Holocene palaeoecological changes recorded in mollusc-bearing cave sediments, the Cave above the Słupska Gate (southern Poland)
Holocene palaeoecological changes recorded in mollusc-bearing cave sediments, the Cave above the Słupska Gate (southern Poland)
Geologica Acta: an international earth science journal, vol. 14, no. 3, pp. 283-298, 2016
Universitat de Barcelona

This work is licensed under Creative Commons Attribution-ShareAlike 4.0 International.
Received: 15 September 2015
Accepted: 15 January 2016
Funding
Funding source: University of Warsaw
Contract number: 166901
Award recipient: Holocene palaeoecological changes recorded in mollusc-bearing cave sediments, the Cave above the Słupska Gate (southern Poland)
Funding
Funding source: Polish Ministry of Science and Higher Education
Contract number: 691-N/2010/0
Award recipient: Holocene palaeoecological changes recorded in mollusc-bearing cave sediments, the Cave above the Słupska Gate (southern Poland)
Abstract: The Cave above the Słupska Gate (southern Poland) contains about 2m depth of mollusc-bearing deposits. Radiocarbon and archaeological dating indicate that these deposits accumulated during the Holocene (Preboreal to Subatlantic), although the earliest layers may date from the end of the Pleistocene. Eight layers of silts, sands and loess-like deposits were distinguished at the site. Seven of them contained identiÀable snail shells, sometimes in large numbers, and sparse remains of vertebrates and archaeological artefacts. The molluscan assemblages retrieved from the cave contain over 40 taxa and 1,200 specimens. The balance of species distributed among 11 zoogeographical groups enabled us to identify four assemblages which differ in their ecological structure and in the composition of the fauna. The oldest fauna (Late Glacial/Preboreal and/or Preboreal) with many shade-loving species is typical of a cool climate. Episodes of drying are evidenced by the loess-like deposits and the occurrence of open-country snails such as the glacial relic Vallonia tenuilabris. This species disappeared in the younger part of the Early Holocene, which is the most distinctive feature of the Słupsko Hill sequence. The Middle Holocene climatic optimum is characterised by abundantand diverse fauna which is typical of mixed and deciduous forests with distinct oceanic influences. The critical Discus ruderatus and Discus rotundatus succession reÁects the general trends in European malacofaunas. The Late Holocene record may bear some hiatuses, but the shift away from a complete forest fauna is evident.
Keywords: Malacofauna, Cave sediment, Early Holocene, Middle Holocene, Palaeoenvironment.
INTRODUCTION
The Pleistocene/Holocene transition and the Early Holocene were a time of important climatic and environmental changes in Central Europe, which marked a transition from glacial to interglacial conditions. Significant changes, mostly connected with the increase of temperature and varying humidity, can be seen in plant cover, the development of water bodies and transformation of terrestrial environments.
Molluscs are one of the best indicators of environmental change. They document changes of many habitats, starting from the freshwater to dry land conditions. An excellent record of development of water bodies during the Late Vistulian/ Holocene boundary and the Holocene was provided by the malacofaunas of many lakes of northern Poland. The abundant proÀles of lacustrine chalk and calcareous gyttja yielded characteristic mollusc assemblages and their succession could be associated with certain climate and environmental conditions (e.g.Wojciechowski, 2000; Alexandrowicz, 2013a). In terrestrial environments, molluscan records from the late Quaternary come mainly from the calcareous tufas and loess deposits of southern Poland (e.g Alexandrowicz, 1983; 2001a, 2004, 2013b, 2014; Alexandrowicz et al., 2014). In Poland, loess sequences usually document severe conditions of the Late Vistulian (or older) with characteristic species Pupilla loessica, Pupilla muscorum, Vallonia tenuilabris, Semilimax kotulae, Columella columella, Succinea oblonga and the others (Alexandrowicz and Alexandrowicz, 1995a, b; Kruk et al., 1996), whereas those from tufas and travertines reveal faunas typical of different parts of the Holocene and the Pleistocene/Holocene transition. One of the best defined boundaries of these periods in Poland was noted in Groń (the Podhale Basin) (Alexandrowicz, 2013b), but abundant comparative material of that age is also available at many proÀles throughout Europe (e.g.Limondin-Lozouet and Rousseau, 1991; Meyrick and Preece, 2001; Meyrick, 2002; Gedda, 2001; Limondin-Lozouet and Preece, 2004; Limondin-Lozouet, 2011).
Despite the many malacofauna sequences reported from cave sediments, few of these go back as far as the Ànal part of last glaciation (Stworzewicz, 1973, 1988; Alexandrowicz et al., 1992; Svoboda et al., 2000; Alexandrowicz and Stworzewicz, 2003; Ložek, 2012). In southern Poland most such stratigraphic proÀles are only a few dozens of centimetres thick, covering the middle or just the youngest part of the Holocene (e.g.Bocheński et al., 1985; Alexandrowicz, 1992, 1997, 2000a, b, 2001b; Alexandrowicz and Rudzka, 2006).
Hence, the relatively thick sequence of mollusc-bearing sediments from the Cave above the Słupska Gate can provide a more complete account of local environmental changes over the whole of the Holocene. Here, we characterise the succession of mollusc assemblages of the Cave above the Słupska Gate and relate them to the conditions of their development and the accumulation of cave deposits. Finally, the palaeoenvironment of surrounding area and the climate variability is reconstructed and compared with regional trends. The interpretation is supplemented by analysis of vertebrates.
GEOLOGICAL SETTING
The Cave above the Słupska Gate (N: 50º35’27.8” E: 19º34’15.2”) is situated on the western slope of Słupsko, the easternmost hill of the Kroczyckie Rocks ridge in the central part of the Cracow-Częstochowa Upland, ca. 40km south-east of Częstochowa (Fig. 1). It was not mentioned in literature before the excavation in 2011-2012. Its name (Jaskinia nad Bramą Słupską in Polish) was proposed by spelaeologist K. Mazik during the spelaeological symposium in Olsztyn, Poland, in 2013 (Krajcarz et al., 2014a). The area is built of the Upper Jurassic massive limestone, partially covered by the red desert sands and loams dated to Paleogene/Neogene (Gradziński, 1977) and sandy deposits attributed to the Áuvio-periglacial facies of the Middle Pleistocene (Lewandowski, 1994). The karstic cavities and caves are Àlled with the Late Pleistocene and Holocene cave and colluvial sediments. The Cave above the Słupska Gate is not an isolated site in the region, but several others are known from the Kroczyckie Rocks, including Deszczowa Cave, Shelter on Słupsko Hill, Cave Wisielców and Krucza Skała Rockshelter (Cyrek, 1994; Madeyska, 1996; Cyrek et al., 2000; Nadachowski et al., 2009; Krajcarz and Madeyska, 2010; Krajcarz et al., 2014b).
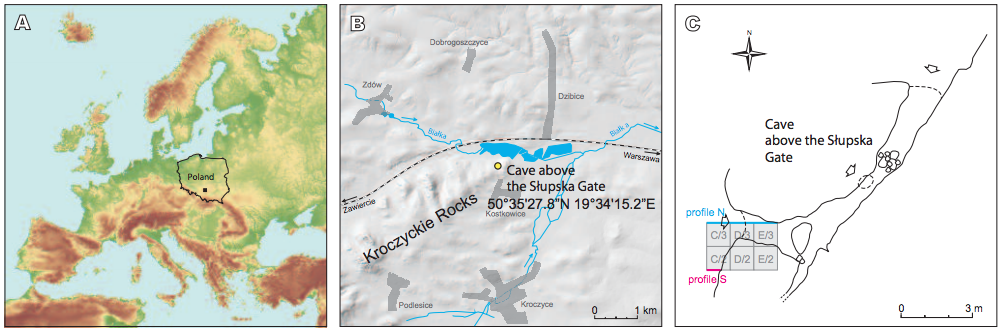
FIGURE 1
A, B) Localisation of the Cave above the Słupska Gate. C) Plan of the cave with situation of archaeological trench in 2012.
