New palaeobotanical data from Carboniferous Culm deposits constrain the age of the Variscan deformation in the eastern Pyrenees
New palaeobotanical data from Carboniferous Culm deposits constrain the age of the Variscan deformation in the eastern Pyrenees
Geologica Acta: an international earth science journal, vol. 16, núm. 2, pp. 107-123, 2018
Universitat de Barcelona
Recepción: 15 Julio 2017
Aprobación: 15 Enero 2018
Publicación: 15 Abril 2018
Abstract: The pre-Variscan rocks of the Pyrenees exhibit a polyphase deformation linked to the Variscan crustal shortening and a low-pressure–high-temperature metamorphism. However, there is scarce chronostratigraphic evidence of this Variscan deformation. In the Pyrenean low-grade metamorphic domains, maximum ages have been provided by the synorogenic Carboniferous Culm deposits. In medium- to high-grade metamorphic areas, the Variscan regional metamorphism or intrusive magmatic bodies constrain the age of the main Variscan deformation structures. However, these data usually provide a minimum age. Here, we present new palaeobotanical records that assign a Namurian age to the base of the Culm deposits of la Cerdanya in the eastern Pyrenees. This dating is based on the co-occurrence of the sphenopsids Archaeocalamites radiatus, Mesocalamites cistiiformis and the seed of Cardiocarpus sp. The plant remains were found in sandstone facies produced by high-density turbidity flows of a deep-sea fan system. The new biostratigraphic information constrains the age of the Carboniferous Culm succession in the eastern Pyrenees.
Keywords: Biostratigraphy, Sphenophyta, Turbidites, Lower Carboniferous, Variscan Orogeny.
Cite as:
Martín-Closas, C., Trias, S., Casas, J.M., 2018. New palaeobotanical data from Carboniferous Culm deposits constrain the age of the Variscan deformation in the eastern Pyrenees. Geologica Acta, 16(2), 107-123. DOI: 10.1344/GeologicaActa2018.16.2.1
INTRODUCTION
Knowing the precise age of deformation events is important in understanding the evolution of orogenic processes. Geochronological methods are useful in determining the age of regional metamorphic or magmatic processes, while synorogenic sediments, if present, can provide additional inform
Knowing the precise age of deformation events is important in understanding the evolution of orogenic processes. Geochronological methods are useful in determining the age of regional metamorphic or magmatic processes, while synorogenic sediments, if present, can provide additional information to constrain the age of the deformation in low-grade rocks. Like the other pre-Variscan rocks of Central and Western Europe, the pre-Variscan basement rocks of the Pyrenees were involved in the Variscan Orogeny linked to the closure of the Rheic Ocean and the collision of Laurussia and Gondwana in the Late Palaeozoic (Matte, 2002; Nance et al., 2010). In the northwest of Iberia, a continuous exposure enables the reconstruction of the different stages of the Variscan evolution, from the end of the oceanic closure in the late Devonian to the onset of the continental collision in the Tournaisian-Visean and the end of the contractional deformation in the Moscovian-Kasimovian (Martínez Catalán et al., 2009 and references therein). By contrast, evidence of deformation or high-pressure metamorphism linked to subduction during the early Variscan convergence is lacking in the pre-Variscan rocks of the Pyrenees. There, pre-Variscan rocks only display a polyphase deformation linked to the main period of Variscan crustal thickening and a low-pressure–high-temperature metamorphism.
Chronostratigraphic evidence for the Variscan shortening event in the Pyrenees is scarce. In low-grade metamorphic domains, data may be provided by the synorogenic Carboniferous deposits, which generally developed in turbidite or flysch basins and are known regionally as the Culm facies. However, the age of these deposits is difficult to ascertain. It has been estimated mainly using biostratigraphic data of conodonts from older rocks corresponding to limestones underlying the Culm deposits and from reworked carbonate blocks and olistostromes occurring within Culm conglomerates in other areas of the Pyrenees and Montagne Noire (Engel and Raymond, 1983; Delvolvé and Perret, 1989; Delvolvé et al., 1998). It should be noted that these methods provide maximum ages. Recently, a maximum depositional age was also provided by the U-Pb ages of detrital zircons (Martínez et al., 2015). A minimum age was provided by the dating of crosscutting granites (Romer and Soler, 1995; Maurel et al., 2004; Pereira et al., 2014). In medium- to high-grade metamorphic areas, the geochronological ages of regional metamorphism or intrusive magmatic bodies constrain the age of the main Variscan deformation structures. This method also provides a minimum age, as the main Variscan structures predate both thermal events.
Here, we present new palaeobotanical data constraining the age of the Culm deposits from la Cerdanya area, in the eastern Pyrenees, using plant remains that were coeval with the deposition. The study area is situated near the village of Bellver de Cerdanya (Catalonia), to the North of the Segre River, in the Carboniferous rocks located in the hangingwall of the Neogene normal fault bounding the la Cerdanya Basin (Figs. 1 and 2). These are the first data obtained from rocks in the eastern Pyrenees that directly provide an age for the Culm deposits, shedding light on the date of the Variscan deformation in the low-grade metamorphic rocks of this part of the chain.
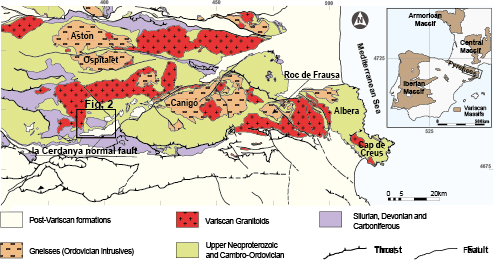
Figure 1.
Simplified geological map of the eastern Pyrenees showing the study area (see in the upper right corner for context map) (Fig. 2). Geographical coordinates are provided in UTM.
GEOLOGICAL SETTING OF THE STUDY AREA
The pre-Variscan rocks of the Pyrenees are involved in the Alpine lower thrust sheets and crop out extensively along the backbone of the mountain chain (Fig. 1). These rocks form an elongated strip, unconformably overlaid by Mesozoic and Cenozoic rocks that are not continuous with the neighbouring outcrops of the Catalan Coastal Range to the South or the Mouthoumet, Montagne Noire and French Central massifs to the North (Fig. 1). In the eastern Pyrenees, a complete pre-Variscan succession ranging in age from the Ediacaran to the late Mississippian/early Pennsylvanian can be identified.
