence for differing environmental conditions in the two regions (Casanovas-Vilar and Agustí, 2007). In any case, using murids in the zonation of the Vallès-Penedès Basin complicates the correlations because, in this basin, they are scarce and may not be recorded at all in a given site if sample size is small. Therefore, we decided to base our zonation in more common taxa. Nevertheless, the murid succession between both areas shows some coincidences. In subzone J1 of the Teruel Basin, P. hispanicus is the only murid present. Therefore, the C. sabadellensis + P. hispanicus and the R. montisrotundi subzones of the Vallès-Penedès would be biostratigraphically equivalent to this subzone, even though P. hispanicus is recorded much earlier in inner Spain. In subzone J2, the murid assemblage is enriched with Progonomys cathalai, whereas Huerzelerimys minor defines the lower boundary of subzone J3 (Van Dam et al., 2001). In the Vallès-Penedès, P. cathalai is only recorded at ROS-D6, which also records a primitive Parapodemus. Interestingly, the first occurrence of the latter genus in the Vallès-Penedès is older than in Teruel, where Parapodemus lugdunensis is not recorded until subzone J4 (MN11, earliest Turolian; Van Dam et al. 2006). Concerning Huerzelerimys minor, in the VallèsPenedès it is only recorded in Can Cruset and Can Cruset 4B, which are correlated to the R. bressanus subzone. There are no magnetostratigraphic data associated with any of these sites, so that their age is not well constrained. ROS-D6 includes scarce remains of the cricetid R. montisrotundi, thus suggesting a correlation with the R. montisrotundi subzone. However, R. montisrotundi also persists (although it is rare) in the R. bressanus subzone, and in ROS-D6 there are recorded a number of taxa that are only known from the R. bressanus zone in the Vallès-Penedès, including Neocricetodon ambarrensis and Muscardinus heintzi (see Electronic Appendix I). Therefore, we favor a correlation of ROS-D6 with the R. bressanus zone, although an older correlation is also possible. However, we assume that the site is certainly older than Can Cruset and Can Cruset 4B so that the record of P. cathalai would precede that of its putative descendant H. minor in the Vallès-Penedès (Fig. 7).
The Turolian rodent record from the Vallès-Penedès is restricted to just two sites that have delivered markedly different faunas, ROS-A1 and Torrentet de Traginers, which can be tentatively correlated to the local zones K and L of the Teruel Basin, respectively (Van Dam et al., 2001, 2006). A better sampling of the Turolian part of the record will confirm these tentative correlations or rather provide the basis for a local zonation for that age in the Vallès-Penedès.
CONCLUSIONS
The detailed local biostratigraphy and magnetostratigraphy presented here allows an unprecedented dating accuracy for the late Aragonian and Vallesian of the Vallès-Penedès Basin. The rodent-based biozones permit the correlation of sites with no associated magnetostratigraphical data, even when the fossil sample size is very limited. On the other hand, a comparison with other records, such as those from the nearby Calatayud-Montalbán and Teruel basins, reveals that the order of the faunal events is roughly the same but that they are diachronic, with some events occurring earlier (i.e. first record of Megacricetodon ibericus and Parapodemus) or later (i.e. first record of Cricetulodon and Progonomys) in the Vallès-Penedès Basin. In addition, some taxa that are diagnostic of the Vallès-Penedès local zones, such as the various species of Rotundomys, are not recorded in these basins, thus hampering a direct biostratigraphic correlation. The faunal differences between the Vallès-Penedès Basin and those of east-central Spain are particularly strong during the late Vallesian, with markedly different rodent faunas occurring in both areas.
ELECTRONIC APPENDIX I: DATABASE OF RODENT SITES MENTIONED IN THIS WORK
CONTENTS
How to use this database III
Late Atagonian-Turolian rodent sites from the Vallès-Penedès Basin mentined in this work III
Abocador de Can Mata/Barranc de Can Vila 1 III
Abocador de Can Mata / Bassa de Lixiviats 2 III
Abocador de Can Mata / Cel·la 1-A4 IV
Abocador de Can Mata / Cel·la 1-D1 IV
Abocador de Can Mata / Cel·la 1-E9 IV
Abocador de Can Mata / Cel·la 3-A6 IV
Abocador de Can Mata / Cel·la 4-A1 IV
Abocador de Can Mata / Cel·la 5-D1 V
Abocador de Can Mata / Cel·la 8-A1 V
Abocador de Can Mata / Cel·la 8-B1 V
Abocador de Can Mata / Cel·la 8-Bx V
Abocador de Can Mata / Cel·la 9-A1 V
Autopista de Rubí-Terrassa 3B V
Autopista de Rubí-Terrassa 6E VI
Autopista de Rubí-Terrassa 7C VI
Autopista de Rubí-Terrassa 8 VI
Autopista de Rubí-Terrassa 10 VI
Autopista de Rubí-Terrassa 11 VI
Autopista de Rubí-Terrassa 12 VI
Autopista de Rubí-Terrassa km 11 VI
Autovia Orbital de Barcelona, tram Olesa de Montserrat-Viladecavalls /Sector 5 locality C VII
Cal Turu 4A VII
Camí de Can Tarumbot 2 VII
Camí de Can Tarumbot 3 VII
Can Casablanca VII
Can Cruset VII
Can Feu 2 VII
Can Llobateres 1 VIII
Can Llobateres 2 VIII
Can Missert VIII
Can Pallars de Llobateres IX
Can Poncic 1 IX
Castell de Barberà IX
Ceràmiques Viladecavalls IX
Creu Conill 20 IX
Creu Conill 22 X
Ecoparc de Can Mata / Variant de la Carretera de l’Ecoparc B2 X
Ecoparc de Can Mata / Variant de la Carretera de l’Eoparc C1 X
Estació Depuradora d’Aigües Residuals Sabadell − Riu Ripoll 2 X
Estació Depuradora d’Aigües Residuals Sabadell − Riu Ripoll 6 X Hostalets Superior X
La Tarumba 1 XI
Ronda Oest de Sabadell A1 XI
Ronda Oest de Sabadell D6 XI
Santiga XI
Sant Quirze A XI
Torrent de Febulines XII
Torrent de Febulines 3 XII
Torrentet dels Traginers XII
Trinxera de Can Llobateres 0 XII
Trinxera de Can Llobateres 1 XIII
Trinxera Nord Autopista XIII
Trinxera Sud Autopista 2 XIII
Viladecavalls km 7 XIII
REFERENCES
HOW TO USE THIS DATABASE
In this Appendix we provide an updated database of all the sites mentioned in the main text arranged in alphabetical order. Here the reader can find additional details regarding their location, age as well as an updated list of the rodent taxa. In case the rodent fauna has been described, the appropriate references are given, but this is rarely the case. When no references are given, the identifications of the rodent taxa should be taken as preliminary. 45 rodent sites from the Vallès-Penedès Basin ranging from the late Aragonian (MN7+8) to the middle Turolian (MN11) are included. The main criterion for their inclusion in this database is the richness of the rodent sample, which in almost all cases is well above 200 identifiable specimens. In addition, we have preferentially selected those sites with well constrained ages. The sites listed in this database comprise about one fourth of all the rodent sites known for this time interval. All the material is kept within the collections of the Institut Català de Paleontologia Miquel Crusafont (ICP) at Sabadell.
Many of these localities also include other macrovertebrates and microvertebrates besides rodents, but these have not been included in the faunal lists. For more complete, although not updated, faunal lists including all mammal orders the reader is referred to the synthetic papers by Agustí et al. (1985, 1997) and to the NOW database of fossil mammals (Fortelius, 2015).
For each record the information is arranged as follows:
Name of the locality [in bold]. In case of synonymy the name used in the collections of the ICP is given.
Acronym: Standard acronym used in maps, stratigraphical series, biostratigraphic charts and other figures.
Locality synonyms: Other names given to the locality, usually referring to older names used by some authors.
Municipality: Municipality to which the site belongs to.
Age: An absolute age can be estimated for the sites with associated magnetostratigraphical data using the methods of Barry et al. (2002). The calculations are based on the stratigraphic position of a given site in the local magnetostratigraphic series. The interpolation must be done between two points of known ages, namely polarity changes in the section that correspond to chron boundaries recorded in the same section. When one of the chron boundaries is not recorded in this section, then the absolute age (in Ma) can be calculated using the composite section which does record the chron boundaries.
Correlation to GPTS: Magnetostratigraphical correlation of the site to the Geomagnetic Polarity Time Scale. Ages for chron boundaries are after Ogg (2012).
Correlation to local biostratigraphy: Local subzone to which the site is correlated (see main text for details). Reference localities for these subzones are indicated.
Correlation to the MN zonation: MN zone to which the site is correlated. The diagnosis of MN zones used here is based in Mein (1999) and Agustí et al. (2001).
Faunal list: Rodent taxa present. Until formal descriptions of the specimens are published their identification should be considered preliminary.
Taxonomic references: Literature (including PhD thesis) where the rodent fauna of this site (or part of it) is described. In those works the reader can find further reference on the rest of the fauna recovered at a particular site.
Remarks: Additional remarks that usually refer to usual confusions or controversies regarding the name or the correlation of a particular site.
LATE ARAGONIAN-TUROLIAN RODENT SITES FROM THE VALLÈS-PENEDÈS BASIN MENTIONED IN THIS WORK
Abocador de Can Mata / Barranc de Can Vila 1
Acronym: ACM/BCV1
Locality synonyms: -
Municipality: els Hostalets de Pierola
Age: 11.97Ma Correlation to GPTS: C5r.3r
Correlation to local biostratigraphy: Megacricetodon crusafonti – Democricetodon crusafonti Interval Subzone [reference locality]
Correlation to the MN zonation: MN7+8
Faunal list: Spermophilinus bredai, Miopetaurista cf. crusafonti, Glirudinus undosus, Muscardinus aff. sansaniensis, Bransatoglis cf. astaracensis, Microdyromys complicatus, Eomyops sp., Eumyarion leemanni, Hispanomys aff. aguirrei, Megacricetodon minor, Democricetodon brevis, Democricetodon larteti.
Taxonomic references: Casanovas-Vilar (2007); Casanovas-Vilar et al. (2010).
Remarks: -
Abocador de Can Mata / Bassa de Lixiviats 2
Acronym: ACM/BDL2
Locality synonyms: -
Municipality: els Hostalets de Pierola
Age: 12.47Ma. Correlation to GPTS: C5Ar.1r
Correlation to local biostratigraphy: Megacricetodon crusafonti + Democricetodon larteti Concurrent Range Subzone
Correlation to the MN zonation: MN6
Faunal list: Spermophilinus bredai, Albanensia quiricensis, Myoglis cf. meini, Muscardinus aff. sansaniensis, Muscardinus cf. hispanicus, Bransatoglis astaracensis, Myomimus cf. dehmi, Eumyarion leemanni, Hispanomys decedens, Megacricetodon cf. minor, Democricetodon cf. brevis, Democricetodon larteti.
Taxonomic references: -
Remarks: -
Abocador de Can Mata / Cel·la 1-A4
Acronym: ACM/C1-A4
Locality synonyms: -
Municipality: els Hostalets de Pierola
Age: 12.15Ma.
Correlation to GPTS: C5An.1n
Correlation to local biostratigraphy: Megacricetodon crusafonti + Democricetodon crusafonti Interval Subzone
Correlation to the MN zonation: MN7+8
Faunal list: Spermophilinus bredai, Heteroxerus cf. rubricati, Miopetaurista cf. neogrivensis, Muscardinus thaleri, Eumyarion leemanni, Hispanomys decedens, Megacricetodon minor, Democricetodon cf. brevis, Democricetodon larteti.
Taxonomic references: -
Remarks: -
Abocador de Can Mata / Cel·la 1-D1
Acronym: ACM/C1-D1
Locality synonyms: -
Municipality: els Hostalets de Pierola Age: 12.45Ma.