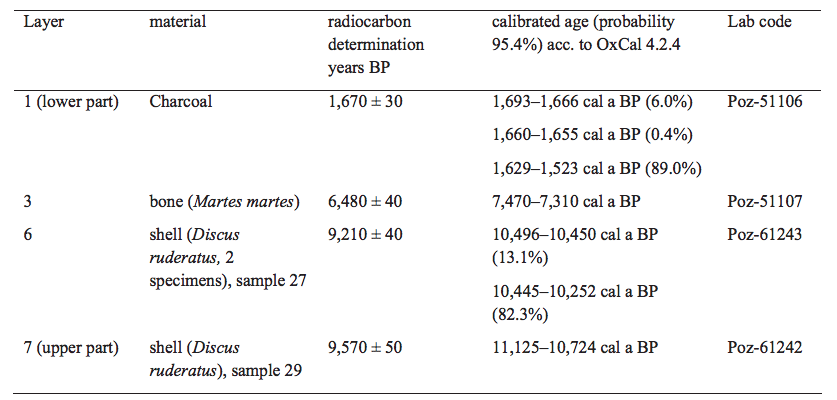
The sedimentary sequence in the Cave above the Słupska Gate is up to 2m thick. Altogether 8 layers of silts, sands and loess-like deposits were distinguished (Fig. 2). Seven of them bear abundant and diverse malacofauna and sparse remains of vertebrates. The sediments under study represent only near-entrance facies of sedimentary Àll, with intercalations of colluvial sediments. The lithology and stratigraphy of sediments is presented in Table 1. The chronology of deposition is shown in Table 2.
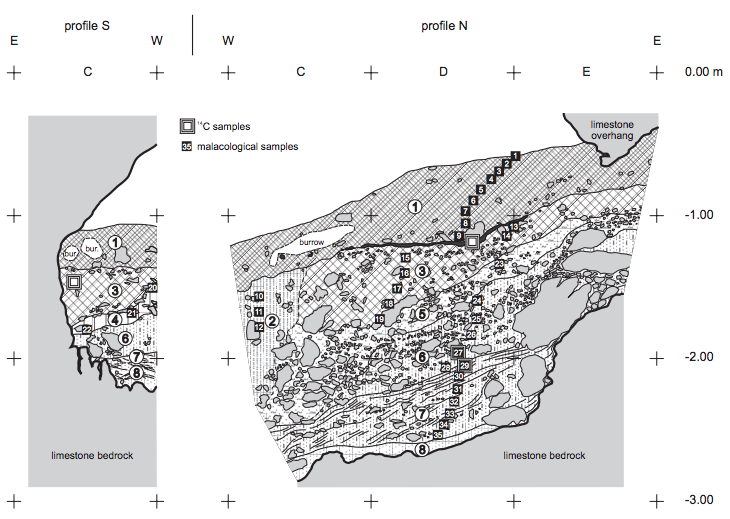
FIGURE 2
Geological cross-section through the sedimentary fill of the Cave above the Słupska Gate (profiles S and N of archaeological trench). For lithological description see Table 1.

MATERIAL AND METHODS
Mollusc analysis
Mollusc analysis was conducted using 35 samples (weight 3-5kg) taken at ca. 10cm intervals from 7 layers of the cave sediments during the archaeological excavation conducted in 2012. The excavation was conducted on a N-oriented archaeological metre grid (Fig. 1) and the material was collected from five square meters (C/2, C/3, D/2, D/3, E/3), excavated in the near-entrance facies of the sedimentary fill. Malacological investigations were carried on in accordance with the schemes proposed by Ložek (1964, 1986, 2000) and Alexandrowicz and Alexandrowicz (2011). Samples were sieved on 0.5mm mesh and carefully cleaned. Mollusc remains, picked up from the samples, were taxonomically determined under a binocular microscope at magnifications up to 64× using taxonomical keys (Kerney et al., 1983; Wiktor, 2004; Welter-Schultes, 2012) and comparative collection of the Department of Climate Geology, Faculty of Geology, University of Warsaw. All completely preserved shells and their identifiable fragments were picked from sediments and counted applying schemes for broken individuals (Ložek, 1964; Alexandrowicz and Alexandrowicz, 2011). Damaged specimens were determined only to the genus or family levels, and the calcareous plates of slugs were counted together under the heading of Limacidae.
Identification of vertebrate taxa
Larger remains of vertebrates were collected directly from the sediment during excavation. The small bones and teeth were extracted from the same sediment samples studied for molluscs. Specimens were identified to skeletal element and species where possible. The remains of birds were identified with the use of the comparative collection of bird bones of the Institute of Systematics and Evolution of Animals PAS (Kraków, Poland), and those of mammals using the comparative collection of mammalian bones of the Institute of Geological Sciences PAS (Warsaw, Poland). The quantification of examined material follows Lyman (1994, 2008). The abundance of fossil mammal and bird material is very low and it occurred mainly in the uppermost layers 1–3, and only single remains in the lower layers. Altogether, only 72 identifiable remains were discovered.
Palaeoecological analysis
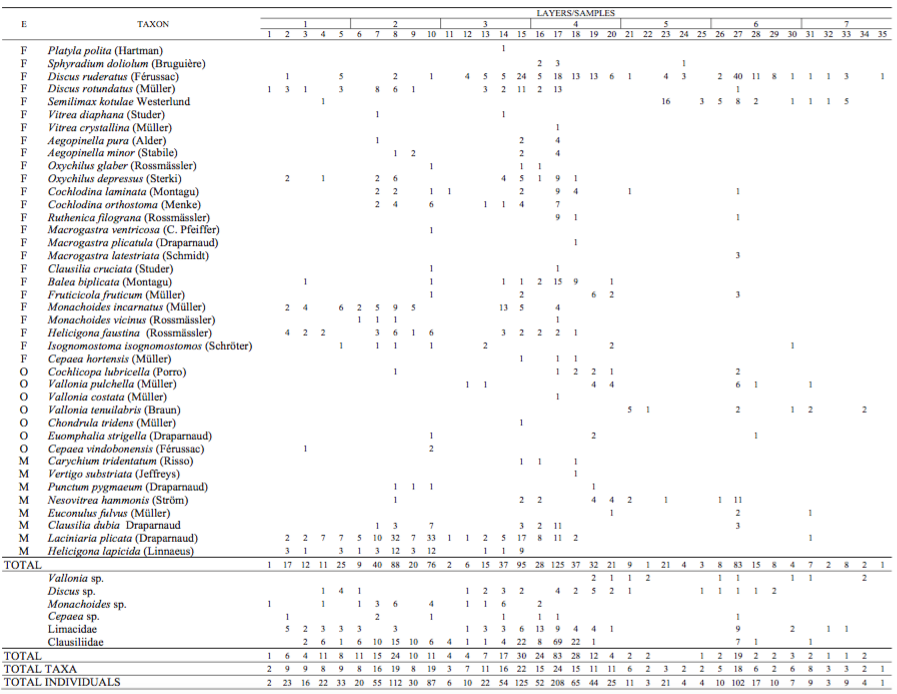
All mollusc species and specimens were grouped according to their environmental preferences (Table I, after Kerney et al., 1983; Wiktor, 2004; Alexandrowicz and Alexandrowicz, 2011; Welter-Schultes, 2012) and zoogeographical distribution (Table 3, after Alexandrowicz and Alexandrowicz, 2011). The structure of malacocoenoses was presented on the Malacological Spectra of Species (MSS) and Individuals (MSI) (Fig. 3). Because the spectra are affected by discrepancies in numbers of shells in each sample, the average percentages were also drawn for molluscan assemblages (Fig. 3). Malacological succession was illustrated by the frequency bar diagram (Fig. 4) providing the picture of environmental and climatic changes during the deposition. For samples with more than 50 specimens a percentage of the total sum was calculated, whereas for those containing less than 50 specimens only absolute numbers of shells were declared (Fig. 4). Mollusc assemblages were distinguished subjetively on the basis of their structure and composition. Statistical clustering appeared inadequate due to variable numbers of recorded shells. Zoogeographical distribution was illustrated by diagram of absolute number of species for each zoogeographical group (Fig. 5). Species with corresponding ranges were listed in Table 3. Both the character and succession of the assemblages were used in the palaeoecological and palaeoclimatic reconstructions.