Ediacaran to Lower Ordovician rocks constitute a thick (3,000 meters) sequence of metasedimentary rocks, with gneissic bodies, and layers of marbles, quartzites and calc-silicates interbedded in its lower part. A well-dated Upper Ordovician succession (Cavet, 1957; Hartevelt, 1970) lies unconformably over the sequence (Santanach, 1972; García-Sansegundo and Alonso, 1989; Casas and Fernández, 2007). Although it is difficult to evaluate the magnitude of this unconformity, it can be assumed that there was considerable erosion before the Upper Ordovician deposition. During the Silurian, black shales were deposited, which grade upwards to an alternation of black limestone and black shales. The Devonian is represented by a limestone sequence with common facies and changes in thickness along the chain. In the study area, Sanz-López (2002, 2004) identified a complete succession from the Lochkovian to the Famennian. The upper part of the Devonian succession consists of characteristic nodular limestone widely occurring throughout the Pyrenees. It should be noted that the first Carboniferous conodonts can be recorded on the uppermost part of these limestones (Sanz-López, 2002). Black radiolarian, chert and shales from the Tournaisian lie above the nodular limestone and are, in turn, overlain by an intercalation of chert with nodular to massive limestones of the Aspe-Brousset Formation (Fm.), that housed the conodonts from the Upper Tournaisian to Lower Serpukhovian. Siliciclastic rocks corresponding to the Culm facies, formally defined as Bellver Fm. in the study area (Brouwer in Hartevelt, 1970), overlie unconformably the Aspe-Brousset Fm. (Delvolvé et al., 1998; Sanz-López, 2002, 2004). The age of the Culm deposits is based on scarce fossils and varies along the Pyrenees (Delvolvé and Perret, 1989), although a Late Visean–Early Namurian (Serpukhovian) age for the Culm deposits in the eastern and central Pyrenees is accepted by most authors (Cygan et al., 1981; Engel and Raymond, 1983; Delvolvé et al., 1993; Delvolvé et al., 1998; see Discussion in Sanz-López, 2002).
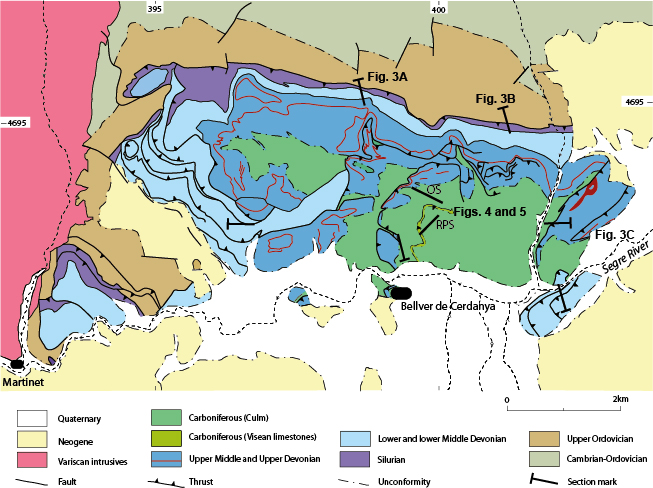
Figure 2.
Geological map of the study area in la Cerdanya, modified from Institut Cartogràfic i Geològic de Catalunya (2006). OS, Ordèn Section; RPS, Roca Punxenta Section. Location of cross-sections in Figure 3, and stratigraphic columns in Figures 4 (OS) and 5 (RPS) are shown. Geographical coordinates are provided in UTM.
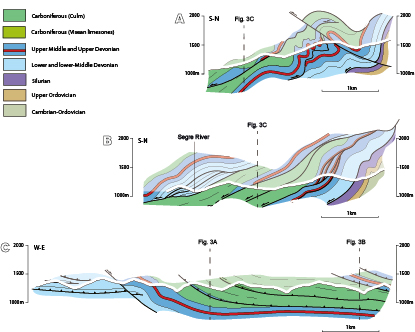
A pervasive cleavage (S3) linked to East-West trending folds (D3) is the main Variscan deformation mesostructure observed in low-grade pre-Variscan rocks of eastern Pyrenees. S3 is the main cleavage in the Pallaresa Massif, the Tor-Casamanya and Llavorsí synclines and the Rabassa and Orri domes (Speksnijder, 1986; Poblet, 1991; Clariana and García-Sansegundo, 2009; Margalef and Casas, 2016). The Culm rocks of the study area exhibit a well-developed cleavage that can be correlated with this S3 mesostructure. Clariana et al. (2009) and Clariana and García-Sansegundo (2009) indicated that a similar subvertical S3 cleavage is a crenulation that deforms a previously formed S1-2 foliation in the pre-Silurian rocks of the eastern part of the Pallaresa Massif, South of the Aston Massif and North of the Tor-Casamanya syncline. Additionally, in the study area, several thrust sheets involving Devonian and Carboniferous rocks can be observed (Figs. 2 and 3). Their basal detachment is located in the Silurian black shales and although most of the thrusts are South-directed, some North-directed thrusts can be observed (Figs. 2 and 3A; B). At outcrop scale, thrusts cut D3 folds and at kilometric scale South-verging D3 folds deform the thrusts and the lower detachment. The thrusts dip to the South and southeast in the southern limb of D3 megastructures, such as the Rabassa dome (Fig. 3A). Thus, thrust development must have been broadly synchronous with the D3 folds in the supra-Silurian rocks.
MATERIALS AND METHODS
The stratigraphy and sedimentology of the Bellver Fm. (Culm facies) were studied in two stratigraphic sections located in the thrust sheet cropping out North of the Bellver de Cerdanya Village (Fig. 2). These sections, Ordèn and Roca Punxeta sections, correspond to laterally equivalent facies within the same deep-sea fan system. The Ordèn Section crops out along the local road from Bellver to Talltendre, ca. 2.8km southeast of the village of Ordèn (base coordinates 42º23’12.01’’N, 1º46’52.54’’E and top coordinates 42º46’52.54’’N, 01º46’52.54’’E). It represents the base of the Bellver Fm. and shows one complete retrograding megasequence of a deep-sea fan. The Roca Punxenta Section was studied in the same road about 1.8km northeast of the junction with the main road N-260 (base coordinates 42º22’47.57’’N, 1º46’51.82’’E and top coordinates 42º22’52.36’’N, 1º46’59.82’’E). This section also contains the base of the Bellver Fm., but includes a more developed record of the channelised facies than the Ordèn Section. The Roca Punxenta Section yielded most of the plant remains studied here.
Plant remains were obtained after systematic sampling of all the beds, particularly sandstones, where they were most abundant. Plant fossils were prepared, analysed and photographed in the palaeontology laboratory of the Departament de Dinàmica de la Terra i de l’Oceà at the University of Barcelona, and are housed in the Museu de Ciències Naturals de Barcelona (MCNB) with the reference codes MGB 77557 to MGB 77585.
Stratigraphy, sedimentology and plant taphonomy
The study area corresponds to the stratotype of the Bellver Fm. The two sections analysed provide an insight into the stratigraphic framework and sedimentary facies of the lower part of this formation, helping to determine the transport and depositional conditions of the fossil plant remains studied.