Correlation to GPTS: C5An.2n
Correlation to local biostratigraphy: Megacricetodon crusafonti + Democricetodon larteti Concurrent Range Subzone
Correlation to the MN zonation: MN6
Faunal list: Spermophilinus bredai, Glirudinus undosus, Muscardinus thaleri, Bransatoglis astaracensis, Microdyromys complicatus, Eumyarion leemanni, Hispanomys decedens, Megacricetodon crusafonti, Democricetodon cf. brevis, Democricetodon larteti.
Taxonomic references: -
Remarks: -
Abocador de Can Mata / Cel·la 1-E9
Acronym: ACM/C1-E9
Locality synonyms: -
Municipality: els Hostalets de Pierola
Age: 12.30Ma.
Correlation to GPTS: C5An.2n
Correlation to local biostratigraphy: Megacricetodon crusafonti – Democricetodon crusafonti Interval Subzone
Correlation to the MN zonation: MN7+8
Faunal list: Hispanomys cf. decedens, Megacricetodon ibericus, Megacricetodon cf. minor, Democricetodon cf. larteti.
Taxonomic references: -
Remarks: -
Abocador de Can Mata / Cel·la 3-A6
Acronym: ACM/C3-A6
Locality synonyms: -
Municipality: els Hostalets de Pierola
Age: 11.95Ma.
Correlation to GPTS: C5r.3r
Correlation to local biostratigraphy: Megacricetodon crusafonti – Democricetodon crusafonti Interval Subzone
Correlation to the MN zonation: MN7+8
Faunal list: Spermophilinus bredai, Heteroxerus grivensis, Muscardinus hispanicus, Microdyromys complicatus, Hispanomys decedens, Megacricetodon cf. ibericus, Megacricetodon minor, Democricetodon larteti.
Taxonomic references: -
Remarks: -
Abocador de Can Mata / Cel·la 4-A1
Acronym: ACM/C4-A1
Locality synonyms: -
Municipality: els Hostalets de Pierola
Age: 11.72Ma.
Correlation to GPTS: C5r.3r
Correlation to local biostratigraphy: Democricetodon crusafonti – Hippotherium Interval Subzone
Correlation to the MN zonation: MN7+8
Faunal list: Spermophilinus bredai, cf. Tamias sp., Albanensia quiricensis, Miopetaurista cf. neogrivensis, Neopetes cf. hoeckarum, Myoglis meini, Muscardinus aff. sansaniensis, Muscardinus hispanicus, Muscardinus thaleri, Bransatoglis astaracensis, Microdyromys complicatus, Eomyops cf. oppligeri, Keramidomys cf. carpathicus, Eumyarion leemanni, Hispanomys aff. aguirrei, Megacricetodon minor, Democricetodon sp.(small size), Democricetodon cf. crusafonti, Anomalomys gaudryi.
Taxonomic references: -
Remarks: -
Abocador de Can Mata / Cel·la 5-D1
Acronym: ACM/C5-D1
Locality synonyms: -
Municipality: els Hostalets de Pierola
Age: 11.64Ma. Correlation to GPTS: C5r.2n
Correlation to local biostratigraphy: Democricetodon crusafonti – Hippotherium Interval Subzone [reference locality]
Correlation to the MN zonation: MN7+8
Faunal list: Spermophilinus bredai, Albanensia quiricensis, Miopetaurista neogrivensis, cf. Neopetes sp., Muscardinus aff. sansaniensis, Muscardinus hispanicus, Bransatoglis astaracensis, Microdyromys complicatus, Paraglirulus werenfelsi, Myomimus dehmi, Eumyarion leemanni, Hispanomys aff. aguirrei, Megacricetodon cf. ibericus, Megacricetodon minor, Democricetodon brevis, Democricetodon crusafonti, Anomalomys gaudryi.
Taxonomic references: -
Remarks: -
Abocador de Can Mata / Cel·la 8-A1
Acronym: ACM/C8-A1
Locality synonyms: -
Municipality: els Hostalets de Pierola
Age: 11.65Ma. Correlation to GPTS: C5r.3r
Correlation to local biostratigraphy: Democricetodon crusafonti – Hippotherium Interval Subzone
Correlation to the MN zonation: MN7+8
Faunal list: Spermophilinus bredai, Albanensia cf. quiricensis, Miopetaurista neogrivensis, Myoglis meini, Muscardinus hispanicus, Eomyops cf. oppligeri, Eumyarion leemanni, Hispanomys aff. aguirrei, Democricetodon cf. nemoralis, Democricetodon crusafonti, Anomalomys cf. gaudryi.
Taxonomic references: -
Remarks: -
Abocador de Can Mata / Cel·la 8-B1
Acronym: ACM/C8-B1
Locality synonyms: -
Municipality: els Hostalets de Pierola
Age: 11.59Ma.
Correlation to GPTS: C5r.2r
Correlation to local biostratigraphy: Democricetodon crusafonti – Hippotherium Interval Subzone
Correlation to the MN zonation: MN7+8
Faunal list: Spermophilinus bredai, Myomimus dehmi, Hispanomys lavocati, Megacricetodon cf. minutus, Democricetodon cf. brevis, Democricetodon crusafonti.
Taxonomic references: -
Remarks: -
Abocador de Can Mata / Cel·la 8-Bx
Acronym: ACM/C8-Bx
Locality synonyms: -
Municipality: els Hostalets de Pierola
Age: 11.63Ma.
Correlation to GPTS: C5r.2n
Correlation to local biostratigraphy: Democricetodon crusafonti – Hippotherium Interval Subzone
Correlation to the MN zonation: MN7+8
Faunal list: Spermophilinus bredai, cf. Tamias sp., Microdyromys cf. koenigswaldi, cf. Eliomys sp., Myomimus dehmi, Eumyarion leemanni, Hispanomys aff. aguirrei, Megacricetodon cf. ibericus, Megacricetodon minutus, Democricetodon crusafonti, Anomalomys gaudryi.
Taxonomic references: -
Remarks: -
Abocador de Can Mata / Cel·la 9-A1
Acronym: ACM/C9-A1
Locality synonyms: -
Municipality: els Hostalets de Pierola
Age: 12.46Ma.
Correlation to GPTS: C5An.2n
Correlation to local biostratigraphy: Megacricetodon crusafonti + Democricetodon larteti Concurrent Range Subzone [reference locality]
Correlation to the MN zonation: MN6
Faunal list: Spermophilinus bredai, Albanensia cf. quiricensis, cf. Forsythia sp., Myoglismeini, Muscardinus aff. sansaniensis, Muscardinus thaleri, Bransatoglis astaracensis, Microdyromys complicatus, Paraglirulus werenfelsi, Eumyarion leemanni, Hispanomys decedens, Megacricetodon crusafonti, Megacricetodon minor, Democricetodon brevis, Democricetodon larteti.
Taxonomic references: -
Remarks: -
Autopista de Rubí-Terrassa 3B
Acronym: RT3B
Locality synonyms: -
Municipality: Sant Quirze del Vallès
Age: 10.02Ma.
Correlation to GPTS: C5n.2n
Correlation to local biostratigraphy: Cricetulodon hartenbergeri Range Subzone
Correlation to the MN zonation: MN9
Faunal list: Spermophilinus bredai, Euroxenomys minutus, Chalicomys jaegeri, Eumyarion leemanni, Megacricetodon cf. minutus, Cricetulodon hartenbergeri, Anomalomys gaillardi.
Taxonomic references: -
Remarks: -
Autopista de Rubí-Terrassa 6E
Acronym: RT6E
Locality synonyms: -
Municipality: Rubí
Age: 10.09Ma
Correlation to GPTS: C5n.2n
Correlation to local biostratigraphy: Cricetulodon hartenbergeri Range Subzone
Correlation to the MN zonation: MN9
Faunal list: Spermophilinus bredai, Muscardinus aff. sansaniensis, Myomimus dehmi, Eumyarion leemanni, Hispanomys cf. thaleri, Cricetulodon cf. hartenbergeri.
Taxonomic references: -
Remarks: -
Autopista de Rubí-Terrassa 7C
Acronym: RT7C
Locality synonyms: -
Municipality: Terrassa
Age: 9.73Ma
Correlation to GPTS: C4Ar.3r
Correlation to local biostratigraphy: Cricetulodon sabadellensis + Progonomys hispanicus Concurrent Range Subzone
Correlation to the MN zonation: MN10
Faunal list: Spermophilinus bredai, Albanensia aff. grimmi, Muscardinus hispanicus, Paraglirulus werenfelsi, Hispanomys cf. thaleri, Rotundomys freirensis, Progonomys hispanicus.
Taxonomic references: -
Remarks: -
Autopista de Rubí-Terrassa 8
Acronym: RT8
Locality synonyms: -
Municipality: Terrassa Age: 10.34Ma
Correlation to GPTS: C5n.2n
Correlation to local biostratigraphy: Cricetulodon hartenbergeri Range Subzone
Correlation to the MN zonation: MN9
Faunal list: Muscardinus hispanicus, Eumyarion leemanni, Megacricetodon cf. minutus, Cricetulodon cf. hartenbergeri.
Taxonomic references: -
Remarks: -
Autopista de Rubí-Terrassa 10
Acronym: RT10
Locality synonyms: -
Municipality: Terrassa
Age: 9.53Ma
Correlation to GPTS: C4Ar.2r
Correlation to local biostratigraphy: Rotundomys montisrotundi Lineage Subzone
Correlation to the MN zonation: MN10
Faunal list: Rotundomys cf. montisrotundi.
Taxonomic references: -
Remarks: -
Autopista de Rubí-Terrassa 11
Acronym: RT11
Locality synonyms: -
Municipality: Terrassa?
Age: -
Correlation to GPTS: -
Correlation to local biostratigraphy: Cricetulodon sabadellensis + Progonomys hispanicus Concurrent Range Subzone
Correlation to the MN zonation: MN10
Faunal list: Spermophilinus bredai, Myoglis meini, Muscardinus hispanicus, Microdyromys complicatus, Glirulus lissiensis, Hispanomys thaleri, Rotundomys freirensis, Progonomys hispanicus.
Taxonomic references: -
Remarks: The exact location of this site is unknown.
Autopista de Rubí-Terrassa 12
Acronym: RT12
Locality synonyms: -
Municipality: Terrassa
Age: 9.73Ma
Correlation to GPTS: C4Ar.3r
Correlation to local biostratigraphy: Cricetulodon sabadellensis + Progonomys hispanicus Concurrent Range Subzone
Correlation to the MN zonation: MN10
Faunal list: Muscardinus hispanicus, Rotundomys cf. freirensis.
Taxonomic references: -
Remarks: -
Autopista de Rubí-Terrassa km 11
Acronym: RK11
Locality synonyms: -
Municipality: Terrassa
Age: 10.98Ma
Correlation to GPTS: C5n.2n
Correlation to local biostratigraphy: Hippotherium – Cricetulodon hartenbergeri Interval Subzone
Correlation to the MN zonation: MN9
Faunal list: Myomimus dehmi, Hispanomys cf. dispectus, Megacricetodon cf. ibericus.
Taxonomic references: -
Remarks: -
Autovia Orbital de Barcelona, tram Olesa de MontserratViladecavalls /Sector 5 locality C
Acronym: B40/S5C
Locality synonyms: -
Municipality: Viladecavalls
Age: -
Correlation to GPTS: -
Correlation to local biostratigraphy: Cricetulodon hartenbergeri Range Subzone
Correlation to the MN zonation: MN9
Faunal list: Spermophilinus bredai, Myoglis cf. meini, Muscardinus hispanicus, Myomimus dehmi, Hispanomys thaleri, Megacricetodon cf. minutus, Cricetulodon hartenbergeri.
Taxonomic references: Pineda-Muñoz et al. (2011).