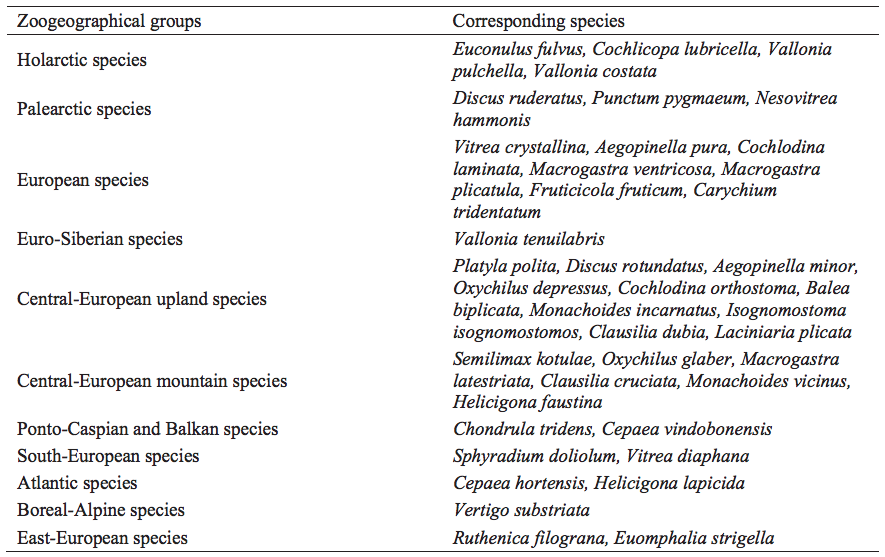
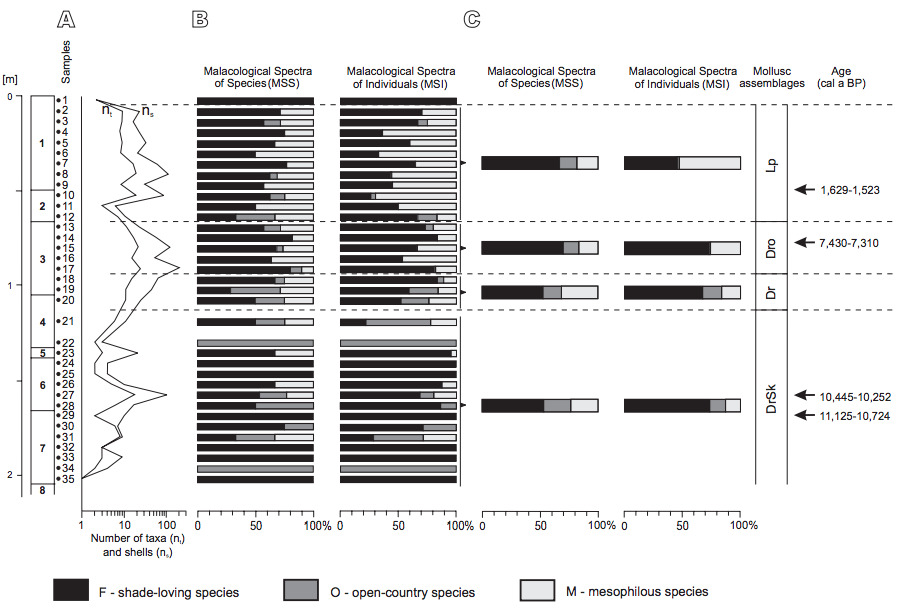
FIGURE 3
Ecological composition of molluscs in the Cave above the Słupska Gate. A) Numbers of taxa and shells per sample. B) Malacological spectra of individual samples. C) Malacological spectra of mollusc assemblages. Mollusc assemblages, DrSk: assemblage with Discus ruderatus and Semilimax kotulae; Dr: assemblage with Discus ruderatus; Dro: assemblage with Discus rotundatus; Lp: assemblage with Laciniaria plicata.

FIGURE 4
Malacological diagram of the Cave above the Słupska Gate. In samples with less than 50 individuals dots indicate single shells and numbers in brackets indicate a number of shells. Mollusc assemblages, DrSk: assemblage with Discus ruderatus and Semilimax kotulae; Dr: assemblage with Discus ruderatus; Dro: assemblage with Discus rotundatus; Lp: assemblage with Laciniaria plicata.

FIGURE 5
Zoogeographical distribution of molluscs from the Cave above the Słupska Gate. Zoogeographical groups after Alexandrowicz and Alexandrowicz (2011).
Radiocarbon dating
Radiocarbon dating was conducted in the Poznan Radiocarbon Laboratory (Poznań, Poland) to establish the chronology of the stratigraphic profile. The Accelerator Mass Spectrometry (AMS) method was applied to shells of the snail Discus ruderatus representing layers 7 and 6 (samples 29 and 27), the bone of pine marten Martes martes from layer 3 (sample SŁ W-26) and the charcoal from the Àreplace found on the boundary of layers 2 and 1 (sample SŁ P-6) (Table 2). Analysed snail shells were composed of aragonite, with no measurable admixture of calcite evidenced by X-ray diffraction analysis, thus the negative effect of recrystallisation was excluded. In case of bone the extracted collagen was dated. All dates presented were calibrated with OxCal v. 4.2.4 (Bronk Ramsey and Lee, 2013) versus the IntCal’13 radiocarbon calibration curve (Reimer et al., 2009).
RESULTS
Mollusc assemblages
Altogether 46 taxa of land snails, including 40 species, 4 genera, one family (Clausiliidae) and a collective group of Limacidae were recognised in the Cave above the Słupska Gate (Table I). They are represented by 1,226 specimens classiÀed in three ecological groups, F: shadeloving taxa (25 species), O: open-country taxa (7 species), M: mesophilous taxa (8 species). The number of taxa and specimens varies between 1 and 24 and 1 and 208 per sample, respectively, with the greatest abundance in layer 3 (Fig. 3; Table I). Four molluscan assemblages can be identified (Fig. 4), differing in their ecological profile.
The assemblage with Discus ruderatus and Semilimax kotulae (DrSk) occurs between samples 35 and 21 (layers 7, 6, 5 and the lower part of layer 4) (Fig. 4). The concentration of mollusc shells is rather low with one peak in sample 27 (layer 6), mostly related to large numbers of these two species. They are forest species characteristic of a cool, continental climate. The occurrence of glacial relic Vallonia tenuilabris is characteristic of this assemblage.
This is typical loess species usually connected with the severe climatic conditions and open areas of so-called “coldsteppes” (Ložek, 1964), but based on recent data from Asia, it may also occur in alpine grasslands, taiga, hemiboreal forest and wooded fens (Schileyko, 1984; White et al., 2008; Horsák et al., 2010, 2015). Its highest frequency is noted in sample 21, layer 4. The assemblage is composed mainly of shade-loving gastropods, with the proportion of 53 and 74% of all species and all individuals, respectively (Fig. 3). In the middle of the layer 6, the meadow species Vallonia pulchella and some mesophilous species of wide tolerance occur frequently - Nesovitrea hammonis, Euconulus fulvus and the thermotolerant, calciphile Clausilia dubia, common on shady and humid rocks. The occurrence of a single shell of a species of higher thermal demands, Discus rotundatus, in this assemblage is also noteworthy (Fig. 4; Table I). Two radiocarbon dates relating to this assemblage indicate that it is of Preboreal age (Table 2).
The assemblage dominated by Discus ruderatus (Dr) occurs in samples 20–18 (the uppermost part of layer 4 and the lowermost of layer 3). D. ruderatus is characteristic of taiga-type coniferous forests. It is accompanied by other shade-demanding gastropods (53% of species and 68% of individuals) Balea biplicata, Fruticicola fruticum, Isognomostoma isognomostomos and others (Figs. 3; 4). Gastropods of open habitats constitute averagely 16% of all species and 17% of all individuals (Fig. 3), with the most numerous being Vallonia pulchella, Cochlicopa lubricella and Euomphalia strigella. Amongst mesophilous species, N. hammonis dominates. From this assemblage upwards V. tenuilabris does not occur in studied cave deposits (Fig. 4; Table I).
Above the layer bearing Dr fauna there is a rich assemblage with Discus rotundatus (Dro) in the middle and upper part of layer 3 (samples 17–13) (Fig. 4). It is the richest and most abundant assemblage in the succession. D. ruderatus still dominates, but forest species requiring humid- and warm conditions: D. rotundatus, Oxychilus depressus, Monachoides incarnatus, Helicigona faustina and Cochlodina orthostoma become commoner. The assemblage is supplemented by abundant taxa of wide ecological tolerance, which represent 17% of all species and 25% of all individuals (Fig. 3). The most frequent Laciniaria plicata and Helicigona lapicida become a main component of the fauna in the uppermost part of the profile (Fig. 4; Table I). Distinct reduction in numbers of shells of open-country taxa from 17 to 1% is noteworthy (Fig. 3). The radiocarbon age of a pine marten from this layer (Table 2) indicates that this assemblage corresponds to the climatic optimum of the Holocene, the Atlantic phase.