The Ordèn Section
Description
The Ordèn Section shows one complete fining-upward sedimentary megasequence, about 135m-thick, belonging to the Bellver Fm. (Fig. 4). This megasequence occurs on top of a small sandstone-to-shale sequence that, in turn, lies unconformably on the limestone of the Compte Fm. (Hartevelt, 1970), which is rich in crinoids. The base of the megasequence is a massive conglomerate, about 5m-thick, with a lateral continuity about 100m and an erosive convex base. The clasts, supported by a grey pelitic matrix, are of up to 10cm in diameter and poorly sorted. They comprise mostly white hydrothermal quartz (rounded clasts) and Tournaisian black chert (angular clasts) and, rarely, granite clasts but no carbonate lithologies. These facies correspond to what Mutti and Ricci Lucchi (1972) and Walker and Mutti (1973) call unorganised conglomerate in a pelitic matrix (facies A2). On top of the basal matrix-supported conglomerate, there is a succession, up to 20m-thick, containing a number of small sequences, beginning with lenticular layers of massive 1–4m-thick clast-supported conglomerate that pass upward to massive grey sandstone beds, a few decimeters-thick. Most clasts share a similar lithology to that of the basal conglomerate, their diameters decreasing upward in the section from up to 20cm in the first sequences to up to 10cm in the other sequences. The clasts occasionally show imbrications, but the conglomerate beds do not generally show any sedimentary structure. These conglomerates correspond to facies A1 of Mutti and Ricci Lucchi (1972) and Walker and Mutti (1973). On top of the clast-supported conglomerate, there is a 25m-thick interval of sandstones, formed of poorly sorted medium- to coarse-grained sand, containing mainly whitish quartz and black chert lithoclasts, but no carbonate. They are stratified in massive beds a few decimeters thick. These sandstones contain abundant plant remains, mainly unidentifiable woody axes and comminuted plant debris. The upper part of the megasequence consists of more than 60m of fine to medium-grained sandstones intercalated with shales, the latter becoming more abundant towards the top. These deposits show a centimeter-to-decimeter recurrence of sandstone intervals, with plane-parallel lamination and ripple marks passing upward to grey finely-laminated shales.
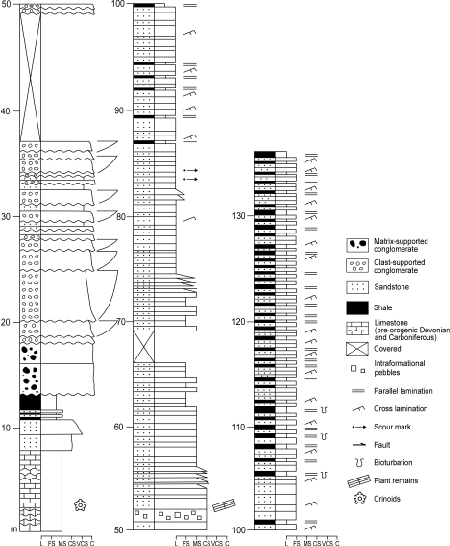
Figure 4.
Stratigraphic log of the base of Bellver Formation (Culm facies) at Ordèn (location shown in Fig. 2). Legend: L, Lutite/Shale; FS, Fine Sandstone; MS, Medium-grained Sandstone; CS, Coarse-grained Sandstone; VCS, Very Coarse-grained Sandstone; C, Conglomerate.
Interpretation
The fining-upward megasequence is attributed to deposition in a retrograding deep-sea fan, with facies corresponding to slope aprons, upper-fan channelised facies and interchannel facies, according to the classification of Mutti and Ricci Lucchi (1972), Walker and Mutti (1973) and Stow et al. (1985). The basal matrix-supported conglomerate is attributed to a single sedimentation event of debris flow of high-viscosity in the slope apron, as shown by the poor-sorting and the abundant pelitic matrix that supports the clasts. The overlying sequences of clast-supported conglomerate grading into massive sandstones are attributed to the infilling of deep-sea fan channels, as attested by the lenticular shape of the conglomerate beds, its erosive base and the occasional imbrication of clasts, indicating transport by traction. The lithology of the conglomerate clasts suggests that they arose from two very different sources: rounded whitish quartz may represent polycyclic clasts from an old hydrothermal, probably an Ordovician, source, while angular black chert represents the immediately pre-orogenic Tournaisian chert. The absence of limestone clasts suggests that the underlying largely Devonian Compte Fm. was still unexposed in the source area of the Culm conglomerates in Bellver, refuting the assumption that the limestone intercalations in the Culm deposits represent olistostromes from dismantled coeval carbonate platforms as described in other areas of the Pyrenees (Delvolvé and Perret, 1989). Instead, these intercalations have been interpreted as repeated thrust sheets cutting the top of the pre-orogenic carbonatic rocks (Institut Cartogràfic i Geològic de Catalunya, 2006). Most of the medium- to coarse-grained sandstone deposits overlying the basal conglomerate correspond to single events of high-density flow depositions, as indicated by their tabular shape and massive structure. Finally, the upper part of the megasequence, formed by fine- to medium-grained sandstones intercalated with laminated shales, are attributed to interchannel turbidite facies.
The Roca Punxenta Section
Description
This section unconformably overlies pre-orogenic Visean limestone (Aspe-Brousset Fm.) and chert. The section contains a number of thinning- and fining-upward sequences, each up to 30m-thick, identical in facies and vertical evolution (Fig. 5). They begin with 1–4m-thick clast-supported conglomerate organised in beds 0.5–2m-thick. These beds are massive and show a lenticular shape, an erosive base and a flat top. The clasts are up to 10cm in diameter and mainly composed of white hydrothermal quartz (rounded) and black chert (angular), rarely of granite or schist, being limestone absent. They correspond to A1 facies of Mutti and Ricci Lucchi (1972) and Walker and Mutti (1973). The basal conglomerate in each sequence is usually overlain by medium- to coarse-grained sandstone intervals, several meters-thick, organised in tabular beds ranging from a few decimeters to 4 meters in thickness. Most of these sandstones are massive, showing a microconglomeratic lag deposit, and some contain abundant woody axes, providing the plant material studied herein. On top of the sandstone interval, some of the sequences display an interval of fine sandstones grading to finely-laminated grey shales (Bouma sequences Tabc and Tabcd). These sequences become increasingly enriched in finer deposits upward in the section.
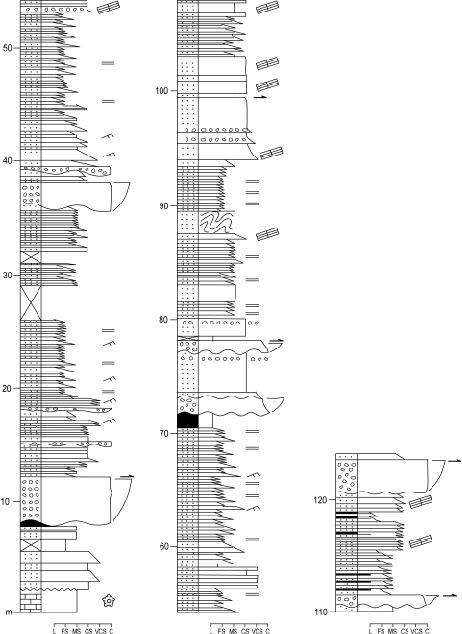
Figure 5.
Stratigraphic log of the base of the Bellver Formation (Culm facies) at Roca Punxenta (location shown in Fig. 2). Legend as in Figure 4.