Remarks: -
Cal Turu 4A
Acronym: CT4A
Locality synonyms: -
Municipality: Viladecavalls
Age: 9.11Ma
Correlation to GPTS: C4Ar.1r
Correlation to local biostratigraphy: Rotundomys bressanus Lineage Subzone
Correlation to the MN zonation: MN10
Faunal list: Rotundomys bressanus.
Taxonomic references: -
Remarks: -
Camí de Can Tarumbot 2
Acronym: CCT2
Locality synonyms: -
Municipality: Viladecavalls
Age: 9.64Ma
Correlation to GPTS: C4Ar.2r
Correlation to local biostratigraphy: Rotundomys montisrotundi Lineage Subzone [reference locality]
Correlation to the MN zonation: MN10
Faunal list: Hispanomys cf. thaleri, Rotundomys montisrotundi, Progonomys hispanicus.
Taxonomic references: -
Remarks: -
Camí de Can Tarumbot 3
Acronym: CCT3
Locality synonyms: -
Municipality: Viladecavalls
Age: 9.59Ma
Correlation to GPTS: C4Ar.2r
Correlation to local biostratigraphy: Rotundomys montisrotundi Lineage Subzone
Correlation to the MN zonation: MN10
Faunal list: Rotundomys montisrotundi, Progonomys hispanicus.
Taxonomic references: -
Remarks: -
Can Casablanca
Acronym: CSB
Locality synonyms: Can Casablanques
Municipality: Sant Quirze del Vallès
Age: -
Correlation to GPTS: -
Correlation to local biostratigraphy: Rotundomys montisrotundi Lineage Subzone
Correlation to the MN zonation: MN10
Faunal list: Myoglis meini, Hispanomys thaleri, Rotundomys montisrotundi, Anomalomys cf. gaillardi, Progonomys cf. hispanicus.
Taxonomic references: Agustí (1981a).
Remarks: -
Can Cruset
Acronym: CRU
Municipality: Torroelles de Foix
Locality synonyms: -
Age: -
Correlation to GPTS: -
Correlation to local biostratigraphy: Rotundomys bressanus Lineage Subzone
Correlation to the MN zonation: MN10
Faunal list: Muscardinus cf. heintzi, Myomimus cf. dehmi, Rotundomys bressanus, Neocricetodon cf. ambarrensis, Progonomys hispanicus, Huerzelerimys minor.
Taxonomic references: -
Remarks: This site is the only late Vallesian site known from the Penedès sector of the Basin and is located near the Vallès-Penedès master fault.
Can Feu 2
Acronym: CFE2
Municipality: Sant Quirze del Vallès
Locality synonyms: -
Age: -
Correlation to GPTS: -
Correlation to local biostratigraphy: Cricetulodon hartenbergeri – Progonomys hispanicus Interval Subzone
Correlation to the MN zonation: MN9
Faunal list: Muscardinus hispanicus, Hispanomys cf. thaleri, Democricetodon cf. nemoralis, Cricetulodon sabadellensis.
Taxonomic references: Casanovas-Vilar et al. (2012).
Remarks: -
Can Llobateres 1
Acronym: CL1
Locality synonyms: Can Llobateres (classical site)
Municipality: Sabadell
Age: 9.76Ma
Correlation to GPTS: C4Ar.3r
Correlation to local biostratigraphy: Cricetulodon hartenbergeri – Progonomys hispanicus Interval Subzone [reference locality]
Correlation to the MN zonation: MN9 [reference locality]
Faunal list: Spermophilinus bredai, Heteroxerus rubricati, Albanensia aff. grimmi, Miopetaurista neogrivensis, Miopetaurista crusafonti, Blackia miocaenica, cf. Pliopetaurista sp., Euroxenomys minutus, Chalicomys jaegeri, Glirudinus cf. undosus, Myoglis meini, Muscardinus vallesiensis, Muscardinus hispanicus, Bransatoglis astaracensis, Paraglirulus werenfelsi, Glirulus lissiensis, Eomyops catalaunicus, Keramidomys pertesunatoi, Eumyarion leemanni, Hispanomys thaleri, Megacricetodon cf. minutus, Democricetodon cf. nemoralis, Cricetulodon sabadellensis, Anomalomys gaillardi.
Taxonomic references: Agustí (1981a); Aldana Carrasco (1991, 1992a, b, c); Casanovas-Vilar et al. (2015); Hartenberger (1965, 1966); Hartenberger and Thaler (1963); Mein (1970).
Remarks: The classic collection of this site includes a single molar of an undetermined murid (Hartenberger and Thaler, 1963; Michaux, 1971, Mein et al., 1993) and two molars of Rotundomys cf. montisrotundi (Agustí, 1981a). No further material of these taxa has been found among the thousands of rodent specimens recovered after successive field campaigns at Can Llobateres 1 during the late 1980s, early 1990s and from 2010 on. Therefore, we consider that these specimens from the old collections were recovered from different levels of the Can Llobateres section or even from different site of the basin. Accordingly, we have excluded them from the updated faunal list presented here.
Can Llobateres 2
Acronym: CL2
Locality synonyms: -
Municipality: Sabadell
Age: 9.62Ma
Correlation to GPTS: C4Ar.2r
Correlation to local biostratigraphy: Cricetulodon sabadellensis + Progonomys hispanicus Concurrent Range Subzone
Correlation to the MN zonation: MN10
Faunal list: Spermophilinus bredai, Heteroxerus cf. grivensis, Muscardinus hispanicus, Paraglirulus werenfelsi, Hispanomys cf. thaleri, Cricetulodon sabadellensis, Progonomys hispanicus.
Taxonomic references: Mein et al. (1993).
Remarks: Crusafont Pairó (1964) used the name ‘Can Llobateres 2’ for a small outcrop placed near the spring of Can Llobateres, on the side of the road opposite to Can Llobateres 1 and stratigraphically below it. Moyà-Solà and Köhler (1993, 1996) used the same name for a site closer to Can Llobateres 1 and located in the upper levels of the same section, which delivered a partial skeleton with cranium of the hominoid Hispanopithecus laietanus. The faunal list given here refers to the latter site and not to the one reported by Crusafont Pairó (1964) and includes the samples CL2C and CL2D. These samples presumably belong to very close stratigraphical levels.
Can Missert
Acronym: CMI
Locality synonyms: -
Municipality: Terrassa
Age: -
Correlation to GPTS: -
Correlation to local biostratigraphy: Hippotherium – Cricetulodon hartenbergeri Interval Subzone
Correlation to the MN zonation: MN9
Faunal list: Spermophilinus bredai, Muscardinus aff. sansaniensis, Muscardinus hispanicus, Paraglirulus werenfelsi, Eumyarion leemanni, Hispanomys daamsi, Megacricetodon cf. ibericus, Megacricetodon cf. minutus, Democricetodon nemoralis, Democricetodon cf. crusafonti.
Taxonomic references: Agustí et al. (2005); Aldana Carrasco (1991, 1992a).
Remarks: This locality was previously correlated to the late Aragonian (MN 7+8; Agustí et al., 1985, 1997, 2005). The correlation was based on scarce Hippotherium remains found in the nearby site of Can Poal, which is stratigraphically below Can Missert (Robles et al., 2011).
Can Pallars de Llobateres
Acronym: CPL
Locality synonyms: -
Municipality: Sant Quirze del Vallès
Age: -
Correlation to GPTS: -
Correlation to local biostratigraphy: Cricetulodon hartenbergeri – Progonomys hispanicus Interval Subzone
Correlation to the MN zonation: MN9
Faunal list: Spermophilinus cf. bredai, Heteroxerus cf. grivensis, Keramidomys pertesunatoi, Eumyarion leemanni, Hispanomys thaleri, Democricetodon cf. nemoralis, Cricetulodon sabadellensis.
Taxonomic references: -
Remarks: -
Can Poncic 1
Acronym: CP1
Locality synonyms: Can Poncic, Can Ponsic, Can Ponsich Municipality: Sant Quirze del Vallès
Age: -
Correlation to GPTS: -
Correlation to local biostratigraphy: Cricetulodon hartenbergeri Range Subzone [reference locality]
Correlation to the MN zonation: MN9
Faunal list: Spermophilinus bredai, Heteroxerus rubricati, Albanensia aff. grimmi, Miopetaurista crusafonti, Euroxenomys minutus, Chalicomys jaegeri, Myoglis meini, Muscardinus hispanicus, Myomimus dehmi, Eomyops cf. catalaunicus, Hispanomys thaleri, Megacricetodon minutus, Cricetulodon hartenbergeri.
Taxonomic references: Agustí (1981a, b); Aldana Carrasco (1991, 1992a, b, c); Hartenberger and Crusafont (1979).
Remarks: Golpe Posse (1974) introduced a second site near Can Poncic 1: Can Poncic 2. This site is approximately 6-10m below Can Poncic 1 in stratigraphic series and did not delivered Hippotherium or any giraffid remains, that according to Golpe Posse (1974) would indicate a late Aragonian age. However, given the short stratigraphic distance this correlation seems unlikely, and Can Poncic 2 would also have a Vallesian age. Unfortunately, the exact location of both sites is unknown, and detailed stratigraphical information is lacking.
Castell de Barberà
Acronym: CB
Locality synonyms: -
Municipality: Barberà del Vallès
Age: -
Correlation to GPTS: -
Correlation to local biostratigraphy: Hippotherium – Cricetulodon hartenbergeri Interval Subzone
Correlation to the MN zonation: MN9
Faunal list: Spermophilinus bredai, Albanensia quiricensis, Miopetaurista neogrivensis, Euroxenomys minutus, Chalicomys jaegeri, Glirudinus undosus, Myoglis meini, Muscardinus aff. sansaniensis, Muscardinus hispanicus, Bransatoglis astaracensis, Paraglirulus werenfelsi, Myomimusdehmi, Eumyarion leemanni, Hispanomysdaamsi, Megacricetodon minutus, Democricetodon nemoralis
Taxonomic references: Aguilar et al. (1979); Agustí (1981a, b); Aldana Carrasco (1991, 1992a, b, c).
Remarks: Despite some controversy (De Bruijn et al., 1992), this locality had been previously correlated to the late Aragonian (MN 7+8; Agustí et al., 1985, 1997; Casanovas-Vilar et al., 2011). This correlation was mostly based on scarce Hippotherium remains found few meters above the main fossiliferous horizon (Crusafont-Pairó and Golpe-Posse, 1974; Rotgers and Alba, 2011; see Alba and Moyà-Solà, 2012, for a more detailed discussion).
Ceràmiques Viladecavalls
Acronym: CV
Locality synonyms: -
Municipality: Viladecavalls
Age: 9.36Ma
Correlation to GPTS: C4Ar.1n
Correlation to local biostratigraphy: Rotundomys montisrotundi Lineage Subzone
Correlation to the MN zonation: MN10
Faunal list: Hispanomys cf. thaleri.
Taxonomic references: Agustí (1981a).
Remarks: The exact provenance of the material Progonomys hispanicus from Viladecavalls described by Schaub (1947b) is unknown, although it is generally assigned to this site.
Creu Conill 20
Acronym: CCN20
Locality synonyms: -
Municipality: Terrassa
Age: 11.18Ma
Correlation to GPTS: C5r.1n
Correlation to local biostratigraphy: Hippotherium – Cricetulodon hartenbergeri Interval Subzone [reference locality]
Correlation to the MN zonation: MN9
Faunal list: Euroxenomys minutus, Muscardinus hispanicus, Hispanomys dispectus, Megacricetodon ibericus.
Taxonomic references: Casanovas-Vilar et al. (2006).
Remarks: The large mammals have not been described, but a faunal list is given in Agustí et al. (1997) and Casanovas-Vilar et al. (2006).