There is a significant change in the character of the mollusc assemblages in the uppermost part of layer 2, and in the whole of layer 1 (samples 11–2) (Fig. 4). These tend to be dominated by Laciniaria plicata (Lp). The assemblage is quite rich. The most characteristic of the association is expansion of mesophilous snails (52% of all individuals, Fig. 3) represented mostly by L. plicata and H. lapicida, most frequent in the humid rocks in open environments and deciduous forests and a shady rocky substrate, respectively. Among shade-loving species the most numerous are M. incarnatus, H. faustina and D. rotundatus also connected with humid rocks and humid forest communities. D. ruderatus, characteristic of continental conditions, is still present but in low numbers (Fig. 4; Table I). Sample 1, at the top of the succession, is very poor. The single shell of D. rotundatus found cannot be interpreted. Charcoal from the lowest part of layer 1 yields a radiocarbon age of ca. 1,650BP (1,629-1,523cal a BP; Table 2) indicating that layer 1 was deposited within the Subatlantic phase.
Zoogeographical analysis
Species found in the malacofauna of the Cave above the Słupska Gate could be assigned to 11 zoogeographical groups (Fig. 5; Table 3). Although there are minor fluctuations in the proportions of individuals of species belonging to each even in neighbouring samples, there is a broadly bipartite division. The lower part of the profile (layers 7–4) contains mostly Palearctic and Central European mountain taxa supplemented by Holarctic, European, South-European, East-European, CentralEuropean upland and Euro-Siberian species represented by Vallonia tenuilabris (Table 3). In the upper part of the section (layers 3–1), by contrast, Central-European upland species dominate and Euro-Siberian species disappear being replaced by the Atlantic and Ponto-Caspian taxa connected with better climatic conditions (Fig. 5).
Vertebrates
Altogether, 14 mammal taxa and 7 bird taxa have been recognised in the Cave above the Słupska Gate (Table 4). Most of the mammal remains are of rodents.
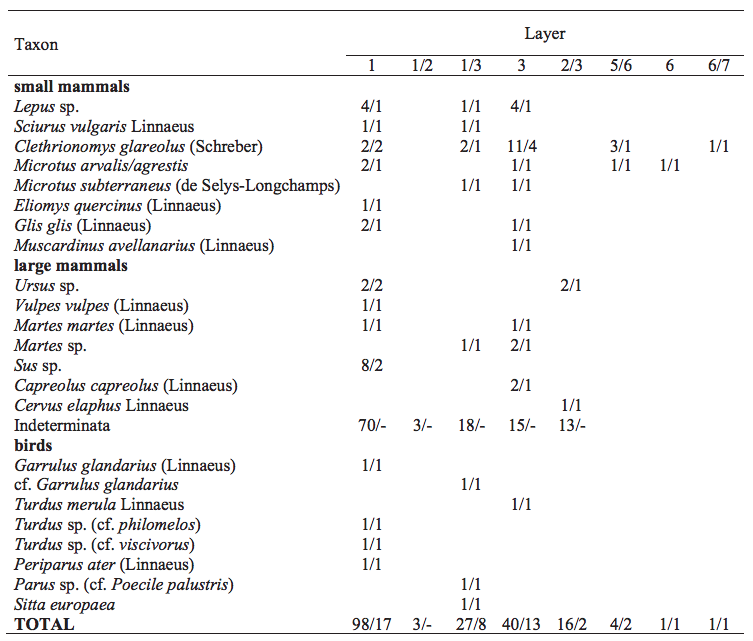
The forest rodents are represented by 3 species of the family Gliridae (Glis glis, Eliomys quercinus and Muscardinus avellanarius) and Sciuridae with one taxa - Sciurus vulgaris. The remains of these species were found in layers 1 and 3, and at the boundary between these layers (1/3). The remains of voles are slightly more abundant. They are represented by woodland and bushy area species (Clethrionomys glareolus) and eurytopic species (Microtus arvalis/agrestis and Microtus subterraneus). Remains of C. glareolus and M. arvalis/agrestis are the only found in the lower part of the sequence (layers 7–4). Lagomorphs are represented by several remains of Lepus sp.
Larger mammals are represented by isolated teeth and fragments of bones. Among them are 4 forest species (Cervus elaphus, Capreolus capreolus, Martes martes and Ursus sp.) and one eurytopic species (Vulpes vulpes). The bird bones are sparse and represent forest species. Remains of birds and large mammals were found only in the upper part of sedimentary profile, in layers 1–3.
DISCUSSION
Age of deposition
Although we have no dated material from the lowest layer (8) in this cave, its composition of yellowish quartz sands is similar to those underlying Holocene deposits in other caves in the region. In Deszczowa Cave (Madeyska, 1996; Cyrek et al., 2000) these were dated at ca. 19,000BP, and in Krucza Skała Rockshelter to between 11,000 and 13,000BP (Nadachowski et al., 2009; Lorenc, 2013). Both radiocarbon dating and archaeological evidence (Krajcarz et al., 2014a) confirm that the sequence of layers 7–1 in the Cave above the Słupska Gate was accumulated during the Holocene from Preboreal to mid-Subatlantic times. Erosional boundaries and hiatuses are expected in the bottom of layers 7 and 1.
Palaeoecological changes of mollusc fauna
Although there are some discontinuities in the pattern of deposition in the cave, both radiocarbon dating and the comparison with other cave deposits in the region (Krajcarz et al., 2016) indicate that the sequence of changes in the malacofauna reflect a chronological sequence of local environmental changes. Despite their polygenetic character the studied thanatocoenoses appear to be representative of the sediments and stratigraphical levels where they occur. The deposits contain shells of some shade-loving species penetrating the cave, of those living in the surface near the cave entrance and shells dropped from the surrounding rocks during sedimentation. Some shells, especially those in laminated deposits (layer 7), might have been washed into the cave from the slope or blown with the wind, but these transportations were rather short in time and distances (cf.Alexandrowicz and Alexandrowicz, 2011). The shells represent the closest neighbourhood being a good indicator of diversity of habitats around the cave (cf.Alexandrowicz et al., 1985; Juřičková et al., 2014). Intrusions from younger deposits caused by animal and/or human activities also occur, but are of minor importance as indicated by the rather environmentally coherent composition of the fauna. Traces of zoogenic and anthropogenic disturbances were visible during excavation only in layer 1.
The oldest ages of ca. 11,125–10,724cal a BP, corresponding to the Preboreal phase of the Holocene (Starkel et al., 2013), are represented by the middle part of the assemblage dominated by Discus ruderatus and Semilimax kotulae (Fig. 6), in the transition between layers 7 and 6. Layer 7 was formed by slope wash, as shown by its lamination and inclination. Despite quick deposition in relatively high energy these sediments are devoid of shell detritus. The rather good state of preservation of shells suggests that they did not undergo intensive transportation, but were accumulated during the deposition. They represent the cave surroundings and it appears that the extremely poor assemblage resulted from severe condition for mollusc development (cf.Ložek, 2012; Lorenc, 2013), as Late Glacial origin of the base of this layer cannot be excluded. The latter is not unequivocal, especially in the light of colluvial character of deposits, but similar malacocoenoses of Late Glacial/Holocene transition predominated by D. ruderatus and S. kotulae, with the occurrence of species of wide thermal tolerance (e.g.Euconulus fulvus) were noted in the landslide deposits and calcareous tufas of southern Poland during the Alleröd interphase (e.g.Alexandrowicz, 1997, 2001a, 2004; Alexandrowicz and Alexandrowicz, 1999). Within layer 6 the lamination disappears and limestone rubble becomes abundant, indicating a change from slope processes to the accumulation of cave debris. The inclination exhibited by layers 6 and 4 suggests that slope processes, such as creep, were still active, but not intensive. The stratigraphy of layers 5 and 4 is uncertain but their position in the proÀle suggests correlation with the Boreal phase (Fig. 6). According to palaeobotanical data coniferous forests with pine, spruce and larch dominated in the Cracow-Częstochowa Upland of that time. During the periods of deterioration of the climate some transformations to sparse forests with larch and a “park tundra with abundant steppe-like communities” might occurred (Madeyska, 1998). These changes can be hardly interpreted in the poor mollusc assemblage of the sample set 35–21, but some general environmental characteristic of these units may be defined, as malacocoenoses of the Early Holocene with high proportions of D. ruderatus and S. kotulae are also known from other sites (e.g.Alexandrowicz et al., 1985). Both species indicate taigatype communities, relatively low temperatures and humid conditions. Rather cool climate is also evidenced by the glacial relic V. tenuilabris which usually disappeared in Central Europe in the Pleistocene/Holocene boundary (e.g.Dagnan-Ginter et al., 1992; Wiktor, 2004; White et al., 2008; Horsák et al., 2010, 2015), but here constitute very characteristic element of the fauna. Accumulation of loess-like deposits with the poor fauna with V. tenuilabris may indicate some transformation towards more open areas around the cave, but due to the occurrence of only few individuals this interpretation is open to discussion. Presence of single shells of Cochlodina laminata and D. rotundatus is not typical of such conditions nor their assumed age, and thus appears to result from re-deposition caused by the activity of burrowing animals. Small mammals in the DrSk assemblage are represented only by eurytopic Microtus arvalis/agrestis and Clethrionomys glareolus mostly associated with arboreal and shrub vegetation but not indicative for certain conditions. Although those species are not diagnostic, similar faunal compositions and biological disturbances are frequent in the Early Holocene cave deposits of the Moravian Karst in Czech Republic (Svoboda et al., 2000).