Interpretation
Each of the cycles described, composed of a clast-supported conglomerate at the base followed by a massive sandstone interval and fine sandstones and shales (Bouma sequences), represent a single event of channelised turbidites as described by Mutti and Ricci Lucchi (1972) and Walker and Mutti (1973). The succession shows progressively less energetic and less channelised facies. The basal conglomerate represents the infilling of the turbidite channel, as indicated by their lenticular shape, erosive base and multilayered internal structure. Tabular, medium- to coarse-grained sandstone beds with a lag deposit and massive internal structure were produced by single high-viscosity turbidity flow expanding into laminar fluxes over the infilled channel. These flows carried most of the plant remains studied herein. Finally, the recurrences of fine- to medium-grained sands and laminated shales indicate gravity-mediated deposition of the suspended sediment load. These sediments accumulated under too low energy conditions to carry any plant macro-remains.
The Ordèn Section includes a single fining- and thinning-upward cycle with relatively more proximal facies (slope apron facies and proximal channelised facies) than the Roca Punxenta Section, which contains abundant fining- and thinning-upward cycles, but only facies corresponding to channel and interchannel areas of the deep-sea fan. Thus, the Ordèn Section represents the upper deep-sea fan area, while the Roca Punxenta Section represents the middle part of the fan.
Plant taphonomy
The studied plant remains were found in the middle part of the deep-sea fan, in the sandstone beds corresponding to high-density flows. These beds contain two types of plant taphofacies (Fig. 6). One of these types involves abundant non-oriented woody axes tridimensionally preserved as casts within the sandstone, and rare lignified seeds. The axes show large variations in size (from comminuted debris that are millimeters-long to 20cm-long axes), but less variation in their plant organ type and shape (most are elongated axes and rarely seeds). Assemblages of selected axes and seeds deposited in marine environments indicate a long transportation involving floating in the sea (Martín-Closas and Gomez, 2004). When the empty internal spaces of these remains became saturated with seawater, they sank to the seafloor and became incorporated into the turbidity flows that generated the sandstone layer, parts of which were filled in with sediment and buried. The infilling of the pith cavity of sphenophyte axes follows the model developed experimentally by Rex (1985), who showed that the quite regular infilling of the pith is due to the relatively short length of the axis parts preserved compared to their diameter. Most of the pith casts show signs of early diagenetic deformation, such as flattening, with the compression borders or depressions resulting from the incomplete infilling of the pith. Corresponding external casts, although also flattened, are much more regular in shape.

Figure 6.
Plant taphofacies found in the turbiditic deposits (Culm facies) of Bellver Formation. A) Plant taphofacies of tridimensionally preserved axes (internal casts) within the turbiditic sandstone. B) Plant taphofacies formed by adpressed axes at top of a turbiditic sandstone.
The other type of taphofacies corresponds to adpressed axes found on top of the sandstone beds. These remains, left on sand deposits after the turbidity flow ceased, were not filled in with sediment, thus hindering the formation of pith casts. They were completely flattened during the early burial and show parallel orientation.
Systematic palaeobotany
The first records of the occurrence of plant remains in the Culm deposits of Bellver were provided by Solé-Sabaris and Llopis-Lladó (1947), who had already reported the presence of Archaeocalamites (?) in “greywackes” of the section near Cal Codolet. More recently, Gómez-Alba (2007) noticed the occurrence of “calamitalean remains” near Bellver, corresponding to sample 86-1/2 of Lluís-Marià Vidal’s collection housed in the MCNB. They are poorly-preserved adpressions of a single sphenophyte axis. The present study provides the first taxonomic description of the plant remains found in the Culm deposits from Bellver de Cerdanya.
Phylum: Sphenophyta
Class: Sphenopsida
Order: Calamitales
Family: Archaeocalamitaceae
Genus ArchaeocalamitesStur, 1875
Type species: Archaeocalamites radiatus (Brongniart, 1828)
Material. One single portion of pith cast (specimen MGB 77557) about 15.5cm-long and 3.9cm-wide.
Description. Axis organised in nodes and internodes, the latter generally having greater widths than lengths (nodes are separated from each other by 1.8 to 2.5cm). Internodes display up to 40 clearly marked longitudinal and parallel ribs that are 1.3–2mm-wide, striated and continuous at each side of the nodes, forming a clearly visible straight line. Infranodal thickenings are generally absent, usually elongated and poorly visible when present. No branch scars are visible.
Chronostratigraphic range. The genus Archaeocalamites mainly occurs in the uppermost Devonian and lowermost Carboniferous deposits (Stewart and Rothwell, 1993; Dimichele et al., 2001; Taylor et al., 2009). However, Mamay and Bateman (1991) described the occurrence of Archaeocalamites lazarii in the Permian of Texas. This unique and extemporary record has been attributed to a relict occurrence of the group in particular facies (Cleal and Thomas, 1994) and is generally not considered for biostratigraphic purposes.

Figure 7.
Pith cast of Archaeocalamites radiatus (MGB 77557) from Roca Punxenta Section.
Family: Calamitaceae
Genus MesocalamitesHirmer, 1927
Type species: Mesocalamites cistiiformis (Stur, 1877)
Material. Four well-preserved pith casts, up to 10cm-long and 2cm-wide attributed to first order axes (MGB 77558–77561). Six smaller pith casts, up to 7cm-long and 8mm-wide attributed to second order axes (MGB 77562–77566, 77568).
Description. Axis organised in nodes and internodes. Internodes generally have greater lengths than widths. Two types of axes are preserved: i) first-order axes, which are up to 2cm-wide with nodes separated from each other by 6.0–6.3cm (Fig. 8), and ii) second-order axes that are up to 8mm-wide with nodes separated from each other by 2.8–3.0cm (Fig. 9).

Figure 8.
Pith casts of first-order axes of Calamites (Mesocalamites) cistiiformis from Roca Punxenta Section. A–C) General view of pith casts, D–F) details of the nodes showing both the continuous and alternating ribs along the nodal line. A, D) Specimen MGB 77559; B, E) Specimen MGB 77558; C, F) Specimen MGB 775561.
First-order axes display about 40 well marked longitudinal and parallel ribs that are 0.8–1.6mm-wide, sometimes finely striated. Ribs, either alternating or continuous at each side of the nodes, end in rounded tips, giving a relatively straight nodal line that is sometimes undulated. No visible branch scar. Infranodal thickenings are small, rounded and poorly visible.
Second-order axes display around 20–30 ribs, with similar features as those of first-order axes, but with predominantly continuous ribs at both sides of the nodal line. Two of these second-order axes (specimens MGB 77565 and MGB 77566) show 1–2 branches at each node, emerging at an acute angle and suggesting that the branching was opposite rather than alternate (Fig. 9B).

Figure 9.
Pith casts of Calamites (Mesocalamites) cistiiformis from Roca Punxenta Section. A–B) Second-order axes with branch bases (arrows); A) Specimen MGB 77566; B) Specimen MGB 77565; C) Axis between two nodes, specimen MGB 77562; D–E) Specimens MGB 77563 and MGB 77568, respectively, showing prevailing continuous ribs at both sides of the nodal line.