Creu Conill 22
Acronym: CCN22
Locality synonyms: -
Municipality: Terrassa
Age: 11.13Ma
Correlation to GPTS: C5r.1r
Correlation to local biostratigraphy: Hippotherium – Cricetulodon hartenbergeri Interval Subzone
Correlation to the MN zonation: MN9
Faunal list: Spermophilinus bredai, Euroxenomys minutus, Myomimus dehmi, Hispanomys dispectus, Megacricetodon ibericus, Megacricetodon cf. minutus, Democricetodon cf. nemoralis.
Taxonomic references: Casanovas-Vilar et al. (2006).
Remarks: See corrections on the rodent faunal list in Casanovas-Vilar et al. (2007, p.76).
Ecoparc de Can Mata / Variant de la Carretera de l’Ecoparc B2
Acronym: ECM/VCE-B2
Locality synonyms: -
Municipality: els Hostalets de Pierola
Age: 10.60Ma
Correlation to GPTS: C5n.2n
Correlation to local biostratigraphy: Hippotherium – Cricetulodon hartenbergeri Interval Subzone
Correlation to the MN zonation: MN9
Faunal list: Spermophilinus bredai, Muscardinus hispanicus, Myomimusdehmi, Hispanomysaragonensis, Megacricetodon cf. ibericus, Democricetodon cf. nemoralis, Democricetodon cf. crusafonti.
Taxonomic references: -
Remarks: -
Ecoparc de Can Mata / Variant de la Carretera de l’Eoparc C1
Acronym: ECM/VCE-C1
Locality synonyms: -
Municipality: els Hostalets de Pierola
Age: 10.60Ma
Correlation to GPTS: C5n.2n
Correlation to local biostratigraphy: Hippotherium – Cricetulodon hartenbergeri Interval Subzone
Correlation to the MN zonation: MN9
Faunal list: Spermophilinus bredai, Heteroxerus rubricati, Muscardinus hispanicus, cf. Eliomys sp., Myomimus dehmi, Hispanomys cf. aragonensis, Megacricetodon minutus, Democricetodon nemoralis.
Taxonomic references: -
Remarks: -
Estació Depuradora d’Aigües Residuals Sabadell − Riu Ripoll 2
Acronym: EDAR2
Locality synonyms: -
Municipality: Sabadell
Age: -
Correlation to GPTS: -
Correlation to local biostratigraphy: Cricetulodon hartenbergeri Range Subzone
Correlation to the MN zonation: MN9
Faunal list: Glirudinus undosus, Myoglis meini, Paraglirulus werenfelsi, Eomyops catalaunicus, Eumyarion leemanni, Hispanomys cf. thaleri, Cricetulodon hartenbergeri, Anomalomys cf. gaillardi.
Taxonomic references: -
Remarks: -
Estació Depuradora d’Aigües Residuals Sabadell − Riu Ripoll 6
Acronym: EDAR6
Locality synonyms: -
Municipality: Sabadell
Age: -
Correlation to GPTS: -
Correlation to local biostratigraphy: Cricetulodon hartenbergeri Range Subzone
Correlation to the MN zonation: MN9
Faunal list: Spermophilinus bredai, Chalicomys jaegeri, Paraglirulus werenfelsi, Cricetulodon hartenbergeri.
Taxonomic references: -
Remarks: -
Hostalets Superior
Acronym: HS
Locality synonyms: see remarks.
Municipality: els Hostalets de Pierola
Age: -
Correlation to GPTS: -
Correlation to local biostratigraphy: Hippotherium – Cricetulodon hartenbergeri Interval Subzone
Correlation to the MN zonation: MN9
Faunal list: Spermophilinus bredai, Heteroxerus rubricati, Heteroxerus cf. grivensis, Chalicomys cf. jaegeri, Myomimus dehmi, Eumyarion cf. leemanni, Hispanomys lavocati, Hispanomys dispectus, Megacricetodon ibericus.
Taxonomic references: Agustí (1977a, 1980a, b, 1981a, b); Agustí and Gibert (1982a); Aldana Carrasco (1991, 1992a); Bataller Calatayud (1938); Hartenberger (1967); Schaub (1944, 1947a).
Remarks: The classical collections of small vertebrates labeled as “Hostalets Superior” include material from different sites from the upper (Vallesian) levels of els Hostalets de Pierola. As far as the micromammals are concerned, these comprise Can Mata 3, “bloc de marga”, Can Flaquer and Barranc del Mastodont. Unfortunately, the provenance of the material is not always known, since most rodent specimens are just labeled as “Hostalets Superior”. Most of the specimens for which the locality is known come from “bloc de marga” (“the marl block”; also mentioned in the literature as “bloc de marne”, its French translation), a fallen marl block extremely rich in microvertebrate remains that is already mentioned by Schaub (1947a), which apparently came from Can Flaquer (Villalta in Agustí, 1981a: 22; Agustí et al., 1985). In accordance, most of the small vertebrate collections labeled as “Hostalets Superior” would belong to this site.
La Tarumba 1
Acronym: LTR1
Locality synonyms: -
Municipality: Viladecavalls
Age: 9.57Ma
Correlation to GPTS: C4Ar.2r
Correlation to local biostratigraphy: Rotundomys montisrotundi Lineage Subzone
Correlation to the MN zonation: MN10
Faunal list: Spermophilinus bredai, Rotundomys montisrotundi.
Taxonomic references: -
Remarks: -
Ronda Oest de Sabadell A1
Acronym: ROS-A1
Locality synonyms: -
Municipality: Sabadell
Age: -
Correlation to GPTS: -
Correlation to local biostratigraphy: Huerzelerimys vireti Range Zone (zone K of the Teruel Basin)
Correlation to the MN zonation: MN11
Faunal list: Spermophilinus bredai, Hispanomys cf. peralensis, Neocricetodon fahlbuschi, Huerzelerimys vireti.
Taxonomic references: -
Remarks: -
Ronda Oest de Sabadell D6
Acronym: ROS-D6
Locality synonyms: -
Municipality: Sabadell
Age: -
Correlation to GPTS: -
Correlation to local biostratigraphy: Rotundomys bressanus Lineage Subzone?
Correlation to the MN zonation: MN10
Faunal list: Spermophilinus bredai, Muscardinus heintzi, Paraglirulus werenfelsi, Glirulus aff. lissiensis, Myomimus dehmi, Eomyops catalaunicus, Keramidomys mohleri, Rotundomys cf. montisrotundi, Neocricetodon ambarrensis, Progonomys cathalai, Parapodemus n. sp.
Taxonomic references: -
Remarks: The rodent fauna from ROS-D6 is significantly different from that of other late Vallesian sites of the basin and includes a number of taxa tha are only known from this site: Keramidomys mohleri, Progonomys cathalai and Parapodemus sp. interestingly, the fauna is dominated by murids. The genus Rotundomys, the usual major component, is so rare that even species identification is uncertain given that only two specimens have been recovered in a sample of 290 rodent cheek teeth. Finally, some rare taxa (Muscardinus heintzi, Neocricetodon ambarrensis, Eomyops catalaunicus) are relatively abundant in ROS-D6. The correlation to the R. bressanus subzone is uncertain because no remains of this taxon have been recovered, so that the locality may well belong to the previous R. montisrotundi subzone. However, the murid fauna is more diverse than in the R. montisrotundi subzone and the rodent assemblage includes M. heintzi and N. ambarrensis, which are only recorded in the R. bressanus subzone. Therefore, we favor this younger correlation.
Santiga
Acronym: SA
Locality synonyms: Santiga Provasa
Municipality: Barberà del Vallès
Age: -
Correlation to GPTS: -
Correlation to local biostratigraphy: Cricetulodon hartenbergeri Range Subzone
Correlation to the MN zonation: MN9
Faunal list: Spermophilinus cf. bredai, Euroxenomys minutus, Chalicomys jaegeri, Muscardinus hispanicus, Eomyops catalaunicus, Eumyarion leemanni, Cricetulodon hartenbergeri.
Taxonomic references: Agustí (1981a); Aldana Carrasco (1991, 1992b).
Remarks: -
Sant Quirze A
Acronym: SQA
Locality synonyms: -
Municipality: Sant Quirze del Vallès
Age: -
Correlation to GPTS: -
Correlation to local biostratigraphy: Democricetodon crusafonti – Hippotherium Interval Subzone
Correlation to the MN zonation: MN7+8
Faunal list: Spermophilinus bredai, Miopetaurista cf. crusafonti, Euroxenomys minutus, Chalicomys catalaunicus, Glirudinus undosus, Myoglis meini, Muscardinus aff. sansaniensis, Muscardinus hispanicus, Bransatoglis astaracensis, Microdyromys complicatus, Paraglirulus werenfelsi, Myomimus dehmi, Eomyops catalaunicus, Eumyarion leemanni, Hispanomys lavocati, Megacricetodon cf. ibericus, Megacricetodon minutus, Democricetodon brevis, Democricetodon crusafonti.
Taxonomic references: -
Remarks: This locality is close but not equivalent to the classical site of Sant Quirze (also known as Trinxera del Ferrocarril - Sant Quirze). Descriptions of the rodent fauna of the classical site can be found in Schaub (1944, 1947a), Crusafont Pairó et al. (1948), Freudenthal (1966), Villalta Comella (1950), Mein (1970), Agustí (1977a, b, 1978, 1980a, 1981a, b) and Aldana Carrasco (1991, 1992a, 1992b).
Torrent de Febulines
Acronym: TF
Locality synonyms: Terrassa (pro parte, see remarks)
Municipality: Terrassa
Age: 9.10Ma
Correlation to GPTS: C4Ar.1r
Correlation to local biostratigraphy: Rotundomys bressanus Lineage Subzone
Correlation to the MN zonation: MN10
Faunal list: Spermophilinus bredai, Chalicomys adroveri, Hispanomys thaleri, Rotundomys bressanus, Anomalomys gaillardi, Progonomys hispanicus, Progonomys woelferi, Muridae sp. 3.
Taxonomic references: Agustí (1981a, b); Agustí and Gibert (1982b); Aldana Carrasco (1991, 1992a, b); Casanovas-Vilar and Alba (2011).
Remarks: This site is placed in the Can Jofresa section near Terrassa. In the NOW database (Fortelius, 2015) the site known as ‘Terrassa’ includes a composite faunal list of the various sites of the Can Jofresa section. Such list includes the faunas of Trinxera Nord Autopista, Trinxera Sud Autopista and Torrent de Febulines, among other sites. Such confusion probably stems from Agustí et al. (1984), who published a composite faunal list of these sites under the name ‘Terrassa’. Later on, Morales et al. (1999) followed this study and gave a composite faunal list for the macromammals. ‘Terrassa’ is not a single locality, but includes around ten different sites from the Can Jofresa section, and therefore mixes material ranging from 9.3 to 9.1 Ma.
Torrent de Febulines 3
Acronym: TF3
Locality synonyms: Terrassa (pro parte, see remarks)
Municipality: Terrassa
Age: 9.10Ma
Correlation to GPTS: C4Ar.1r
Correlation to local biostratigraphy: Rotundomys bressanus Lineage Subzone
Correlation to the MN zonation: MN10
Faunal list: Spermophilinus bredai, Myoglis meini, Muscardinus heintzi, Paraglirulus werenfelsi, Eomyops catalaunicus, Hispanomys thaleri, Rotundomys bressanus, Anomalomys gaillardi, Progonomys hispanicus, Progonomys woelferi.
Taxonomic references: Agustí (1981a, b); Agustí and Gibert (1982b); Aldana Carrasco (1991, 1992a, b); Casanovas-Vilar and Alba (2011).
Remarks: See remarks for Torrent de Febulines.
Torrentet dels Traginers
Acronym: TT
Locality synonyms: Piera (sensu lato)
Municipality: Piera
Age: -
Correlation to GPTS: -
Correlation to local biostratigraphy: Parapodemus barbarae – Stephanomys ramblensis Interval Zone (zone L of the Teruel Basin)
Correlation to the MN zonation: MN12
Faunal list: Occitanomys adroveri.