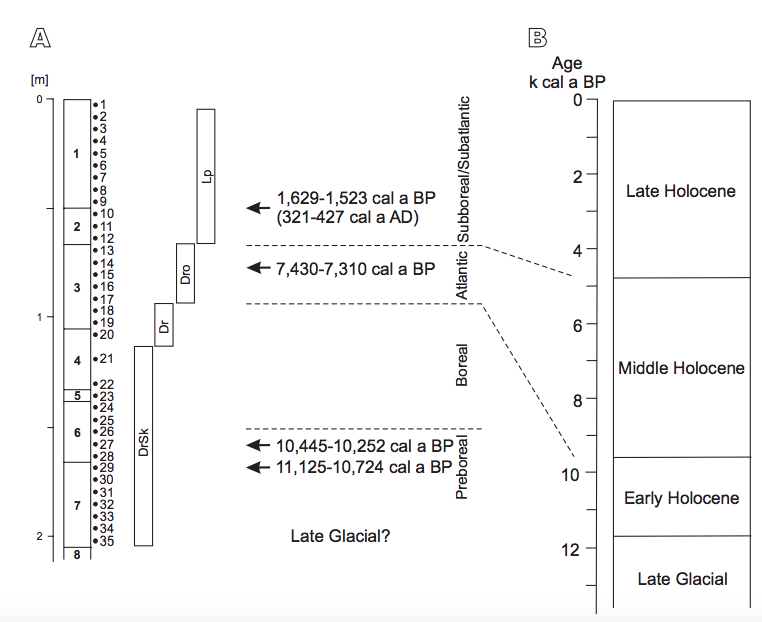
FIGURE 6
A) Stratigraphy and succession of molluscan fauna in the Cave above the Słupska Gate. B) Holocene subdivision according to Starkel et al. (2013). Mollusc assemblages, DrSk: assemblage with Discus ruderatus and Semilimax kotulae; Dr: assemblage with Discus ruderatus; Dro: assemblage with Discus rotundatus; Lp: assemblage with Laciniaria plicata.
The final retreat of V. tenuilabris population on the Słupsko Hill appears to be correlated with the Boreal phase and the expansion of D. ruderatus. This faunal shift is accompanied by many shade-loving species (e.g. Fruticicola fruticum, Balea biplicata, Isognomostoma isognomostomos), which indicate an increase of humidity and favourable thermal conditions. Occurrence of weathered, smoothed and pebble-like limestone rubble, starting at level 4, also points to climatic warming and moistening (Krajcarz and Madeyska, 2010). The composition of the assemblage resembles the Ruderatus-fauna described by Dehm (1967) and noted in many terrestrial deposits of Early Holocene across Europe between the Preboreal and the older part of the Atlantic phase (e.g. Ložek, 1964, 2000; Preece and Day, 1994; Preece and Bridgland, 1999; Gedda, 2001; Meyrick, 2002; Limondin-Lozouet and Preece, 2004; Limondin Lozouet, 2011).
The climatic optimum of the Holocene is represented by Discus rotundatus assemblage (Fig. 6). Large numbers of both species and shells result from abundant mollusc communities existing around the cave in favourable thermal and moisture conditions (cf.Svoboda et al., 2000). Mixed or deciduous forests prevailed in the studied area, as evidenced among others by Oxychilus depressus, Monachoides incarnatus, Helicigona faustina, Cochlodina orthostoma and forest rodents Glis glis and Muscardinus avellanarius. This is in agreement with Áoristic changes in Central Europe described by, among others, Starkel (1977) and Ralska-Jasiewiczowa (1991). Gradual replacing of D. ruderatus by D. rotundatus indicates both the progressive warming of the climate and a transition from continental to oceanic circulation (e.g.Alexandrowicz, 2004, 2013b; Alexandrowicz and Rybska, 2013; Alexandrowicz et al., 2014), supported in the proÀle by the expansion of Atlantic and PontoCaspian species. The structure of the assemblage resembles Discus rotundatus and Aegopinella pura fauna typical of somewhat cooler periods during the climatic optimum of the Holocene, usually in the beginning or in its Ànal part (e.g.Ložek, 1964; Alexandrowicz, 1983, 1992, 2004; Alexandrowicz and Alexandrowicz, 1995b). It may be also supported by still high frequency of D. ruderatus. The conditions around the cave were probably not suitable for development of characteristic warmthdemanding Perspectivus-fauna noted in many European sites (e.g. Ložek, 1964; Fuhrmann, 1973; Dehm, 1987; Füköh, 1995; Meyrick, 2002; Alexandrowicz and Rybska, 2013; Alexandrowicz et al., 2014) or this part of the optimum is not recoded in the proÀle of the Cave above the Słupska Gate.
Significant reduction of sediment thickness above layer 3 suggests restricted deposition and/or possible hiatuses in the sequence of mollusc assemblages. Layer 2 may represent the lateral facies of layer 3, accumulated outside of the cave. However, its upper part is probably younger than layer 3. Layer 1, the youngest, originated after intensive human settlement in the cave (1,629– 1,523cal a BP; 321-427cal a AD), and its lower boundary with layers 2 and 3 is anthropogenic (Krajcarz et al., 2014a) and there is a hiatus between layers 2–3 and layer 1. Some burrows occur in the sediment of layer 1 but mollusc assemblage appears to be free of intrusive elements, as its composition, despite some irregular distribution of shells, is rather constant. Although gradual retreat of forest species noted in Lp assemblage is observed since Subboreal in Central Europe (e.g.Füköh, 1995), the detailed biostratigraphical subdivision for the Late Holocene is practically impossible due to lack of substantial changes in the composition of the mollusc assemblage. Compared to Dro assemblage, the transformation of fauna is evidenced by the expansion of gastropods of wide ecological tolerance with predominance of Laciniaria plicata in the Protohistorical and historical periods. Rodents and birds of woodland areas are still present in the assemblage, but abundant L. plicata indicates the exposing of open rocky walls near the site. More intensive human activity in the region is possible (Cyrek, 1994; Kruk et al., 1996; Alexandrowicz, 2004; Rozmus et al., 2006; Krajcarz et al., 2014a), but estimation of its inÁuence on malacofauna of the Słupsko Hill is not suficiently evidenced.