Remarks. According to Josten (1983), M. cistiiformis is very closely related to M. ramifer (Stur, 1877), but differs by the absence of the interdigitated ribs at the node forming a zig-zag line.
Chronostratigraphic range. Stewart and Rothwell (1993) indicated that Mesocalamites is typically reported from the middle to Upper Carboniferous (i.e. the Namurian); however, some species have also been documented from the Visean (Villalba-Breva and Martín-Closas, 2009). Specifically M. cistiiformis was reported by Josten (1983) from the top of the Namurian A until the upper part of the Namurian C in the Ruhr district (Germany), being very abundant in the Namurian B-C. These two old regional substages are equivalent to the modern Kinderscoutian, Marsdenian and Yeadonian substages according to Wagner and Winkler-Prins (2016) after Ramsbottom et al. (1978).
Phylum: Tracheophyta
Class: Pinopsida
Order: Cordaitales
Family: Cordaitaceae
Genus CardiocarpusBrongniart, 1881
Cardiocarpus sp.
Material. A single seed preserved as a limonitised adpression (MGB 77567).
Description. Rounded, bilaterally symmetrical, broadly heart-shaped seed with a maximum diameter of 1.5cm. The apex is rounded, slightly pointed, and the base concave (emarginated). The wing (sarcotesta) is well-developed, smooth and up to 2mm-wide in the equatorial area, decreasing in width towards the apex and base.
Remarks. The specimen found is similar in shape and size to that of C. sartisensisStockmans and Willière, 1961. However, the wing is not wider in the apex than in the equator in the material studied here, as it is in the holotype of this species.
Chronostratigraphic range. The organ genus Cardiocarpus is usually attributed to cordaitalean seeds (Taylor et al., 2009). Cordaitales are considered to have ranged mainly from the Lower Namurian to the Lower Permian (Stewart and Rothwell, 1993), although some isolated remains such as wood (Bell, 1992) and leaves (Mosseichik and Ruban, 2010) suggest that they might have an earlier origin.
DISCUSSION
Biostratigraphy of the Culm deposits of Bellver de Cerdanya based on plant remains
The Carboniferous biostratigraphy of plant macro-remains is based mainly on assemblages consisting of fern and pteridosperm foliage and lycopsid bark, e.g. the classical biozonations of Wagner (1984) and Cleal (1991). Sphenopsids have been usually considered less precise biostratigraphic indicators, given their wide chronostratigraphic ranges. Nevertheless, this plant group shows a relatively well-known chronostratigraphic succession of morphologies that remains consistent over large biogeographic areas and is extremely useful when other fossils are absent due to taphonomic factors. Although a formal biozonation based on these fossils has not been proposed yet, the succession of the sphenophyte species has been used by some authors, such as Josten (1983), for the biostratigraphic characterisation of the Namurian. In the case of the Culm facies and similar flysch deposits, sphenopsid remains are especially useful since they can withstand long-distance transport by flotation or traction even in very distal and deep marine settings. Furthermore, they are easily fossilised as adpressions or casts, the former without further reworking. Another type of plant remains that can be easily transported and preserved as allochthonous assemblages are pteridosperm and cordaitalean seed. The combination of both axes and seeds enable a relatively reliable biostratigraphic characterisation of the Culm deposits, based on coeval, rather than reworked fossils.

Figure 10.
Seed of Cardiocarpus sp. from Roca Punxenta section (specimen MGB 77567).
The biostratigraphic succession of Carboniferous sphenopsid genera is composed of three intervals. The first interval, represented by assemblages of Archaeocalamites, ranged from the Upper Devonian (Famennian), to Middle-Upper Visean, from where the first records of Mesocalamites have been reported (e.g. Villalba-Breva and Martín-Closas, 2009). The second interval (Upper Visean and Namurian) is represented by the concurrence of Archaeocalamites and Mesocalamites (e.g. Feist and Galtier, 1985), although Mesocalamites may have dominated or even been the only type of sphenopsid axis present (Josten, 1983). This interval ends with the disappearance of Archaeocalamites from most of the floras during the lowermost Pennsylvanian (e.g. Dimichele and Phillips, 1996; Taylor et al., 2009) and the change from Mesocalamites to Calamites (Boureau, 1964). The third interval, corresponding to the remaining part of the Pennsylvanian (largely the Westphalian and Stephanian), is where mainly Calamites sp. occurred among the sphenopsid axes (Stewart and Rothwell, 1993; Taylor et al., 2009). The base of the Pyrenean Culm deposits from Bellver de Cerdanya belonged to the second interval.
The reported concurrent ranges of Archaeocalamites radiatus, Mesocalamites cistiiformis and Cardiocarpus sp. suggest a Namurian age for the base of these Culm deposits (Fig. 11).
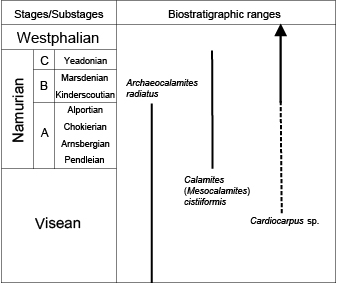
Figure 11.
Biostratigraphic range of the three taxa found in the Bellver Formation, based on Josten (1983), Stewart and Rothwell (1983) and Taylor et al. (2009). Regional substages according to Wagner and Winkler-Prins (2016).
Onset of the Variscan deformation in the eastern Pyrenees
The new palaeobotanical data presented here are the first to directly assign a Namurian (Serpukhovian to Bashkirian) age to the base of the Bellver Fm. (Culm deposits) in the eastern Pyrenees. The results clarify and constrain a previous age provided East of the study area by Sanz-López et al. (2006), who used foraminifers to attribute a latest Visean or Serpukhovian age to the Camprodon Fm. (sensu Trow in Boersma, 1973). This formation presumably corresponds, at least in part, to the Carboniferous flysch (Culm facies). Taken together, these data are compatible with the Serpukhovian age of the limestones underlying the Culm deposits in the Garona River (Ariège, France), confirming the westward migration of the Culm deposits from the eastern and central Pyrenees during the Namurian to the westernmost Cinco Villas Massif during the Middle Westphalian (Moscovian) (Delvolvé, 1981; Delvolvé and Perret, 1989; Delvolvé et al., 1993). Moreover, there was a southward migration (in present-day coordinates) of the Culm deposits from the Montagne Noire and Mouthoumet Massif (late Visean-early Namurian) to the eastern and central Pyrenees (Namurian) (Bessière and Schulze, 1984; Feist and Galtier, 1985; Delvolvé and Perret, 1989; Delvolvé et al., 1993; this study). Therefore, we can confirm a northeast-southwest migration, from internal to external zones, of the Variscan deformation, as proposed previously (Feist and Galtier, 1985; Delvolvé et al., 1993; Sanz-López, 2002).