Taxonomic references: -
Remarks: The fauna from the sites located in the town of Piera (Plana del Castell, Torrent del Gall Mullat, Finca Llopart, Teuleria de les Flandes and Torrentet dels Traginers) is usually considered altogether with that of Torrent de Traginers and referred to in the literature as ‘Piera’ (see for example Agustí et al., 1985, 1997). Nevertheless, it is likely that these sites have slightly different ages.
Trinxera de Can Llobateres 0
Acronym: TCL0
Locality synonyms: -
Municipality: Sabadell
Age: -
Correlation to GPTS: -
Correlation to local biostratigraphy: Cricetulodon sabadellensis + Progonomys hispanicus Concurrent Range Subzone
Correlation to the MN zonation: MN10
Faunal list: Heteroxerus cf. grivensis, Muscardinus vallesiensis, Muscardinus hispanicus, Microdyromys complicatus, Paraglirulus werenfelsi, Eumyarion leemanni, Hispanomys cf. thaleri, Cricetulodon sabadellensis, Rotundomys cf. freirensis, Progonomys hispanicus.
Taxonomic references: -
Remarks: -
Trinxera de Can Llobateres 1
Acronym: TCL1
Locality synonyms: -
Municipality: Sabadell
Age: -
Correlation to GPTS: -
Correlation to local biostratigraphy: Cricetulodon sabadellensis + Progonomys hispanicus Concurrent Range Subzone [reference locality]
Correlation to the MN zonation: MN10
Faunal list: Spermophilinus bredai, Heteroxerus grivensis, Muscardinus vallesiensis, Muscardinus hispanicus, Microdyromys complicatus, Eomyops catalaunicus, Eumyarion leemanni, Hispanomys thaleri, Cricetulodon sabadellensis, Progonomys hispanicus.
Taxonomic references: -
Remarks: -
Trinxera Nord Autopista
Acronym: TNA
Locality synonyms: Trinchera Norte Autopista, Terrassa (pro parte, see remarks)
Municipality: Terrassa
Age: 9.30Ma
Correlation to GPTS: C4Ar.1r
Correlation to local biostratigraphy: Rotundomys bressanus Lineage Subzone [reference locality]
Correlation to the MN zonation: MN10
Faunal list: Spermophilinus bredai, Muscardinus cf. heintzi, Hispanomys cf. thaleri, Rotundomys cf. montisrotundi, Rotundomys bressanus, Anomalomys gaillardi, Progonomys hispanicus, Progonomys woelferi.
Taxonomic references: Agustí (1981a, b); Agustí and Gibert (1982b); Aldana Carrasco (1991, 1992a).
Remarks: See remarks in Torrent de Febulines.
Trinxera Sud Autopista 2
Acronym: TSA2
Locality synonyms: Terrassa (pro parte, see remarks)
Municipality: Terrassa
Age: 9.30Ma
Correlation to GPTS: C4Ar.1r
Correlation to local biostratigraphy: Rotundomys bressanus Lineage Subzone
Correlation to the MN zonation: MN10
Faunal list: Spermophilinus bredai, Heteroxerus grivensis, Hispanomys thaleri, Rotundomys cf. montisrotundi, Rotundomys bressanus, Anomalomys gaillardi, Progonomys hispanicus, Progonomys woelferi.
Taxonomic references: Agustí (1981a, b); Agustí and Gibert (1982b); Aldana Carrasco (1991, 1992a).
Remarks: See remarks in Torrent de Febulines.
Viladecavalls km 7
Acronym: VK7
Locality synonyms: Can Trullàs 7
Municipality: Viladecavalls
Age: 9.36Ma
Correlation to GPTS: C4Ar.1n
Correlation to local biostratigraphy: Rotundomys montisrotundi Lineage Subzone
Correlation to the MN zonation: MN10
Faunal list: Rotundomys cf. montisrotundi, Progonomys cf. hispanicus.
Taxonomic references: Agustí (1981a).
Remarks: -
Agradecimientos
This work has been supported by the Spanish Ministerio de Economía y Competitividad (CGL2014-54373-P and RYC2013-12470 to ICV; CGL2014-55900-P), and the Agència de Gestió d’Ajuts Universitaris i de Recerca (2014 SGR 416) of the Generalitat de Catalunya. IG-P acknowledges a JAE-Doc contract (CSIC), co-funded by the European Social Fund, and the support of the Research group UCM-910607 “Evolution of Cenozoic Mammals and Continental Paleoenvironments”. We acknowledge J. Agustí, L. Cabrera and M. Llenas for their help in the geographical and stratigraphical location of several sites that provided material kept in the collections of the ICP. We sincerely thank the insightful comments by the two reviewers of an earlier version of this manuscript, P. Peláez-Campomanes and F.J. RuizSánchez, which surely helped to improve the final result.
REFERENCES
Aguilar, J.-P., 1980. Nouvelle interprétation de l’évolution du genre Megacricetodon au cours du Miocène. Palaeovertebrata, 9 Ext., 355-364.
Agustí, J., 1981. Roedores Miomorfos del Neógeno de Cataluña. PhD Thesis. Barcelona, Universitat de Barcelona, 288pp.
Agustí, J., 1982. Biozonación del Neógeno continental de Cataluña mediante roedores. Acta Geologica Hispanica, 17(1-2), 21-26.
Agustí, J., Moyà-Solà, S., 1991. Spanish Neogene Mammal succession and its bearing on continental biochronology. Neswletters on Stratigraphy, 25(2), 91-114.
Agustí, J., Moyà-Solà, S., Gibert, L., 1984. Mammal distribution dynamics in the Eastern margin of the Iberian Peninsula during the Miocene. Paléobiologie Continentale, 14(2), 33-46.
Agustí, J., Cabrera, L., Moyà-Solà, S., 1985. Sinopsis estratigráfica del Neógeno de la fosa del Vallès-Penedès. Paleontologia i Evolució, 18, 57-81.
Agustí, J., Cabrera, L., Garcés, M., Parés, J.M., 1997. The Vallesian mammal succession in the Vallès-Penedès basin (northeast Spain): paleomagnetic calibration and correlation with global events. Palaeogeography, Palaeoclimatology, Palaeoecology, 133(3-4), 149-180. DOI: https://doi.org/10.1016/S0031-0182(97)00084-9
Agustí, J., Cabrera, L., Garcés, M., Krijgsman, W., Oms, O., Parés, J.M., 2001. A calibrated mammal scale for the Neogene of Western Europe. State of the art. Earth Science Reviews, 52(4), 247-260. DOI: https://doi.org/10.1016/S0012-8252(00)00025-8
Agustí, J., Casanovas-Vilar, I., Furió, M., 2005. Rodents, insectivores and chiropterans (Mammalia) from the late Aragonian of Can Missert (Middle Miocene, Vallès-Penedès Basin, Spain). Geobios, 38(5), 575-583. DOI: https://doi.org/10.1016/j.geobios.2003.11.011
Agustí, J., Garcés, M., Krijgsman, W., 2006. Evidence for African-Iberian exchanges during the Messinian in the Spanish mammalian record. Palaeogeography, Palaeoclimatology, Palaeoecology, 238(1-4), 5-14. DOI: https://doi.org/10.1016/j.palaeo.2006.03.013
Alba, D.M., Moyà-Solà, S., Casanovas-Vilar, I., Galindo, J., Robles, J.M., Rotgers, C., Furió, M., Angelone, C., Köhler, M., Garcés, M., Cabrera, L., Almécija, S., Obradó, P., 2006. Los vertebrados fósiles del Abocador de Can Mata (els Hostalets de Pierola, l’Anoia, Cataluña), una sucesión de localidades del Aragoniense superior (MN6 y MN7+8) de la cuenca del Vallès-Penedès. Campañas 2002-2003, 2004 y 2005. Estudios Geológicos, 62(1), 295-312. DOI: https://doi.org/10.3989/egeol.0662127
Alba, D.M., Carmona, R., Bolet, A., Robles, J.M., Casanovas- Vilar, I., Furió, M., Sanisidro, Ó., Rivà, E., Balaguer, J., Checa, L., Luján, Á.H., Tomàs, M., Moyà-Solà, S., 2010. Intervención paleontológica en la Autovía Orbital de Barcelona B-40, tramo Olesa de Montserrat-Viladecavalls: Resultados preliminares. Cidaris, 30, 51-59.
Alba, D.M., Casanovas-Vilar, I., Robles, J.M., Moyà-Solà, S., 2011. Parada 3. El Aragoniense superior y la transición con el Vallesiense: Can Mata y la exposición paleontológica de els Hostalets de Pierola. Paleontologia i Evolució, Memòria Especial núm. 6, 95-109.
Alba, D.M., Carmona, R., Bertó Mengual, J.V., Casanovas- Vilar, I., Furió, M., Garcés, M., Galindo, J., Luján, A.H., 2012. Intervenció paleontològica a l’Ecoparc de Can Mata (els Hostalets de Pierola, conca del Vallès-Penedès). Tribuna d’Arqueologia, 2010–2011, 115-130.
Álvarez-Sierra, M.Á., Calvo, J.P., Morales, J., Alonso-Zarza, M.A., Azanza, B., García Paredes, I., Hernández Fernández, M., Van der Meulen, A.J., Peláez-Campomanes, P., Quiralte, V., Salesa, M.J., Sánchez, I.M., Soria, D., 2003. El tránsito Aragoniense- Vallesiense en el área de Daroca-Nombrevilla (Zaragoza, España). Coloquios de Paleontología, 1 (Volumen Extraordinario), 25-33.
Barry, J.C, Flynn, L.J., 1990. Key biostratigrapic events in the Siwalik sequence. In: Lindsay, E.H., Fahlbusch, V., Mein, P. (eds.). European Neogene mammal chronology. New York, Plenum Press, 557-571.
Barry, J.C., Lindsay, E.H., Jacobs, L.L., 1982. A biostratigraphic zonation of the middle and upper Siwaliks of the Potwar Plateau of northern Pakistan. Palaeogeography, Palaeoclimatoogy, Palaeoecology, 37, 95-130. DOI: https://doi.org/10.1016/0031-0182(82)90059-1
Barry, J.C., Johnson, N.M., Mahmood Raza, S., Jacobs, L.L. 1985. Neogene mammalian change in southern Asia: correlations with climatic, tectonic, and eustatic events. Geology, 13, 637-640. DOI: https://doi.org/10.1130/0091-7613(1985)13<637:NMFCIS>2.0.CO;2 (Error 14: El enlace externo https://doi.org/10.1130/0091-7613(1985)13<637:NMFCIS>2.0.CO;2 debe ser una URL) (Error 15: La URL https://doi.org/10.1130/0091-7613(1985)13<637:NMFCIS>2.0.CO;2 no esta bien escrita)
Barry, J.C., Morgan, M.E., Flynn, L.J., Pilbeam, D., Beherensmeyer, A.K., Mahmood Raza, S., Khan, I.A., Badgley, C., Hicks, J., Kelley, J., 2002. Faunal and environmental change in the Late Miocene Siwaliks of Northern Pakistan. Paleobiology, 28 (2, supplement), 1-71. DOI: http://dx.doi.org/10.1666/0094-8373(2002)28[1:FAECIT]2.0.CO;2
Bartrina M.T., Cabrera, L., Jurado, M.J., Guimerà, J., Roca, E., 1992. Evolution of the central Catalan margin of the Valencia trough (western Mediterranean). Tectonophysics, 203(1-4), 219-247. DOI: https://doi.org/10.1016/0040-1951(92)90225-U
Bernor, R.L., Kovar-Eder, J., Lipscomb, D., Rögl, F., Sen, S., Tobien, H., 1988. Systematic, stratigraphic and paleoenvironmental contexts of first appering Hipparionin the Vienna Basin, Austria. Journal of Vertebrate Paleontology, 8, 427-452. DOI: https://doi.org/10.1080/02724634.1988.10011729
Bernor, R.L., Koufos, G.D., Woodburne, M.O., Fortelius, M., 1996. The evolutionary history and biochronology of European and Southwest Asian Late Miocene and Pliocene Hipparionine horses. In: Bernor, R.L., Fahlbusch, V., Mittman, H.-W. (eds.). The Evolution of Western Eurasian Neogene Faunas. New York, Columbia University Press, 307-338.