Regional context and comparison with other records
Compared to other records of cave sediments and rocky shelters of Central Europe (Stworzewicz, 1973, 1988; Ložek, 1980, 2012; Alexandrowicz et al., 1985; Bocheński et al., 1985; Alexandrowicz, 1992, 1997, 2000a, b, 2001b; Füköh, 1995; Svoboda et al., 2000; Alexandrowicz and Stworzewicz, 2003) the malacofauna from the Cave above the Słupska Gate, with their over 40 taxa and 1,200 specimens, appears to be signiÀcant for palaeoecological studies of the Holocene and the Late Glacial of the Vistulian. Accumulations of ca. 2m of mollusc-bearing cave deposits is rather unique in the region and occurs only in few sites such as e.g. Veľká Drienčanská Cave in Slovakia (Ložek, 2012) and the Pekárna and Barová caves in the Moravian Karst in Czech Republic (Svoboda et al., 2000). Among dozens of caves, rock-shelters and rocky niches of southern Poland, just a few have more than 20-50cm thick sedimentary fills with mollusc remains, usually corresponding to the Late Holocene (Alexandrowicz, 1997; Alexandrowicz, 2000a, b; Alexandrowicz, 2001b; Alexandrowicz and Rudzka, 2006). The local variability of shell-bearing deposits and of cave thanatocoenoses hinder the regional comparisons, but the main traits of malacofauna evolution over the central European region may be highlighted. Despite some considerable differences in mollusc composition of different regions, the environmental changes and mollusc succession of the Słupsko Hill correlates well with those inferred for Central-European uplands and karstic mountains (e.g.Starkel, 1977; Ralska-Jasiewiczowa, 1991; Alexandrowicz and Alexandrowicz, 1995a; Füköh, 1995; Madeyska, 1998; Alexandrowicz, 2000b; Svoboda et al., 2000; Ložek, 2012). In the Early Holocene most of mid-European cave and other terrestrial records are characterised by low mollusc recoveries (Füköh, 1995; Svoboda et al., 2000; Gedda, 2001; Ložek, 2012). The distinctive feature of Preboreal and Boreal assemblages is the occurrence of Discus ruderatus. At Słupsko Hill and in the cave in the Sobczański Gully (Pieniny Mts.) (Alexandrowicz et al., 1985) it is accompanied by Semilimax kotulae, but many local variations occur in mollusc composition of that time throughout Europe (Füköh, 1995; Preece and Bridgland, 1999; Svoboda et al., 2000; Gedda, 2001; Meyrick and Preece, 2001; Ložek, 2012). The mid-Holocene optimum in the Cave above the Słupska Gate with a quite rich Dro assemblage and declined open-country species is correlative with abundant forest communities of Moravian, Slovakian Hungarian and German records (Füköh, 1995; Svoboda et al., 2000; Meyrick, 2002; Ložek, 2012). Well marked species-rich optimum is also characteristic of many british and french sequences, but the proportion of woodland species of those is considerably lower than in midEuropean assemblages (Juřičková et al., 2014). Those far distant sequences are usually hardly comparable with Central Europe, but record the most critical change of mollusc communities during the Atlantic period noted across many European land sequences. Starting from ca. 900014C BP Discus rotundatus expanded throughout Europe and gradually replaced D. ruderatus, reaching its highest abundance at approximately 800014C BP. This transformation was not synchronous and even a one-thousand-years delay may occur between several countries (Preece and Day, 1994; Meyrick and Preece, 2001; Meyrick, 2002). In Cave above the Słupska Gate the upper part of D. rotundatus zone indicated by the date of 7,430–7,310cal a BP (radiocarbon age of 6,480±40BP) occurs, but its detailed chronology stays open as its lower boundary have not been dated. It is noteworthy that D. ruderatus was still abundant around the cave and in smaller numbers survived until today. This is common in mid-European sequences, whereas in Britain it disappeared at radiocarbon age of ca. 7000BP and in France today occurs only in isolated sites in the Pyrenees (Preece and Bridgland, 1999; Welter-Schultes, 2012). The Late Holocene succession is characterised by the limited occurrence of shade-loving and expansion of mesophilous taxa (cf.Füköh, 1995; Svoboda et al., 2000; Ložek, 2012). The mollusc community with the most numerous Laciniaria plicata and Helicigona lapicida appears to be characteristic of the Słupsko Hill and at the moment is not correlated with any regional trends.
The most distinctive feature in the Cave above the Słupska Gate is the abundance and long occurrence of the glacial relic Vallonia tenuilabris, unknown both from the Holocene and most of the cave deposits of Central Europe. Nowadays this species occurs in Siberia, Central Asia and northern China (Dagnan-Ginter et al., 1992; Gerber, 1996; Meng, 2008, 2009; White et al., 2008; Horsák et al., 2010, 2015). It is common in the Late Vistulian loess and loess-loams of southern Poland, but in cave sediments has only been described in the Zawalona Cave and Duża Cave (southern part of the Cracow-Częstochowa Upland) (Alexandrowicz et al., 1992; Dagnan-Ginter et al., 1992; Alexandrowicz and Alexandrowicz, 1995a). In the latter it was accompanied by abundant forest species, thus some reworking and intermixing of the mollusc material was postulated (Dagnan-Ginter et al., 1992). It can never be excluded in cave sediments, but geological situation and sediment features in the Cave above the Słupska Gate do not indicate significant reworking of sediments and shell material, and a vast occurrence of V. tenuilabris from dry steppes to various woodland habitats in Asia, provides alternative ways of interpretation. Another appearance of this species in recently studied caves and shelters of the Cracow-Częstochowa Upland (Krajcarz et al., 2016) may confirm microclimate favourable for longer existence of this species in the Early Holocene and offers a perspective on further studies of the problem.
CONCLUSIONS
Sediments of the Cave above the Słupska Gate contain differentiated mollusc assemblages. Their composition and structure reflect the depositional conditions around the cave during the Holocene and possibly the final part of the Late Vistulian. The detailed stratigraphic subdivision is not unequivocal, some hiatuses occur, but the Early, Middle and Late Holocene are quite well marked in the succession. The development of malacofauna indicates both the environmental and climatic changes of that time. The oldest, rather poor fauna (Late Glacial/Preboreal and/or Preboreal) is typical of a cold climate and light coniferous forests. The shade-loving taxa were the main component of mollusc assemblages, but episodes of drying and cooling of the climate are evidenced by the loess-like deposition and expansion of open-country snails with glacial relic V. tenuilabris. The latter disappeared in the younger part of the Early Holocene. The main phase of development of malacocoenoses correlates with the Middle Holocene. Radiocarbon dating provides age characteristic of the Atlantic phase. The climatic optimum is characterised by the abundant, differentiated fauna characteristic of mixed and deciduous forests and distinct oceanic influences. Malacological record of the Late Holocene may bear some gaps. Forests still existed in the surroundings of the site, but more areas appear to be relatively poorly vegetated, especially rocky walls around the cave. Gastropods of wide ecological amplitude dominated in the assemblage being the main component of the fauna during the Protohistorical and historical period noted in the uppermost part of the succession. The cave sequence on the Słupsko Hill corresponds to the malacological records of Central European uplands bearing some individual characteristics, especially the long occurrence of glacial relic V. tenuilabris.
ELECTRONIC APPENDIX I
Agradecimientos
This research has been Ànanced by the University of Warsaw through the Faculty of Geology Grant number BST 166901. Field works were supported by Polish Ministry of Science and Higher Education, grant number 691-N/2010/0 Ukraina. Authors are thankful to Teresa Tomek and Krzysztof Wertz (Institute of Systematics and Evolution of Animals PAS in Cracow, Poland) for identiÀcation of bird remains. Valuable comments on the manuscript by two anonymous reviewers are highly appreciated.
REFERENCES
Alexandrowicz, S.W., 1983. Malacofauna of the Holocene calcareous sediments of the Cracow Upland. Acta Geologica Polonica, 33(3), 117-158.
Alexandrowicz, S.W., 1992. Malacofauna from a rock-shelter in Ruskie Skały Tors (Będkowska Valley, Cracow Upland). Folia Quaternaria, 63, 27-34.
Alexandrowicz, W.P., 1997. Malacofauna of Quaternary deposits and environmental changes of the Podhale Basin during the Late Glacial and Holocene. Folia Quaternaria, 68, 7-132.
Alexandrowicz, S.W., 2000a. Malacofauna of Holocene cave sediments of the Cracow Upland (Southern Poland). Folia Quaternaria, 71, 85-112.
Alexandrowicz, W.P., 2000b. Molluscan assemblages from cave and slope sediments of the Częstochowa Upland (Poland). Folia Quaternaria, 71, 113-137.
Alexandrowicz, W.P., 2001a. Late Vistulian and Holocene molluscan assemblages from calcareous tufa at Ostrysz Hill (Podhale Basin). Folia Malacologica, 9, 159-169.
Alexandrowicz, W.P., 2001b. Molluscan assemblages from deposits filling small karst forms in the Tatra Mountains (Southern Poland). Acta Carsologica, 30, 125-142.
Alexandrowicz, W.P., 2004. Molluscan assemblages of Late Glacial and Holocene calcareous tufas in Southern Poland. Folia Quaternaria, 75, 1-309.
Alexandrowicz, W.P., 2013a. Late Glacial and Holocene molluscan assemblages in deposits filling palaeolakes in Northern Poland. Studia Quaternaria, 30, 5-17.