The biostratigraphic results obtained in the present study constrain the age of the Variscan deformation in the low-grade supra-Silurian rocks of the eastern Pyrenees between 330 and 319Ma. This time span is consistent with the maximum depositional ages of 336±2Ma and 356±9Ma obtained from U-Pb dating of the detrital zircons in Carboniferous rocks at Guils de Cerdanya and Bagà, not far fom the study area (Martínez et al., 2015) (Fig. 12). A minimum age can be deduced from the age of the crosscutting Andorra-Montlluís granodiorite, dated as 305±3Ma by Romer and Soler (1995), 305±5Ma by Maurel et al. (2004) and 301.5±1.9Ma by Pereira et al. (2014), or more to the West by the radiometric age of the Maladeta and Lys-Caillaouas plutons, dated as 301.7±7.3Ma and 300±2Ma by Martínez et al. (2015) and Esteban et al. (2015), respectively. It is also compatible with the age of the Bagà rhyolite (287.7±3.7Ma) proposed by Martínez et al. (2015) and the age of the Castellar de n’Hug ignimbrite (283.4±1.9Ma) provided by Pereira et al. (2014) (Fig. 12). Both of these igneous rocks overly unconformably the Culm deposits near the study area. The presence of undeformed cobbles of igneous rocks in Culm conglomerates (Solé-Sabarís and Llopis-Lladó, 1947; Fontboté and Julivert, 1952; Hartevelt, 1970) confirms pre-latest Visean igneous activity in the Pyrenees (Soulcem and Bosost granites, ca. 339–336Ma old, according to Mezger and Gerdes, 2016) (Fig. 12), which significantly predates the much younger late Carboniferous magmatism (ca. 312–305Ma). By contrast, the presence of strongly foliated schist and gneiss cobbles (Solé-Sabarís and Llopis-Lladó, 1947; Hartevelt, 1970) indicates the pre-Culm Variscan ductile deformation (see discussion in Martínez et al., 2015). The age of this pre-Culm deformation is not well constrained, although it may be linked to the formation of the S1-2 foliation in medium- to high-grade pre-Silurian rocks and to the early Carboniferous Variscan metamorphism (ca. 360–350Ma) proposed by Martínez et al. (2015), based on the radiometric age of the monazite and metamorphic rims in zircons in gneiss cobbles. A different, younger, deformation event (D3 deformation), synchronous with the Culm sedimentation, could be responsible for the folds, cleavage and thrust formation in the Silurian, Devonian and Lower Carboniferous rocks. However, the correlation between the structures developed in both events (D1-2 and D3) is hampered by the different expression of each of these events in the low-grade and medium- to high-grade metamorphic rocks.
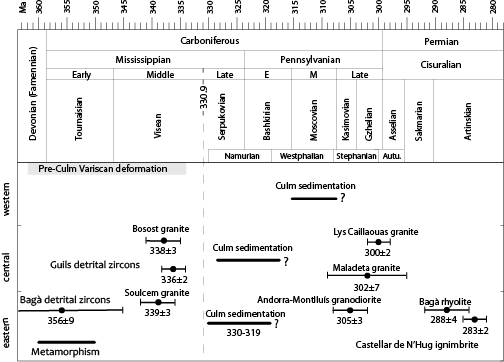
Figure 12.
Age of the Variscan deformation in the Pyrenees based on the Culm sedimentation. Culm sedimentation biostratigraphic data of the eastern Pyrenees based on this study. Central Pyrenean data based on Delvolvé (1981), Delvolvé and Perret (1989) and Delvolvé et al. (1993). Western Pyrenean data based on Delvolvé and Perret (1989), the crosscutting Andorra-Montlluís granodiorite (Romer and Soler, 1995), Maladeta granodiorite (Martínez et al., 2015) and Lys-Caillaouas granite (Esteban et al., 2015), the unconformable Bagà rhyolite (Martínez et al., 2015) and Castellar de N’Hug ignimbrite (Pereira et al., 2014), the youngest detrital zircon population from Guils and Bagà (Martínez et al., 2015), the Bossost and Soulcem granites that may provide the undeformed cobbles of igneous rocks (Mezger and Gerdes, 2016), and the Mississippian Variscan metamorphism (Martínez et al., 2015).
CONCLUSIONS
Vascular plant macro-remains, one cordaitalean seed and several sphenopsid axes, have been useful for the chronostratigraphic characterisation of the deep-sea fan deposits in Culm facies, which were previously characterised using only indirect evidence. These types of remains can withstand long transportation by floatation or traction, even in marine environments, but generally cannot withstand further reworking, being one of the few palaeontological remains coeval to the sedimentation found in Culm flysch deposits. The biostratigraphic succession of Archaeocalamites, Mesocalamites and Calamites provides a broad approximation of the flysch succession age, which is possible to determine even with only field data. Further refinement can be obtained by a detailed taxonomic study and biostratigraphic analysis. The age obtained in the Culm deposits of Bellver by the concurrent ranges of A. radiatus, M. cistiiformis and Cardiocarpus sp. is Namurian.
This new biostratigraphic information constrains the age of the Variscan deformation in the low-grade supra-Silurian rocks of the eastern Pyrenees to 330–319Ma. The data presented here agree with the occurrence of two different Variscan ductile deformation events that are pre-Culm and Culm in age, respectively, extending the time during which the Variscan deformation phases occurred in the Pyrenees to between the Visean and Namurian.
Acknowledgments
This work was funded by the grants CGL2010-21298, CGL2015-66335-C2-1-R of the Spanish Ministry of Science and Innovation (MINEICO) and 2017SGR-284 “Geologia Sedimentària” from the Catalan Research Authority. The authors are indebted to Prof. Jean Galtier (CIRAD, Montpellier) for his help in the determination of the fossil plant remains described herein. The two reviewers, Dr. C. Álvarez-Vázquez (Jardín Botánico de Córdoba) and Dr. F. Martínez (Universitat Autònoma de Barcelona), as well as the editor Dr. X. Delclòs (Universitat de Barcelona) are acknowledged for their constructive criticisms. The English text of the manuscript was revised by Dr. Ch. Evans (University of Barcelona). Aixa Tosal is acknowledged for photographing some of the plant remains figured.
References
Bell, P.R., 1992. Green Plants: their origin and diversity, 1st edition. Cambridge, Cambridge University Press, 349pp.
Bessière, G., Schulze, H., 1984. Le Massif Mouthoumet (Aude, France): nouvelle définition des unités structurales et essai d’une reconstruction paléogéographique. Bulletin de la Société Géologique de France, 26, 885-894.
Boersma, K.T., 1973. Devonian and Lower Carboniferous conodont biostratigraphy, Spanish Central Pyrenees. Leidse Geologische Mededelingen, 49, 303-377.
Boureau, É., 1964. Traité de botanique, Tome 3. Sphenophyta, Noeggerathiophyta. Paris, Masson et Compagnie Éditeurs, 544pp.