Cabrera, L., 1981a. Estratigrafía y características sedimentológicas generales de las formaciones continentales de la cuenca del Vallès-Penedès (Barcelona, España). Estudios Geológicos, 37(1), 35-43.
Cabrera, L., 1981b. Influencia de la tectónica en la sedimentación continental de la cuenca del Vallès-Penedès (provincia de Barcelona, España) durante el Mioceno Inferior. Acta Geologica Hispanica, 16(3), 163-169.
Cabrera, L., Calvet, F., 1996. Onshore Neogene record in NE Spain: Vallès-Penedès and El Camp half grabens (NW Mediterranean). In: Friend, P.F., Dabrio, C.J. (eds.). Tertiary Basins of Spain the stratigraphic record of crustal kinematics. Cambridge, Cambridge University Press, 97-105.
Cabrera, L., Calvet, F., Guimerà, J., Permanyer, A., 1991. El registro sedimentario miocénico en los semigrabens del Vallès- Penedès y de El Camp: Organización secuencial y relaciones tectónica sedimentación. In: Colombo, F. (ed.). Libro Guía no. 4 del I Congreso Grupo Español del Terciario, Vic, 132pp.
Cabrera, L., Roca, E., Garcés, M., Porta, J. de, 2004. Estratigrafía y evolución tectonosedimentaria oligocena superior-neógena del sector central del margen catalán (Cadena Costero- Catalana). In: Vera, J.A. (ed.). Geología de España. Madrid, Sociedad Geológica de España y Instituto Geológico y Minero de España (SGE-IGME), 569-573.
Casanovas-Vilar, I., 2007. The rodent assemblages from the Late Aragonian and the Vallesian (Middle to Late Miocene) of the Vallès-Penedès Basin (Catalonia, Spain). PhD Thesis. Universitat Autònoma de Barcelona, 282pp.
Casanovas-Vilar, I., Agustí, J., 2007. Ecogeographical stability and climate forcing in the Late Miocene (Vallesian) rodent record of Spain. Palaeogeography, Palaeoclimatology, Palaeoecology, 248(1-2), 169-189. DOI: https://doi.org/10.1016/j.palaeo.2006.12.002
Casanovas-Vilar, I., Alba, D.M., Moyà-Solà, S., Galindo, J., Cabrera, L., Garcés, M., Furió, M., Robles, J.M., Köhler, M., Angelone, C., 2008. Biochronological, taphonomical and paleoenvironmental background of the fossil great ape Pierolapithecus catalaunicus (Primates, Hominidae). Journal of Human Evolution, 55(4), 589-603. DOI: https://doi.org/10.1016/j.jhevol.2008.05.004
Casanovas-Vilar, I., Alba, D.M., Robles, J.M., Moyà-Solà, S., 2011a. Registro paleontológico continental del Mioceno de la Cuenca del Vallès-Penedès. Paleontologia i Evolució, 6 (Memòria Especial), 55-80.
Casanovas-Vilar, I., Alba, D.M., Garcés, M., Robles, J.M., Moyà- Solà, S., 2011b. An updated chronology for the Miocene hominoid radiation in Western Eurasia. Proceedings of the National Academy of Sciences U.S.A., 108(14), 5554-5559. DOI: https://doi.org/10.1073/pnas.1018562108
Casanovas-Vilar, I., Van den Hoek Ostende, L.W., Furió, M., Madern, P.A., 2014. The range and extent of the Vallesian Crisis (Late Miocene): new prospects based on the micromammal record from the Vallès-Penedès basin (Catalonia, Spain). Journal of Iberian Geology, 40(1), 29-48. DOI: https://doi.org/10.5209/rev_JIGE.2014.v40.n1.44086
Casanovas-Vilar, I., Madern, A., Alba, D.M., Cabrera, L., García-Paredes, I., Van den Hoek Ostende, L.W., DeMiguel, D., Robles, J.M., Furió, M., Van Dam, J.A., Garcés, M., Angelone, C., Moyà-Solà, S., 2015. The Miocene mammal record of the Vallès-Penedès Basin (Catalonia). Comptes Rendus Palevol. DOI: https://doi.org/10.1016/j.crpv.2015.07.004
Checa Soler, L., Rius Font, L., 2003. Intervenció paleontològica a l’EDAR Sabadell-Riu Ripoll (Can Llobateres, Sabadell, Vallès Occidental). Tribuna d’Arqueologia, 1999-2000, 7-22.
Crusafont Pairó, M., 1950. La cuestión del llamado Meótico español. Arraona, 1-2, 1ª Època, 41-48.
Crusafont Pairó, M., Villalta Comella, J.F. de, 1947. Sobre la caracterización de los niveles meóticos en el Vallés. Museo de Sabadell, 3, 33-42.
Crusafont Pairó, M., Truyols Santonja, J., 1951. Hallazgo del Plesiodymylus chantrei Gaillard en el Meótico del Vallés. Notas y Comunicaciones del Instituto Geológico y Minero de España, 22, 97-126.
Crusafont Pairó, M., Truyols Santonja, J., 1960. Sobre la caracterización del Vallesiense. Notas y Comunicaciones del Instituto Geológico y Minero de España, 60, 109-126.
Crusafont-Pairó, M., Golpe-Posse, J.M., 1974a. Asociación de AnchitheriumMey., 1834, con Hipparion Christ, 1832, en el Alto Mioceno del Vallés. Boletín de la Real Sociedad Española de Historia Natural (Geología), 72(1-4), 75-93.
Crusafont-Pairó, M., Golpe-Posse, J.M., 1974b. Nuevos yacimientos del Terciario continental del N.E. de España. Acta Geológica Hispánica, 9(3), 81-83.
Daams, R., Freudenthal, M., 1988a. Synopsis of the Dutch- Spanish collaboration program in the Aragonian type area. Scripta Geologica, 1 (Special Issue), 3-18.
Daams, R., Freudenthal, M., 1988b. Cricetidae (Rodentia) from the type Aragonian; the genus Megacricetodon. Scripta Geologica, 1 (Special Issue), 39-132.
Daams, R., Van der Meulen, A.J., Álvarez Sierra, M.Á., Pélaez-Campomanes, P., Calvo, J.P., Alonso Zarza, M.A., Krijgsman, W., 1999. Stratigraphy and sedimentology of the Aragonian (Early to Middle Miocene) in its type area (North-Central Spain). Newsletters on Stratigraphy, 37(3), 103-139.
De Bruijn, H., Daams, R., Daxner-Höck, G., Fahlbusch, V., Ginsburg, L., Mein, P., Morales, J., 1992. Report of the RCMNS working group on fossil mammals, Reisensburg 1990. Newsletters on Stratigraphy, 26(2/3), 65-118.
De Bruijn, H., Ünay, E., Hordijk, K., 2013. A review of the Neogene succession of the Muridae and Dipodidae from Anatolia with special reference to taxa known from Asia and/or Europe. In: Wang, X., Flynn, L.J., Fortelius, M. (eds.). Fossil Mammals of Asia. Neogene Biostratigraphy and Chronology. New York, Columbia University Press, 564-580.
De Gibert, J.M., Casanovas-Vilar, I., 2011. Contexto geológico del Mioceno de la Cuenca del Vallès-Penedès. Paleontolologia i Evolució, 6 (Memòria Especial) 6, 39-45.
Eronen, J.T., Ataabadi, M.M., Micheels, A., Karme, A., Bernor, R.L., Fortelius, M., 2009. Distribution history and climatic controls of the Late Miocene Pikermian chronofauna. Proceedings of the National Academy of Sciences U.S.A., 106(29), 11867-11871. DOI: https://doi.org/10.1073/pnas.0902598106
Flynn, L.J., Barry, J.C., Morgan, M.E., Pilbeam, D., Jacobs, L.L., Lindsay, E.H., 1995. Neogene Siwalik mammalian lineages: species longevities, rates of change and mode of speciation. Palaeogeography, Palaeoclimatolology, Palaeoecology, 115(1-4), 249-264. DOI: https://doi.org/10.1016/0031-0182(94)00114-N
Flynn, L.J., Lindsay, E.H., Pilbeam, D., Mahmood Raza, S., Morgan, M.E., Barry, J.C., Badgley, C.E., Behrensmeyer, A.K., Cheema, I.U., Rahim Rajpar, A., Opdyke, N.D., 2013. The Siwaliks and Neogene evolutionary biology in South Asia. In: Wang, X., Flynn, L.J., Fortelius, M. (eds.). Fossil Mammals of Asia. Neogene Biostratigraphy and Chronology. New York, Columbia University Press, 353-372.
Garcés, M., Agustí, J., Cabrera, L., Parés, J.M., 1996. Magnetostratigraphy of the Vallesian (late Miocene) in the Vallès-Penedès Basin (northeast Spain). Earth and Planetary Science Letters, 142(3), 381-396. DOI: https://doi.org/10.1016/0012- 821X(96)00110-0
Garcés, M., Cabrera, L., Agustí, J., Parés, J.M., 1997. Old World first appearance datum of “Hipparion” horses: Late Miocene large mammal dispersals and global events. Geology, 25(1), 19-22. DOI: https://doi.org/10.1130/0091-7613(1997)025<0019:OWFADO>2.3.CO;2
Garcés, M., Krijgsman, W., Peláez-Campomanes, P., Álvarez Sierra, M.A., Daams, R., 2003. Hipparion dispersal in Europe: Magnetostratigraphic constraints from the Daroca area (Spain). Coloquios de Paleontología, 1 (Volumen Extraordinario), 171-178.
García-Paredes, I., Álvarez-Sierra, M.A., Van den Hoek Ostende, L.W., Hernández-Ballarín, V., Hordijk, K., López-Guerrero, P., Oliver, A., Pélaez-Campomanes, P., in press. The Aragonian and Vallesian high-resolution micromammal succession from the Calatayud-Montalbán Basin (Aragón, Spain). Comptes Rendus Palevol. DOI: https://doi.org/10.1016/j.crpv.2015.09.014.
Hartenberger, J.-L., Thaler, L., 1963. Sur les rongeurs fossiles du Vallésien (Miocène supérieur) de Can Llobateres (Sabadell, Espagne). Comptes Rendus de l’Académie des Sciences, Sciences de la Terre et des Planets, 256, 3333-3336.
Hilgen, F.J., Lourens, L.J., Van Dam, J.A., 2012. The Neogene period. In: Gradstein, F.M., Ogg, J.G., Schmitz, M., Ogg, G. (eds.). The Geologic Time Scale 2012. Amsterdam, Elsevier, 923-978.
Jacobs, L.J., 1977. A new genus of murid rodent from the Miocene of Pakistan and comments on the origin of the Muridae. PaleoBios, Contributions from the University of California Museum of Paleontology, 25, 1-11.
Kälin, D., Kempf, O., 2009. High-resolution stratigraphy from the continental record of the Middle Miocene Northern Alpine Foreland Basin of Switzerland. Neues Jahrbuch für Geologie un Paläontologie Abhandlungen, 254(1/2), 177-235. DOI: https://doi.org/10.1127/0077-7749/2009/0010
Kappelman, J., Duncam, A., Feseha, M., Lunkka, J.-P., Ekart, D., McDowell, F., Ryan, T.M., Swisher III, C.C., 2003. Chronology. In: Fortelius, M., Kappelman, J., Sen, S., Bernor, R.L. (eds.). Geology and paleontology of the Miocene Sinap Formation, Turkey. New York, Columbia University Press, 41-66.