Alexandrowicz, W.P., 2013b. Malacological sequence from profile of calcareous tufa in Groń (Podhale Basin, southern Poland) as an indicator of the Late Glacial/ Holocene boundary. Quaternary International, 293, 196-206.
Alexandrowicz, W.P., 2014. Molluscan assemblages in Late Holocene tufa cones in the Pieniny Mountains (southern Poland). Geological Quarterly, 58(2), 269-280.
Alexandrowicz, S.W., Alexandrowicz, W.P., 1995a. Molluscan fauna of the Upper Vistulian and Early Holocene sediments of South Poland. Biuletyn Peryglacjalny, 34, 5-19.
Alexandrowicz, S.W., Alexandrowicz, W.P., 1995b. Quaternary molluscan assemblages of the Polish Carpathians. Studia Geomorphologica Carpatho-Balcanica, 29, 41-54.
Alexandrowicz, S.W., Alexandrowicz, Z., 1999. Recurrent Holocene landslides: a case study of the Krynica landslide in the Polish Carpathians. The Holocene, 9(1), 91-99.
Alexandrowicz, W.P., Stworzewicz, E., 2003. Snails (Gastropoda). In: Valde-Nowak, P., Nadachowski, A., Madeyska, T. (eds.). Obłazowa cave. Human activity, stratigraphy and palaeoenvironment. Cracow, Institute of Archaeology and Ethnology PAS, 91-94.
Alexandrowicz, W.P., Rudzka, D. 2006. Molluscan communities from cave and slope deposits of the limestone rocky hills in the eastern part of Podhale Basin (Southern Poland). Folia Malacologica, 14, 191-201.
Alexandrowicz, S.W., Alexandrowicz, W.P., 2011. Analiza malakologiczna metody badań i interpretacji. (Malacological analyses methods of investigation and interpretation). Rozprawy Wydziału Przyrodniczego Polskiej Akademii Umiejetności, 3, 5-302.
Alexandrowicz, W.P., Rybska, E., 2013. Environmental changes of intramontane basins derived from malacological analysis of profile of calcareous tufa in Niedzica (Podhale Basin, Southern Poland). Carpathian Journal of Earth and Environmental Sciences, 8(4), 13-26.
Alexandrowicz, S.W., Nadachowski, A., Rydlewski, J., Valde- Nowak, P., Wołoszyn, B., 1985. Subfossil fauna from cave sediments in the Sobczański Guly (Pieniny Mts. Poland). Folia Quaternaria, 56, 57-78.
Alexandrowicz, S.W., Drobniewicz, B., Gintner, B., Kozłowski, J.K., Madeyska, T., Nadachowski, A., Pawlikowski, M., Sobczyk, K., Szyndlar, Z., Wolsan, M., 1992. Excavations in the Zawalona Cave at Mników (Cracow Upland, Southern Poland). Folia Quaternaria, 63, 43-76.
Alexandrowicz, W.P., Szymanek M., Rybska, E., 2014. Changes to the environment of intramontane basins in the light of malacological research of calcareous tufa: example of the Podhale Basin (Carpathians, Southern Poland). Quaternary International, 353, 250-265.
Bocheński, Z., Gintner, B., Kozłowski, J.K., Mook, W.G., Muszyński, M., Nadachowski, A., Stworzewicz, E., Szyndlar, Z., 1985. Excavations of the rock-shelters in Zalas near Cracow. Folia Quaternaria, 56, 3-56.
Bronk Ramsey, C., Lee, S., 2013. Recent and planned developments of the program OxCal. Radiocarbon, 55, 720-730.
Cyrek, K., 1994. Wyniki badań sondażowych na Górze Słupsko (stanowisko 3) w Kostkowicach, województwo częstochowskie, w 1991 r. In: Tomczak, E. (ed.). Badania archeologiczne na Górnym Śląsku i w Zagłębiu Dąbrowskim w latach 1991-1992. Katowice, Centrum Dziedzictwa Kulturowego Górnego Śląska, 26-33.
Cyrek, K., Nadachowski, A., Madeyska, T., Bocheński, Z., Tomek, T., Wojtal, P., Miękina, B., Lipecki, G., Garapich, A., Rzebik- Kowalska, B., Stworzewicz, E., Wolsan, M., Godawa, J., Kościów, R., Fostowicz-Frelik, L., Szyndler, Z., 2000. Excavation in the Deszczowa Cave (Kroczyckie Rocks, Częstochowa Upland, Central Poland). Folia Quaternaria, 71, 5-84.
Dagnan-Ginter, A., Drobniewicz, B., Godawa, J., Miękina, B., Sobczyk, K., Stworzewicz, E., 1992. Excavations in the Duża Cave at Mączna Skała near Cracow (Southern Poland). Folia Quaternaria, 63, 3-25.
Dehm, R., 1967. Die landschnecke Discus ruderatusim Postglazial Süddeutschlands. Mitteilungen der Bayerische Staatssammlung für Paläontologie und Historische Geologie, 7, 135-155.
Dehm, R., 1987. Die landschnecke Discus perspectivus im Postglazial Südbayerns. Mitteilungen der Bayerische Staatssammlung für Paläontologie und Historische Geologie, 27, 21-30.
Fuhrmann, R., 1973. Die spätweichselglaziale und holozäne Molluskenfauna Mittel – und Westsachsens. Freiberger Forschungshefte, Reihe, C, 278, 1-121.
Füköh, L., 1995. History of the Hungarian Holocene mollusc fauna. Geojournal, 36(3-2), 255-259.
Gedda, B., 2001. Environmental and climatic aspects of the Early and Mid Holocene calcareous tufa and land mollusc fauna in southern Sweden. Lundqua Thesis, 45, 1-50.
Gerber, J., 1996. Revision der Gattung Vallonia Risso 1826 (Mollusca: Gastropoda: Valloniidae). Schriften zur Malakozoologie aus dem Haus der Natur-Cismar, 8, 1-227.
Gradziński, R., 1977. Sedymentacja piasków formierskich na skrasowiałym podłożu środkowej części Jury Krakowsko- Częstochowskiej. Kras i speleologia, 1(10), 59-70.
Horsák, M., Chytrý, M., Pokryszko, B.M., Danihelka, J., Ermakov, N., Hájek, M., Hájkova, P., Kintrová, K., Kočí, M., Kubešova, S., Lustyk, P., Otýpková, Z., Pelánková, B., Valachovič, M., 2010. Habitats of relict terrestrial snails in southern Siberia: lessons for the reconstruction of palaeoenvironments of full- glacial Europe. Journal of Biogeography, 37, 1450-1462.
Horsák, M., Chytrý, M., Hájkova, P., Hájek, M., Danihelka, J., Horsáková, V., Ermakov, N., German, D.A., Kočí, M., Lustyk, P., Nekola, J.C., Preislerová, Z., Valachovič, M., 2015. European glacial relict snails and plants: environmental context of their modern refugial occurrence in southern Siberia. Boreas, 44(4), 638-657. DOI: 10.1111/bor.12133
Juřičková, L., Horsák, M., Horáčková, J., Abraham, V., Ložek, V., 2014. Patterns of land-snail succession in Central Europe over the last 15,000 years: main changes along environmental, spatial and temporal gradients. Quaternary Science Reviews, 93, 155-166.
Kerney, M.P., Cameron, R.A.D., Jungbluth, J.H., 1983. Die Landschnecken Nord- und Mitteleuropas. Hamburg-Berlin, Veralg Paul Parey, 384pp.
Krajcarz, M.T., Madeyska, T., 2010. Application of the weathering parameters of bones to stratigraphical interpretation of the sediments from two caves (Deszczowa Cave and Nietoperzowa Cave, Kraków-Częstochowa Upland, Poland). Studia Quaternaria, 27, 43-54.
Krajcarz, M., Wojenka, M., Krajcarz, M.T., Cyrek, K., 2014a. Wyniki badań wykopaliskowych w Schronisku nad Bramą Słupską w Kostkowicach oraz Jaskini Deszczowej w Kroczycach, województwo śląskie. In: Glanc-Zagaja, G. (ed.). Badania archeologiczne na Górnym Śląsku i ziemiach pogranicznych w latach 2011-2012. Katowice, Śląskie Centrum Dziedzictwa Kulturowego w Katowicach, 22-29.
Krajcarz, M.T., Krajcarz, M., Goslar, T., Nadachowski, A., 2014b. The first radiocarbon dated steppe polecat (Mustela eversmanii) from the Pleistocene of Poland. Quaternary International, 357, 237-244.