Brongniart, A., 1828. Histoire des végétaux fossiles ou recherches botaniques et géologiques sur les végétaux renfermés dans les couches du globe. Paris, D. Dufour Editeurs, 488pp.
Brongniart, A., 1881. Recherche sur les graines fossiles silicifiées. Paris, Masson, 93pp.
Casas, J.M., Fernández, O., 2007. On the Upper Ordovician unconformity in the Pyrenees: New evidence from the La Cerdanya area. Geologica Acta, 5(2), 193-198.
Cavet, P., 1957. Le Paléozoïque de la zone axiale des Pyrénées orientales françaises entre le Roussillon et l’Andorre. Bulletin Service Carte Géologique de France, 55, 303-518.
Clariana, P., García-Sansegundo, J., 2009. Variscan structure in the eastern part of the Pallaresa massif, Axial Zone of the Pyrenees (NW Andorra). Tectonic implications. Bulletin de la Société Géologique de France, 180, 501-511.
Clariana, P., García-Sansegundo, J., Gavaldà, J., 2009. The structure in the Bagneres de Luchon and Andorra cross sections (Axial Zone of the central Pyrenees). Trabajos de Geología, Universidad de Oviedo, 29, 175-181.
Cleal, C.J., 1991. Carboniferous and Permian biostratigraphy. In: Cleal, C.J. (ed.). Plant fossils in Geological Investigation: the Palaeozoic. Ellis Horwood, Chichester, 182-209.
Cleal, C.J., Thomas, B.A., 1994. Plant fossils of the British Coal Measures. The Palaeontological Association, London, 222pp.
Cygan, C., Perret, M.F., Raymond, D., 1981. Le Dévonien et le Carbonifère du “Synclinal de Villefranche-de-Conflent” (Pyrénées orientales, France): datation par Conodontes et conséquences structurales. Bulletin du Bureau des Recherches Géologiques et Minières, 2, 13-118.
Delvolvé, J.J., 1981. Arguments en faveur de l’âge namurien du Culm des Pyrénées Centrales françaises. Comptes Rendus de l’Académie des Sciences, Paris, 293, 219-222.
Delvolvé, J.J., Perret, M.F., 1989. Variations de l’âge des sédiments calcaires et “Culm” carbonifère dans la chaîne varisque du Sud de la France: migration de l’orogenèse varisque. Geodinamica Acta, 3, 117-126.
Delvolvé, J.J., Souquet, P., Vachard, D., Perret, M.F., Aguirre, P., 1993. Caractérisation d’un bassin d’avant-pays dans le Carbonifère des Pyrénées: faciès, chronologie de la tectonique synsédimentaire. Comptes Rendus de l’Académie des Sciences, Paris, 316(2), 959-966.
Delvolvé, J.J., Vachard, D., Souquet, P., 1998. Stratigraphic record of thrust propagation, Carboniferous foreland basin, Pyrenees, with emphasis on Pays-de-Sault (France/Spain). International Journal of Earth Sciences Geologische Rundschau, 87(3), 363-372.
Dimichele, W.A., Phillips, T.L., 1996. Clades, ecological amplitudes and ecomorphs: Phylogenetic effects and persistence of primitive plant communities in the Pennsylvanian age tropical wetlands. Palaeogeography, Palaeoclimatology, Palaeoecology, 127, 83-105.
Dimichele, W.A., Stein, W.E., Bateman, R.M., 2001. Ecological sorting of vascular plant classes during the Paleozoic evolutionary radiation. In: Allmon, W.D., Bottjer D.J. (eds.). Evolutionary Paleoecology: The ecological context of Macroevolutionary Change. Columbia University Press, New York, 285-335.
Engel, W., Raymond, D., 1983. Phénomènes resédimentation dans le Carbonifère à faciès Culm du Haut Pays de Sault (Est de la Zone Primaire Axial pyrénéenne, France); signification géodynamique dans le branche sud de l’orogène varisque. Bulletin de la Société Géologique de France, 25, 921-926.
Esteban J.J., Aranguren, A., Cuevas, J., Hilario, A., Tubía, J.M., Larionov, A., Sergeev, S., 2015. Is there a time lag between the metamorphism and emplacement of plutons in the Axial Zone of the Pyrenees? Geological Magazine, 152, 935-941.
Feist, R., Galtier, J., 1985. Découverte de flores d’âge namurien probable dans le flysch à olistolites de Cabrières (Hérault). Implications sur la durée de la sédimentation synorogénique dans la Montagne Noire. Comptes Rendus de l’Académie des Sciences, Paris, 300(2), 207-213.
Fontboté, J.M., Julivert, M., 1952. Algunas precisiones sobre la cronología de los plegamientos hercinianos en Cataluña. Congrès Geologique International d’Alger, Comptes Rendus, XIXème Session, 13(15) (Questions diverses de Géologie Génerale), 575-591.
García-Sansegundo, J., Alonso, J.L., 1989. Stratigraphy and structure of the southeastern Garona Dome. Geodinamica Acta, 3, 127-134.
Gómez-Alba, J., 2007. La cuenca carbonífera de Surroca-Ogassa (Ripollés, Cataluña, España). Monografies del Museu de Ciències Naturals de Barcelona, 4, 263pp.
Hartevelt, J.J.A., 1970. Geology of the upper Segre and Valira valleys, central Pyrenees, Andorra/Spain. Leidse Geologische Mededelingen, 45, 167-236.
Hirmer, M., 1927. Handbuch der Paläobotanik. Band I: Thallophyta, Bryophyta, Pteridophyta. Germany, Eicklingen, unbekannt, 708pp.
Institut Cartogràfic i Geològic de Catalunya, 2006. Mapa geològic comarcal de la Cerdanya. 1:50,000. Institut Cartogràfic i Geològic de Catalunya, 15.
Josten, K.H., 1983. Die fossilen Floren im Namur des Ruhrkarbons. Fortschritte in der Geologie von Rheinland und Westfallen, 31,1-327.
Mamay, S.H., Bateman, R.M., 1991. Archaeocalamites lazarii, sp. nov.: the range of Archaeocalamitaceae extended from the lowermost Pennsylvanian to the mid-lower Permian. American Journal of Botany, 78, 489-496.
Margalef, A., Casas, J.M., 2016. Corte geológico compensado del sur de Andorra: aportaciones a la estructura varisca del Pirineo central. Geo-Temas, 16, 61-63.
Martín-Closas, C., Gomez, B., 2004. Taphonomie des plantes et interprétations paléoécologiques. Une synthèse. Geobios, 37, 65-88.
Martínez, F.J., Dietsch, C., Aleinikoff, J., Cirés, J., Arboleya M.L., Reche, J., Gómez-Gras, D., 2015. Provenance, age, and tectonic evolution of Variscan flysch, southeastern France and northeastern Spain, based on zircon geochronology. Bulletin of the Geological Society of America, 128, 242-289.