Koufos, G.D., 2003. Late Miocene mammal events and biostratigraphy in the Eastern Mediterranean. Deinsea, 10, 343-372.
López-Antoñanzas, R., Mein, P., 2011. First detailed description of Hispanomys decedens (Rodentia) from the Middle Miocene of La Grive-Saint Alban (France). Swiss Journal of Geosciences, 104(2), 345-353. DOI: https://doi.org/10.1007/s00015-011-0066-0
López-Guerrero, P., García-Paredes, I., Álvarez-Sierra, M.A., Peláez-Campomanes, P., 2014. Cricetodontini from the Calatayud-Daroca Basin (Spain): A taxonomical description and update of their stratigraphical distributions. Comptes Rendus Palevol, 13(8), 647-664.
Mein, P., 1975. Résultats du Groupe de Travail des Vertébrés. In: Senes, J. (ed.). Report on Activity of the RCMNS Working Groups. Bratislava, 6th Congress of the Regional Committee of Mediterranean Neogene Stratigraphy, 1 (Proceedings), 78-81.
Mein, P., 1999. European Miocene mammal biochronology. In: Rössner, G.E., Heissig, K. (eds.). The Miocene Land Mammals of Europe. Munich, Verlag Dr. Friedrich Pfeil, 25-38.
Mein, P., Ginsburg, L., 2002. Sur l’âge relatif des différents dépôts karstiques miocènes de La Grive Saint-Alban (Isère). Cahiers scientifiques, 2/2002, 7-47.
Mein, P., Martín-Suárez, E., Agustí, J., 1993. ProgonomysSchaub, 1938 and Huerzelerimys gen. nov. (Rodentia); their evolution in Western Europe. Scripta Geologica, 103, 41-64.
Michaux, J., 1971. Muridae (Rodentia) néogènes d’Europe sud- occidentale. Évolution et rapports avec les formes actuelles. Paléobiologie Continentale, 2(1), 1-67.
Muridae (Rodentia) néogènes d’Europe sud- occidentale. Évolution et rapports avec les formes actuelles.Paléobiologie Continentale, 2(1), 1-67.
Morales, J., Nieto, M., Köhler, M., Moyà-Solà, S., 1999. Large mammals from the Vallesian of Spain. In: Agustí, J., Rook, L., Andrews, P. (eds.). Hominoid evolution and climatic change in Europe. The evolution of Neogene terrestrial ecosystems in Europe. Cambridge, Cambridge University Press, 1, 113-126.
Moyà-Solà, S., Köhler, M., Alba, D.M., Casanovas-Vilar, I., Galindo, J., Robles, J.M., Cabrera, L., Garcés, M., Almécija, S., Beamud, E., 2009. First partial face and upper dentition of the Middle Miocene hominoid Dryopithecus fontani from Abocador de Can Mata (Vallès-Penedès Basin, Catalonia, Spain): taxonomic and phylogenetic implications. American Journal of Physical Anthropology, 139, 126- 145. DOI: https://doi.org/10.1002/ajpa.20891
Murphy, M.A., Salvador, A. (eds.), 1999. International Stratigraphic Guide, An abridged version. Episodes, Newsmagazine of the International Union of Geological Sciences, 22(4), 255-271.
Ogg, J.G., 2012. Geomagnetic Polarity Time Scale. In: Gradstein, F.M., Ogg, J.G, Schmitz, M., Ogg, G. (eds.). The Geologic Time Scale 2012. Amsterdam, Elsevier, 85-113.
Pirlot, P.L., 1956. Les formes européennes du genre Hipparion. Memorias y Comunicaciones del Instituto Geológico, 14, 1-121.
Robles, J.M., Alba, D.M., Casanovas-Vilar, I., Galindo, J., Cabrera, L., Carmona, R., Moyà-Solà, S., 2011. On the age of the paleontological site of Can Missert (Terrassa, Vallès- Penedès, NE Iberian Peninsula). In: Pérez-García, A., Gascó, F., Gasulla, J.M., Escaso, F. (eds.). Viajando a mundos pretéritos. Morella, Ayuntamiento de Morella, 339-346.
Roca, A., Miranda, J., 2010. Atles Geológic de Catalunya. Barcelona, Institut Geològic de Catalunya - Institut Cartogràfic de Catalunya, 463 pp.
Roca, E., Guimerà, J., 1992. The Neogene structure of the eastern Iberian margin: structural constraints on the crustal evolution of the Valencia Trough (western Mediterranean). Tectonophysics, 203(1-4), 203-218. DOI: https://doi.org/10.1016/0040-1951(92)90224-T
Roca, E., Sans, M., Cabrera, L., Marzo, M., 1999. Oligocene to Middle Miocene evolution of the central Catalan margin (northwestern Mediterranean). Tectonophysics, 315(1-4), 209-233. DOI: https://doi.org/10.1016/S0040-1951(99)00289-9
Rotgers, C., Alba, D.M., 2011. The genus Anchitherium (Equidae: Anchitheriinae) in the Vallès-Penedès Basin (Catalonia, Spain). In: Pérez-García, A., Gascó, F., Gasulla, J.M., Escaso, F. (eds.). Viajando a mundos pretéritos. Morella, Ayuntamiento de Morella, 347-354.
Steininger, F., 1999. Chronostratigraphy, geochronology and biochronology of the Miocene “European Land Mammal Mega- Zones (ELMMZ)” and the Miocene “Mammal Zones (MN- Zones)”. In: Rössner, G.E., Heissig, K. (eds.). The Miocene Land Mammals of Europe. Munich, Verlag Dr. Friedrich Pfeil, 9-24.
Ünay, E., De Bruijn, H., 1984. On some Neogene rodent assemblages from both sides of the Dardanelles, Turkey. Newsletters on Stratigraphy, 13(3), 119-132.
Van Dam, J.A. 2003. European Neogene mammal chronology: past, present and future. DEINSEA, 10, 85-95.
Van Dam, J.A., Alcalá, L., Alonso Zarza, A., Calvo, J.P., Garcés, M., Krijgsman, W., 2001. The upper Miocene mammal record from the Teruel‒Alfambra region (Spain). The MN system and continental stage/age concepts discussed. Journal of Vertebrate Paleontology, 21(2), 367-385. DOI: https://doi.org/10.1671/0272-4634(2001)021[0367:TUMMRF]2.0.CO;2
Van Dam, J.A., Abdul Aziz, H., Álvarez-Sierra, M.Á., Hilgen, F.J., Van den Hoek Ostende, L.W., Lourens, L.J., Mein, P., Van der Meulen, A.J., Pélaez-Campomanes, P., 2006. Long- period astronomical forcing of mammal turnover. Nature, 443, 687-691. DOI: https://doi.org/10.1038/nature05163
Van Dam, J.A., Krijgsman, W., Abels, H.A., Álvarez-Sierra, M.Á., García-Paredes, I., López-Guerrero, P., Peláez- Campomanes, P., Ventra, D., 2014. Updated chronology for Middle to Late Miocene mammal sites of the Daroca area (Calatayud-Montalbán Basin, Spain). Geobios, 47(5), 325-334. DOI: https://doi.org/10.1016/j.geobios.2014.07.002
Van der Meulen, A.J., García-Paredes, I., Álvarez-Sierra, M.A., Van den Hoek Ostende, L.W., Hordijk, K., Oliver, A., López-Guerrero, P., Hernández-Ballarín, V., Peláez- Campomanes, P., 2011. Biostratigraphy or biochronology? Lessons from the Early and Middle Miocene small Mammal Events in Europe. Geobios, 44(2-3), 309-321. DOI: https://doi.org/10.1016/j.geobios.2010.11.004
Van der Meulen, A.J., García-Paredes, I., Álvarez-Sierra, M.A., Van den Hoek Ostende, L.W., Hordijk, K., Oliver, A., Peláez-Campomanes, P., 2012. Updated Aragonian biostratigraphy: small mammal distribution and its implications for the Miocene European Chronology. Geologica Acta, 10(2), 159-179. DOI: https://doi.org/10.1344/105.000001710
Villalta Comella, J.F. de, Crusafont Pairó, M., 1946. Les gisements de Mammifères du Néogène espagnol. III. Bassin du Vallés-Penedés (Catalogne). Pontien. Comptes Rendus Sommaires de la Societé Géologique de France, 1946 (11/12), 224-227.
Walsh, S.L. 1998. Fossil datum and paleobiological event terms, paleontostratigraphy, chronostratigraphy and the definition of Land Mammal ‘Age’ boundaries. Journal of Vertebrate Paleontology, 18(1), 150-179. DOI: https://doi.org/10.1080/02724634.1998.10011041
Wessels, W., 2009. Miocene rodent evolution and migration: Muroidea from Pakistan, Turkey and Northern Africa. Geologica Ultraiectina, 307, 1-290.
Woodburne, M.O., 2009. The early Vallesian vertebrates from Atzelsdorf (Late Miocene, Austria). 9. Hippotherium (Mammalia, Equidae). Annalen des Naturhistorischen Museums in Wien, 111A, 585−604.
Woodburne, M.O., Swisher, C.C., 1995. Land mammal high resolution goechronology, intercontinental overland dispersals, sea level, climate and vicariance. In: Berggren, W.A., Kent, D.V., Aubry, M.P., Hardenbol, J. (eds.). Geochronology, time scales and global stratigraphic correlations: Uni ed temporal framework and historical geology. Society of Sedimentary Geology (SEPM), 54 (special publication), 335-364.
Woodburne, M.O., MacFadden, B.J., Skinner, M.F., 1981. The North American “Hipparion” datum and implicatons for the Neogene of the Old World. Géobios, 14(4), 493-524. DOI: https://doi.org/10.1016/S0016-6995(81)80124-6
Zachos, J., Pagani, M., Sloan, L., Thomas, E., Billups, K., 2001. Trends, rhythms and aberrations in global climate 65 Ma to present. Science, 292(5517), 686-693. DOI: https://doi.org/10.1126/science.1059412
REFERENCES
Aguilar, J.-P., Agustí, J., Gibert, J., 1979. Rongeurs miocènes dans le Vallés-Penedés: Les Rongeurs de Castell de Barberá. Palaeovertebrata, 9(1), 17-31.
Agustí, J., 1977a. Contribución al conocimiento del género Hispanomys (Cricetidae, Rodentia, Mammalia) en la cuenca miocénica del Vallès-Penedès (Resumen). Butlletí Informatiu de l’Institut de Paleontologia de Sabadell, 1(9), 30-32.
Agustí, J., 1977b. Descripción de Cricetodon lavocati FREUD., 1966 de St. Quirze (Trinxera). Paleontologia i Evolució, 12, 65-67.
Agustí, J., 1978. Fahlbuschia crusafonti n. sp., cricétido nuevo del Astaraciense superior del Vallés-Penedés. Butlletí Informatiu de l’Institut de Paleontologia de Sabadell, 10(1-2), 63-68.
Agustí, J., 1980a. Sobre Megacricetodon ibericus (Schaub) (Rodentia, Mammalia) de Hostalets de Pierola (Penedès). Butlletí Informatiu de l’Institut de Paleontologia de Sabadell, 12(1-2), 35-39.
Agustí, J., 1980b. La asociación de Hispanomys y Cricetodon (Rodentia, Mammalia) en el Mioceno Superior del Vallès- Penedès (Cataluña, España). Acta Geologica Hispanica, 15(2), 51-60.
Agustí, J., 1981a. Roedores Miomorfos del Neógeno de Cataluña.PhD Thesis. Universitat de Barcelona, 288pp.
Agustí, J., 1981b. Roedores Miomorfos del Neógeno de Cataluña. Barcelona, Centre de Publicacions Intercanvi Científic i Extensió Universitària, 60pp.