Krajcarz, M.T., Cyrek, K., Krajcarz, M., Mroczek, P., Sudoł, M., Szymanek, M., Tomek, T., Madeyska, T., 2016. Loess in a cave: Lithostratigraphic and correlative value of loess and loess-like layers in caves from the Kraków-Częstochowa Upland (Poland). Quaternary, International, 399, 13-30. DOI: 10.1016/j.quaint.2015.08.069
Kruk, J., Milisauskas, S., Alexandrowicz, S.W., Śnieszko, Z., 1996. Environmental changes and settlement on the loess uplands. An archaeological and palaeogeographical study on the neolithic in the Nidzica basin. Kraków, Instytut Archeologii i Etnologii Polskiej Akademii Nauk, 139pp.
Lewandowski, J., 1994. Pokrywy fluwioperyglacjalne Wyżyny Częstochowskiej. Przegląd Geologiczny, 42(12), 1009-1113.
Limondin-Lozouet, N., 2011. Successions malacologiques à la charnière Glaciaire/ Interglaciaire: du modèle Tardiglaciaire-Holocène aux transitions du Pleistocène. Quaternaire, 22(3), 211-220.
Limondin-Lozouet, N., Rousseau, D.D., 1991. Holocene climate as reflected by a malacological sequence at Verriers, France. Boreas, 20, 207-229.
Limondin-Lozouet, N., Preece, R.C., 2004. Molluscan successions from the Holocene tufa of St Germain-le-Vasson, Normandy (France) and their biogeographical significance. Journal of Quaternary Science, 19(1), 55-71.
Lorenc, M., 2013. Radiocarbon ages of bones from Vistulian (Weichselian) cave deposits in Poland and their stratigraphy. Acta Geologica Polonica, 63(3), 399-424.
Ložek, V., 1964. Quartärmollusken der Tschechoslowakei.Rozpravy Ustředniho Ustavu Geologického, 31, 1-374.
Ložek, V., 1980. Quaternary molluscs and stratigraphy of the Mažarna Cave. Československy Kras, 30, 67-80.
Ložek, V., 1986. Mollusca Analysis. In: Berglund, B.E. (ed.). Handbook of Holocene Palaeoecology and Palaeohydrology. Caldwell, New Jersey, The Blackburn Press, 729-740.
Ložek, V., 2000. Palaeoecology of Quaternary Mollusca. Antropozoikum, 24, 35-59.
Ložek, V., 2012. Molluscan and Vertebrate Successions from the Veľká Drienčanská Cave. Malacologica Bohemoslovaca, 11, 39-44.
Lyman, R.L., 1994. Vertebrate Taphonomy. Cambridge, Cambridge University Press, 524pp.
Lyman, R.L., 2008. Quantitative Paleozoology. Cambridge, Cambridge University Press, 374pp.
Madeyska, T., 1996. Osady schroniska w Kruczej Skale w Skałach Kroczyckich (środkowa część Jury Polskiej). Kras i speleologia, 8(17), 57-65.
Madeyska, T., 1998. Vegetation in Poland at the close of last glaciation. Studia Geologica Polonica, 113, 137-180.
Meng, S., 2008. Neue Daten zur Verbreitung der Vertiginidae (Gastropoda: Pulmonata) in Zentralasien. Mollusca, 26, 207-219.
Meng, S., 2009. Rezente zentralasiatische und pleistozäne mitteleuropäische Faunen mit Vallonia tenuilabris (A. Braun, 1843). Mollusca, 27, 61-82.
Meyrick, R.A., 2002. Holocene molluscan faunal history and environmental change at Kloster Mühle, Rheinland-Pfalz, western Germany. Journal of Quaternary Science, 18(2), 121-132.
Meyrick, R.A, Preece, R.C., 2001. Molluscan succesions from two Holocene tufas near Northampton, English Midlands. Journal of Biogeography, 28, 77-93.
Nadachowski, A., Żarski, M., Urbanowski, M., Wojtal, P., Miękina, B., Lipecki, G., Ochman, K., Krawczyk, M., Jakubowski, G., Tomek, T., 2009. Late Pleistocene Environment of the Częstochowa Upland (Poland) Reconstructed on the Basis of Faunistic Evidence from Archaeological Cave Sites. Kraków, Institute of Systematics and Evolution of Animals, Polish Academy of Sciences, 112pp.
Preece, R.C., Day, S.P., 1994. Comparison of Post-glacial molluscan and vegetational successions from a radiocarbon- dated tufa sequence in Oxfordshire. Journal of Biogeography, 21(4), 463-468.
Preece, R.C., Bridgland, D.R., 1999. Holywell Coombe, Folkestone: a 13,000 year history of an English chalkland valley. Quaternary Science Reviews, 18, 1075-1125.
Ralska-Jasiewiczowa, M., 1991. Ewolucja szaty roślinnej. In: Starkel, L. (ed.). Geografia Polski, środowisko przyrodnicze. Warszawa, Panstwowe Wydawnictwo Naukowe (PWN), 106-127.
Reimer, P.J., Baillie, M.G.L., Bard, E., Bayliss, A., Beck, J.W., Blackwell P.G., Bronk Ramsey, C., Buck, C.E., Burr, G.S., Edwards, R.L., Friedrich, M., Grootes, P.M., Guilderson,T.P., Hajdas, I., Heaton,T.J., Hogg, A.G., Hughen, K.A., Kaiser, K.F., Kromer, B., McCormac, F.G., Manning, S.W., Reimer, R.W., Richards, D.A., Southon, J.R., Talamo, S., Turney, C.S.M., van der Plicht, J., Weyhenmeyer, C.E., 2009. IntCal09 and Marine09 radiocarbon age calibration curves, 0–50,000 years cal BP. Radiocarbon, 51(4), 1111-1150.
Rozmus, D., Szmoniewski, B., Troncik, A., 2006. An early medieval assemblage of iron artefacts from ‘Góra Slupsko’ hillfort, site 2 at Kostkowice, Zawiercie district, slaskie Voivodeship. Acta Archaeologica Carpathica, 41, 101-113.
Schileyko, A.A., 1984. Land mollusc suborder Pupillina of USSR fauna (Gastropoda, Pulmonata, Geophila) (in Russian). Leningrad (Russia), Fauna of the USSR, New Series, Nauka, 130, 399pp.
Starkel, L., 1977. Paleogeografia holocenu. Warszawa, Panstwowe Wydawnictwo Naukowe (PWN), 361pp.
Starkel, L., Michczyńska, D.J., Krąpiec, M., Margielewski, W., Nalepka, D., Pazdur, A., 2013. Progress in the Holocene chrono-climatostratigraphy of Polish territory. Geochronometria, 40(1), 1-21.
Stworzewicz, E., 1973. Kopalna fauna ślimaków (Gastropoda) ze schroniska nad Jaskinią Niedostępną w okolicach Ojcowa. Acta Zoologica Cracoviensia, 18, 301-309.
Stworzewicz, E., 1988. Fauna mięczaków jaskiń i schronisk Doliny Sąspowskiej. In: Chmielewski, W. (ed.). Jaskinie Doliny Sąspowskiej. Prace Instytutu Archeologii, Uniwersytetu Warszawskiego (UW), 39-45.
Svoboda, J., Horáček, I., Ložek, V., Svobodová, H., Šilar, J., 2000. The Pekárna Cave. Magdalenian stratigraphy, environment, and the termination of the loess formation in Moravian Karst. Antropozoikum, 24, 61-79.
Welter-Schultes, F., 2012. European non-marine molluscs, a guide for species identification. Goettingen, Planet Poster Editions, 760pp.
White, D., Preece, R., Shchetnikov, A.A., Parfitt, S.A., Dlussky, K.G., 2008. A Holocene molluscan succession from floodplain sediments of the upper Lena River (Lake Baikal region), Siberia. Quaternary Science Reviews, 27, 962-987.
Wiktor, A., 2004. Ślimaki lądowe Polski. Olsztyn, Wydawnictwo Mantis, 302pp.
Wojciechowski, A., 2000. Palaeohydrological changes in the Central Wielkopolska Lowland during the last 12 000 years on the basis of deposits of the Kórnik- Zaniemyśl Lakes. Poznań, Wydawnictwo Naukowe, Uniwersytetu im. Adama Mickiewicza w Poznaniu (UAM), 236pp.