Martínez Catalán, J.R., Arenas, R., Abati J., Sánchez Martínez, S., Díaz García, F., Fernández Suárez, J., González Cuadra, P., Castiñeiras, P., Gómez Barreiro, J., Díez Montes, A., González Clavijo, E., Rubio Pascual, F.J., Andonaegui, P., Jeffries, T.E., Alcock, J.E., Díez Fernández, R., López Carmona, A., 2009. A rootless suture and the loss of the roots of a mountain chain: The Variscan belt of NW Iberia. Comptes Rendus Geoscience, 341, 114-126.
Matte, Ph., 2002. Variscides between the Appalachians and the Urals: Similarities and differences between Paleozoic subduction and collision belts. In: Martínez Catalán, J.R., Hatcher, R.D., Arenas, R., Díaz García, F. (eds.). Variscan-Appalachian dynamics: The building of the late Paleozoic basement. Boulder, Colorado. Geological Society of America, Special Paper, 364, 239-251.
Maurel, O., Respaut, J.P., Monié, P., Arnaud, N., Brunel, M., 2004. U-Pb emplacement and 40Ar/39Ar cooling ages of the eastern Mont-Louis granite massif (Eastern Pyrenees, France). Comptes Rendus Geoscience, 336, 1091-1098.
Mezger, J., Gerdes, A., 2016. Early Variscan (Visean) granites in the core of central Pyrenean gneiss domes: implications from laser ablation U-Pb and Th-Pb studies. Gondwana Research, 29, 181-198.
Mosseichik, Y.W., Ruban, D.A., 2010. Viséan flora from the Moscow Coal Basin (Baltic Plate; European Russia): Local evolution in the context of global tendencies. Palaeogeography, Palaeoclimatology, Palaeoecology, 292, 168-183.
Mutti, E., Ricci Lucchi, F., 1972. Le torbiditi dell’Appennino settentrionale: introduzione all’analisi di facies. Memoria della Societá Geologica Italiana, 11, 161-199.
Nance, R.D., Gutiérrez-Alonso, G., Keppie, J.D., Linnemann, U., Murphy, J.B., Quesada, C., Strahan, R.A., Woodcock, N.H., 2010. Evolution of the Rheic Ocean. Gondwana Research, 17, 194-222.
Pereira, M.F., Castro, A., Chichorro, M., Fernández, C., Díaz-Alvarado, J., Martí, J., Rodríguez, C., 2014. Chronological link between deep-seated processes in magma chambers and eruptions: Permo-Carboniferous magmatism in the core of Pangaea (Southern Pyrenees). Gondwana Research, 25, 290-308.
Poblet, J., 1991. Estructura herciniana i alpina del vessant sud de la zona axial del Pirineu central. PhD Thesis, Universitat de Barcelona, 603pp.
Ramsbottom, W.H.C., Calver, M.A., Eagar, R.M.C., Hodson, F., Holliday, D.W., Stubblefield, C.J., Wilson, R.B., 1978. A correlation of Silesian rocks in the British Isles. Geological Society of London, Special Report, 10, 1-82.
Rex, G., 1985. A laboratory flume investigation of the formation of fossil stem fills. Sedimentology, 32, 245-255.
Romer, R.L., Soler, A., 1995. U-Pb age and lead isotopic characterization of Au-bearing skarn related to the Andorra granite (central Pyrenees, Spain). Mineralium Deposita, 30, 374-383.
Santanach, P.F., 1972. Sobre una discordancia en el Paleozoico inferior de los Pirineos orientales. Acta Geologica Hispanica, 7, 129-132.
Sanz-López, J., 2002. Devonian and Carboniferous pre-Stephanian rocks from the Pyrenees. In: García-López S., Bastida F. (eds.). Paleozoic conodonts from Northern Spain, Cuadernos del Museo Geominero, Instituto Geológico y Minero de España, 1, Madrid, 367-389.
Sanz-López, J., 2004. Silúrico, Devónico y Carbonífero pre- y sin-varisco de los Pirineos. In: Vera, J.A. (ed.). Geología de España, Sociedad Geológica de España – Instituto Geológico y Minero de España (SGE-IGME), Madrid, 250-254.
Sanz-López, J., Perret M.F., Vachard D., 2006. Silurian to Mississipian series of the eastern Catalan Pyrenees (Spain), updated by conodonts, foraminifers and algae. Geobios, 39, 709-725.
Solé-Sabaris, L., Llopis-Lladó, N., 1947. Mapa geológico de España, 1:50.000, Explicación de la Hoja nº 216, Bellver. Instituto Geológico y Minero de España, Madrid, 109pp.
Speksnijder, A., 1986. Geological analysis of Paleozoic large-scale faulting in the south-central Pyrenees. Geologica Ultraiectina, 43, 209.
Stewart, W.N., Rothwell, G.W., 1993. Paleobotany and the Evolution of Plants. 2nd Edition. Cambridge, Cambridge University Press, 521pp.
Stockmans, F., Willière, Y., 1961. Végétaux du Westphalien A de la Belgique. Graines, inflorescences et synanges. Publication num. 4, Centre National de Géologie Houillère, Brussels, 118pp.
Stow, D., Howell, D., Nelson, C.H., 1985. Sedimentary, Tectonic and Sea-Level Controls. In: Bouma, A.H., Normak, W.R., Barnes, N.E. (eds.). Submarine Fans and related Turbidite Systems. Springer-Verlag, New York, 15-22.
Stur, D., 1875. Die Culmflora des Mährisch-Schlesischen Dachschiefers. Abhandlungen der Kaiserlich-Königlichen Geologischen Reichsanstalt, 8, 1-106. [In German]
Stur, D., 1877. Die Culmflora der ostrauer und Waldenburger Schichten. Abhandlungen der Kaiserlich-Königlichen Geologischen Reichsanstalt, 8, 195-213. [In German]
Taylor, T.N., Taylor, E., Krings, M., 2009. Paleobotany.The Biology and Evolution of Fossil Plants, 2nd Edition. Academic Press, New York, 1230pp.
Villalba-Breva, S., Martín-Closas, C., 2009. Plant taphonomy from the Mississipian Flysch Facies of the El Priorat Massif (Catalonia, Spain). Journal of Taphonomy, 7, 249-262.
Wagner, R.H., 1984. Megafloral zones of the Carboniferous. Comptes rendus 9ème Congrès de Stratigraphie et de Géologie du Carbonifère (Washington and Urbana, 1979), 2, 109-134.
Wagner, R.H., Winkler-Prins, C.F., 2016. History and present status of the Pennsylvanian chronostratigraphic units: problems of definition and interregional correlation. Newsletter on Stratigraphy, 49, 281-320.
Walker, R.G., Mutti E., 1973. Turbidite facies and facies associations. In: Middleton, G.V., Bouma, A.H. (eds.). Turbidites and Deep Water Sedimentation. Society of Economic Palaeontologists and Mineralogists, Pacific Section, Short Course, Anaheim, 119-157.
Notas de autor
cmartinclosas@ub.edu
Enlace alternativo
http://revistes.ub.edu/index.php/GEOACTA/article/view/GeologicaActa2018.16.2.1/23516.pdf (pdf)