Agustí, J., Gibert, J., 1982a. Roedores e insectívoros del Mioceno superior dels Hostalets de Pierola (Vallès-Penedès, Catalunya). Butlletí Informatiu de l’Institut de Paleontologia de Sabadell, 14(1-2), 19-37.
Agustí, J., Gibert, J., 1982b. Roedores e insectívoros (Mammalia) del Mioceno superior de Can Jofresa y Can Perallada (Vallès- Penedès, Cataluña). Paleontologia i Evolució, 17, 29-41.
Agustí, J., Moyà-Solà, S., Gibert, L., 1984. Mammal distribution dynamics in the Eastern margin of the Iberian Peninsula during the Miocene. Paléobiologie Continentale, 14(2), 33-46.
Agustí, J., Cabrera, L., Moyà-Solà, S., 1985. Sinopsis estratigráfica del Neógeno de la fosa del Vallès-Penedès. Paleontologia i Evolució, 18, 57-81.
Agustí, J., Cabrera, L., Garcés, M., Parés, J.M., 1997. The Vallesian mammal succession in the Vallès-Penedès basin (northeast Spain): paleomagnetic calibration and correlation with global events. Palaeogeography, Palaeoclimatology, Palaeoecology, 133(3-4), 149-180. DOI: https://doi.org/10.1016/S0031-0182(97)00084-9
Agustí, J., Cabrera, L., Garcés, M., Krijgsman, W., Oms, O., Parés, J.M., 2001. A calibrated mammal scale for the Neogene of Western Europe. State of the art. Earth Science Reviews, 52(4), 247-260. DOI: https://doi.org/10.1016/S0012-8252(00)00025-8
Agustí, J., Casanovas-Vilar, I., Furió, M., 2005. Rodents, insectivores and chiropterans (Mammalia) from the late Aragonian of Can Missert (Middle Miocene, Vallès-Penedès Basin, Spain). Geobios, 38(5), 575-583. DOI: https://doi.org/10.1016/j.geobios.2003.11.011
Alba, D.M., Moyà-Solà, S., 2012. A new pliopithecid genus (Primates: Pliopithecoidea) from Castell de Barberà (Vallès- Penedès Basin, Catalonia, Spain). American Journal of Physical Anthropology, 147(1), 88-112. DOI: https://doi.org/10.1002/ajpa.21630
Aldana Carrasco, E.J., 1991. Roedores esciuromorfos del Neógeno de Cataluña. Doctoral Thesis. Universitat Autònoma de Barcelona, 304pp.
Aldana Carrasco, E.J., 1992a. Los Sciurinae (Rodentia, Mammalia) del Mioceno de la Cuenca del Vallès-Penedès (Cataluña, España). Treballs del Museu de Geologia de Barcelona, 2, 69-97.
Aldana Carrasco, E.J., 1992b. Los Castoridae (Rodentia, Mammalia) del Neógeno de Cataluña (España). Treballs del Museu de Geologia de Barcelona, 2, 99-141.
Aldana Carrasco, E.J., 1992c. Los Esciuróperos del Mioceno de la Cuenca del Vallès-Penedès (Cataluña, España). Geogaceta, 11, 114-116.
Bataller Calatayud, J.R., 1938. Els Ratadors Fòssils de Catalunya. Barcelona, Impremta de la Casa d’Assistència President Macià, 64pp.
Casanovas-Vilar, I., 2007. The rodent assemblages from the Late Aragonian and the Vallesian (Middle to Late Miocene) of the Vallès-Penedès Basin (Catalonia, Spain). Doctoral Thesis. Universitat Autònoma de Barcelona, 282pp.
Casanovas-Vilar, I., Alba, D.M., 2011. The never-ending problem of Miocene beaver taxonomy. Acta Palaeontologica Polonica, 56(1), 217-220. DOI: https://doi.org/10.4202/app.2010.0051
Casanovas-Vilar, I., Furió, M., Agustí, J., 2006. Rodents insectivores and paleoenvironment associated to the first-appearing hipparionine horses in the Vallès-Penedès Basin (northeastern Spain). Beiträge zur Paläontologie, 30, 89-107.
Casanovas-Vilar, I., Angelone, C., Alba, D.M., Moyà-Solà, S., Köhler, M., Galindo, J., 2010. Rodents and lagomorphs from the hominoid site of Barranc de Can Vila 1, with remarks on the structure and diversity of Late Aragonian and Vallesian rodent assemblages from the Vallès-Penedès Basin (Catalonia, Spain). Neues Jahrbuch für Geologie und Paläontologie-Abhandlungen, 257(3), 297–315. DOI: https://doi.org/10.1127/0077-7749/2010/0077
Casanovas-Vilar, I., Alba, D.M., Garcés, M., Robles, J.M., Moyà- Solà, S., 2011. An updated chronology for the Miocene hominoid radiation in Western Eurasia. Proceedings of the National Academy of Sciences U.S.A., 108(14), 5554-5559. DOI: https://doi.org/10.1073/pnas.1018562108
Casanovas-Vilar, I., Furió, M., Alba, D.M., Moyà-Solà, S., Méndez, J.M., 2012. Rodents and insectivores from the hominoid-bearing site of Can Feu (Vallès-Penedès Basin, Catalonia, Spain). Journal of Vertebrate Paleontology, 32(1), 225-230. DOI: https://doi.org/10.1080/02724634.2012.633948
Casanovas-Vilar, I., Almécija, S., Alba, D.M., 2015. Late Miocene flying squirrels from Can Llobateres 1 (Vallès-Penedès Basin, Catalonia): systematics and palaeobiogeography. Palaeobiodiversity and Palaeoenvironments, 95(3), 353-372. DOI: https://doi.org/10.1007/s12549-015-0192-1
Crusafont Pairó, M., 1964. La biota de Can Llobateres (Sabadell) y su significación biológica. Cursillos y Conferencias del Instituto “Lucas Mallada”, 9, 177-179.
Crusafont-Pairó, M., Golpe-Posse, J.M., 1974a. Asociación de Anchitherium Mey., 1834, con Hipparion Christ, 1832, en el Alto Mioceno del Vallés. Boletín de la Real Sociedad Española de Historia Natural (Geología), 72(1-4), 75-93.
Crusafont Pairó, M., Villalta, J.F. de, Bataller, J.R., 1948. Los castores fósiles de España. I. Parte general y descriptiva. Boletín del Instituto Geológico y Minero de España, 61, 321-423.
De Bruijn, H., Daams, R., Daxner-Höck, G., Fahlbusch, V., Ginsburg, L., Mein, P., Morales, J., Heizmann, E., Mayhew, D.F., Van der Meulen, A.J., Schmidt-Kittler, N., Telles Antunes, M., 1992. Report of the RCMNS working group on fossil mammals, Reisenburg 1990. Newsletters on Stratitgraphy, 26(2-3), 65-118.
Fortelius, M. (coordinator), 2015. New and Old Worlds Database of Fossil Mammals (NOW). University of Helsinki. Accessed 21 April 2015. Website: https://www.helsinki.fi/science/now/
Freudenthal, M., 1966. On the mammalian fauna of the Hipparion- beds in the Calatayud-Teruel Basin. Part 1: The genera Cricetodon and Ruscinomys (Rodentia). Proceedings of the Koninklijke Nederlandse Akademie van Wetenschappen Series B, 69(2), 296-317.
Golpe Posse, J.M. 1974. Faunas con yacimientos de suiformes en el Terciario español. Paleontología y Evolución, 8, 1-87.
Hartenberger, J.-L., 1965. Les Cricetidae (Rodentia) de Can Llobateres (Néogène d’Espagne). Bulletin de la Société Géologique de France, 7(7), 487-498.
Hartenberger, J.-L., 1966. Les rongeurs du Vallésien (Miocène superieur) de Can Llobateres (Sabadell, Espagne): Gliridae et Eomyidae. Bulletin de la Société Géologique de France, 7(8), 596-604.
Hartenbeger, J.-L., 1967. Contributions à l’étude de l’anatomie crânienne des Rongeurs. I. – Pricipaux types de Cricétodontinés. Palaeovertebrata, 1(2), 47-64.
Hartenberger, J.-L., Thaler, L., 1963. Sur les rongeurs fossiles du Vallésien (Miocène supérieur) de Can Llobateres (Sabadell, Espagne). Comptes Rendus de l’Académie des Sciences, Sciences de la Terre et des Plantes, 256, 3333-3336.
Mein, P., 1970. Les sciuroptères (Mammalia, Rodentia) Néogènes d’Europe Occidentale. Geobios, 3(3), 7-77. DOI: https://doi.org/10.1016/S0016-6995(70)80001-8
Mein, P., 1999. The Late Miocene small mammal succession from France, with emphasis on the Rhône Valley localities. In: Agustí, J., Rook, L., Andrews, P. (eds.). Hominoid evolution and climatic change in Europe. The evolution of Neogene terrestrial ecosystems in Europe. Cambridge, Cambridge University Press, 1, 140-164.
Mein, P., Martín-Suárez, E., Agustí, J., 1993. Progonomys Schaub, 1938 and Huerzelerimys gen. nov. (Rodentia); their evolution in Western Europe. Scripta Geologica, 103, 41-64.
Morales, J., Nieto, M., Köhler, M., Moyà-Solà, S., 1999. Large mammals from the Vallesian of Spain. In: Agustí, J., Rook, L., Andrews, P. (eds.). Hominoid evolution and climatic change in Europe. The evolution of Neogene terrestrial ecosystems in Europe. Cambridge, Cambridge University Press, 1, 113-126.
Moyà-Solà, S., Köhler, M., 1993. Recent discoveries of Dryopithecus shed new light on evolution of great apes. Nature, 365(6446), 543-545. DOI: https://doi.org/10.1038/365543a0
Moyà-Solà, S., Köhler, M. 1996. A Dryopithecus skeleton and the origins of great-ape locomotion. Nature, 379(6561), 156-159. DOI: https://doi.org/10.1038/379156a0
Pineda-Muñoz, S., Casanovas-Vilar, I., Alba, D.M., Carmona, R., Rifà, E., 2011. Cricetinae (Rodentia) from site B40/ S5C (Vallès-Penedès Basin, NE Iberian Peninsula). Biostratigraphic implications. In: Pérez-García, A., Gascó, F., Gasulla, J.M., Escaso, F. (eds.). Viajando a mundos pretéritos. Morella, Ayuntamiento de Morella, 289-296.
Robles, J.M., Alba, D.M., Casanovas-Vilar, I., Galindo, J., Cabrera, L., Carmona, R., Moyà-Solà, S., 2011. On the age of the paleontological site of Can Missert (Terrassa, Vallès- Penedès, NE Iberian Peninsula). In: Pérez-García, A., Gascó, F., Gasulla, J.M., Escaso, F. (eds.). Viajando a mundos pretéritos. Morella, Ayuntamiento de Morella, 339-346.
Rotgers, C., Alba, D.M., 2011. The genus Anchitherium (Equidae: Anchitheriinae) in the Vallès-Penedès Basin (Catalonia, Spain). In: Pérez-García, A., Gascó, F., Gasulla, J.M., Escaso, F. (eds.). Viajando a mundos pretéritos. Morella, Ayuntamiento de Morella, 347-354.
Schaub, S., 1944. Cricetodontiden der Spanischen Halbinsel. Ber. Schweizerische Paläontologie Eclogae geologicae Helvetiae, 37(2), 453-457.
Schaub, S., 1947a. Los Cricetodóntidos del Vallés-Penedés. Estudios Geológicos, 6, 55-67.
Schaub, S., 1947b. Primer hallazgo de un ratón fósil en el Pontiense español. Publicaciones del Museo de la Ciudad, 3, 31-33.
Villalta Comella, J.F. de, 1950. Sobre un esciuróptero del Vindoboniense del Vallés-Penedés. Boletín de la Real Sociedad Española de Historia Natural, 48(1), 53-60.